

Abstract
Objective:
Exposure to maternal obesity in utero predisposes offspring to obesity and metabolic disease. This study investigated whether maternal obesity is associated with alterations in expression of fetal microRNA (miRNA).
Study Design:
A cohort study of women with body mass index (BMI) ≥35 kg/m2 (n = 16) versus those with normal BMI 20 to 24.9 (n = 20) was performed. All participants had normal glucose tolerance (1-hour glucose challenge test <130) and normally grown neonates (2700-3500 g). Umbilical cord samples were collected immediately after delivery. Expression of miRNA was assessed using Affymetrix GeneChip miRNA 3.0 Arrays. Differential miRNA expression was determined using Student t tests with Benjamini-Hocherg correction.
Results:
For 1733 human mature miRNAs, the expression levels were not statistically different in umbilical cord blood samples from pregnancies of obese women compared to controls.
Conclusion:
Expression of fetal miRNA is not altered in umbilical cord blood in response to in utero exposure to obesity. Alternate mechanisms underlying the fetal effects of maternal obesity should be explored.
Keywords: fetal metabolic programming, microRNA, obesity
Introduction
In the United States, more than one-half of pregnant women are overweight or obese, putting them at a greater risk of pregnancy complications.1 In addition to the known maternal risks, infants born to obese women are at increased risk of prematurity, stillbirth, congenital anomalies, macrosomia with possible birth injury, and childhood obesity.2,3 In utero exposure to maternal obesity may lead to long-term consequences, predisposing children to metabolic disease later in life.
Nutritional stress during critical periods of early development permanently alters the physiology and metabolism of an organism, affecting health in later life via a mechanism known as fetal metabolic programming. Fetal overnutrition also predisposes offspring to obesity, type 2 diabetes, hypertension, and cardiovascular disease. Studies of human mother–offspring cohorts have demonstrated an association between high maternal body mass index (BMI) or excessive gestational weight gain and disrupted childhood metabolism and cardiovascular function.4,5
The mechanisms underlying the fetal effects from maternal overnutrition are poorly understood, yet several pathways have been proposed.6,7 These mechanisms include altered organ development, cellular signaling responses, and epigenetic modifications.8,9 Emerging data have revealed that microRNA (miRNA) controls key cellular processes such as development, growth, apoptosis and inflammation. MicroRNAs are now implicated in varied disease states including cancer,10 cardiovascular disease,11 obesity,12,13 and type 2 diabetes mellitus,14 but few studies have investigated the role of miRNA in pregnancy-related pathology.15
Recent pregnancy-related studies have demonstrated a potential role of miRNAs in the pathogenesis of preeclampsia,16,17 preterm labor,18 and gestational diabetes mellitus.19 Although studies have proven the significance of miRNAs in the pathogenesis of disease and the value of miRNAs as reliable biomarkers of disease, there remains a paucity of information regarding the role of miRNAs in fetal overnutrition.
We hypothesized that epigenetic alterations would be present in fetuses exposed to an obesogenic in utero environment. Traditionally, umbilical cord blood has been studied as a reflection of the fetal state. In the present study, we explored the expression profiles of miRNA in umbilical cord blood obtained from obese and nonobese women to determine whether an obesogenic in utero environment is associated with specific alterations in fetal miRNA profiles.
