

Abstract
Advanced glycation end products (AGEs) constitute a complex group of compounds produced endogenously during the aging process and under conditions of hyperglycemia and oxidative stress. AGEs also have an emerging exogenous origin. Cigarette smoke and diet are the two main exogenous sources of AGEs (glycotoxins). Modern Western diets are rich in AGEs which have been implicated in the pathogenesis of several metabolic and degenerative disorders. Accumulating evidence underlies the beneficial effect of the dietary restriction of AGEs not only in animal studies but also in patients with diabetic complications and metabolic diseases. This article reviews the evidence linking dietary glycotoxins to several disorders from diabetic complications and renal failure to liver dysfunction, female reproduction, eye and cognitive disorders as well as cancer. Furthermore, strategies for AGE reduction are discussed with a focus on dietary modification.
Keywords: AGEs, dietary glycotoxins, dietary restriction, PCOS, MSR-1, RAGE
Introduction
Advanced glycation end products (AGEs) represent a complex group of compounds derived from the nonenzymatic glycation of proteins, lipids, and nucleic acids formed endogenously but also from exogenous supplies (also referred as glycotoxins).1 Tobacco smoke2 and certain foods rich in glycotoxins that are formed by specific cooking methods3 represent the two major exogenous sources of glycotoxins. The pool of body AGEs reflects the balance between endogenous production and exogenous intake, with the effective mechanisms of AGE detoxification systems as well as their excretion from the kidneys.4,5
AGEs are heterogeneous molecules (Figure 1) that share some common characteristics including the formation of covalent cross-links between proteins, the effect of turning some foodstuffs a yellow-brown color (the ‘browning’ effect), and the ability to generate fluorescence. They have diverse chemical structures but have a common characteristic lysine residue in their molecule. Based on their properties, AGEs can be categorized as: (a) fluorescent and cross-linking AGEs, including pentosidine, crossline, 2-(2-furoyl)-4(5)-(2-furanyl)-1H-imidazole, glyoxal-lysine dimer, and methylglyoxal-lysine dimer (MOLD); and (b) nonfluorescent and non-cross-linking agents such as N-carboxymethyl lysine (CML), N3-(carboxyethyl)lysine, and pyrraline.6,7
Figure 1.
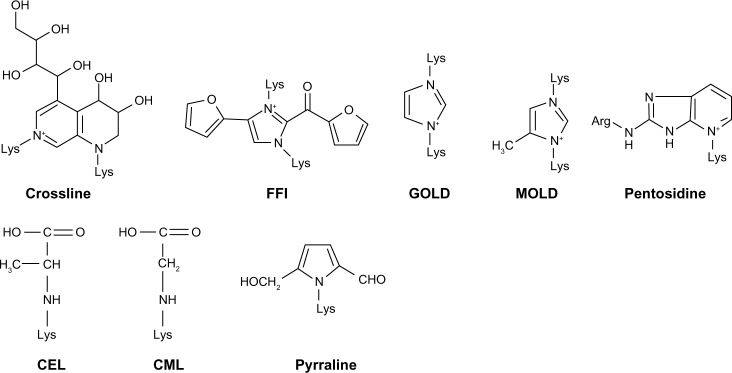
Chemical structure of some advanced glycation end products (AGEs): crossline, 2-(2-furoyl)-4(5)-(2-furanyl)-1H-imidazole (FFI), glyoxal-lysine dimer (GOLD), methylglyoxal-lysine dimer (MOLD), pentosidine, N3-(carboxyethyl)lysine (CEL), N-carboxymethyl lysine (CML), and pyrraline.
AGEs’ actions are mainly mediated through two classes of AGE receptors with different biological effects (Figure 2). The receptors responsible for their clearance are macrophage scavenger receptor-1 (MSR-1) and the AGE-specific receptors AGE-R1, AGE-R2, and AGE-R3. These receptors are responsible for maintaining AGE homeostasis through regulation of their degradation and removal. The multi-ligand receptor for advanced glycation end products (RAGE) mediates pro-atherogenic, inflammatory, and immune responses via activation of nuclear factor-kappa B (NF-κB), resulting in the increased expression/synthesis of proinflammatory cytokines such as interleukin (IL)-6, IL-1a, and tumor necrosis factor-alpha.6 Furthermore, activation of NF-κB leads to increased reactive oxygen species (ROS) production1,8,9 through molecules mediating inflammation, vasoconstriction, and coagulation such as vascular cell adhesion molecule-1, endothelin-1, and thrombomodulin (Figure 2).6 Apart from the AGE receptor-mediated mechanisms, AGEs can form cross-links with basic components in the basement membrane of the extracellular matrix, modifying its structural characteristics (Figure 2).10
Figure 2.

Schematic representation of advanced glycation end product (AGE) mechanisms of action.
Notes: Receptor-mediated mechanisms include two different types of receptors: receptors responsible for AGE degradation/detoxification – namely, MSR and AGE-specific receptors – and the multi-ligand receptor for AGEs – receptor for advanced glycation end products (RAGE) – that mediates pro-atherogenic, inflammatory, and immune responses. Receptor-independent mechanisms of action include formation of cross-links with basic components in the basement membrane of the extracellular matrix permanently modifying its structural characteristics.6,10
Abbreviations: IL-1a, interleukin-1a; IL-6, interleukin-6, MSR, macrophage scavenger receptor; NF-κB, nuclear factor-kappa B, RAGE, receptor for advanced glycation end products; TNF-α, Tumor necrosis factor-alpha; VCAM-1, vascular cell adhesion molecule-1.
When defense mechanisms are damaged, as in patients with diabetes mellitus, or when inflammatory mechanisms are upregulated, as in tissues in which AGEs are accumulated,11 the deleterious impact of AGEs prevails. This involves pro-oxidant and inflammatory effects that increase intracellular oxidative stress and ROS production; therefore, chronic exposure to AGEs may be implicated in a broad spectrum of diseases through inflammation and propagation of tissue damage.
This review focuses on the impact of dietary glycotoxins in the pathogenesis of several diseases. We also review therapeutic management, focusing mainly on food restriction, which appears to have beneficial effects on the adverse effects of these harmful molecules.
Dietary glycotoxins
Approximately 10% of consumed AGEs are absorbed though the intestine. One-third of the absorbed AGEs are excreted within 48 hours in the urine, whereas the other two-thirds of those absorbed remain in the tissues.12 The exogenous formation of AGEs takes place spontaneously in food under specific cooking methods – predominantly, cooking at high temperature of short duration. Nutrient composition (high lipid and protein content), presence or absence of moisture, and pH are also parameters that affect AGE content. For instance, in dry-heat cooking (grilling, frying, roasting, baking, and barbecuing) over short periods of time, more AGEs are formed than in high-moisture cooking (boiling, steaming, poaching, stewing, and slow cooking) over longer periods of time. Table 1 presents the AGE contents of selected food items from a large database provided by Goldberg et al.3
Table 1.
Advanced glycation end product (AGE) content in commonly consumed foodsa
| Food item | AGE contentb |
|---|---|
| Salmon, raw (90 g) | 502 |
| Salmon, broiled (10 minutes, 90 g) | 1,348 |
| Beef, boiled (1 hour, 90 g) | 2,000 |
| Beef, broiled (15 minutes, 90 g) | 5,367 |
| Beef, stir fried (20 minutes) and broiled (15 minutes, 90 g) | 6,166 |
| Chicken, boiled (1 hour, 90 g) | 1,011 |
| Chicken, broiled (15 minutes, 90 g) | 5,245 |
| Beans, red kidney, raw (100 g) | 116 |
| Beans, red kidney, canned (100 g) | 191 |
| White potato, boiled (25 minutes, 100 g) | 17 |
| French fries (100 g) | 1,522 |
| Broccoli (100 g) | 226 |
| Tomato (100 g) | 23 |
| Apple (100 g) | 13 |
Notes:
Data derived from Goldberg et al;3
AGE content denotes kilounits per serving; AGE measured by enzyme-linked immunosorbent assay using an antibody against N-carboxymethyl lysine.
