

Abstract
The NUP98-NSD1 fusion, product of the t(5;11)(q35;p15.5) chromosomal translocation, is one of the most prevalent genetic alterations in cytogenetically normal pediatric acute myeloid leukemias and is associated with poor prognosis. Co-existence of an FLT3-ITD activating mutation has been found in more than 70% of NUP98-NSD1-positive patients. To address functional synergism, we determined the transforming potential of retrovirally expressed NUP98-NSD1 and FLT3-ITD in the mouse. Expression of NUP98-NSD1 provided mouse strain-dependent, aberrant self-renewal potential to bone marrow progenitor cells. Co-expression of FLT3-ITD increased proliferation and maintained self-renewal in vitro. Transplantation of immortalized progenitors co-expressing NUP98-NSD1 and FLT3-ITD into mice resulted in acute myeloid leukemia after a short latency. In contrast, neither NUP98-NSD1 nor FLT3-ITD single transduced cells were able to initiate leukemia. Interestingly, as reported for patients carrying NUP98-NSD1, an increased Flt3-ITD to wild-type Flt3 mRNA expression ratio with increased FLT3-signaling was associated with rapidly induced disease. In contrast, there was no difference in the expression levels of the NUP98-NSD1 fusion or its proposed targets HoxA5, HoxA7, HoxA9 or HoxA10 between animals with different latencies to develop disease. Finally, leukemic cells co-expressing NUP98-NSD1 and FLT3-ITD were very sensitive to a small molecule FLT3 inhibitor, which underlines the significance of aberrant FLT3 signaling for NUP98-NSD1-positive leukemias and suggests new therapeutic approaches that could potentially improve patient outcome.
Introduction
Acute myeloid leukemia (AML) is a heterogeneous group of myeloid malignancies that are driven by a variety of genetic alterations which target signaling mediators, transcriptional regulators, as well as genes involved in mRNA processing or chromatin organization.1–3
Internal tandem duplications (ITD) in the cytoplasmic domain of FLT3 that lead to constitutive kinase activation are found in up to 20% of AMLs and are generally associated with elevated white blood cell counts and overall poor prognosis.4–6 Clinical observations and experimental studies have shown that FLT3-ITD can co-operate strongly in leukemia induction with a variety of leukemia-initiating gene fusions such as MLL-AF9, MLL-GAS7, AML1-ETO, PML-RARα or NUP98-HOXD13.7–13 Several studies have shown that FLT3-ITD alone can cause a myeloproliferative disorder in mice, but is not sufficient to induce AML.14–16
NUP98-NSD1 is the product of a cytogenetically silent chromosomal translocation t(5;11) (q35;p15.5) that generates a fusion between the N-terminus of nucleoporin 98 (NUP98) and the C-terminal part of the nuclear receptor-binding SET-domain-containing protein NSD1. NUP98-NSD1 is the most frequent gene fusion in cytogenetically normal (CN) pediatric AML, and is generally associated with an aggressive disease and poor prognosis.17–19 In adult AML patients, NUP98-NSD1 is rarely detected, but, if present, also confers a poor AML outcome.20,21 Interestingly, several reports have shown that the vast majority of NUP98-NSD1 positive patients also bear an FLT3-ITD mutation, suggesting functional co-operation.19–22 Previous studies have shown that retroviral overexpression of NUP98-NSD1 alone is able to induce an AML-like disease in mice after a latency of several months.23 Here we provide experimental evidence for a potent co-operation between the NUP98-NSD1 fusion and the FLT3-ITD mutation for the transformation of murine hematopoietic cells in vitro and in vivo.
Methods
Retroviral expression vectors
The pMSCV-NUP98-NSD1-neo and pMSCV-FLT3-ITD-IRES-EGFP retroviral expression vectors were kindly provided by Mark Kamps (San Diego, USA) and Justus Duyster (Freiburg, Germany) and have been described previously.15,23
In vitro transduction, proliferation and immortalization assays
1.5 × 105 lineage-surface marker negative (Lin-) bone marrow cells (MagCellect Kit, R&D systems, Minneapolis, USA) from 6–10 week old Balb/C or C57BL/6 mice were transduced with retrovirus on a 48-well fibronectin-coated plate according to the manufacturer’s protocol (Retronectin™, TaKaRa, Bio Inc., Shiga, Japan). The cells were selected with neomycin (0.8 mg/mL, Life Technologies, Paisley, UK) and cultured in medium containing murine stem cell factor (mSCF), FLT3 ligand, IL-11, thrombopoietin (PeproTech, Rocky Hill, NJ, USA). Growth in liquid culture was monitored for seven days by counting using dead exclusion dye, trypan blue (Life Technologies, Paisley, UK) in medium (RPMI 1640, 10% FBS) containing growth factors. The PKC-412 (Midostaurin) FLT3 inhibitor was purchased from Sigma-Aldrich (M1323, St Louis, USA). Transduced progenitors were plated in methylcellulose containing IL-3, IL-6 and mSCF (M3534, Stem Cell Technologies, Vancouver, Canada) and colonies were counted after seven days.