Materials and Methods
Study Design
A cohort study was conducted to examine expression of miRNA in umbilical cord blood from obese women (n = 16) and normal weight women (n = 20). This sample size was selected based on studies of gene expression microarray showing that 10 to 15 replicates yield results that are stable, with little increase in stability with increasing sample size.20 Body mass index was calculated at first prenatal visit (earliest visit ≤22 weeks). Women were categorized based on the World Health Organization classifications, with normal defined as a BMI of 20 to 24.9 kg/m2 and obese as a BMI ≥35 kg/m2. Women with obesity class II and above were chosen to ensure that there was a clear phenotypic difference between the normal weight and obese groups, increasing the possibility of finding a difference in expression of maternal weight-related miRNA. Women who met the following criteria were included in the study: age 18 to 45 years, singleton pregnancy, normal glucose tolerance (1 hour glucose challenge test [GCT] ≤130 mg/dL), term delivery (37-41 6/7 weeks), and normally grown neonates (2700-3500 g). Normal birth weight between the 10th and 90th percentiles at 37 to 41 weeks of gestation were identified using the Alexander curve.21 To ensure that the effect of obesity alone was examined independent of glucose intolerance, a threshold of 130 mg/dL was used, which is the lowest value that has been proposed as a screening cutoff for gestational diabetes mellitus.22
Women with comorbid conditions that could independently affect fetal growth were excluded, including women with chronic hypertension, preeclampsia, chronic steroid use, diabetes, major fetal anomaly, connective tissue disorder requiring medication, and active HIV or hepatitis C. The study was approved by the institutional review board of the University of Pennsylvania. All study participants provided written informed consent.
RNA Isolation
Umbilical cord blood samples were collected immediately after delivery, serum was isolated, and samples stored at −80°C until further use. Cell-free total RNA was isolated from the cord blood serum, using Qiagen miRNeasy Serum Kits according to the manufacturer’s instructions (Qiagen, Valencia, California). Briefly, umbilical cord blood cells were lysed, RNA was extracted by phenol/chloroform extraction, followed by silica membrane-based purification of total RNA. RNA concentration was determined with the NanoDrop spectrophotometer (NanoDrop 2000 Spectrophotometer; Nanodrop, Rockland, Delaware).
Microarray Methods and Analyses
Expression of miRNA was determined using Affymetrix GeneChip miRNA 3.0 Arrays (Affymetrix, Santa Clara, California) according to the manufacturer’s instructions. Briefly, the FlashTag Biotin HSR RNA (Affymetrix, Santa Clara, California) labeling procedure was used to label total RNA. This involves a brief tailing reaction followed by ligation of the biotinylated signal molecule to the target RNA sample. The labeled samples were hybridized to the Affymetrix GeneChip miRNA 3.0 array and scanned according to the manufacturer’s instructions. Hybridization images were then scanned and digitized with the Genechip Scanner 3000 (Affymetrix). The normalized signal intensity was log2 transformed, and data analysis was performed with the Partek 6.6 Genomic Suite software (Copywrite; Partek Inc, St. Louis, Missouri). Differential miRNA expression was determined using Student’s t test with Benjamini-Hocherg (BH) correction. The Affymetrix GeneChip miRNA 3.0 array contains 19 724 total mature miRNA probe sets, representing 153 organisms. We focused our analysis to the 1733 human mature miRNAs on the arrays, given that these were most relevant to our study of human fetal umbilical cord blood. MicroRNAs associated with absolute Z-scores ≥2 or BH < .05 were considered significant. Global sample variation was assessed by principal components analysis.
Statistical Analysis
For demographic data and clinical characteristics, statistical analyses were performed using Stata version 10.1 (StataCorp LP, College Station, Texas). Categorical data were compared using chi-square tests. Means and medians of continuous data were compared using Student t tests and Wilcoxon rank-sum tests, as indicated.
Results
Maternal characteristics including age, GCT value, delivery mode, and smoking status were similar between the groups (Table 1). The mean BMI at the first prenatal visit prior to 22 weeks, “First BMI,” was 22.8 kg/m2 for the normal weight group, while mean BMI at first prenatal visit was 41.9 kg/m2 for the obese group. The median gestational age at first prenatal visit was similar between cases (9.4 weeks) and controls (9.7 weeks). Normal weight women gained significantly more weight than their obese counterparts. However, the mean weight gain of both groups was within the Institute of Medicine (IOM) recommendations, based on their initial BMI.23 Neonatal characteristics, including infant birth weight, gestational age at delivery, and gender, were also similar between the groups (Table 1).
Table 1.