It seems obvious that the modification of cooking methods is a sufficient way of reducing a meal’s AGE content. For instance, cooking foods at low temperatures for long time periods in the presence of higher water content – boiling, steaming, poaching, stewing – minimizes the AGE content, compared with broiling or frying. This is of great significance given that dietary AGEs have been directly associated with serum and tissue AGE levels in healthy individuals as well as in patients suffering from diabetes and chronic kidney disease.12–14 This was even more profound in a study of 26 patients with renal failure undergoing peritoneal dialysis who followed a diet with high versus low AGE content for a short period of time (4 weeks).15 A significant reduction in serum AGE levels was observed in the group following a low AGE-content diet, whereas in the group on a high AGE diet, a significant increase in serum AGE levels was reported.
Nowadays, conventional diets are considered to provide 25 to 75 mg of AGEs, estimated mainly with pyrraline and N-carboxymethyl lysine.16 Vlassara et al13 and Uribarri et al15 reported a significant reduction in serum AGE levels of diabetics as well as renal failure patients due to diets low in dietary glycotoxins. In a human study analyzing the metabolism of Amadori products, only approximately 5% was recovered from the urine and feces.17
These data underlie the contribution of nutritional AGEs to total body burden and, furthermore, imply a possible mechanism of reducing circulating AGE levels through dietary interventions.
AGE metabolism and excretion
The kidney is the organ predominantly responsible for AGE removal. Here the filtered AGEs are partially degraded in the tubule system, while those remaining are excreted in the urine.18 Renal impairment leads to decreased clearance of AGEs and their accumulation in tissues,19,20 while their plasma levels progressively rise. AGEs are shown to affect almost all renal components,21,22 with the proximal tubule responsible for AGE catabolism.23 Data from experimental animal and human studies have confirmed the crucial role of the kidneys in the metabolic regulation and clearance of glycotoxins.12,15,24
It has been shown that AGEs in the kidney originate either from the circulation, are generated from intrarenal proteins, or are due to impaired AGE-removal mechanisms. The role of the kidney in trapping AGEs was shown after chronic intravenous infusion of AGE (CML)-modified rat albumin in normal rats. The AGE content of the kidneys increased by 50% in the treated animals compared with in the control ones.25 Interestingly, this process resulted in widening of the basement membrane, increased glomerular tuft volume, mesangial extracellular matrix expansion, glomerulosclerosis, and albuminuria.25
In addition to renal tissues, the liver is also a site of metabolism and degradation of AGEs, predominantly through a scavenger receptor mechanism expressed in hepatic sinusoidal cells and to lesser extent Kupffer cells.26,27 After the intravenous administration of AGE-modified bovine serum albumin in a rat experimental model, Smedsrød et al demonstrated a rapid and almost exclusive accumulation in liver endothelial cells (60% uptake) and to lesser extent Kupffer cells (25% uptake), while parenchymal cells accounted only for approximately 10%–15% of hepatic deposition.26 This endocytic uptake is mediated by a scavenger receptor27 whose function has been shown to be negatively modulated by AGEs.28 Besides its pivotal role in AGE degradation, the liver has the potency to generate low levels of inflammatory molecules, causing oxidative stress and, in the case of AGEs, is presumed to contribute to inflammatory processes through the activation of RAGE after interaction with its ligands (AGEs) in hepatocytes and hepatic stellate cells.29
Implications for health
Nowadays, AGEs are implicated in the pathogenesis of several disorders from diabetic complications and renal failure to female reproduction, liver disorders, neurodegenerative, and eye diseases as well as cancer. The pathological role of food-derived AGEs in various types of disorders is analyzed following.
AGEs and diabetes
It is now well-documented that AGEs are related to the progression of diabetic complications including diabetic nephropathy, diabetic peripheral neuropathy, diabetic cardiomyopathy and peripheral arterial disease, diabetic ocular disease, and atherosclerotic disease.30 Dietary glycotoxins have been shown experimentally to contribute to the development of type 2 (T2D) diabetes,5 and animal studies with rodents following a high AGE diet for a long period of time document the induction of insulin resistance and T2D.31,32
In diabetes patients, restriction of dietary-glycotoxin intake has been shown to decrease inflammatory markers,13 improve insulin resistance, and lower plasma insulin levels.33 Furthermore, it has been associated with lower leptin and increased adiponectin levels, as well as altered AGE-receptor expression (decreased RAGE and increased AGE-R1 expression),5 while impaired vascular and endothelial function has been reported following a high AGE meal.34,35
AGEs and kidney disease
Chronic kidney disease and AGEs participate in what has been termed a “vicious cycle”, as this condition is associated with increased oxidative and carbonyl stress – that is, the production of reactive carbonyl compounds and AGEs. AGEs and other carbonyl-modified proteins, in turn, contribute to the further decline of renal function.36 This vicious cycle is implied, as in patients with end-stage renal disease, serum AGE levels are elevated,37,38 and, in turn, AGEs contribute to further decrease of the glomerular filtration rate. Several studies have demonstrated the mechanisms via which AGEs can damage the kidneys, either through AGE–RAGE interaction, deposition of AGEs, or by in situ glycation.39,40
Kidney disease has been linked to the decreased excretion of an oral load of AGEs, resulting in an inverse correlation between renal function and serum levels of AGEs.41 AGE-induced renal injury is due to renal cellular-signaling alterations, mainly through the interaction of AGEs with their receptor, RAGE. Protein kinase C, mitogen-activated protein kinase, and NF-κB are some of the signaling molecules involved in the tissue injury related to AGE pathways.39 In addition, tubular expression of connective tissue growth factor via the tissue growth factor-beta-independent RAGE-ERK/p38-mitogen-activated protein kinase-Smad3 cross-talk pathway is another way through which AGEs are involved in renal injury.40 Moreover, AGEs directly affect the structural integrity of the renal tissue through the cross-linking of matrix proteins (collagen).42,43
In podocytes, the AGE–RAGE interaction induces the upregulation of monocyte chemoattractant protein-1 expression, while MCP-1 gene transcription has been shown to be regulated by NF-κB and SP1 protein, and this interaction results in the generation of intracellular ROS.44
Dietary AGEs have also been shown to contribute to kidney disease, as demonstrated in studies of patients with renal failure with dietary restriction of glycotoxin intake. In patients with renal failure, dietary glycotoxins were positively associated with high AGE serum levels.14 Thus, dietary restriction of AGEs may contribute to the reduction of AGE-related renal injury and associated mortality, through several mechanisms including the reduction of oxidative stress and inflammation.
Moreover, a study which included a female rat model that exerted the metabolic and hormonal characteristics of women with hyperandrogenemia, suggested that dietary glycotoxins, in combination with increased androgen exposure, exert a more profound negative impact on the kidney.45 In addition, the study demonstrated that dietary glycotoxins and androgen excess induce an inflammatory environment for the kidney, which could further aggravate its structure and function.45
AGEs and liver disease
Several human and experimental studies have shown an association between AGEs with several hepatic disorders from simple steatosis, and biochemical aberrations to hepatic cirrhosis.29,46–48 In patients with nonalcoholic steatohepatitis, AGEs were histochemically documented in the liver, and serum levels of AGEs were related to the severity of liver dysfunction.48
Experimental studies of AGE administration in rodents confirm their increased liver tissue deposition parallel to an increase in the hepatic expression of RAGE and vascular endothelial growth factor (VEGF), which is implicated in hepatic fibrosis.49 In another study of male mice, the long-term administration of a low AGE-content diet was related to enhanced expression of AGE-R1 and decreased RAGE expression in the liver tissue parallel to a beneficial effect on oxidative stress and extended lifespan, compared to rodents following an isocaloric diet of standard AGE content.50 Furthermore, in a mouse model of high-versus-regular AGE-content diet, signs of liver inflammation were observed in the high AGE content diet subgroup.51
AGEs and polycystic ovary syndrome (PCOS)
The elevated levels of serum AGEs observed in PCOS women associated positively with insulin-resistance indices, testosterone, and anti-Müllerian hormone levels when compared with healthy age- and body mass index-matched women.52,53 This is a unique characteristic in women with PCOS as opposed to those who experience only some of the clinical features of the syndrome.54 These observations, combined with the immunohistochemical evidence of increased AGE deposition in human polycystic ovaries,55 imply a possible direct impact of AGEs in the ovarian physiology of women with PCOS.