In vivo leukemogenesis study
3.5 × 105 in vitro expanded cells (expressing NUP98-NSD1, FLT3-ITD, or both) were injected into sub-lethally irradiated syngeneic Balb/C mice (1×500 rad) by tail vein injection. All experiments were carried out according to Swiss laws for animal welfare and approved by the Swiss Cantonal Veterinary Office of Basel-Stadt.
Quantitative RT-PCR
Total RNA was extracted using the RNA extraction kit (Macherey-Nagel, GmbH & Co, Duren, Germany) according to the manufacturer’s protocol. cDNA synthesis was carried out using the high capacity cDNA reverse transcription kit (Applied Biosystems, Foster City, CA, USA). mRNA expression was assessed by quantitative real-time PCR using SYBR Green on an ABI prism 7500 Sequence Detection System (Applied Biosystems). Results were normalized to Gapdh expression.
Retroviral integration cloning
Splinkerette-based PCR was performed as previously described.24 PCR amplicons were separated on a 2% agarose gel, purified and cloned into pCR2.1-TOPO (Invitrogen) and sequenced. The obtained sequences were analyzed using BLAST (NCBI).
Flow cytometry
For the immunophenotyping of the surface markers of the bone marrow cells, the following antibodies were used: c-kit-PE-Cy7, Mac-1-APC, FcγRII/III-PE (eBioscience, San Diego, USA) CD45R-B220-APC, Gr-1-APC-Cy7 (BD Biosciences), CD34-PE-Cy5 (Biolegend, San Diego, USA). For phospho-flow analysis, the cells were fixed (1.5% paraformaldehyde (PFA) for 10 min), permeabilized with 100% ice-cold methanol, and incubated with phospho-specific antibodies (pFLT3 (9351), pSTAT5 (3461); Cell signaling, Danvers, USA) followed by a secondary anti-rabbit, Alexa Fluor 405 labeled antibody (Life Technologies, Oregon, USA) The samples were run on a Cyan-ADP-Flow Cytometer (Dako Cytomation, Glostrup, Denmark) using Summit software (Summit Software Solutions Inc., Poway, CA, USA). Data were analyzed using FlowJo (Tree Star Inc., Ashland, OR, USA
Microscopy
Colonies were observed with an Olympus IX50 and cytospin preparations and histopathological sections were observed with an Olympus BX61 using the Cell Sens software (Olympus).
Statistical analysis
Statistical significance among groups was determined by t-test (two-tailed, unpaired/unequal variances). Comparisons of survival curves were conducted with the log rank test using Prism software v.5.0 (GraphPad Software, La Jolla, California USA).
Results
To study the co-operation between NUP98-NSD1 and FLT3-ITD, we transduced bone marrow stem and progenitor cells from two different mouse strains (Balb/C, C57BL/6) with retroviruses expressing NUP98-NSD1 or FLT3-ITD alone or together (Figure 1A). Cells transduced with NUP98-NSD1 underwent antibiotic selection for seven days and the expression of both transgenes was determined by RT-PCR (data not shown). Serial plating in methylcellulose showed that expression of NUP98-NSD1 alone increased the self-renewal capacity of Balb/C-derived cells. (Figure 1B, top panel). In contrast, expression of NUP98-NSD1 did not increase colony formation of C57BL/6-derived cells (Figure 1B, bottom panel). FLT3-ITD expression resulted in only limited re-plating activity of the cells of both strains. However, co-expression of NUP98-NSD1 and FLT3-ITD allowed serial plating of the cells. In addition, co-expression of NUP98-NSD1 and FLT3-ITD also resulted in significantly increased rates of proliferation of cells of both strains in liquid cultures (Figure 1C). Balb/C-derived progenitors solely transduced with the NUP98-NSD1 expressing virus formed dense colonies. Cytospin preparations revealed an immature blast-like morphology, whereas NUP98-NSD1 and FLT3-ITD co-expressing cells appeared slightly more differentiated (Figure 2A). In contrast, C57BL/6-derived cells expressing solely NUP98-NSD1 or FLT3-ITD completely lost their capacity of forming colonies and were differentiating towards mast cells and macrophages (Figure 2B). Cells that co-expressed NUP98-NSD1 and FLT3-ITD formed dense colonies containing cells with a blast-like morphology (Figure 2B). Immunophenotyping of Balb/C-derived cells showed that cells expressing solely NUP98-NSD1 exhibited high levels of the stem cell associated surface marker c-kit, the early myeloid FcγRII/III, but only low levels of the myelomonocytic differentiation markers Gr-1 and Mac-1 (Figure 2C, left panel). Co-expression of FLT3-ITD resulted in lower expression of c-kit and higher expression of FcγRII/III, Gr-1 and Mac-1 (Figure 2C, middle panel). In contrast, cells expressing only FLT3-ITD exhibited a more differentiated myeloid phenotype with high levels of Gr-1 and Mac-1 and lower levels of c-kit (Figure 2C, right panel). In summary, these data show that co-expression of NUP98-NSD1 with FLT3-ITD increased proliferation and the clonogenic activity of murine bone marrow cells in vitro. Unlike C57BL/6, Balb/C-derived bone marrow cells expressing solely NUP98-NSD1 could be serially re-plated.
