

Significance
IL-2 and IL-15, members of the gamma chain family of cytokines, are prominently deregulated in human T-cell lymphotropic virus type 1 (HTLV-1)-associated myelopathy/tropical spastic paraparesis (HAM/TSP) and underlie many of the characteristic immune abnormalities such as spontaneous lymphocyte proliferation, increased STAT5 phosphorylation in the lymphocytes, and increased frequency and cytotoxicity of virus-specific CD8+ T lymphocytes (CTLs). In this in vitro study, we demonstrate that selective and simultaneous blockade of IL-2 and IL-15, with a γ-chain antagonistic peptide, reduces spontaneous lymphocyte proliferation (SP), STAT5 phosphorylation, and more important, the degranulation of CD8+ T cells and the frequency of HTLV-1-specific CTLs. Thus, selective cytokine blockade with antagonistic peptides might be a therapeutic approach in HAM/TSP and is potentially applicable to multiple other conditions in which cytokines are pathogenic.
Keywords: human T-cell lymphotropic virus type 1, HAM/TSP, gamma chain, IL-15
Abstract
Human T-cell lymphotropic virus type 1 (HTLV-1)-associated myelopathy/tropical spastic paraparesis (HAM/TSP) is a progressive inflammatory myelopathy occurring in a subset of HTLV-1-infected individuals. Despite advances in understanding its immunopathogenesis, an effective treatment remains to be found. IL-2 and IL-15, members of the gamma chain (γc) family of cytokines, are prominently deregulated in HAM/TSP and underlie many of the characteristic immune abnormalities, such as spontaneous lymphocyte proliferation (SP), increased STAT5 phosphorylation in the lymphocytes, and increased frequency and cytotoxicity of virus-specific cytotoxic CD8+ T lymphocytes (CTLs). In this study, we describe a novel immunomodulatory strategy consisting of selective blockade of certain γc family cytokines, including IL-2 and IL-15, with a γc antagonistic peptide. In vitro, a PEGylated form of the peptide, named BNZ132-1-40, reduced multiple immune activation markers such as SP, STAT5 phosphorylation, spontaneous degranulation of CD8+ T cells, and the frequency of transactivator protein (Tax)-specific CD8+ CTLs, thought to be major players in the immunopathogenesis of the disease. This strategy is thus a promising therapeutic approach to HAM/TSP with the potential of being more effective than single monoclonal antibodies targeting either IL-2 or IL-15 receptors and safer than inhibitors of downstream signaling molecules such as JAK1 inhibitors. Finally, selective cytokine blockade with antagonistic peptides might be applicable to multiple other conditions in which cytokines are pathogenic.
Human T-cell lymphotropic virus type 1 (HTLV-1)-associated myelopathy/tropical spastic paraparesis (HAM/TSP) is a central nervous system (CNS) disease occurring in up to 3% of HTLV-1-infected individuals (1). Clinically, it is a spontaneous lymphocyte proliferation (SP) with sphincter dysfunction, pointing to predominant involvement of the lower thoracic spinal cord. However, many other syndromes [such as cognitive and autonomic dysfunction, amyotrophic lateral sclerosis (ALS)-like syndrome, peripheral neuropathy, and myositis] have been reported, suggesting more widespread neurologic damage (2). Typically, the onset is insidious, and significant disability occurs within years of symptom manifestation. However, a rapidly progressive form with acute onset and complete paraplegia in less than 2 y can also occur (3).
There is substantial evidence that HAM/TSP is an immune-mediated disorder, the pathogenesis of which is associated with deregulated immune activation. Pathologically, there is perivascular and parenchymal inflammatory infiltration in the watershed areas of the CNS with associated axonal loss, demyelination, reactive astrocytosis, and fibrillary gliosis (4). Early in the course of the disease, the inflammatory infiltrate contains equal numbers of CD4+, CD8+ T cells, and foamy macrophages; over time, the lesions become less inflammatory with a predominance of CD8+ T cells and high levels of IFN-γ (2, 5). Immunologically, ex vivo spontaneous proliferation of the peripheral blood mononuclear cells (PBMCs) is a hallmark of ongoing lymphocyte activation in HAM/TSP. Both CD4+ and CD8+ T cells are involved in SP, but the CD8+ T cells, and in particular, HTLV-1-specific CD8+ cytotoxic T lymphocytes (CTLs), proliferate more vigorously (6). These virus-specific CTLs infiltrate CNS lesions, where they could function to eliminate HTLV-1-infected CD4+ T cells, resulting in the secretion of damaging proinflammatory factors (such as TNF-α and IFN-γ) and bystander CNS damage (7). Notably, transactivator protein (Tax)-specific CTLs are found at higher frequency in the peripheral blood and cerebrospinal fluid of patients with HAM/TSP compared with in asymptomatic carriers (8, 9); these cells are predominantly of the memory effector phenotype and have increased cytotoxicity (10).The phenomenon of SP has been suggested to be a surrogate marker for the expansion and persistence of such virus-specific pathogenic CTLs in the CNS (6).