Dietary AGEs are also considered to have an impact on PCOS pathophysiology, since evidence has been provided from animal studies with AGE-enriched diets. In those experiments, animals fed with enriched AGE diets had an increased immunochemically documented accumulation of AGEs in ovarian tissue and elevated serum levels and this was associated with an impaired hormonal and metabolic profile expressed as elevated insulin and androgen levels, compared to animals fed a low AGE-content diet.56,57 The potential involvement of dietary AGEs to PCOS pathophysiology is implied from a recent study of women with the syndrome who followed specific, consecutive, nutritional interventions for short period of time based on diet AGE content (high and subsequently low AGE content).58 As shown by researchers, a disturbed metabolic and hormonal profile expressed as elevated insulin and testosterone and increased markers of insulin resistance and oxidative stress was observed in women with PCOS following the diet high in AGE content compared to preceding levels during a regular hypocaloric diet.58 On the other hand, the low AGE diet seemed to have a beneficial effect on oxidative stress.58 Along these lines, in vivo studies of PCOS-like animal models, PCOS-like models, fed with high AGE diets showed that androgens and dietary AGEs have a synergistic effect on the intra-ovarian detoxifying system operated by glyoxalase-1, resulting in the increased deposition of AGEs in ovarian tissues.57 In addition, a high AGE diet compromises the expression of the scavenger receptor in peripheral blood monocytes and therefore interferes with the clearance mechanisms and subsequently contributes to loading the tissues with AGEs.59
A recent study of human granulosa KGN cells cultured with variable concentrations of human glycated albumin or insulin revealed a novel interference between AGEs with insulin intracellular signaling and glucose transport in the ovarian microenvironment.60 The AGEs showed a direct inhibitory effect on insulin signaling in the human granulosa cells, resulting in the prevention of the membrane translocation of GLUT4. Therefore, intra-ovarian AGE accumulation, from endogenous or exogenous sources, may contribute to the pathophysiology of states characterized by anovulation and insulin resistance such as PCOS.60
These in vitro observations may prove to be even more important on clinical grounds, since AGEs have been shown to be present in the umbilical cord blood. In pregnant women with PCOS and with gestational diabetes, the mother’s AGE serum levels have been shown to correlate with the infant’s levels as well as with other oxidative molecules, findings which suggest that the mother’s oxidative stress affects the infant’s environment.61
AGEs and neurological disorders
AGEs have been related to the pathogenesis of several neurological degenerative diseases, including Alzheimer’s disease, Parkinson’s disease, and amyotrophic lateral sclerosis as well as peripheral diabetic polyneuropathies.62 The pathophysiological mechanism involved is believed to be related to pathological amyloid glycation and the induction of oxidative stress leading to potential neurotoxic effects.62
A probable involvement of oral AGEs in the initiation and progression of cognitive aberrations is proposed by animal data showing systemic and brain changes in old mice following chronic exposure to a neurotoxic methyl-glyoxal diet.63 Moreover, these effects are probably mediated through an inhibitory effect on Sirtuin 1 (SIRT1) gene expression that has been shown to be related to cognitive decline and Alzheimer’s disease. In humans, a reduced intake of food-derived AGEs has been considered an effective strategy to prevent neurodegenerative diseases given that in subjects over 60 years old an association has been shown between high serum AGEs and cognitive decline.63
AGEs and eye diseases
AGEs are also involved in vision loss, and have been linked with macula degeneration, cataract formation, diabetic retinopathy, and glaucoma.64
The pathogenetic mechanisms involved include AGE accumulation in ocular tissues where they mediate the aberrant cross-linking of extracellular matrix proteins and the disruption of endothelial junctional complexes that affects cell permeability and mediates angiogenesis. Moreover, AGEs severely affect cellular metabolism by disrupting adenosine triphosphate production, enhancing oxidative stress, and modulating the gene expression of anti-angiogenic and anti-inflammatory genes.64 A recent study demonstrated the direct impact of a high AGE diet on the ocular tissues of normal rats of different ages.65 In that study, it was demonstrated that increased peripheral AGE levels were positively correlated with significant increased tissue immunoreactivity of AGEs and RAGE in retinal and uveal tissues associated with increased retinal VEGF-A expression.65 Furthermore the upregulation of RAGE and VEGF-A expression was observed in the ocular tissue of both baby and adult animals fed with a high AGE content diet, suggesting that dietary AGEs can have negative effects on eye tissues regardless of age.65
AGEs and cancer
The potential involvement of AGEs in cancer risk, development, and disease progression is largely undetermined and remains to be elucidated. In vitro studies of breast, prostate, and lung cancer cell lines imply a possible contribution of AGEs to cancer proliferation, migration, invasion, and survival.66–68 Human studies are yet inconclusive but significant observations have been reported. Detection of AGEs has been reported in various human tumors69 and, interestingly, a higher accumulation of AGEs has been observed in malignant tissues (such as prostate cancer) compared to benign ones.70 Some epidemiologic studies suggest an inverse association between soluble AGEs and the risk of pancreatic71 and colorectal cancer,72 while other studies have failed to report similar effects.73 A suggested molecular link between glycotoxins and cancer biology involves the activation of an inflammatory cycle of persistent oxidative stress that favors cancer development.74
Strategies for AGE reduction
As previously described, accumulating evidence from animal studies as well as clinical studies in diabetic patients and healthy individuals converge on the observation that the dietary intake of foods with a high AGE content predisposes to inflammation and deteriorating biological effects, whereas AGE restriction seems to have an opposite, beneficial impact. In line with this, several researches have been conducted over the last decades to investigate possible strategies to influence the AGE pathway (Table 2). However, despite the wide range of promising pharmacologic agents for AGE reduction, like blockage of AGE formation, cross-link breakers, RAGE blockade, and others, human studies are scanty and are in an experimental state, including clinical trial studies, therefore their clinical application has not been established. Most experimental data – both in vitro and in vivo – as well as clinical studies, converge to suggest the significance of dietary AGE restriction. Therefore, the current strategy to avoid the body accumulation of AGEs is mainly to decrease their intake (via diet restriction and tobacco avoidance or cessation).
Table 2.
Interventions targeting the advanced glycation end product (AGE) pathway
| Mechanism of action | Therapeutic agent | Biological effects |
|---|---|---|
| Dietary restriction of AGE | Animal studies31,50,75–77 | |
| Prevents/Improves | ||
| Insulin resistance | ||
| Abdominal obesity/body weight | ||
| Diabetes mellitus type 2 | ||
| Diabetic nephropathy | ||
| Diabetic-impaired wound healing | ||
| Extends life span | ||
| Human studies | ||
| Healthy subjects78–80 | ||
| ↓ Basal oxidative stress | ||
| ↓ Inflammation | ||
| ↓ Serum AGEs | ||
| Improves markers of insulin resistance | ||
| Subjects with diabetes or renal failure13,15,34 | ||
| ↓ Serum AGEs | ||
| ↓ Inflammation | ||
| Improves vascular function and insulin resistance | ||
| Blockage of AGE formation | Aminoguanidine83–85 | Animal studies |
| ↓ Retinopathy | ||
| ↓ Nephropathy | ||
| ↓ Neuropathy | ||
| N-(2-Acetamido Methyl) hydrazinecarboximide amide hydrochloride (ALT-946)86,87 | ↓ Nephropathy | |
| 3-benzyloxycarbonylmethyl-4-methyl-thiazol-3-ium bromide (C36)88 | ↓ Diabetic cardiovascular dysfunction | |
| Pyridoxamine90,91 | ↓ Nephropathy | |
| ↓ Dyslipidemia | ||
| Cross-link breakers | N-phenacylthiazolium (PTB)92 | Animal studies |
| ↓ AGE | ||
| Alagebrium (ALT-711)93,94,96 | Animal studies | |
| ↓ Diabetic cardiomyopathy and atherosclerosis | ||
| ↓ Nephropathy | ||
| Human studies | ||
| ↓ Diastolic heart function | ||
| RAGE blockade | sRAGE100–103 | Animal studies |
| ↓ Diabetic atherosclerosis | ||
| and retinopathy (sRAGE) | ||
| Antihypertensive drugs99 | Antioxidative properties – prevention of diabetic vascular complications? | |
| PEDF99 | Antioxidative properties | |
| Statins99 | Antioxidative properties | |
| Bisphosphonates99 | Antioxidative properties | |
| PARP inhibitors104,105 | Animal studies | |
| ↓ Early peripheral diabetic neuropathy Improve endothelial and myocardial function | ||
| Other agents | Kremezin (AST-120)106 | ↓ AGE levels in chronic renal failure |
Abbreviations: PARP, poly(ADP-ribose) polymerase; PEDF, pigment-epithelium-derived factor; RAGE, receptor for advanced glycation end products; sRAGE, soluble receptor for advanced glycation end products; ↓, decrease/reduce.