Figure 1.
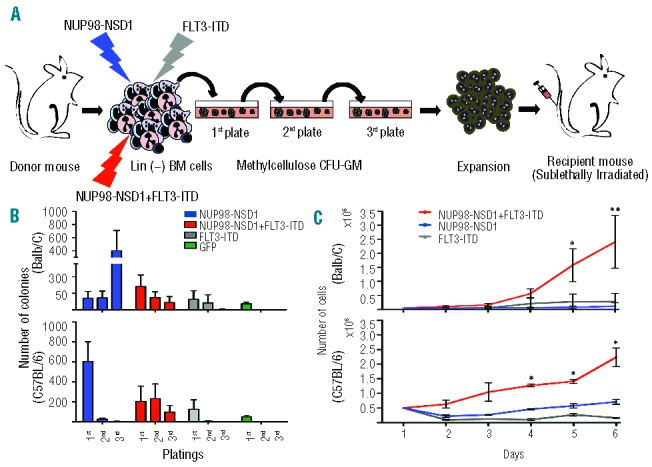
Functional co-operation of NUP98-NSD1 and FLT3-ITD in vitro. (A) Schematic outline of the experimental strategy to address the functional co-operation between NUP98-NSD1 and FLT3-ITD. (B) Serial plating in methylcellulose of cells expressing NUP98-NSD1, FLT3-ITD or both, of Balb/C (upper panel) and C57BL/6 mice (lower panel): NUP98-NSD1 increased colony formation of Balb/C-but not of C57BL/6-derived cells, whereas co-expression of FLT3-ITD supported colony formation in both strains. No significant re-plating activity was observed upon expression of FLT3-ITD or the control vector (GFP) in cells of both strains. (C) Proliferation of cells expressing NUP98-NSD1, FLT3-ITD or both in liquid culture. Co-expression of NUP98-NSD1 and FLT3-ITD significantly increased proliferation compared to single transduced cells. Proliferation was assessed in triplicates using trypan blue staining. Error bars represent the standard deviations from 5 independent experiments with Balb/C and 3 independent experiments with C57BL/6-derived cells (t-test, two-tailed, unpaired/unequal variances, *P ≤ 0.05, **P≤0.01).
Figure 2.

Phenotypic analysis of NUP98-NSD1 and FLT3-ITD expressing progenitors. (A, B) Photomicrographs (x100) of cytospots of cells and their colonies (smaller picture, x4) formed in methylcellulose cultures of Balb/C (A) and of C57BL/6 (B) -derived cells. NUP98-NSD1 expressing Balb/C progenitors formed dense colonies and exhibited a blast-like morphology. NUP98-NSD1 + FLT3-ITD co-expressing cells formed less dense colonies and had a more differentiated phenotype, whereas FLT3-ITD-only-expressing progenitors differentiated into mast cells and macrophages. C57BL/6 –derived cells co-expressing NUP98-NSD1 + FLT3-ITD formed dense colonies and exhibited a blast-like morphology. Cells expressing only NUP98-NSD1 or FLT3-ITD differentiated into mast cells and macrophages. (C) Flow cytometry revealed that NUP98-NSD1 expressing cells were c-kit+, FcγRII/III+, Mac-1lo, Gr-1lo. Co-expression of FLT3-ITD resulted in lower c-kit and higher FcγRII/III, Gr-1 and Mac-1 expression. FLT3-ITD single transduced cells were c-kitlo, FcγRII/III+, Gr-1+, Mac-1+.
We next examined a potential functional co-operation between NUP98-NSD1 and FLT3-ITD in vivo. For the in vivo experiments, we chose two independently generated sets of cell populations expressing NUP98-NSD1, FLT3-ITD or both, that we had analyzed in vitro. We focused on Balb/C-derived cells, as they were immortalized in vitro by NUP98-NSD1 alone and in combination with FLT3-ITD. We harvested bone marrow progenitor cells from methyl-cellulose cultures after 2 platings and transplanted them into sub-lethally irradiated syngeneic mice (Figure 1A). As shown in Figure 3A, all mice that received NUP98-NSD1 and FLT3-ITD co-expressing cells rapidly developed AML that was fully transplantable into secondary recipients (Online Supplementary Figure S1). The diseased mice exhibited enlarged spleens and significantly increased circulating white blood counts (Figure 3B). Histopathology showed widespread infiltration of the bone marrow, spleen and multiple solid organs (Figure 3C) with leukemic blasts that displayed myeloid surface antigens (Mac-1+/Gr-1+/FcγRII/III+/c-kitlo/CD34−) (Figure 3D). In contrast, mice transplanted with cells expressing NUP98-NSD1 alone developed some signs of distress after a significantly longer latency (Figure 3A). Histopathology of these mice showed an increase in the number of myeloid cells in the bone marrow and spleen (Figure 4A). The majority of the mice showed a mildly increased number of circulating neutrophils (Online Supplementary Figure S1C). Flow cytometry revealed an expansion of Mac-1+/Gr-1+ positive cells in the bone marrow, compatible with a phenotype of myeloid hyperplasia (Figure 4B). Mice receiving FLT3-ITD-only-expressing cells did not show any signs of disease over several months post-transplant.