IL-2 and IL-15 are pleiotropic cytokines that play pivotal roles in inflammation and immune responses to infections. Both are glycoprotein members of the four α-helix bundle-containing cytokines and signal through receptor complexes that use the gamma chain (γc; CD132) and the β chain [CD122; IL-2/IL-15 receptor (R)]. γc is additionally shared with IL-4, IL-7, IL-9, and IL-21, all of which are members of the common γc family of cytokines (11). Cytokine binding to the β/γc complex results in heterodimerization of their cytoplasmic domains with activation of the Janus family tyrosine kinases, JAK1 (in association with β chain) and JAK3 (associating with the γc). Activated JAK1 and JAK3 then phosphorylate signal transducer and activator of transcription proteins STAT3 and STAT5, respectively, to mediate IL-2 and IL-15 effects in immune cells.
IL-2 is primarily produced by activated CD4+ T cells and, at lower amounts, by other immune cells (CD8+ T cells, NK cells, activated dendritic cells, and mast cells). It signals via combinations of IL-2 R subunits: IL-2 Rα (CD25), IL-2/IL-15 Rβ, and the γc. The combination IL-2 Rβ with γc forms intermediate-affinity IL-2 R; all three subunits together form the high-affinity IL-2 R. Both these classes of receptors are functional and are differentially expressed on immune cell subsets. In addition to being a potent T-cell growth factor, IL-2 is also an important modulator of the effector cell response. In particular, it is involved in the differentiation of Th1 and Th2, promotes antibody production and proliferation of B cells, and is essential for activation-induced cell death and for the development of CD4+FOXP3+ regulatory T cells (T-reg) (12).
IL-15 shares not only structural and signaling components with IL-2 but also many functional redundancies. In particular, both induce the proliferation and increase the cytolytic activity of NK and CD8+ T cells, and both can induce the proliferation and differentiation of stimulated human B cells. However, certain important immune functions are exclusively mediated by IL-15. In particular, IL-15 is critical for the development of NK cells and antigen-specific memory CD8+ T cells and is best characterized for its role in maintaining memory pools of CD8+ T cells (13).
The IL-2/IL-2 Rα and IL-15/IL-15 Rα cytokine loops are deregulated in HAM/TSP and contribute to a variety of immunological abnormalities characteristic of this disorder, providing a strong rationale for anticytokine therapy in this condition (14). Such therapy could potentially reduce clonal expansion and activation of the infected cells in the peripheral circulation, and subsequently decrease their infiltration and expansion in the CNS. On the basis of this rationale, monoclonal antibody therapy in HAM/TSP using humanized anti-Tac antibody (against IL-2 Rα) was conducted with encouraging results (i.e., reduction in the proviral load and spontaneous proliferation), and a clinical trial of humanized Mik β1, monoclonal antibody against IL-2/IL-15 Rβ, is currently underway, with promising early results (15).
However, we reasoned that simultaneous inhibition of the two IL-2/IL-15 cytokine loops is potentially a more effective anticytokine therapy than antibody monotherapy. Signaling molecules that are shared by the two loops, namely, the JAK3 molecule and the common γc, provide reasonable targets for this strategy. CP-690, 550, which was originally developed as a JAK3 inhibitor molecule, was shown to reduce in vitro immune activation in HAM/TSP PBMCs (16) and is currently approved for the treatment of psoriasis and rheumatoid arthritis. However, this agent also inhibits JAK1 and lacks cytokine specificity, which probably accounts for the high incidence of adverse effects observed with its clinical use; namely, bone marrow suppression, lymphocytopenia, and high incidence of serious infections (17).
A novel and potentially more effective and safe targeted therapeutic approach to HAM/TSP might consist of γc blockade with an antagonizing peptide that selectively inhibits IL-2, IL-9, and IL-15, but not other γc cytokines. BNZ132-1 is an anticytokine peptide that was designed on the basis of the presence of a moderately conserved region among all of the cytokines sharing the γc receptor and overlapping with the d-helix. It is a 19-mer peptide that, according to proliferation assays in cytokine-dependent human and murine cells lines, was found to selectively block signaling of IL-2, IL-15, and IL-9.
In this study, we examined the effects of BNZ132-1 on multiple in vitro markers of immune activation in HAM/TSP. The peptide significantly suppressed SP of the PBMCs and reduced the frequency of Tax-specific CD8+ T cells, the degree of spontaneous CD8+ T-cell degranulation, and the degree of STAT5 phosphorylation. Compared with a combination of monoclonal antibodies against CD25 and CD122, targeting the IL-2 and IL-15 cytokine loops, this novel antagonist peptide was found to be as effective in reducing STAT5 phosphorylation and more effective in inhibiting SP. Therefore, BNZ132-1 is a potential therapy for HAM/TSP.
Results
BNZ132-1 Inhibits HAM/TSP PBMC SP in a Dose-Dependent Manner.