Dietary restriction of AGEs
Dietary restriction of the consumption of AGEs from exogenous sources through modifications in eating habits may reflect an initial, effective therapeutic intervention in the management of several AGE-related disorders as indicated, so far, by many animal and human studies of dietary AGE restriction.15,75–77
In experimental studies, a reduction in the intake of exogenous, food-derived glycotoxins has been linked to a beneficial effect on insulin sensitivity, abdominal obesity, and body weight regulation, and, furthermore, has been shown to have a protective effect against T2D.76,77 Moreover, in diabetic rodents, dietary AGE restriction was shown to improve diabetic complications (macroangiopathy vascular disease, diabetic nephropathy, recover healing ability)31,50,75–77 and has been related to prolonged life expectancy – probably through reduced oxidant stress and organ dysfunction.50
AGE restriction in individuals with diabetes and kidney failure for a short period of time decreases inflammation and has a beneficial effect on the endothelium and insulin sensitivity.13,15,34 In healthy individuals, lower AGE intake has been related to lower serum AGE levels78 and improvement in basal oxidative status, inflammation,79 and insulin-resistance markers.80 It should be highlighted that, as far as we are aware, AGE restriction studies in humans so far have been performed for relatively short periods of time (approximately 4 weeks),78–80 and clinical studies exploring long-term effects are missing.
In conclusion, accumulating evidence suggests the modification of dietary habits as an effective strategy for the alleviation of many AGE-related disorders; however, the evidence, though highly suggestive, is still insufficient to prompt precautions.81 The restriction of glycotoxin consumption and the recognition of foods with high AGE contents require constant communication between the scientific community, authorities, and public in order to inform consumers about the potential hazardous effects of AGE consumption and reinforce the importance of reducing dietary AGEs.
General practices for lowering dietary AGE intake (Table 3) include the consumption of foods with low AGE contents (such as fruits and vegetables, seafood, whole grains, bread, low-fat milk) while avoiding or reducing the consumption of high AGE content food (including sugary items such as candy, cookies, and beverages; highly processed foods, such as packaged meats, cheese, and snack-type foods; and fats, including butter, full-fat cheeses, and fried foods), adaptation of specific cooking methods (such as cooking foods at low temperatures with high moisture – boiling, steaming, poaching, stewing – for a relatively long period of time) and adoption of healthy lifestyle habits (such as exercise, maintaining normal body weight, and cessation of tobacco consumption).
Table 3.
Recommendations for lowering advanced glycation end product (AGE) consumption
| Recommendations | Permit | Avoid |
|---|---|---|
| Foods | Fresh fruits and vegetables Seafood Whole grains Bread Low-fat milk | Sugary items such as candy, cookies, and beverages Processed foods such as packaged meats, cheese, and snack-type foods High-fat meats Fats, such as butter, margarine, full-fat cheeses Fried foods |
| Cooking methods | Low temperatures for long time periods Presence of high water content: boiling, steaming, poaching, stewing | High temperatures for short time periods Dry heat: grilling, frying, roasting, baking, and barbecuing |
| Lifestyle modifications | Exercise Normal weight | Obesity Tobacco smoke |
Blockage of AGEformation
Blockage of AGE formation has been considered as a potential anti-AGE therapeutic strategy targeting the initial steps of AGE formation. Aminoguanidine was first studied as a compound with potential AGE-inhibiting properties82 and has been shown to be effective in preventing diabetic complications in various animal studies, such as those on retinopathy83 and nephropathy.84 Further, it has been shown to ameliorate neuropathy even in normal rodents.85
N-(2-Acetamidoethyl) hydrozinecarboximidamide hydrochloride (ALT-946) is another compound that has been studied with reference to its renal protective effects,86,87 while 3-benzyloxycarbonylmethyl-4-methyl-thiazol-3-ium bromide (C36) was shown to improve markers of cardiovascular system function in a group of streptozotocin-induced diabetic rats.88 Hydroxymethylglutaryl-coenzyme A reductase inhibitors (statins) and pyridoxamine, the natural form of vitamin B6, are among the substances shown to be effective at inhibiting AGEs in diverse ways.89–91
Putative cross-link breakers
Breaking collagen cross-links has been thought a possible therapeutic approach against AGE-provoked unfavorable biological outcomes. The first study on AGE breakers with a rational design was published in 1996 by Vasan et al.92 The strategy was based on the design of compounds able to covalently react and then cleave the alpha-dicarbonyl derivatives which were proposed as intermediates of AGE formation. The prototype was identified as N-phenacylthiazolium and was shown to cleave cross-links.92 A consecutive derivative of N-phenacylthiazolium (alagebrium [3-phenyacyl-4,5-dimethyl-thiazolium chloride] or ALT-711) has been examined in view of its ability to attenuate diabetes-associated myocardial structural abnormalities in rats.93 In a series of studies with diabetic animal models, alagebrium was demonstrated to have a beneficial effect on diabetic complications – renal disease, diabetic cardiomyopathy, and atherosclerosis.94 AGE breakers have been shown to improve arterial compliance95 and several parameters of cardiac function in patients with stable diastolic heart failure.96
RAGE blockade
Blockage of the ligand–RAGE axis is another promising therapeutic approach based on the fact that a significant part of oxidative stress in cells is provoked by exposure to AGEs via RAGE. Blockage of this axis can be achieved either by inactivating RAGE using high-molecular-weight substrate analogs, low-molecular-weight inhibitors, or anti-RAGE antibodies, or by inhibiting the signal transduction pathway that follows the ligand–RAGE interaction.97 Substances believed to have a suppressive effect on RAGE expression are several categories of antihypertensive drugs (calcium-channel blockers, angiotensin converting enzyme inhibitors, angiotensin II receptor blockers), antidiabetic agents (thiazolidinediones), and cholesterol-lowering drugs (statins).98 Theoretically, the group of agents that could potentially suppress the RAGE axis consists of the soluble form of RAGE (soluble receptor for advanced glycation end products [sRAGE]); antihypertensive drugs – telmisartan, ramipril, olmesartan, candesartan, losartan, valsartan, nifedipine; pigment epithelium-derived factor; statins, inhibitors of cholesterol synthesis; and nitrogen-containing bisphosphonates used as anti-osteoporotic drugs.99 The beneficial effects of sRAGE on the development and progression of diabetic atherosclerosis have been documented in animal models of diabetic apolipoprotein E-null mice.100,101 Furthermore, a favorable effect of sRAGE administration on diabetic retinopathy has also been described.102,103
Other agents
Poly(ADP-ribose) polymerase (PARP) inhibitors have been shown to ameliorate early peripheral diabetic neuropathy as well as endothelial and myocardial function in experimental studies of laboratory animals.104,105
Another possible therapeutic approach against the AGE pathway is through agents inhibiting glycotoxin absorption. An example of such is provided by AST-120, an oral adsorbent, which has been shown to exert a beneficial effect on chronic kidney disease by eliminating uremic toxins.106 Furthermore, its ability to lower serum AGE levels by binding to CML has been reported in patients with chronic renal failure.106
Orlistat, an inhibitor of intestinal lipases, has been shown to have a beneficial effect on serum AGE levels and metabolic profile in obese women with PCOS and in obese control women.107 Finally, inhibitors of advanced glycation acting as potent endoplasmic reticulum stress modulators have been reported to have beneficial effects on metabolic dysfunction.108
Conclusion
Dietary AGEs represent a class of proinflammatory and oxidative stress-promoting agents implicated not only in normal aging and diabetes, but also in a wide spectrum of diseases. A common pathogenetic mechanism in many AGE-related disorders is the induction or deterioration of oxidative stress that is mainly mediated through the activation of the AGE–RAGE axis and/or through the suppression of internal defensive systems. Furthermore, chronic exposure to dietary AGEs is speculated to act as an additional, aggravating factor for tissue injury and deleterious biological effects. In the modern society, AGE consumption – as component of modern Westernized fast and high-temperature cooked diets – is excessive. Therefore, AGE restriction is now recognized as a necessary intervention. Prospective studies are needed to clarify whether dietary AGEs significantly contribute to oxidant stress. Furthermore, it is of great significance to determine whether the modulation of dietary AGE content has beneficial effects on certain clinical conditions. Restriction of food-derived AGEs through diet and tobacco consumption reflects an efficient approach to avoid body accumulation of AGEs. It remains to be proven whether therapeutic agents targeting AGE formation, AGE cross-linking, and AGEs–RAGE interaction will be approved for use in clinical practice.