Figure 3.

Functional co-operation of NUP98-NSD1 and FLT3-ITD in vivo. (A) Kaplan-Meier survival plot of mice transplanted with cells expressing NUP98-NSD1, NUP98-NSD1 + FLT3-ITD, or solely FLT3-ITD harvested after two rounds of plating in methylcellulose and expanded in liquid culture. Transplantation of cells co-expressing NUP98-NSD1 + FLT3-ITD resulted in AML after a median latency of 12 days (experiment “A”), and 91 days (experiment “B”). Mice receiving cells expressing only NUP98-NSD1 developed some signs of disease after a median latency of 334 days. (B) Diseased mice transplanted with cells co-expressing NUP98-NSD1 + FLT3-ITD showed significantly increased spleen weights and peripheral white blood cell counts. Error bars represent the standard deviation of 5 mice per group (t-test, two-tailed, unpaired/unequal variances, *P ≤ 0.05, **P≤0.01). (C) Representative images of peripheral blood smears stained with Wright-Giemsa (x60) (i), histopathological sections of bone marrow (x20) (ii), spleen (x10) (iii), and liver (x10) (iv) of diseased mice transplanted with NUP98-NSD1 + FLT3-ITD co-expressing cells. Leukemic blasts were found in the peripheral blood and extensively infiltrated the bone marrow, spleen and liver. (D) Leukemic blasts from mice transplanted with NUP98-NSD1 + FLT3-ITD transduced cells exhibited a myeloid phenotype, expressing Mac-1+, Gr-1+, FcγRII/III+, CD34−, c-kitlo as assessed by flow cytometry.
Figure 4.
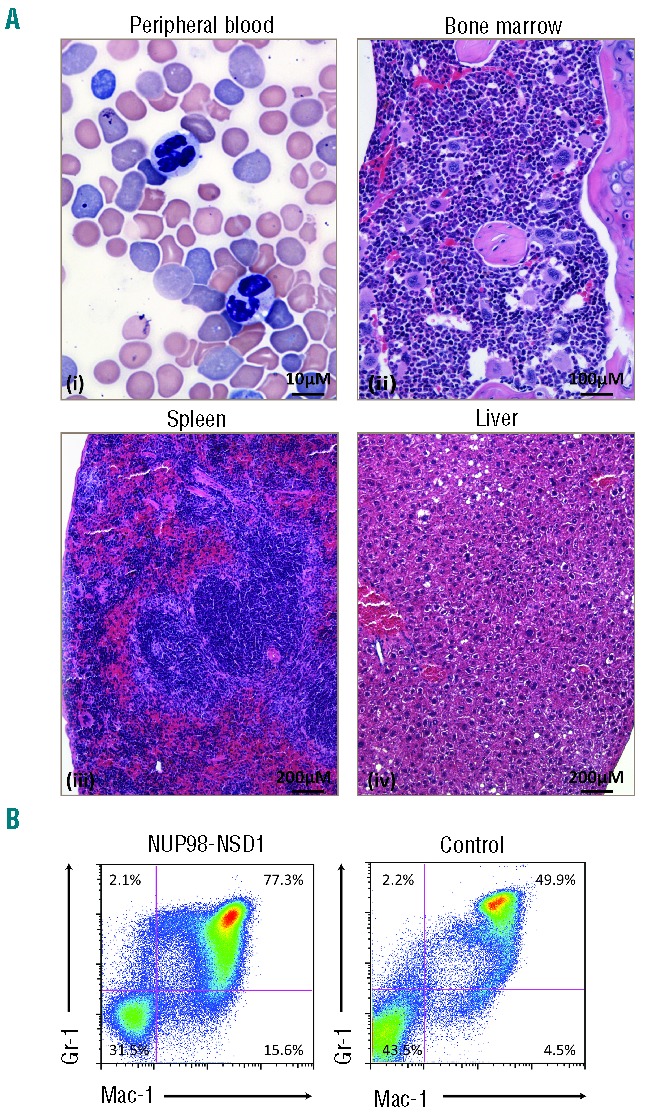
Mice transplanted with NUP98-NSD1 expressing progenitors develop myeloid hyperplasia. (A) Representative images of Wright-Giemsa stained peripheral blood smears (x60) (i), and histopathological sections of bone marrow (x20) (ii), spleen (x10) (iii) and liver (x10) (iv) of diseased mice transplanted with cells expressing only NUP98-NSD1. Histopathology revealed expansion of differentiating myeloid elements in the bone marrow and the spleen. (B) Mice transplanted with cells expressing only NUP98-NSD1 showed significant expansion of the myeloid bone marrow compartment (Mac-1+, Gr-1+) as assessed by flow cytometry compared to same aged control mice (t-test, two-tailed, unpaired/unequal variances, P=0.0043). The flow cytometry panel is representative for 5 transplanted mice compared to age-matched controls.