Given the important role of IL-2 and IL-15 in SP of HAM/TSP PBMCs, the effects of the γc-blocking peptide BNZ132-1 on this phenomenon were examined. PBMCs from six patients with HAM/TSP were cultured either in complete RPMI alone or with BNZ132-1 at 1 and 10 μM. At day 5 of culture, the peptide reduced the degree of SP, as quantified by radioactive thymidine incorporation, in five of six samples (Fig. 1A) in a dose-dependent manner. No decrease in cell viability was observed at these concentrations. A PEGylated form of the peptide, BNZ132-1-40, was then developed and used in the subsequent experiments. At a concentration as low as 0.01 μM, BNZ132-1-40 inhibited the proliferation of CTLL-2, an IL2-dependent T-cell line that also responds to IL-15, and the inhibition was almost complete at a 10 μM concentration of BNZ132-1-40 (Fig. S1). Subsequently, a dose-titration experiment in one patient with HAM/TSP, HAM1, known to have IL-2/IL-15-dependent SP (based on the inhibition by BNZ132-1; Fig. 1A), showed that at 10 μM, BNZ132-1-40 inhibited SP by 60% (Fig. 1B, summarized in Fig. 1D). On the basis of these results, additional studies were conducted with more HAM/TSP PBMC samples. As shown in Fig. 1C, at 10 μM, BNZ132-1-40 inhibited the SP in all six patients with HAM/TSP tested. Again, no decrease in cell viability was observed with the PEGylated peptide. The percentage inhibition of SP by BNZ132-1-40 is summarized in Fig. 1D and ranged from 20% to 85%.
Fig. 1.

BNZ132-1 and its PEGylated form BNZ132-1-40 inhibit HAM/TSP PBMC spontaneous proliferation in a dose-dependent manner. (A) Spontaneous proliferation of PBMCs from six patients with HAM/TSP cultured with or without BNZ132-1 at 1 and 10 μM. (B) SP of PBMCs from a representative patient with HAM/TSP with and without BNZ132-1-40 at three different concentrations (1, 3, and 10 μM). (C) SP of PBMC from six patients with HAM/TSP with and without BNZ132-1-40 at 10 μM. (D) Percentage inhibition of SP by BNZ132-1-40 at 10 μM in six patients with HAM/TSP.
Fig. S1.

BNZ132-1-40 inhibits, in a dose-dependent manner, IL-2- and IL-15-mediated proliferation of CTLL-2. (A) Proliferation of CTLL-2 in the presence of IL-2 (10 IU/mL) and various concentrations of BNZ132-1-40. (B) Proliferation of CTLL-2 in the presence of IL-15 (5 ng/mL) and various concentrations of BNZ132-1-40. Proliferation of CTLL-2 was characterized at 24 h of culture with a metabolic assay using WST-1 reagent (Clontech) and quantified by an ELISA reader as the absorbance at 450 nm.
BNZ132-1-40 Inhibits the Proliferation of Both CD4+ and CD8+ T Cells.
Because CD4+ and CD8+ T cells are both involved in HTLV-1-induced SP, the effects of BNZ132-1-40 on the proliferation of each of the two subsets were examined by flow cytometry, using a carboxyfluorescein diacetate succinimidyl ester dilution assay in three patients. Fig. 2A is a representative histogram of the inhibitory effects of BNZ132-1-40 at 10 μM on the proliferation of CD4+ and CD8+ T-cell subsets from a patient with HAM/TSP (HAM5). In all three patients with HAM/TSP tested, BNZ132-1-40 reduced the SP of both CD4+ and CD8+ T cells (Fig. 2 A and B). Notably, there was no significant difference in the levels of inhibition of SP between CD4+ and CD8+ T cells (Fig. 2B), although the degree of proliferation of CD4+ T cells in the untreated samples was minimal compared with CD8+ cells (e.g., Fig. 2A).
Fig. 2.

BNZ132-1-40 (10 μM) inhibits SP in CD4+ and CD8+ T cells. (A) Representative histograms of carboxyfluorescein diacetate succinimidyl ester in CD4+ and CD8+ T cells of one patient with HAM/TSP (HAM5) cultured with or without BNZ132-1-40. (B) Percentage inhibition of SP of CD4+ and CD8+ T cells in three HAM/TSP samples by BNZ132-1-40. (C) Percentage inhibition of Ki-67 expression in CD4+ and CD8+ T-cell subsets from six patients with HAM/TSP.
Similar results were found when the differential effects on T-cell subset proliferation were examined by Ki-67 expression, another marker of proliferation. As summarized in Fig. 2C, BNZ132-1-40 at 10 μM reduced Ki-67 expression in both CD4+ and CD8+ T-cell subsets in all six HAM/TSP PBMCs. Again, although there was no statistically significant difference in the inhibition between CD8+ and CD4+ T cells, in four of six studied samples, the reduction of Ki-67 was more prominent in the CD8+ T-cell subset (Fig. 2C). Moreover, additional markers of activation such as pSTAT5 and CD25 expression were significantly more reduced in the CD8+ compared with the CD4+ T-cell subset (Fig. 3 A and B). The degree of inhibition of STAT5 phosphorylation in the CD8+ T cells at day 5 of culture strongly correlated with the inhibition of Ki-67 expression in this subset (Fig. 3C).
Fig. 3.