Demographics and Clinical Characteristics.a,b
| Variable | Normal BMI (n = 20) | Obese (n = 16) | P |
|---|---|---|---|
| Race | .04 | ||
| Black | 8 (40) | 14 (87.5) | |
| White | 7 (35) | 1 (6.3) | |
| Asian | 3 (15) | 0 | |
| Latina | 1 (5) | 0 | |
| Other | 1 (5) | 1 (6.3) | |
| Age, years | 26.5 ± 5.8 | 25.6 ± 3.9 | .61 |
| Tobacco use, n (%) | 3 (15) | 3 (18.8) | .76 |
| Prepregnancy BMI, kg/m2 | 22.7 (20.6-23.9) | 37.1 (34.1-46.9) | <.001 |
| First BMI, kg/m2 | 22.8 ± 1.6 | 41.9 ± 6.4 | <.001 |
| Gestational age at first visit, weeks | 9.7 (8.2-11.3) | 9.4 (7.9-13.4) | .91 |
| Last BMI, kg/m2 | 28.8 ± 3.1 | 45.2 ± 6.8 | <.001 |
| Weight gain, kg | 15.5 ± 5.7 | 8.7 ± 6.3 | .002 |
| Infant birth weight, g | 3204.7 ± 185.6 | 3178.5 ± 159.1 | .66 |
| 1 hour GCT, mg/dL | 90.9 ± 19.4 | 99.3 ± 16.2 | .18 |
| Gestational age at delivery, weeks | 40 ± 1.2 | 39.3 ± 0.9 | .04 |
| Female infant, n (%) | 12 (60) | 11 (68.75) | .59 |
| Cesarean delivery, n (%) | 6 (30) | 4 (25) | .74 |
Abbreviations: BMI, body mass index; GCT, glucose challenge test.
a P values were determined by chi-square test (categorical data) and appropriate comparisons of means and medians using t test (for parametric continuous data) and Wilcoxon rank-sum test (for nonparametric continuous data).
bData are n (%), median (IQR), or mean ± standard deviation.
Among the 1733 human mature miRNAs screened by the miRNA microarray, there were no differences in expression between the 2 groups. Principal components analysis demonstrates that the global miRNA expression profiles were overall similar among all samples (Figure 1). Samples were regrouped by maternal weight gain, first pregnancy BMI, last pregnancy BMI, delivery mode, fetal gender, and race, and there remained no significant differences in expression of miRNA between the groups.
Figure 1.
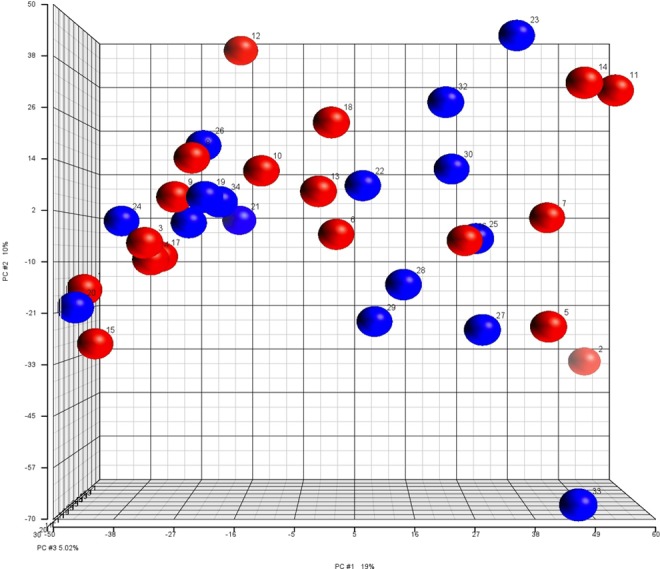
Principal component analysis plot for expression of microRNA (miRNA) in umbilical cord blood. Red spheres represent controls with normal maternal BMI (BMI 20-24.9 at first prenatal visit). Blue spheres represent cases with maternal obesity BMI (BMI ≥ 35 kg/m2 at first prenatal visit). BMI indicates body mass index. (The color version of this figure is available in the online version at http://rs.sagepub.com/.)