Two independent experiments resulted in different latency periods to develop the disease of mice transplanted with NUP98-NSD1 and FLT3-ITD co-expressing progenitors (12 days in experiment “A” vs. 91 days in experiment “B”) (Figure 3A). To explore the clonal composition, we compared the viral integration pattern of the cells harvested from the methylcellulose cultures before transplantation and the resulting AMLs of both experiments. As shown in Online Supplementary Figure S2, splinkerette-PCR revealed overlapping integration pattern after in vitro immortalization and the developing AML. Two viral integrations could be determined in experiment A and one in experiment B. These observations suggest in vivo expansion of an in vitro pre-selected clone. We also compared the expression levels of the HoxA gene cluster previously associated with transformation by NUP98-NSD1.23 As shown in Figure 5A, we observed significant increased levels of HoxA5, HoxA7, HoxA9 and HoxA10 RNA expression in the leukemic cells; however, there was no significant difference between both experiments. In addition, there was no difference in the expression levels of the NUP98-NSD1 fusion mRNA in the leukemic blasts between both experiments.
Figure 5.
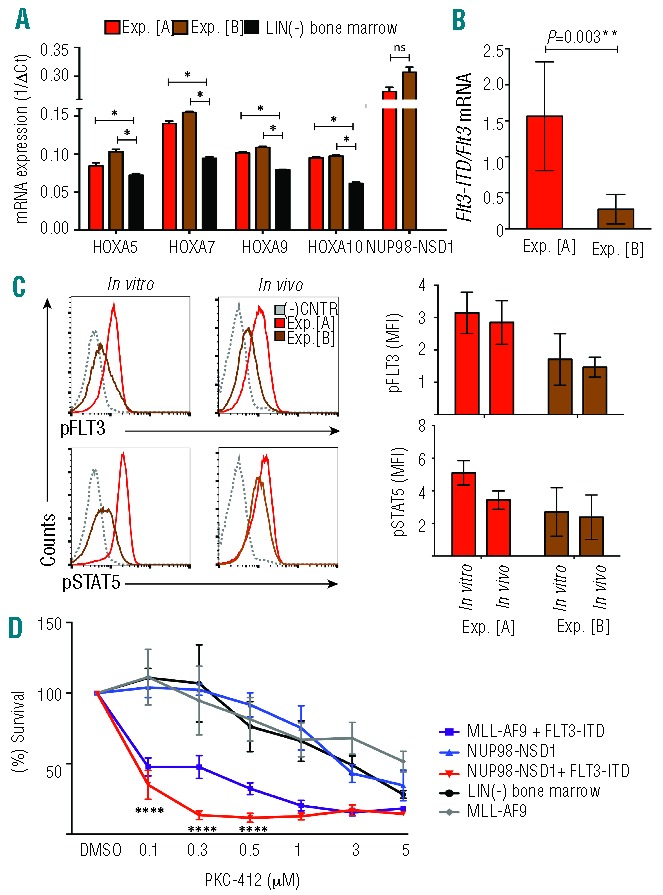
Increased FLT3-signaling in short-latency AML cells and sensitivity to a FLT3 inhibitor. (A) NUP98-NSD1 + FLT3-ITD expressing leukemic blasts from experiments “A” and “B” expressed significantly increased levels of HoxA5, HoxA7, HoxA9 and HoxA10 mRNA levels compared to lineage negative bone marrow cells. The expression of the NUP98-NSD1 was not different between experiments. Error bars represent the standard deviations from 2 experiments run in duplicates (t-test, two-tailed, unpaired/unequal variances, *P≤0.05). (B) NUP98-NSD1 + FLT3-ITD expressing leukemic blasts from transplanted mice express significantly different Flt3-ITD/wild-type Flt3 mRNA levels between experiment “A” and experiment “B” as detected by Q-PCR (P=0.003). A higher FLT3-ITD/wild type Flt3 mRNA ratio correlated with a shorter latency. The error bars indicate the standard deviation out of 5 analyzed mice per group in duplicates (t-test, two-tailed, unpaired/unequal variances, *P≤0.05). (C) Phospho-flow cytometric analysis for pFLT3 revealed that cells from experiment “A” express more pFLT3 than cells from experiment “B” both after in vitro immortalization and after in vivo AML induction. Similar results were found for pSTAT5. Graphs in the lower panel summarize the median fluorescence intensity obtained in 3 independent experiments. (D) Immortalized NUP98-NSD1 + FLT3-ITD co-expressing (Balb/C-derived) bone marrow progenitors are more sensitive to the PKC-412 small molecule FLT3 inhibitor than cells immortalized by NUP98-NSD1, MLL-AF9 fusion, or normal bone marrow cells. The graph represents survival of the cells after a 24-h-exposure to different doses of PKC-412 as assessed by a WST-1 assay performed in quadruplicates (t-test, two-tailed, unpaired/unequal variances, ****P≤0.0001).
In contrast, we observed that the different latency periods in experiments “A” and “B” correlated with an increased expression ratio of mutated Flt3-ITD to wild-type Flt3 mRNA. Higher Flt3-ITD mRNA expression levels were associated with a significantly shorter latency (Figure 5B). To address whether these changes might translate into different activation levels of FLT3-derived cellular signals, we performed phospho-flow analysis. As shown in Figure 5C, cells from experiment “A” after in vitro immortalization, as well as upon AML formation in vivo, exhibited higher levels of pFLT3 than cells from experiment “B”. Similarly, although less pronounced, we observed a moderate increase in the basal pSTAT5 levels in cells from experiment “A”. Collectively, these observations suggest that the shorter latency of experiment “A” is the consequence of a stronger activation of FLT3-ITD-rather than NUP98-NSD1-mediated cellular signals.