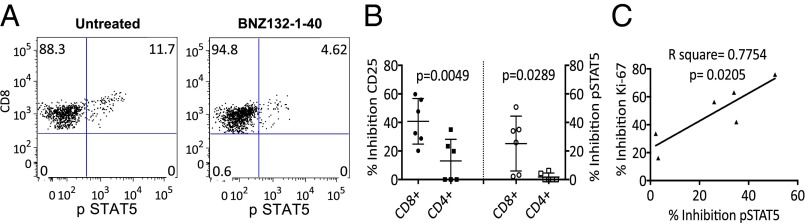
BNZ132-1-40 (10 μm) reduces CD25 expression and STAT5 phosphorylation in the CD8+ T cells. (A) Representative dot plots illustrating the expression of pSTAT5 in CD8+ T cells from one patient with HAM/TSP when PBMCs were cultured with and without BNZ132-1-40. (B) Percentage inhibition of CD25 and pSTAT5 expression in CD8+ and CD4+ T cells of six different patients with HAM/TSP at day 5 of SP by BNZ132-1-40. (C) The inhibition of pSTAT5 in the CD8+ T cells strongly correlates with the reduction in SP of this T-cell subset.
BNZ132-1-40 Reduces the Expression of CD132, the Common γc, in HAM/TSP PBMC.
In addition to sharing the common γc for signal transduction, IL-2 and IL-15 share the cell-surface receptor subunit IL-2/IL-15 Rβ (CD122) known to be critical for IL-15-mediated CD8+ T-cell activation, memory cell survival, and cytotoxic activity (18). In addition, CD25, the IL-2 private receptor chain α, is up-regulated in HAM/TSP and mediates many of the effects of IL-2 in the CD4+ T cells. Thus, we examined the effects of γc blockade by BNZ132-1-40 on the expression of these three receptor subunits (CD25, CD122, and CD132). HAM/TSP PBMCs from three subjects were cultured for 48 h with or without BNZ132-1-40, and the expression of the receptor chains was characterized at 48 h. BNZ132-1-40 significantly reduced the expression of CD132, both in CD4+ and CD8+ T cells (Fig. 4A), whereas there was no change in the expression of CD25 or CD122 on either of the cell subsets. In addition, the degree of reduction in CD132 expression in the CD8+ T cells strongly correlated with the percentage inhibition of day 5 SP by BNZ132-1-40 in the same samples (Fig. 4B).
Fig. 4.

BNZ132-1-40 significantly reduces the expression of CD132 in cultured HAM/TSP PBMC. (A) Inhibition of CD132 expression in CD4+ and CD8+ T cells with BNZ132-1-40 (10 μM) at 48 h of culture. (B) The inhibition of CD132 expression in CD8+ T cells correlates with inhibition of SP by BNZ132-1-40.
BNZ132-1-40 Is More Effective Than a Combination of Anti-Tac and Mikβ1 Antibodies in Inhibiting HAM/TSP Spontaneous Proliferation.
Murine anti-CD25 (anti-Tac) and anti-CD122 (Mikβ1) antibodies are known to inhibit HAM/TSP SP and have been important tools in elucidating the roles of IL-2 and IL-15 in this phenomenon. Anti-Tac targets the CD25 (IL-2 Rα) and inhibits IL-2 action. Mikβ1 binds to the IL-2/15 β chain, but only inhibits IL-15 actions that are mediated by transpresentation through the high-affinity receptor complex (IL-15 Rα, IL-15 Rβ, and IL-15 Rγ). Because each antibody inhibits either the IL-2 or IL-15 pathway, we compared the effects of BNZ132-1-40 peptide on pSTAT5 expression and SP with those of a combination of anti-Tac and MiKβ1. At 10 μΜ, BNZ132-1-40 was as effective as the combination of anti-Tac and MiK β1 (each 5 μM) in inhibiting STAT5 phosphorylation in HAM/TSP lymphocytes (Fig. 5A) and was significantly more effective in inhibiting SP (P = 0.0325). (Fig. 5B).
Fig. 5.

BNZ132-1-40 is more effective than a combination of blocking anti-CD25 and anti-CD122 antibodies in reducing HAM/TSP PBMC SP. (A) Inhibition of pSTAT5 expression in PBMCs from three patients with HAM/TSP with BNZ132-1-40 (10 μM) or with a combination of Anti-Tac and Mikβ1 (each 5 μM). STAT5 phosphorylation in the lymphocytes was examined at 20 h of culture. (B) Inhibition of PBMC SP in three patients with HAM/TSP with BNZ132-1-40 (10 μM) or with a combination of Anti-Tac and Mikβ1 (each 5 μM).
BNZ132-1-40 Reduces the Frequency of Tax-Tetramer-Positive CD8+ T Cells.
IL-15 increases the proliferation, reduces the apoptosis, and enhances the cytotoxicity of HTLV-1-specific CD8+ CTLs (18). The effects of BNZ132-1-40 on the frequency of HTLV-1 Tax-specific CD8+ T cells were therefore examined in two HLA-A*0201 HAM/TSP individuals after culture. As shown in Fig. 6, both HAM/TSP patient 9 and patient 10 had Tax11-19 tetramer reactive cells after 6 d of culture that were reduced in the presence of 10 μM BNZ132-1-40. The frequency of Tax-specific CD8+ T cells was reduced from 14.4% to 4%, and from 4.68% to 1.9%, respectively (Fig. 6).