Comment
This study was performed to discover differences in the expression of miRNA in the umbilical cord of fetuses born to obese women compared to those of women with normal weight, in order to elucidate molecular changes in the fetus related to maternal obesity in the absence of maternal diabetes or other significant comorbidities. We found that expression of umbilical cord miRNA was not statistically different in pregnancies of obese women compared with those of normal weight women.
A few studies have investigated the role of miRNA in fetal states of malnutrition in humans. Differential miRNA expression has been described in the placenta from pregnancies complicated by fetal growth restriction.24 Published human studies have not investigated an association with a maternal obesogenic environment on expression of miRNA, but several animal studies have explored this association. Animal studies have investigated organ-specific changes in the miRNA profile of offspring after exposure to maternal obesity, including miRNA isolated from baboon hearts,25 sheep livers,26 and sheep muscle miRNA.27
In contrast to these studies, which have demonstrated miRNA changes in relation to a maternal obesogenic environment, our study did not find such differences in umbilical cord blood samples. Our negative findings may be accounted for by the isolation of miRNA from umbilical cord blood rather than directly from the fetal organs which may be involved in metabolic programming, such as the liver or pancreas. Additionally, the state of obesity in animal models is inherently different from obesity in the human pregnancy as it is largely defined by a controlled diet.
It is also plausible that changes in the miRNA profile occur earlier in pregnancy and are not evident at term. It is well known that there is likely a temporal nature to fetal programming, since the developing organism passes through multiple critical periods of development at different stages, and the vulnerability of organ systems to certain insults differs according to its stage of development at the time.
Finally, our findings might merely indicate that maternal obesity alone does not significantly alter miRNA profile in the fetus and hence, this epigenetic modification is not a key mechanism in adverse outcomes in offspring born from obese mothers. Recent studies suggest an alternative pathway of altered DNA methylation in umbilical cord blood associated with maternal obesity.28,29 Alterations in methylation of histones and chromatin remodeling are also being investigated as epigenetic contributors to fetal metabolic programming.30
Our study has several notable strengths. We carefully selected an obese population without key confounders, such as glucose intolerance, hypertensive disorders, and fetal growth disturbance. This study design allowed us to study the effects of obesity alone. Although we consider this is a strength of our study, we recognize that excluding other aspects of the metabolic syndrome may have selected an obese population without sufficient metabolic changes to alter expression of miRNA. Although some of these comorbidities may have a greater contribution to fetal programming than obesity in isolation, we thought it was prudent to first determine a possible effect of maternal obesity on expression of miRNA. The effect of comorbidities on expression of miRNA and neonatal outcomes would require further research. For such a discovery study, our sample size was sufficient based on studies of gene expression microarray showing that 10 to 15 replicates yield results that are quite stable, and there is less improvement in stability as the number of replicates is further increased.20 An increase in sample numbers would unlikely alter our findings.
There are noted limitations to this study. We performed a discovery study and not an investigation on focused molecular pathways. However, since so little is known regarding miRNA and fetal programming in the setting of maternal obesity, a discovery study seemed more appropriate. Although we hypothesized that umbilical cord blood would best represent the fetal response to maternal obesity, miRNA profiling in specific tissues might be more revealing as to an effect from maternal obesity. This type of investigation is not possible in human pregnancy. Traditionally, umbilical cord blood has been used in research to reflect the fetal state and is a feasible human biospecimen.
In terms of defining obesity, we used recorded BMI at first prenatal visit to select cases of obesity because prepregnancy weight was self-reported, which has lead to misclassification bias in prior studies.31 A majority of patients in our study presented for prenatal care in the first trimester, so the initial recorded BMI likely accurately reflects their classification prior to weight gain from pregnancy. However, pre-pregnancy BMI may be a better indicator of an obesogenic environment. Additionally, maternal diet or weight gain may have a greater effect on fetal programming than obesity alone. To explore this, we did regroup the samples by maternal pregnancy weight gain but did not find differences in the expression of miRNA between the groups. However, the mean pregnancy weight gain in both the groups was within the IOM recommendations. To assess maternal diet and miRNA profiling, a more intense study would be required that was beyond the scope of this work.