Selective small-molecule-inhibitors of FLT3 are currently being evaluated in clinical AML trials. We, therefore, also assessed the sensitivity of in vitro immortalized progenitors expressing NUP98-NSD1 or NUP98-NSD1 and FLT3-ITD to increasing doses of an FLT3 inhibitor (PKC-412, Midostaurin). As shown in Figure 5D, cells co-expressing NUP98-NSD1 and FLT3-ITD were significantly more sensitive to PKC-412 treatment than cells immortalized by NUP98-NSD1 alone. Likewise, cells immortalized by co-expression of the MLL-AF9 fusion with FLT3-ITD were more sensitive to PKC-412 than cells immortalized by MLL-AF9 or normal lineage marker-depleted mouse bone marrow cells. Taken together, these results demonstrate a potent co-operation between NUP98-NSD1 and FLT3-ITD in leukemic transformation in vitro and in vivo, and revealed that expansion of NUP98-NSD1 and FLT3-ITD co-expressing cells significantly depends on FLT3-derived signals.
Discussion
Previous studies had demonstrated that the vast majority of the NUP98-NSD1 positive AML patients also carry an activating FLT3-ITD mutation, indicating functional cooperation.19–22 In the present study, we provide experimental evidence that co-expression of NUP98-NSD1 and FLT3-ITD co-operate for transformation in vitro and the development of AML in vivo. In contrast, neither NUP98-NSD1 nor FLT3-ITD expression was able to induce the disease.
Previous studies have suggested that retroviral overexpression of the NUP98-NSD1 fusion in murine bone marrow cells could result in an AML-like disease in mice.23 The reasons for the discrepancy with our observations are not fully clear, but could be based on different protocols of retroviral production and transduction, or genetic differences of the mouse strains used. Interestingly, we observed in several independent experiments that expression of NUP98-NSD1 provided in vitro self-renewal capacity to Balb/C but not to C57BL/6-derived bone marrow cells. Co-expression of NUP98-NSD1 with FLT3-ITD was able to immortalize cells from both strains (Figure 1). Several previous studies have clearly demonstrated that Balb/C mice are more prone to transformation of hematopoietic cells, but the underlying mechanism is poorly understood.7 The observed strain-dependent differences of NUP8-NSD1 in hematopoietic cells suggest that Balb/C-derived cells might carry alterations that are functionally linked to NUP98-NSD1-induced transformation in vitro.25,26
In 2 independent experiments, we observed that mice transplanted with NUP98-NSD1 immortalized cells developed myeloid hyperplasia in the bone marrow and spleen, associated with increased neutrophil counts in the majority of the mice. The discrepancy between our results and those of Wang et al. might also be due to a different experimental setup since we transplanted in vitro immortalized and expanded bone marrow progenitor cells following a protocol previously established for the in vivo transforming potential of MLL-fusions.23,27,28 Nevertheless, retroviral overexpression of the NUP98-HOXA9 fusion has been previously shown also to first induce a chronic myeloproliferative disorder with progression to AML after a long latency.29 The observed generally long latency periods for AML development in mice upon transplanting cells overexpressing different NUP98 fusions support the idea of a general indispensable acquirement of secondary mutations such as e.g. FLT3-ITD.30
We and others have previously shown that transplantation of retroviral FLT3-ITD expressing progenitors can cause a myeloproliferative disease in vivo.14,15 In contrast to those experiments, we here transplanted in vitro selected and expanded progenitors. As shown in Figures 1B and 2, expression of FLT3-ITD alone resulted in a poor self-renewal capacity and was associated with differentiation of bone marrow progenitors towards monocytes/macrophages and mast cells, which most probably abrogated their leukemogenic potential in vivo.
Interestingly, expression of FLT3-ITD with NUP98-NSD1 not only led to AML after short latency in vivo, but also influenced the immunophenotype of the in vitro transformed cells, as seen by reduced c-kit and increased Mac-1, Gr-1 expression. Similar observations have been made upon co-expression of FLT3-ITD with the PML-RARα or MLL-AF9 fusions.7,10
In addition, we found that the length of the latency period for AML induction by co-expression of NUP98-NSD1 and FLT3-ITD correlated with the ratio of FLT3-ITD to wild-type Flt3 mRNA expression (Figure 5B). Phospho-flow experiments suggested that increased FLT3-ITD expression resulted in increased FLT3-derived cellular signals (Figure 5C). Interestingly, our observations in the mouse closely phenocopied recent observations demonstrating that the prognosis of AML patients with NUP98-NSD1 could be correlated to a high Flt3-ITD/wild-type Flt3 mRNA expression ratio.31
The reasons for different FLT3-ITD expression levels remain unclear but could be influenced by the type of hematopoietic stem and or progenitor cell targeted by the two viruses, as well as different retroviral integration sites. As shown in Online Supplementary Figure S2, we found an overlapping pattern of retroviral integrations before transplantation and in resulting cells from the induced AML, suggesting in vivo expansion of a pre-selected clone. Two integration sites (in chromosomes 7 and 12) and one (chromosome 15) could be identified in experiments “A” and “B”, respectively. These different integration events could theoretically not only provide clonal advantage, but also result in different expression levels of the viral encoded transgenes.24 However, in contrast to FLT3-ITD, we found no significantly different expression levels of the NUP98-NSD1 fusion or its proposed downstream target the HoxA gene cluster (HoxA5, HoxA7, HoxA9, HoxA10) in cells from both experiments (Figure 5A).