Fig. 6.

BNZ132-1-40 (10 μm) reduces the SP of Tax-specific CD8+ T cells. PBMCs from two HLA-A 0201 HAM/TSP subjects (HAM9 and HAM10) were cultured for 6 d with and without BNZ132-1-40.
BNZ132-1-40 Reduces HTLV-1-Induced Cytolytic T-Cell Degranulation.
It has been reported that patients with HAM/TSP demonstrate increased spontaneous degranulation (CD107a) and IFN-γ secretion by CD8+ CTLs in unstimulated ex vivo culture compared with asymptomatic carriers and healthy donors (19). This response is HTLV-1-specific, as it requires antigen presentation and IL-15 transpresentation to effector CD8+ T cells by infected CD14+ cells. Notably, in HAM/TSP there is increased IL-15 expression on the surface of CD14+ antigen presenting cells (APC) and increased responsiveness of CD8+ CTLs to IL-15 (19).
Therefore, it was of interest to examine the effects of BNZ132-1-40 on the frequency of CD107a (a marker of the mobilization of cytolytic granules to the cell surface) and the production of IFN-γ by CD8+ T cells from patients with HAM/TSP during short-term culture. Fig. 7A is a flow-cytometric dot plot illustrating the inhibitory effect of BNZ132-1-40 on the spontaneous degranulation and IFN-γ expression by CD8+ CTLs from a representative HAM/TSP subject. As shown in Fig. 7 B and C, patients with HAM/TSP displayed variable frequencies of spontaneously degranulating/IFN-γ-expressing CTLs, which were reduced by BNZ132-1-40 by more than 50%, in three of four tested subjects.
Fig. 7.

BNZ132-1-40 (10 μm) inhibits the spontaneous degranulation and IFN-γ secretion by CD8+ cytotoxic T cells. (A) Representative dot plots from one patient with HAM/TSP illustrating spontaneous degranulation and IFN-γ secretion by CD8+ CTLs. (B) Spontaneous degranulation and IFN-γ secretion by CD8+ CTLs from four patients with HAM/TSP. (C) Percentage inhibition, by BNZ132-1-40, of spontaneous degranulation and IFN-γ secretion by CD8+ CTLs from the four tested patients with HAM/TSP.
Collectively, these results demonstrate that common γc blockade by BNZ132-1-40 is effective in reducing a number of IL-2- and IL-15-mediated immunological effects associated with HAM/TSP pathogenesis.
Discussion
In this study, a new strategy to reduce the excessive immune activation that occurs in HAM/TSP was used by simultaneously inhibiting the effects of multiple proinflammatory autocrine and paracrine cytokine loops involved in the pathogenesis of this disease. It has long been known that HAM/TSP PBMCs spontaneously proliferate in ex vivo cultures and that this immunological finding might reflect naturally occurring events in vivo in patients with HAM/TSP, particularly at the site of injury in the CNS. Thus, ex vivo SP is an important tool when evaluating potential interventional strategies and specific therapeutic agents in HAM/TSP.
Many of the immune abnormalities observed in HAM/TSP (SP, differentiation, increased cytotoxicity, and prolonged survival of HTLV-1-specific CTLs) have been linked to the HTLV-1 Tax. HTLV-1 Tax, directly and through activation of NF-κB, induces the expression of IL-2, IL-15, and their specific receptors (IL-2 Rα and IL-15 Rα), leading to excessive signaling through proinflammatory autocrine and paracrine loops (20).
It was previously demonstrated that IL-2/IL-2 Rα and IL-15/IL-15 Rα are overexpressed in HAM/TSP PBMCs and have additive effects on SP, which can be inhibited by the addition of antibodies to the cytokines or their receptors (IL-2 Rα and IL-15 Rβ). IL-9, another Tax-induced γ-cytokine, plays a role in the proliferation of adult T-cell leukemia (ATL) cells. Unlike IL-2 and IL-15, which act through autocrine loops on the T cells, IL-9 stimulates the IL-9 Rα-expressing CD14+ monocytes, which then support the proliferation of ATL cells via a paracrine loop (21). In vitro studies have shown that in addition to its role in SP, IL-15 also contributes to the survival of Tax-specific CD8+ T cells (supporting their proliferation and inhibiting apoptosis) and their cytotoxic function, both of which are increased in HAM/TSP and are thought to be critical mediators of CNS injury, as discussed previously.