Our study suggests that the fetal changes in obesogenic pregnancies are unlikely to be caused by alterations in expression of fetal miRNA at term. Future research should investigate miRNA changes in other tissues, such as the placenta. Additionally, human cohorts with differences in maternal weight gain, maternal diet, or fetal growth disturbance should be investigated. Further investigation is needed to understand the mechanisms underlying the fetal molecular pathways in states of overnutrition, as these processes are known to have far-reaching effects on the health of future generations.
Footnotes
Declaration of Conflicting Interests: The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding: The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The Maternal and Child Health Research Program within the Department of Obstetrics and Gynecology at the University of Pennsylvania Health System provided financial support for this research.
References
- 1. American College of O, Gynecologists. ACOG Committee opinion no. 549: obesity in pregnancy. Obstet Gynecol. 2013;121 (1):213–217. [DOI] [PubMed] [Google Scholar]
- 2. Stothard KJ, Tennant PW, Bell R, Rankin J. Maternal overweight and obesity and the risk of congenital anomalies: a systematic review and meta-analysis. JAMA. 2009;301 (6):636–650. [DOI] [PubMed] [Google Scholar]
- 3. Oken E, Taveras EM, Kleinman KP, Rich-Edwards JW, Gillman MW. Gestational weight gain and child adiposity at age 3 years. Am J Obstet Gynecol. 2007;196 (4):322. e1–e8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Mamun AA, Hayatbakhsh MR, O’Callaghan M, Williams G, Najman J. Early overweight and pubertal maturation—pathways of association with young adults' overweight: a longitudinal study. Int J Obes (lond). 2009;33 (1):14–20. [DOI] [PubMed] [Google Scholar]
- 5. Wen X, Triche EW, Hogan JW, Shenassa ED, Buka SL. Prenatal factors for childhood blood pressure mediated by intrauterine and/or childhood growth? Pediatrics. 2011;127 (3):e713–e721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Brenseke B, Prater MR, Bahamonde J, Gutierrez JC. Current thoughts on maternal nutrition and fetal programming of the metabolic syndrome. J Pregnancy. 2013;2013:368461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Lau C, Rogers JM, Desai M, Ross MG. Fetal programming of adult disease: implications for prenatal care. Obstet Gynecol. 2011;117 (4):978–985. [DOI] [PubMed] [Google Scholar]
- 8. Heijmans BT, Tobi EW, Stein AD, et al. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc Natl Acad Sci USA. 2008;105 (44):17046–17049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Godfrey KM, Sheppard A, Gluckman PD, et al. Epigenetic gene promoter methylation at birth is associated with child’s later adiposity. Diabetes. 2011;60 (5):1528–1534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Choi JD, Lee JS. Interplay between epigenetics and genetics in cancer. Genomics Inform. 2013;11 (4):164–173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Fiedler J, Gupta SK, Thum T. MicroRNA-based therapeutic approaches in the cardiovascular system. Cardiovasc Ther. 2012;30 (1):e9–e15. [DOI] [PubMed] [Google Scholar]
- 12. Esau C, Kang X, Peralta E, et al. MicroRNA-143 regulates adipocyte differentiation. J Biol Chem. 2004;279 (50):52361–52365. [DOI] [PubMed] [Google Scholar]
- 13. Kloting N, Berthold S, Kovacs P, et al. MicroRNA expression in human omental and subcutaneous adipose tissue. PLoS One. 2009;4 (3):e4699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Zampetaki A, Kiechl S, Drozdov I, et al. Plasma microRNA profiling reveals loss of endothelial miR-126 and other microRNAs in type 2 diabetes. Cir Res. 2010;107 (6):810–817. [DOI] [PubMed] [Google Scholar]