Footnotes
Disclosure
The authors declare no conflicts of interest in this work.
References
- 1.Vlassara H, Palace MR. Diabetes and advanced glycation endproducts. J Intern Med. 2002;251(2):87–101. doi: 10.1046/j.1365-2796.2002.00932.x. [DOI] [PubMed] [Google Scholar]
- 2.Cerami C, Founds H, Nicholl I, et al. Tobacco smoke is a source of toxic reactive glycation products. Proc Natl Acad Sci U S A. 1997;94(25):13915–13920. doi: 10.1073/pnas.94.25.13915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Goldberg T, Cai W, Peppa M, et al. Advanced glycoxidation end products in commonly consumed foods. J Am Diet Assoc. 2004;104(8):1287–1291. doi: 10.1016/j.jada.2004.05.214. [DOI] [PubMed] [Google Scholar]
- 4.Vlassara H, Uribarri J, Cai W, Striker G. Advanced glycation end product homeostasis: exogenous oxidants and innate defenses. Ann N Y Acad Sci. 2008;1126:46–52. doi: 10.1196/annals.1433.055. [DOI] [PubMed] [Google Scholar]
- 5.Vlassara H, Striker GE. Advanced glycation endproducts in diabetes and diabetic complications. Endocrinol Metab Clin North Am. 2013;42(4):697–719. doi: 10.1016/j.ecl.2013.07.005. [DOI] [PubMed] [Google Scholar]
- 6.Ahmed N. Advanced glycation endproducts – role in pathology of diabetic complications. Diabetes Res Clin Pract. 2005;67(1):3–21. doi: 10.1016/j.diabres.2004.09.004. [DOI] [PubMed] [Google Scholar]
- 7.Wu CH, Huang SM, Lin JA, Yen GC. Inhibition of advanced glycation endproduct formation by foodstuffs. Food Funct. 2011;2(5):224–234. doi: 10.1039/c1fo10026b. [DOI] [PubMed] [Google Scholar]
- 8.Bierhaus A, Stern DM, Nawroth PP. RAGE in inflammation: a new therapeutic target? Curr Opin Investig Drugs. 2006;7(11):985–991. [PubMed] [Google Scholar]
- 9.Yonekura H, Yamamoto Y, Sakurai S, Watanabe T, Yamamoto H. Roles of the receptor for advanced glycation endproducts in diabetes-induced vascular injury. J Pharmacol Sci. 2005;97(3):305–311. doi: 10.1254/jphs.cpj04005x. [DOI] [PubMed] [Google Scholar]
- 10.Goldin A, Beckman JA, Schmidt AM, Creager MA. Advanced glycation end products: sparking the development of diabetic vascular injury. Circulation. 2006;114(6):597–605. doi: 10.1161/CIRCULATIONAHA.106.621854. [DOI] [PubMed] [Google Scholar]
- 11.Stern D, Yan SD, Yan SF, Schmidt AM. Receptor for advanced glycation endproducts: a multiligand receptor magnifying cell stress in diverse pathologic settings. Adv Drug Deliv Rev. 2002;54(12):1615–1625. doi: 10.1016/s0169-409x(02)00160-6. [DOI] [PubMed] [Google Scholar]
- 12.Koschinsky T, He CJ, Mitsuhashi T, et al. Orally absorbed reactive glycation products (glycotoxins): an environmental risk factor in diabetic nephropathy. Proc Natl Acad Sci U S A. 1997;94(12):6474–6479. doi: 10.1073/pnas.94.12.6474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Vlassara H, Cai W, Crandall J, et al. Inflammatory mediators are induced by dietary glycotoxins, a major risk factor for diabetic angiopathy. Proc Natl Acad Sci U S A. 2002;99(24):15596–15601. doi: 10.1073/pnas.242407999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Uribarri J, Peppa M, Cai W, et al. Dietary glycotoxins correlate with circulating advanced glycation end product levels in renal failure patients. Am J Kidney Dis. 2003;42(3):532–538. doi: 10.1016/s0272-6386(03)00779-0. [DOI] [PubMed] [Google Scholar]
- 15.Uribarri J, Peppa M, Cai W, et al. Restriction of dietary glycotoxins reduces excessive advanced glycation end products in renal failure patients. J Am Soc Nephrol. 2003;14(3):728–731. doi: 10.1097/01.asn.0000051593.41395.b9. [DOI] [PubMed] [Google Scholar]
- 16.Henle T. AGEs in foods: do they play a role in uremia? Kidney Int Suppl. 2003;(84):S145–S147. doi: 10.1046/j.1523-1755.63.s84.16.x. [DOI] [PubMed] [Google Scholar]
- 17.Erbersdobler HF, Faist V. Metabolic transit of Amadori products. Nahrung. 2001;45(3):177–181. doi: 10.1002/1521-3803(20010601)45:3<177::AID-FOOD177>3.0.CO;2-A. [DOI] [PubMed] [Google Scholar]
- 18.Gugliucci A, Bendayan M. Renal fate of circulating advanced glycated end products (AGE): evidence for reabsorption and catabolism of AGE-peptides by renal proximal tubular cells. Diabetologia. 1996;39(2):149–160. doi: 10.1007/BF00403957. [DOI] [PubMed] [Google Scholar]
- 19.Monnier VM, Sell DR, Nagaraj RH, et al. Maillard reaction-mediated molecular damage to extracellular matrix and other tissue proteins in diabetes, aging, and uremia. Diabetes. 1992;41(Suppl 2):36–41. doi: 10.2337/diab.41.2.s36. [DOI] [PubMed] [Google Scholar]
- 20.Niwa T, Katsuzaki T, Momoi T, et al. Modification of beta 2 m with advanced glycation end products as observed in dialysis-related amyloidosis by 3-DG accumulating in uremic serum. Kidney Int. 1996;49(3):861–867. doi: 10.1038/ki.1996.119. [DOI] [PubMed] [Google Scholar]
- 21.Bucala R, Makita Z, Vega G, et al. Modification of low density lipoprotein by advanced glycation end products contributes to the dyslipidemia of diabetes and renal insufficiency. Proc Natl Acad Sci U S A. 1994;91(20):9441–9445. doi: 10.1073/pnas.91.20.9441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Jensen LJ, Østergaard J, Flyvbjerg A. AGE-RAGE and AGE Cross-link interaction: important players in the pathogenesis of diabetic kidney disease. Horm Metab Res. 2005;37(Suppl 1):26–34. doi: 10.1055/s-2005-861360. [DOI] [PubMed] [Google Scholar]
- 23.Saito A, Takeda T, Sato K, et al. Significance of proximal tubular metabolism of advanced glycation end products in kidney diseases. Ann N Y Acad Sci. 2005;1043:637–643. doi: 10.1196/annals.1333.072. [DOI] [PubMed] [Google Scholar]
- 24.Vlassara H. Advanced glycation in health and disease: role of the modern environment. Ann N Y Acad Sci. 2005;1043:452–460. doi: 10.1196/annals.1333.051. [DOI] [PubMed] [Google Scholar]
- 25.Vlassara H, Striker LJ, Teichberg S, Fuh H, Li YM, Steffes M. Advanced glycation end products induce glomerular sclerosis and albuminuria in normal rats. Proc Natl Acad Sci U S A. 1994;91(24):11704–11708. doi: 10.1073/pnas.91.24.11704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Smedsrød B, Melkko J, Araki N, Sano H, Horiuchi S. Advanced glycation end products are eliminated by scavenger-receptor mediated endocytosis in hepatic sinusoidal Kupffer and endothelial cells. Biochem J. 1997;322(2):567–573. doi: 10.1042/bj3220567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Matsumoto K, Sano H, Nagai R, et al. Endocytic uptake of advanced glycation end products by mouse liver sinusoidal endothelial cells is mediated by a scavenger receptor distinct from the macrophage scavenger receptor class A. Biochem J. 2000;352(1):233–240. [PMC free article] [PubMed] [Google Scholar]
- 28.Hansen B, Svistounov D, Olsen R, Nagai R, Horiuchi S, Smedsrød B. Advanced glycation end products impair the scavenger function of rat hepatic sinusoidal endothelial cells. Diabetologia. 2002;45(10):1379–1388. doi: 10.1007/s00125-002-0912-8. [DOI] [PubMed] [Google Scholar]