Therefore, the IL-2/IL-15 cytokine loops constitute a reasonable therapeutic target in HAM/TSP. This is supported by the clinical trial of humanized anti-Tac antibody (against IL-2 Rα), showing a reduction in the proviral load and SP, and is currently being explored in a phase 1/2 clinical trial of humanized Mikβ1Ab directed against IL-2/IL-15 Rβ. As an alternative approach, we investigated the effects of a novel immunomodulatory γc blocking peptide, BNZ132-1, on HAM/TSP PBMCs. BNZ132-1 was designed to selectively inhibit the functions of some of the γc cytokines (IL-2, IL-9, and IL-15), but not others (IL-4, IL-7, and IL-21). BNZ132-1 was demonstrated to be a potent inhibitor of HAM/TSP SP. To improve the pharmacokinetic properties of the peptide, increase its bioavailability, and reduce its immunogenicity for potential future clinical studies, a PEGylated form of BNZ132-1, BZN132-1-40, was developed. BNZ-132-40 was also shown to inhibit SP, using a variety of immunological assays. Notably, the peptide suppressed the proliferation of both CD4+ and CD8+ T cells. In addition, in short-term culture of HAM/TSP PBMCs, the peptide reduced STAT5 phosphorylation, a downstream event of γc signaling, significantly more in CD8+ compared with CD4+ T cells (P = 0.028), suggesting preferential inhibitory effects on this subset. The inhibition of pSTAT5 expression and SP by BNZ132-1-40 peptide was secondary to the reduction in CD132 expression, as this was significantly reduced in both CD4+ and CD8+ T cells, whereas there were no changes in the expression of CD25 or CD122 molecules. It remains unclear, however, whether this reflects down-regulation of the γc or its saturation by the peptide. Remarkably, compared with a combination of anti-Tac and Mikß1 antibodies, BNZ132-1-40 was as effective in inhibiting STAT5 phosphorylation, but significantly more effective in reducing PBMC SP. This is possibly mediated by blocking the effects of another γc cytokine, IL-9, which is also suggested to play a role in HAM/TSP SP. It has been previously shown that IL-9 contributes to the proliferation of malignant HTLV-1-related ATL cells, but its importance in HAM/TSP remains to be established.
Virus-specific cytotoxic CD8+ T cells are thought to be major players in the pathogenesis of HAM/TSP. They are enriched in the peripheral circulation and cerebrospinal fluid of affected patients and demonstrate increased cytotoxicity, which is at least partially mediated by their interaction with IL-15 overexpressing CD14+ cells. BNZ132-1-40 also was found to reduce the SP of Tax-specific CD8+ T cells in two patients with HAM/TSP, which is consistent with the role of IL-15 in the differentiation and prolonged survival of these cells. In addition, the peptide reduced the degree of spontaneous degranulation and IFN-γ secretion by CD8+ CTLs in short-term culture, illustrating the efficacy of the peptide in reducing the stimulation of virus-specific CTL response by antigen-presenting CD14+ cells.
This approach highlights a promising therapeutic strategy for HAM/TSP and overcomes the challenge posed by the deregulation of multiple proinflammatory cytokines. The understanding and potential reversal of immunologic mechanisms implicated in a rare, retrovirus-associated human neurologic condition, such as HAM/TSP, could also help develop treatments for inflammatory conditions of unknown etiology. Similarly, understanding the pathogenic roles of the CD8+ CTLs and other inflammatory cytokines in HAM/TSP can be translated to other conditions in which similar immune abnormalities might be pathogenic. In particular, the anticytokine strategy described in this study is a promising therapeutic approach for many inflammatory disorders related to the γc cytokines; such conditions include but are not limited to myelofibrosis, rheumatoid arthritis, psoriasis, and celiac disease (22–24).
Methods
Samples.
Blood was collected from patients with HAM/TSP seen at the National Institute of Neurologic Disorders and Stroke. PBMCs were isolated by density centrifugation and cryopreserved before use. Written, informed consent was obtained in compliance with the Declaration of Helsinki. The study was reviewed and approved by a National Institute of Neurologic Disorders and Stroke institutional review board.
Cell Culture.
PBMCs were suspended in RPMI supplemented with 1% penicillin/streptomycin, 1% L-glutamine, and 5% (vol/vol) Human AB serum and placed in 96-well round-bottom plates at 3 × 105 cells per well, with or without BNZ132-1 or its pegylated form (BNZ132-1-40), at the indicated concentration. In some experiments, HAM/TSP PBMCs were cultured in the presence of the antibodies murine anti-Tac (5 μM) and Mik-β1 (5 μM) (kind gifts from Thomas A. Waldmann, NIH, Bethesda). Cell viability was measured using GuavaViaCount assay (Millipore) according to the manufacturer’s instructions.
Proliferation Assay.
HAM/TSP PBMCs were cultured as described earlier in triplicate. Proliferation was measured by radioactive thymidine incorporation on day 5 of culture. PHA-treated and untreated healthy donor PBMCs were used as positive and negative controls, respectively.
Alternatively, HAM/TSP PBMCs were labeled with carboxyfluorescein diacetate succinimidyl ester (Life Technologies), according to the manufacturer's instructions, and cultured as described earlier without or with BNZ132-1-40 at 10 μM. At day 5, the cells were washed, stained for cell-surface markers [CD3-pacific blue (PB), CD4-APC, and CD8-PerCP-Cy5.5 (BD Biosciences)], and the data acquired on LSRII flow cytometer (BD Biosciences) and analyzed using FlowJo software (Tree star).