- 15. Zhao Z, Moley KH, Gronowski AM. Diagnostic potential for miRNAs as biomarkers for pregnancy-specific diseases. Clin Biochem. 2013;46 (10-11):953–960. [DOI] [PubMed] [Google Scholar]
- 16. Pineles BL, Romero R, Montenegro D, et al. Distinct subsets of microRNAs are expressed differentially in the human placentas of patients with preeclampsia. Am J Obstet Gynecol. 2007;196 (3):261. e1–e6. [DOI] [PubMed] [Google Scholar]
- 17. Zhang Y, Diao Z, Su L, et al. MicroRNA-155 contributes to preeclampsia by down-regulating CYR61. Am J Obstet Gynecol. 2010;202 (5):466. e1–e7. [DOI] [PubMed] [Google Scholar]
- 18. Montenegro D, Romero R, Kim SS, et al. Expression patterns of microRNAs in the chorioamniotic membranes: a role for microRNAs in human pregnancy and parturition. J Pathol. 2009;217 (1):113–121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Zhao C, Dong J, Jiang T, et al. Early second-trimester serum miRNA profiling predicts gestational diabetes mellitus. PLoS One. 2011;6 (8):e23925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Pavlidis P, Li Q, Noble WS. The effect of replication on gene expression microarray experiments. Bioinformatics. 2003;19 (13):1620–1627. [DOI] [PubMed] [Google Scholar]
- 21. Alexander GR, Himes JH, Kaufman RB, Mor J, Kogan M. A United States national reference for fetal growth. Obstet Gynecol. 1996;87 (2):163–168. [DOI] [PubMed] [Google Scholar]
- 22. Donovan L, Hartling L, Muise M, Guthrie A, Vandermeer B, Dryden DM. Screening tests for gestational diabetes: a systematic review for the U.S. Preventive Services Task Force. Ann Intern Med. 2013;159 (2):115–122. [DOI] [PubMed] [Google Scholar]
- 23. Rasmussen KM, Yaktine AL, eds. Weight Gain During Pregnancy: Reexamining the Guidelines. Washington, DC: National Academies Press; 2009. [PubMed] [Google Scholar]
- 24. Maccani MA, Padbury JF, Marsit CJ. miR-16 and miR-21 expression in the placenta is associated with fetal growth. PLoS One. 2011;6 (6):e21210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Maloyan A, Muralimanoharan S, Huffman S, et al. Identification and comparative analyses of myocardial miRNAs involved in the fetal response to maternal obesity. Physiol Genomics. 2013;45 (19):889–900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Nicholas LM, Rattanatray L, MacLaughlin SM, et al. Differential effects of maternal obesity and weight loss in the periconceptional period on the epigenetic regulation of hepatic insulin-signaling pathways in the offspring. FASEB J. 2013;27 (9):3786–3796. [DOI] [PubMed] [Google Scholar]
- 27. Yan X, Huang Y, Zhao JX, et al. Maternal obesity downregulates microRNA let-7g expression, a possible mechanism for enhanced adipogenesis during ovine fetal skeletal muscle development. Int J Obes (lond). 2013;37 (4):568–575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Gemma C, Sookoian S, Alvarinas J, et al. Maternal pregestational BMI is associated with methylation of the PPARGC1A promoter in newborns. Obesity. 2009;17 (5):1032–1039. [DOI] [PubMed] [Google Scholar]
- 29. Perkins E, Murphy SK, Murtha AP, et al. Insulin-like growth factor 2/H19 methylation at birth and risk of overweight and obesity in children. J Pediatr. 2012;161 (1):31–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Sookoian S, Gianotti TF, Burgueno AL, Pirola CJ. Fetal metabolic programming and epigenetic modifications: a systems biology approach. Pediatr Res. 2013;73 (4 pt 2):531–542. [DOI] [PubMed] [Google Scholar]
- 31. Bodnar LM, Siega-Riz AM, Simhan HN, Diesel JC, Abrams B. The impact of exposure misclassification on associations between prepregnancy BMI and adverse pregnancy outcomes. Obesity. 2010;18 (11):2184–2190. [DOI] [PMC free article] [PubMed] [Google Scholar]