- 29.Santos JC, Valentim IB, de Araújo OR, Ataide Tda R, Goulart MO. Development of nonalcoholic hepatopathy: contributions of oxidative stress and advanced glycation end products. Int J Mol Sci. 2013;14(10):19846–19866. doi: 10.3390/ijms141019846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Goh SY, Cooper ME. Clinical review: The role of advanced glycation end products in progression and complications of diabetes. J Clin Endocrinol Metab. 2008;93(4):1143–1152. doi: 10.1210/jc.2007-1817. [DOI] [PubMed] [Google Scholar]
- 31.Sandu O, Song K, Cai W, Zheng F, Uribarri J, Vlassara H. Insulin resistance and type 2 diabetes in high-fat-fed mice are linked to high glycotoxin intake. Diabetes. 2005;54(8):2314–2319. doi: 10.2337/diabetes.54.8.2314. [DOI] [PubMed] [Google Scholar]
- 32.Cai W, Ramdas M, Zhu L, Chen X, Striker GE, Vlassara H. Oral advanced glycation endproducts (AGEs) promote insulin resistance and diabetes by depleting the antioxidant defenses AGE receptor-1 and sirtuin 1. Proc Natl Acad Sci U S A. 2012;109(39):15888–15893. doi: 10.1073/pnas.1205847109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Uribarri J, Cai W, Ramdas M, et al. Restriction of advanced glycation end products improves insulin resistance in human type 2 diabetes: potential role of AGER1 and SIRT1. Diabetes Care. 2011;34(7):1610–1616. doi: 10.2337/dc11-0091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Negrean M, Stirban A, Stratmann B, et al. Effects of low- and high-advanced glycation endproduct meals on macro- and microvascular endothelial function and oxidative stress in patients with type 2 diabetes mellitus. Am J Clin Nutr. 2007;85(5):1236–1243. doi: 10.1093/ajcn/85.5.1236. [DOI] [PubMed] [Google Scholar]
- 35.Stirban A, Negrean M, Götting C, et al. Dietary advanced glycation endproducts and oxidative stress: in vivo effects on endothelial function and adipokines. Ann N Y Acad Sci. 2008;1126:276–279. doi: 10.1196/annals.1433.042. [DOI] [PubMed] [Google Scholar]
- 36.Miyata T, Kurokawa K, van Ypersele de Strihou C. Relevance of oxidative and carbonyl stress to long-term uremic complications. Kidney Int Suppl. 2000;76:S120–S125. doi: 10.1046/j.1523-1755.2000.07615.x. [DOI] [PubMed] [Google Scholar]
- 37.Raj DS, Choudhury D, Welbourne TC, Levi M. Advanced glycation end products: a Nephrologist’s perspective. Am J Kidney Dis. 2000;35(3):365–380. doi: 10.1016/s0272-6386(00)70189-2. [DOI] [PubMed] [Google Scholar]
- 38.Busch M, Franke S, Müller A, et al. Potential cardiovascular risk factors in chronic kidney disease: AGEs, total homocysteine and metabolites, and the C-reactive protein. Kidney Int. 2004;66(1):338–347. doi: 10.1111/j.1523-1755.2004.00736.x. [DOI] [PubMed] [Google Scholar]
- 39.Tan AL, Forbes JM, Cooper ME. AGE, RAGE, and ROS in diabetic nephropathy. Semin Nephrol. 2007;27(2):130–143. doi: 10.1016/j.semnephrol.2007.01.006. [DOI] [PubMed] [Google Scholar]
- 40.Chung AC, Zhang H, Kong YZ, et al. Advanced glycation end-products induce tubular CTGF via TGF-beta-independent Smad3 signaling. J Am Soc Nephrol. 2010;21(2):249–260. doi: 10.1681/ASN.2009010018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Coresh J, Astor BC, Greene T, Eknoyan G, Levey AS. Prevalence of chronic kidney disease and decreased kidney function in the adult US population: Third National Health and Nutrition Examination Survey. Am J Kidney Dis. 2003;41(1):1–12. doi: 10.1053/ajkd.2003.50007. [DOI] [PubMed] [Google Scholar]
- 42.Tanaka S, Avigad G, Brodsky B, Eikenberry EF. Glycation induces expansion of the molecular packing of collagen. J Mol Biol. 1988;203(2):495–505. doi: 10.1016/0022-2836(88)90015-0. [DOI] [PubMed] [Google Scholar]
- 43.Haitoglou CS, Tsilibary EC, Brownlee M, Charonis AS. Altered cellular interactions between endothelial cells and nonenzymatically glucosylated laminin/type IV collagen. J Biol Chem. 1992;267(18):12404–12407. [PubMed] [Google Scholar]
- 44.Gu L, Hagiwara S, Fan Q, et al. Role of receptor for advanced glycation end-products and signalling events in advanced glycation end-product-induced monocyte chemoattractant protein-1 expression in differentiated mouse podocytes. Nephrol Dial Transplant. 2006;21(2):299–313. doi: 10.1093/ndt/gfi210. [DOI] [PubMed] [Google Scholar]
- 45.Palimeri S, Palioura E, Piperi C, et al. Additive effects of dietary glycotoxins and androgen excess on the kidney of a female rat model. Alex J Med. Epub. [Google Scholar]
- 46.Sebeková K, Kupcová V, Schinzel R, Heidland A. Markedly elevated levels of plasma advanced glycation end products in patients with liver cirrhosis – amelioration by liver transplantation. J Hepatol. 2002;36(1):66–71. doi: 10.1016/s0168-8278(01)00232-x. [DOI] [PubMed] [Google Scholar]
- 47.Yagmur E, Tacke F, Weiss C, et al. Elevation of Nepsilon-(carboxymethyl) lysine-modified advanced glycation end products in chronic liver disease is an indicator of liver cirrhosis. Clin Biochem. 2006;39(1):39–45. doi: 10.1016/j.clinbiochem.2005.07.016. [DOI] [PubMed] [Google Scholar]
- 48.Hyogo H, Yamagishi S, Iwamoto K, et al. Elevated levels of serum advanced glycation end products in patients with non-alcoholic steatohepatitis. J Gastroenterol Hepatol. 2007;22(7):1112–1119. doi: 10.1111/j.1440-1746.2007.04943.x. [DOI] [PubMed] [Google Scholar]
- 49.Sato T, Wu X, Shimogaito N, Takino J, Yamagishi S, Takeuchi M. Effects of high-AGE beverage on RAGE and VEGF expressions in the liver and kidneys. Eur J Nutr. 2009;48(1):6–11. doi: 10.1007/s00394-008-0753-4. [DOI] [PubMed] [Google Scholar]
- 50.Cai W, He JC, Zhu L, et al. Reduced oxidant stress and extended lifespan in mice exposed to a low glycotoxin diet: association with increased AGER1 expression. Am J Pathol. 2007;170(6):1893–1902. doi: 10.2353/ajpath.2007.061281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Patel R, Baker SS, Liu W, et al. Effect of dietary advanced glycation end products on mouse liver. PLoS One. 2012;7(4):e35143. doi: 10.1371/journal.pone.0035143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Diamanti-Kandarakis E, Piperi C, Kalofoutis A, Creatsas G. Increased levels of serum advanced glycation end-products in women with polycystic ovary syndrome. Clin Endocrinol (Oxf) 2005;62(1):37–43. doi: 10.1111/j.1365-2265.2004.02170.x. [DOI] [PubMed] [Google Scholar]
- 53.Diamanti-Kandarakis E, Piouka A, Livadas S, et al. Anti-mullerian hormone is associated with advanced glycosylated end products in lean women with polycystic ovary syndrome. Eur J Endocrinol. 2009;160(5):847–853. doi: 10.1530/EJE-08-0510. [DOI] [PubMed] [Google Scholar]
- 54.Diamanti-Kandarakis E, Katsikis I, Piperi C, et al. Increased serum advanced glycation end-products is a distinct finding in lean women with polycystic ovary syndrome (PCOS) Clin Endocrinol (Oxf) 2008;69(4):634–641. doi: 10.1111/j.1365-2265.2008.03247.x. [DOI] [PubMed] [Google Scholar]
- 55.Diamanti-Kandarakis E, Piperi C, Patsouris E, et al. Immunohistochemical localization of advanced glycation end-products (AGEs) and their receptor (RAGE) in polycystic and normal ovaries. Histochem Cell Biol. 2007;127(6):581–589. doi: 10.1007/s00418-006-0265-3. [DOI] [PubMed] [Google Scholar]