The effects of BNZ132-1–40 on the proliferation of different lymphocyte subsets were also examined by Ki-67 stain at day 5 of culture. For this, PBMCs were cultured without or with BNZ132-1-40 at 10 μM. At day 5, the cells were stained for cell-surface markers [CD3-PB, CD4-Alexa Fluor700, CD8-PerCP-Cy5.5, and CD25-APC (BD Biosciences)] and then fixed and permeabilized using the Foxp3 Fix/Perm kit (eBioscience), according to the manufacturer’s instructions, before staining with FITC-conjugated Ki-67 antibody (BD Biosciences).
Activation Markers Analysis.
For phosphorylated STAT5 (pSTAT5) detection, HAM/TSP PBMCs were cultured as described earlier, with or without BNZ132-1-40, at 10 μM for 20 h or 5 d. The cells were then fixed in 1.5% (vol/vol) paraformaldehyde and permeabilized with 90% (vol/vol) methanol. After washing, the cells were stained with antibodies against pSTAT5-PE, CD3-APC, CD4-FITC, and CD8-PE-Cy7 (BD Biosciences).
The effects of BNZ132-1-40 peptide on the expression of CD122, CD25, and CD132 were examined by flow cytometry. HAM/TSP PBMCs were stained with CD3-PB, CD4-Alexa Fluor700, CD8-PerCP-Cy5.5, CD122-PE, CD25-FITC (BD Biosciences), and CD132-APC (Biolegend) at baseline and after culture at 24 and 48 h.
Analysis of Tax-Specific Cells and CTL Spontaneous Degranulation.
Analysis of antigen-specific CD8+ T cells was performed using a PE-conjugated HTLV-1 Tax11–19 peptide (LLFGYPVYV)-loaded HLA-A*0201 tetramer (provided by National Institutes of Health AIDS Research and Reference Reagent Program). PBMCs from patients with HAM/TSP known to have the allele HLA A*0201 were cultured without or with BNZ132-1-40 at 10 μM. At day 6 of culture, the cells were stained with Tax-tetramer, CD3-PB, CD4-APC, and CD8-PerCP-Cy5.5 (BD Biosciences).
For spontaneous CTL degranulation, HAM/TSP PBMCs were cultured in a 96-well U-bottom plate at 2 × 105 cells per well, with or without BNZ132-1-40 10 μM, and analysis was performed as previously described (19).
Statistical Analysis.
Paired t test was used to compare the differential effects of BNZ132-1-40 on the proliferation (Ki-67) and activation (pSTAT5 and CD25 expression) in the different lymphocyte subsets (CD4+ vs. CD8+ T cells) and to compare the effects of BNZ132-1-40 (on SP and pSTAT5) with those of a combination of monoclonal antibodies (anti-CD25 and anti-CD122). Linear regression analysis was used to examine the correlation between the effects on pSTAT5 and CD132 and the inhibition of SP. All statistical analysis was done using Prism software version 6.0 (Graphpad).
Footnotes
Conflict of interest statement: N.A. and A.B. are employees of Bioniz, the developer of the peptide used in this study.
*This Direct Submission article had a prearranged editor.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1412626112/-/DCSupplemental.
References
- 1.Proietti FA, Carneiro-Proietti AB, Catalan-Soares BC, Murphy EL. Global epidemiology of HTLV-I infection and associated diseases. Oncogene. 2005;24(39):6058–6068. doi: 10.1038/sj.onc.1208968. [DOI] [PubMed] [Google Scholar]
- 2.Araujo AQ, Silva MT. The HTLV-1 neurological complex. Lancet Neurol. 2006;5(12):1068–1076. doi: 10.1016/S1474-4422(06)70628-7. [DOI] [PubMed] [Google Scholar]
- 3.Lima MA, Harab RC, Schor D, Andrada-Serpa MJ, Araújo AQ. Subacute progression of human T-lymphotropic virus type I-associated myelopathy/tropical spastic paraparesis. J Neurovirol. 2007;13(5):468–473. doi: 10.1080/13550280701510096. [DOI] [PubMed] [Google Scholar]
- 4.Umehara F, et al. Immunocytochemical analysis of the cellular infiltrate in the spinal cord lesions in HTLV-I-associated myelopathy. J Neuropathol Exp Neurol. 1993;52(4):424–430. doi: 10.1097/00005072-199307000-00010. [DOI] [PubMed] [Google Scholar]
- 5.Aye MM, et al. Histopathological analysis of four autopsy cases of HTLV-I-associated myelopathy/tropical spastic paraparesis: Inflammatory changes occur simultaneously in the entire central nervous system. Acta Neuropathol. 2000;100(3):245–252. doi: 10.1007/s004019900170. [DOI] [PubMed] [Google Scholar]
- 6.Sakai JA, Nagai M, Brennan MB, Mora CA, Jacobson S. In vitro spontaneous lymphoproliferation in patients with human T-cell lymphotropic virus type I-associated neurologic disease: Predominant expansion of CD8+ T cells. Blood. 2001;98(5):1506–1511. doi: 10.1182/blood.v98.5.1506. [DOI] [PubMed] [Google Scholar]
- 7.Yamano Y, et al. Correlation of human T-cell lymphotropic virus type 1 (HTLV-1) mRNA with proviral DNA load, virus-specific CD8(+) T cells, and disease severity in HTLV-1-associated myelopathy (HAM/TSP) Blood. 2002;99(1):88–94. doi: 10.1182/blood.v99.1.88. [DOI] [PubMed] [Google Scholar]