Similar to what has been previously shown for AML co-expressing FLT3-ITD with MLL-AF9, AML1-ETO or NUP98-HOXD13, leukemic blasts expressing NUP98-NSD1 and FLT3-ITD were very sensitive to treatment with a small-molecule-inhibitor of FLT3 (Figure 5D). This observation further supports the importance of the FLT3-mediated cellular signaling in the development of the disease and underlines the need to explore selective new generation FLT3 inhibitors for patients with aggressive AML carrying NUP98-NSD1 and FLT3-ITD.
Acknowledgments
The authors would like to thank Radek Skoda for critically reading the manuscript. We also thank Sabine Juge and Ueli Schneider for technical support.
Footnotes
The online version of this article has a Supplementary Appendix.
Funding
This work was supported by the Swiss National Science Foundation (SNF-31003A-130661/1), the Swiss Cancer League (OCS 2357-02-2009), and the Swiss Bridge Foundation. J.S. was supported by the Gertrude Von Meissner Foundation Basel.
Authorship and Disclosures
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org
References
- 1.Riva L, Luzi L, Pelicci PG. Genomics of acute myeloid leukemia: the next generation. Front Oncol. 2012;1;2:40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Welch JS, Ley TJ, Link DC, Miller CA, Larson DE, Koboldt DC, et al. The origin and evolution of mutations in acute myeloid leukemia. Cell. 2012;150(2):264–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Walter MJ, Shen D, Ding L, Shao J, Koboldt DC, Chen K, et al. Clonal architecture of secondary acute myeloid leukemia. N Engl J Med. 2012;366(12):1090–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Gilliland DG, Griffin JD. The roles of FLT3 in hematopoiesis and leukemia. Blood. 2002;100(5):1532–42. [DOI] [PubMed] [Google Scholar]
- 5.Meshinchi S, Appelbaum FR. Structural and functional alterations of FLT3 in acute myeloid leukemia. Clin Cancer Res. 2009;15(13):4263–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kelly LM, Liu Q, Kutok JL, Williams IR, Boulton CL, Gilliland DG. FLT3 internal tandem duplication mutations associated with human acute myeloid leukemias induce myeloproliferative disease in a murine bone marrow transplant model. Blood. 2002;99(1):310–8. [DOI] [PubMed] [Google Scholar]
- 7.Kelly LM, Kutok JL, Williams IR, Boulton CL, Amaral SM, Curley DP, et al. PML/RARalpha and FLT3-ITD induce an APL-like disease in a mouse model. Proc Natl Acad Sci USA. 2002;99(12):8283–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Schessl C, Rawat VP, Cusan M, Deshpande A, Kohl TM, Rosten PM, et al. The AML1-ETO fusion gene and the FLT3 length mutation collaborate in inducing acute leukemia in mice. J Clin Invest. 2005;115(8):2159–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ono R, Nakajima H, Ozaki K, Kumagai H, Kawashima T, Taki T, et al. Dimerization of MLL fusion proteins and FLT3 activation synergize to induce multiple-lineage leukemogenesis. J Clin Invest. 2005;115(4):919–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Stubbs MC, Kim YM, Krivtsov AV, Wright RD, Feng Z, Agarwal J, et al. MLL-AF9 and FLT3 cooperation in acute myelogenous leukemia: development of a model for rapid therapeutic assessment. Leukemia. 2008;22(1):66–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Reckzeh K, Bereshchenko O, Mead A, Rehn M, Kharazi S, Jacobsen SE, et al. Molecular and cellular effects of oncogene cooperation in a genetically accurate AML mouse model. Leukemia. 2012;26(7):1527–36. [DOI] [PubMed] [Google Scholar]
- 12.Greenblatt S, Li L, Slape C, Nguyen B, Novak R, Duffield A, et al. Knock-in of a FLT3/ITD mutation cooperates with a NUP98-HOXD13 fusion to generate acute myeloid leukemia in a mouse model. Blood. 2012;119(12):2883–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Tam WF, Hähnel PS, Schüler A, Lee BH, Okabe R, Zhu N, et al. STAT5 is crucial to maintain leukemic stem cells in acute myelogenous leukemias induced by MOZ-TIF2. Cancer Res. 2013;73(1):373–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Grundler R, Miething C, Thiede C, Peschel C, Duyster J. FLT3-ITD and tyrosine kinase domain mutants induce 2 distinct phenotypes in a murine bone marrow transplantation model. Blood. 2005;105(12):4792–9. [DOI] [PubMed] [Google Scholar]