- 56.Diamanti-Kandarakis E, Piperi C, Korkolopoulou P, et al. Accumulation of dietary glycotoxins in the reproductive system of normal female rats. J Mol Med (Berl) 2007;85(12):1413–1420. doi: 10.1007/s00109-007-0246-6. [DOI] [PubMed] [Google Scholar]
- 57.Kandaraki E, Chatzigeorgiou A, Piperi C, et al. Reduced ovarian glyoxalase-I activity by dietary glycotoxins and androgen excess: a causative link to polycystic ovarian syndrome. Mol Med. 2012;18:1183–1189. doi: 10.2119/molmed.2012.00293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Tantalaki E, Piperi C, Livadas S, et al. Impact of dietary modification of advanced glycation end products (AGEs) on the hormonal and metabolic profile of women with polycystic ovary syndrome (PCOS) Hormones (Athens) 2014;13(1):65–73. doi: 10.1007/BF03401321. [DOI] [PubMed] [Google Scholar]
- 59.Chatzigeorgiou A, Kandaraki E, Piperi C, et al. Dietary glycotoxins affect scavenger receptor expression and the hormonal profile of female rats. J Endocrinol. 2013;218(3):331–337. doi: 10.1530/JOE-13-0175. [DOI] [PubMed] [Google Scholar]
- 60.Diamanti-Kandarakis E, Chatzigeorgiou A, Papageorgiou E, Koundouras D, Koutsilieris M. Advanced glycation end-products and insulin signaling in granulosa cells. Exp Biol Med. 2015 May 7; doi: 10.1177/1535370215584937. Epub. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Boutzios G, Livadas S, Piperi C, et al. Polycystic ovary syndrome offspring display increased oxidative stress markers comparable to gestational diabetes offspring. Fertil Steril. 2013;99(3):943–950. doi: 10.1016/j.fertnstert.2012.10.050. [DOI] [PubMed] [Google Scholar]
- 62.Salahuddin P, Rabbani G, Khan RH. The role of advanced glycation end products in various types of neurodegenerative disease: a therapeutic approach. Cell Mol Biol Lett. 2014;19(3):407–437. doi: 10.2478/s11658-014-0205-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Cai W, Uribarri J, Zhu L, et al. Oral glycotoxins are a modifiable cause of dementia and the metabolic syndrome in mice and humans. Proc Natl Acad Sci U S A. 2014;111(13):4940–4945. doi: 10.1073/pnas.1316013111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Kandarakis SA, Piperi C, Topouzis F, Papavassiliou AG. Emerging role of advanced glycation-end products (AGEs) in the pathobiology of eye diseases. Prog Retin Eye Res. 2014;42:85–102. doi: 10.1016/j.preteyeres.2014.05.002. [DOI] [PubMed] [Google Scholar]
- 65.Kandarakis SA, Piperi C, Moschonas DP, Korkolopoulou P, Papalois A, Papavassiliou AG. Dietary glycotoxins induce RAGE and VEGF up-regulation in the retina of normal rats. Exp Eye Res. 2015;137:1–10. doi: 10.1016/j.exer.2015.05.017. [DOI] [PubMed] [Google Scholar]
- 66.Sharaf H, Matou-Nasri S, Wang Q, et al. Advanced glycation endproducts increase proliferation, migration and invasion of the breast cancer cell line MDA-MB-231. Biochim Biophys Acta. 2015;1852(3):429–441. doi: 10.1016/j.bbadis.2014.12.009. [DOI] [PubMed] [Google Scholar]
- 67.Rodriguez-Teja M, Gronau JH, Breit C, et al. AGE-modified basement membrane cooperates with Endo180 to promote epithelial cell invasiveness and decrease prostate cancer survival. J Pathol. 2015;235(4):581–592. doi: 10.1002/path.4485. [DOI] [PubMed] [Google Scholar]
- 68.Takino J, Yamagishi S, Takeuchi M. Cancer malignancy is enhanced by glyceraldehyde-derived advanced glycation end-products. J Oncol. 2010;2010:739852. doi: 10.1155/2010/739852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.van Heijst JW, Niessen HW, Hoekman K, Schalkwijk CG. Advanced glycation end products in human cancer tissues: detection of Nepsilon-(carboxymethyl)lysine and argpyrimidine. Ann N Y Acad Sci. 2005;1043:725–733. doi: 10.1196/annals.1333.084. [DOI] [PubMed] [Google Scholar]
- 70.Foster D, Spruill L, Walter KR, et al. AGE metabolites: a biomarker linked to cancer disparity? Cancer Epidemiol Biomarkers Prev. 2014;23(10):2186–2191. doi: 10.1158/1055-9965.EPI-14-0564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Jiao L, Weinstein SJ, Albanes D, et al. Evidence that serum levels of the soluble receptor for advanced glycation end products are inversely associated with pancreatic cancer risk: a prospective study. Cancer Res. 2011;71(10):3582–3589. doi: 10.1158/0008-5472.CAN-10-2573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Jiao L, Taylor PR, Weinstein SJ, et al. Advanced glycation end products, soluble receptor for advanced glycation end products, and risk of colorectal cancer. Cancer Epidemiol Biomarkers Prev. 2011;20(7):1430–1438. doi: 10.1158/1055-9965.EPI-11-0066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Grote VA, Nieters A, Kaaks R, et al. The associations of advanced glycation end products and its soluble receptor with pancreatic cancer risk: a case-control study within the prospective EPIC Cohort. Cancer Epidemiol Biomarkers Prev. 2012;21(4):619–628. doi: 10.1158/1055-9965.EPI-11-1139. [DOI] [PubMed] [Google Scholar]
- 74.Turner DP. Advanced glycation end-products: a biological consequence of lifestyle contributing to cancer disparity. Cancer Res. 2015;75(10):1925–1929. doi: 10.1158/0008-5472.CAN-15-0169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Hofmann SM, Dong HJ, Li Z, et al. Improved insulin sensitivity is associated with restricted intake of dietary glycoxidation products in the db/db mouse. Diabetes. 2002;51(7):2082–2089. doi: 10.2337/diabetes.51.7.2082. [DOI] [PubMed] [Google Scholar]
- 76.Zheng F, He C, Cai W, Hattori M, Steffes M, Vlassara H. Prevention of diabetic nephropathy in mice by a diet low in glycoxidation products. Diabetes Metab Res Rev. 2002;18(3):224–237. doi: 10.1002/dmrr.283. [DOI] [PubMed] [Google Scholar]
- 77.Vlassara H, Striker GE. AGE restriction in diabetes mellitus: a paradigm shift. Nat Rev Endocrinol. 2011;7(9):526–539. doi: 10.1038/nrendo.2011.74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Pouillart P, Mauprivez H, Ait-Ameur L, et al. Strategy for the study of the health impact of dietary Maillard products in clinical studies: the example of the ICARE clinical study on healthy adults. Ann N Y Acad Sci. 2008;1126:173–176. doi: 10.1196/annals.1433.040. [DOI] [PubMed] [Google Scholar]
- 79.Vlassara H, Cai W, Goodman S, et al. Protection against loss of innate defenses in adulthood by low advanced glycation end products (AGE) intake: role of the antiinflammatory AGE receptor-1. J Clin Endocrinol Metab. 2009;94(11):4483–4491. doi: 10.1210/jc.2009-0089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Birlouez-Aragon I, Saavedra G, Tessier FJ, et al. A diet based on high-heat-treated foods promotes risk factors for diabetes mellitus and cardiovascular diseases. Am J Clin Nutr. 2010;91(5):1220–1226. doi: 10.3945/ajcn.2009.28737. [DOI] [PubMed] [Google Scholar]
- 81.Kellow NJ, Savige GS. Dietary advanced glycation end-product restriction for the attenuation of insulin resistance, oxidative stress and endothelial dysfunction: a systematic review. Eur J Clin Nutr. 2013;67(3):239–248. doi: 10.1038/ejcn.2012.220. [DOI] [PubMed] [Google Scholar]