- 8.Jacobson S, Shida H, McFarlin DE, Fauci AS, Koenig S. Circulating CD8+ cytotoxic T lymphocytes specific for HTLV-I pX in patients with HTLV-I associated neurological disease. Nature. 1990;348(6298):245–248. doi: 10.1038/348245a0. [DOI] [PubMed] [Google Scholar]
- 9.Kubota R, Kawanishi T, Matsubara H, Manns A, Jacobson S. Demonstration of human T lymphotropic virus type I (HTLV-I) tax-specific CD8+ lymphocytes directly in peripheral blood of HTLV-I-associated myelopathy/tropical spastic paraparesis patients by intracellular cytokine detection. J Immunol. 1998;161(1):482–488. [PubMed] [Google Scholar]
- 10.Johnson-Nauroth JM, Graber J, Yao K, Jacobson S, Calabresi PA. Memory lineage relationships in HTLV-1-specific CD8+ cytotoxic T cells. J Neuroimmunol. 2006;176(1-2):115–124. doi: 10.1016/j.jneuroim.2006.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Yamane H, Paul WE. Cytokines of the γ(c) family control CD4+ T cell differentiation and function. Nat Immunol. 2012;13(11):1037–1044. doi: 10.1038/ni.2431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Liao W, Lin JX, Leonard WJ. Interleukin-2 at the crossroads of effector responses, tolerance, and immunotherapy. Immunity. 2013;38(1):13–25. doi: 10.1016/j.immuni.2013.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Verbist KC, Klonowski KD. Functions of IL-15 in anti-viral immunity: Multiplicity and variety. Cytokine. 2012;59(3):467–478. doi: 10.1016/j.cyto.2012.05.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Azimi N, Jacobson S, Leist T, Waldmann TA. Involvement of IL-15 in the pathogenesis of human T lymphotropic virus type I-associated myelopathy/tropical spastic paraparesis: Implications for therapy with a monoclonal antibody directed to the IL-2/15R beta receptor. J Immunol. 1999;163(7):4064–4072. [PubMed] [Google Scholar]
- 15.Lehky TJ, et al. Reduction in HTLV-I proviral load and spontaneous lymphoproliferation in HTLV-I-associated myelopathy/tropical spastic paraparesis patients treated with humanized anti-Tac. Ann Neurol. 1998;44(6):942–947. doi: 10.1002/ana.410440613. [DOI] [PubMed] [Google Scholar]
- 16.Ju W, et al. CP-690,550, a therapeutic agent, inhibits cytokine-mediated Jak3 activation and proliferation of T cells from patients with ATL and HAM/TSP. Blood. 2011;117(6):1938–1946. doi: 10.1182/blood-2010-09-305425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Salgado E, Maneiro JR, Carmona L, Gomez-Reino JJ. Safety profile of protein kinase inhibitors in rheumatoid arthritis: Systematic review and meta-analysis. Ann Rheum Dis. 2014;73(5):871–882. doi: 10.1136/annrheumdis-2012-203116. [DOI] [PubMed] [Google Scholar]
- 18.Azimi N, Nagai M, Jacobson S, Waldmann TA. IL-15 plays a major role in the persistence of Tax-specific CD8 cells in HAM/TSP patients. Proc Natl Acad Sci USA. 2001;98(25):14559–14564. doi: 10.1073/pnas.251540598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Enose-Akahata Y, Oh U, Grant C, Jacobson S. Retrovirally induced CTL degranulation mediated by IL-15 expression and infection of mononuclear phagocytes in patients with HTLV-I-associated neurologic disease. Blood. 2008;112(6):2400–2410. doi: 10.1182/blood-2008-02-138529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Peloponese JM, Yeung ML, Jeang KT. Modulation of nuclear factor-kappaB by human T cell leukemia virus type 1 Tax protein: Implications for oncogenesis and inflammation. Immunol Res. 2006;34(1):1–12. [PubMed] [Google Scholar]
- 21.Chen J, et al. Induction of the IL-9 gene by HTLV-I Tax stimulates the spontaneous proliferation of primary adult T-cell leukemia cells by a paracrine mechanism. Blood. 2008;111(10):5163–5172. doi: 10.1182/blood-2007-09-113654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Abadie V, Discepolo V, Jabri B. Intraepithelial lymphocytes in celiac disease immunopathology. Semin Immunopathol. 2012;34(4):551–566. doi: 10.1007/s00281-012-0316-x. [DOI] [PubMed] [Google Scholar]
- 23.Dunphy S, Gardiner CM. NK cells and psoriasis. J Biomed Biotechnol. 2011;2011:248317. doi: 10.1155/2011/248317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Tefferi A, et al. Circulating interleukin (IL)-8, IL-2R, IL-12, and IL-15 levels are independently prognostic in primary myelofibrosis: A comprehensive cytokine profiling study. J Clin Oncol. 2011;29(10):1356–1363. doi: 10.1200/JCO.2010.32.9490. [DOI] [PubMed] [Google Scholar]