- 15.Grundler R, Brault L, Gasser C, Bullock AN, Dechow T, Woetzel S, et al. Dissection of PIM serine/threonine kinases in FLT3-ITD-induced leukemogenesis reveals PIM1 as regulator of CXCL12-CXCR4-mediated homing and migration. J Exp Med. 2009;206(9):1957–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chu SH, Heiser D, Li L, Kaplan I, Collector M, Huso D, et al. FLT3-ITD knockin impairs hematopoietic stem cell quiescence/homeostasis, leading to myeloproliferative neoplasm. Cell Stem Cell. 2012. 7;11(3):346–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Jaju RJ, Fidler C, Haas OA, Strickson AJ, Watkins F, Clark K, et al. A novel gene, NSD1, is fused to NUP98 in the t(5;11)(q35;p15.5) in de novo childhood acute myeloid leukemia. Blood. 2001;98(4):1264–7. [DOI] [PubMed] [Google Scholar]
- 18.Brown J, Jawad M, Twigg SR, Saracoglu K, Sauerbrey A, Thomas AE, et al. A cryptic t(5;11)(q35;p15.5) in 2 children with acute myeloid leukemia with apparently normal karyotypes, identified by a multiplex fluorescence in situ hybridization telomere assay. Blood. 2002;99(7):2526–31. [DOI] [PubMed] [Google Scholar]
- 19.Hollink IH, van den Heuvel-Eibrink MM, Arentsen-Peters ST, Pratcorona M, Abbas S, Kuipers JE, et al. NUP98/NSD1 characterizes a novel poor prognostic group in acute myeloid leukemia with a distinct HOX gene expression pattern. Blood. 2011;118(13):3645–56. [DOI] [PubMed] [Google Scholar]
- 20.Thol F, Kolking B, Hollink IH, Damm F, van den Heuvel-Eibrink MM, Michel Zwaan C, et al. Analysis of NUP98/NSD1 translocations in adult AML and MDS patients. Leukemia. 2012;27(3):750–4. [DOI] [PubMed] [Google Scholar]
- 21.Fasan A, Haferlach C, Alpermann T, Kern W, Haferlach T, Schnittger S. A rare but specific subset of adult AML patients can be defined by the cytogenetically cryptic NUP98-NSD1 fusion gene. Leukemia. 2013;27(1):245–8. [DOI] [PubMed] [Google Scholar]
- 22.Pigazzi M, Manara E, Bisio V, Aveic S, Masetti R, Menna G, et al. Screening of novel genetic aberrations in paediatric acute myeloid leukemia: a report from the AIEOP AML-2002 study group. Blood. 2012;120(18):3860–2. [DOI] [PubMed] [Google Scholar]
- 23.Wang GG, Cai L, Pasillas MP, Kamps MP. NUP98-NSD1 links H3K36 methylation to Hox-A gene activation and leukaemogenesis. Nat Cell Biol. 2007;9(7):804–12. [DOI] [PubMed] [Google Scholar]
- 24.Liu T, Jankovic D, Brault L, Ehret S, Baty F, Stavropoulou V, Rossi V, Biondi A, Schwaller J. Functional characterization of high levels of meningioma 1 as collaborating oncogene in acute leukemia. Leukemia. 2010;24(3):601–12. [DOI] [PubMed] [Google Scholar]
- 25.Demant P. Cancer susceptibility in the mouse: genetics, biology and implications for human cancer. Nat Rev Genet. 2003; 4(9):721–34. [DOI] [PubMed] [Google Scholar]
- 26.Williams RT, Sherr CJ. The INK4-ARF (CDKN2A/B) locus in hematopoiesis and BCR-ABL-induced leukemias. Cold Spring Harbor Laboratory Press; 2008;73:461–7. [DOI] [PubMed] [Google Scholar]
- 27.Lavau C, Szilvassy SJ, Slany R, Cleary ML. Immortalization and leukemic transformation of a myelomonocytic precursor by retrovirally transduced HRX-ENL. EMBO J. 1997;16(14):4226–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.So CW, Karsunky H, Wong P, Weissman IL, Cleary ML. Leukemic transformation of hematopoietic progenitors by MLL-GAS7 in the absence of Hoxa7 or Hoxa9. Blood. 2004;103(8):3192–9. [DOI] [PubMed] [Google Scholar]
- 29.Kroon E, Thorsteinsdottir U, Mayotte N, Nakamura T, Sauvageau G. NUP98-HOXA9 expression in hemopoietic stem cells induces chronic and acute myeloid leukemias in mice. EMBO J. 2001;20(3):350–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Gough SM, Slape CI, Aplan PD. NUP98 gene fusions and hematopoietic malignancies: common themes and new biologic insights. Blood. 2011;118(24):6247–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Akiki S, Dyer SA, Grimwade D, Ivey A, Abou-Zeid N, Borrow J, Jeffries S, Caddick J, et al. NUP98-NSD1 fusion in association with FLT3-ITD mutation identifies a prognostically relevant subgroup of paediatric acute myeloid leukemia patients suitable for monitoring by real time quantitative PCR. Genes Chromosomes Cancer. 2001; 52(11):1053–64. [DOI] [PubMed] [Google Scholar]