- 82.Brownlee M, Vlassara H, Kooney A, Ulrich P, Cerami A. Aminoguanidine prevents diabetes-induced arterial wall protein cross-linking. Science. 1986;232(4758):1629–1632. doi: 10.1126/science.3487117. [DOI] [PubMed] [Google Scholar]
- 83.Hammes HP, Brownlee M, Edelstein D, Saleck M, Martin S, Federlin K. Aminoguanidine inhibits the development of accelerated diabetic retinopathy in the spontaneous hypertensive rat. Diabetologia. 1994;37(1):32–35. doi: 10.1007/BF00428774. [DOI] [PubMed] [Google Scholar]
- 84.Soulis-Liparota T, Cooper M, Papazoglou D, Clarke B, Jerums G. Retardation by aminoguanidine of development of albuminuria, mesangial expansion, and tissue fluorescence in streptozocin-induced diabetic rat. Diabetes. 1991;40(10):1328–1334. doi: 10.2337/diab.40.10.1328. [DOI] [PubMed] [Google Scholar]
- 85.Nishizawa Y, Wada R, Baba M, Takeuchi M, Hanyu-Itabashi C, Yagihashi S. Neuropathy induced by exogenously administered advanced glycation end-products in rats. J Diabetes Investig. 2010;1(1–2):40–49. doi: 10.1111/j.2040-1124.2009.00002.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Forbes JM, Soulis T, Thallas V, et al. Renoprotective effects of a novel inhibitor of advanced glycation. Diabetologia. 2001;44(1):108–114. doi: 10.1007/s001250051587. [DOI] [PubMed] [Google Scholar]
- 87.Wilkinson-Berka JL, Kelly DJ, Koerner SM, et al. ALT-946 and aminoguanidine, inhibitors of advanced glycation, improve severe nephropathy in the diabetic transgenic (mREN-2)27 rat. Diabetes. 2002;51(11):3283–3289. doi: 10.2337/diabetes.51.11.3283. [DOI] [PubMed] [Google Scholar]
- 88.Cheng G, Wang LL, Long L, et al. Beneficial effects of C36, a novel breaker of advanced glycation endproducts cross-links, on the cardiovascular system of diabetic rats. Br J Pharmacol. 2007;152(8):1196–1206. doi: 10.1038/sj.bjp.0707533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Okamoto T, Yamagishi S, Inagaki Y, et al. Angiogenesis induced by advanced glycation end products and its prevention by cerivastatin. FASEB J. 2002;16(14):1928–1930. doi: 10.1096/fj.02-0030fje. [DOI] [PubMed] [Google Scholar]
- 90.Stitt A, Gardiner TA, Alderson NL, et al. The AGE inhibitor pyridoxamine inhibits development of retinopathy in experimental diabetes. Diabetes. 2002;51(9):2826–2832. doi: 10.2337/diabetes.51.9.2826. [DOI] [PubMed] [Google Scholar]
- 91.Degenhardt TP, Alderson NL, Arrington DD, et al. Pyridoxamine inhibits early renal disease and dyslipidemia in the streptozotocin-diabetic rat. Kidney Int. 2002;61(3):939–950. doi: 10.1046/j.1523-1755.2002.00207.x. [DOI] [PubMed] [Google Scholar]
- 92.Vasan S, Zhang X, Zhang X, et al. An agent cleaving glucose-derived protein crosslinks in vitro and in vivo. Nature. 1996;382(6588):275–278. doi: 10.1038/382275a0. [DOI] [PubMed] [Google Scholar]
- 93.Candido R, Forbes JM, Thomas MC, et al. A breaker of advanced glycation end products attenuates diabetes-induced myocardial structural changes. Circ Res. 2003;92(7):785–792. doi: 10.1161/01.RES.0000065620.39919.20. [DOI] [PubMed] [Google Scholar]
- 94.Coughlan MT, Forbes JM, Cooper ME. Role of the AGE crosslink breaker, alagebrium, as a renoprotective agent in diabetes. Kidney Int Suppl. 2007;(106):S54–S60. doi: 10.1038/sj.ki.5002387. [DOI] [PubMed] [Google Scholar]
- 95.Kass DA, Shapiro EP, Kawaguchi M, et al. Improved arterial compliance by a novel advanced glycation end-product crosslink breaker. Circulation. 2001;104(13):1464–1470. doi: 10.1161/hc3801.097806. [DOI] [PubMed] [Google Scholar]
- 96.Little WC, Zile MR, Kitzman DW, Hundley WG, O’Brien TX, Degroof RC. The effect of alagebrium chloride (ALT-711), a novel glucose cross-link breaker, in the treatment of elderly patients with diastolic heart failure. J Card Fail. 2005;11(3):191–195. doi: 10.1016/j.cardfail.2004.09.010. [DOI] [PubMed] [Google Scholar]
- 97.Aldini G, Vistoli G, Stefek M, et al. Molecular strategies to prevent, inhibit, and degrade advanced glycoxidation and advanced lipoxidation end products. Free Radic Res. 2013;47(Suppl 1):93–137. doi: 10.3109/10715762.2013.792926. [DOI] [PubMed] [Google Scholar]
- 98.Win MT, Yamamoto Y, Munesue S, et al. Regulation of RAGE for attenuating progression of diabetic vascular complications. Exp Diabetes Res. 2012;2012:894605. doi: 10.1155/2012/894605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Yamagishi S, Nakamura K, Matsui T, Ueda S, Fukami K, Okuda S. Agents that block advanced glycation end product (AGE)-RAGE (receptor for AGEs)-oxidative stress system: a novel therapeutic strategy for diabetic vascular complications. Expert Opin Investig Drugs. 2008;17(7):983–996. doi: 10.1517/13543784.17.7.983. [DOI] [PubMed] [Google Scholar]
- 100.Park L, Raman KG, Lee KJ, et al. Suppression of accelerated diabetic atherosclerosis by the soluble receptor for advanced glycation endproducts. Nat Med. 1998;4(9):1025–1031. doi: 10.1038/2012. [DOI] [PubMed] [Google Scholar]
- 101.Bucciarelli LG, Wendt T, Qu W, et al. RAGE blockade stabilizes established atherosclerosis in diabetic apolipoprotein E-null mice. Circulation. 2002;106(22):2827–2835. doi: 10.1161/01.cir.0000039325.03698.36. [DOI] [PubMed] [Google Scholar]
- 102.Barile GR, Pachydaki SI, Tari SR, et al. The RAGE axis in early diabetic retinopathy. Invest Ophthalmol Vis Sci. 2005;46(8):2916–2924. doi: 10.1167/iovs.04-1409. [DOI] [PubMed] [Google Scholar]
- 103.Kaji Y, Usui T, Ishida S, et al. Inhibition of diabetic leukostasis and blood-retinal barrier breakdown with a soluble form of a receptor for advanced glycation end products. Invest Ophthalmol Vis Sci. 2007;48(2):858–865. doi: 10.1167/iovs.06-0495. [DOI] [PubMed] [Google Scholar]
- 104.Li F, Drel VR, Szabó C, Stevens MJ, Obrosova IG. Low-dose poly(ADP-ribose) polymerase inhibitor-containing combination therapies reverse early peripheral diabetic neuropathy. Diabetes. 2005;54(5):1514–1522. doi: 10.2337/diabetes.54.5.1514. [DOI] [PubMed] [Google Scholar]
- 105.Pacher P, Liaudet L, Soriano FG, Mabley JG, Szabó E, Szabó C. The role of poly(ADP-ribose) polymerase activation in the development of myocardial and endothelial dysfunction in diabetes. Diabetes. 2002;51(2):514–521. doi: 10.2337/diabetes.51.2.514. [DOI] [PubMed] [Google Scholar]
- 106.Ueda S, Yamagishi S, Takeuchi M, et al. Oral adsorbent AST-120 decreases serum levels of AGEs in patients with chronic renal failure. Mol Med. 2006;12(7–8):180–184. doi: 10.2119/2005-00034.Ueda. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Diamanti-Kandarakis E, Katsikis I, Piperi C, Alexandraki K, Panidis D. Effect of long-term orlistat treatment on serum levels of advanced glycation end-products in women with polycystic ovary syndrome. Clin Endocrinol (Oxf) 2007;66(1):103–109. doi: 10.1111/j.1365-2265.2006.02693.x. [DOI] [PubMed] [Google Scholar]
- 108.Piperi C, Adamopoulos C, Dalagiorgou G, Diamanti-Kandarakis E, Papavassiliou AG. Crosstalk between advanced glycation and endoplasmic reticulum stress: emerging therapeutic targeting for metabolic diseases. J Clin Endocrinol Metab. 2012;97(7):2231–2242. doi: 10.1210/jc.2011-3408. [DOI] [PubMed] [Google Scholar]