

Abstract
Behavioral research has demonstrated that the shape and texture of single objects can be processed independently. Similarly, neuroimaging results have shown that an object's shape and texture are processed in distinct brain regions with shape in the lateral occipital area and texture in parahippocampal cortex. Meanwhile, objects are not always seen in isolation and are often grouped together as an ensemble. We recently showed that the processing of ensembles also involves parahippocampal cortex and that the shape and texture of ensemble elements are processed together within this region. These neural data suggest that the independence seen between shape and texture in single-object perception would not be observed in object-ensemble perception. Here we tested this prediction by examining whether observers could attend to the shape of ensemble elements while ignoring changes in an unattended texture feature and vice versa. Across six behavioral experiments, we replicated previous findings of independence between shape and texture in single-object perception. In contrast, we observed that changes in an unattended ensemble feature negatively impacted the processing of an attended ensemble feature only when ensemble features were attended globally. When they were attended locally, thereby making ensemble processing similar to single-object processing, interference was abolished. Overall, these findings confirm previous neuroimaging results and suggest that distinct cognitive mechanisms may be involved in single-object and object-ensemble perception. Additionally, they show that the scope of visual attention plays a critical role in determining which type of object processing (ensemble or single object) is engaged by the visual system.
Keywords: object-ensemble perception, single-object perception, texture perception, shape perception, Garner interference
Introduction
Object shape and surface texture are two informative cues to successful object perception and recognition. Although shape is perhaps the most heavily studied cue in object perception and recognition (Marr & Nishihara, 1978; Biederman, 1987; Biederman & Cooper, 1991; Grill-Spector et al., 1998; Wilson & Wilkinson, 1998; Kourtzi & Kanwisher, 2000), evidence from behavioral psychophysics, functional magnetic resonance imaging (fMRI), computational modeling, and neuropsychology has demonstrated that surface texture also plays an important role in object perception and recognition (Rossion & Pourtois, 2004; James, Culham, Humphrey, Milner, & Goodale, 2003; Adelson, 2001, 2008; Cavina-Pratesi, Kentridge, Heywood, & Milner, 2010a, 2010b).
In our everyday phenomenological experience with objects, we do not typically perceive objects as a bundle of isolated visual features, such as outline shape or inner surface texture. Rather, we perceive a unified and whole object. Thus, to gain a deeper understanding of object processing, it is important to investigate how the visual features of the same object are processed with respect to each other. In single-object perception, behavioral research has demonstrated that the shape and texture of objects can be processed independently, specifically in cases in which texture flags the material that an object is made of (Cant, Large, McCall, & Goodale, 2008; but see Cant & Goodale, 2009). Moreover, fMRI research has demonstrated that the shape and the texture of single objects are processed in separate cortical regions with shape being processed in the lateral occipital area (LO; Grill-Spector et al., 1999) and texture being processed in the scene-sensitive parahippocampal place area (PPA; Epstein & Kanwisher, 1998; Cant & Goodale, 2007, 2011; Cant, Arnott, & Goodale, 2009; Cant & Xu, 2012). Taken together, these behavioral and neuroimaging findings indicate that there are separate cognitive and neural mechanisms for the processing of shape and texture in single-object perception.
But in our everyday visual experience, we rarely encounter isolated objects. In many cases, multiple objects are grouped together into a perceptual unit known as an object ensemble (e.g., a bowl filled with fruit, a flock of birds, leaves on a tree). Behavioral investigations have demonstrated that observers can extract summary statistical information from ensembles of objects, such as their mean size, direction of motion, speed, orientation, center location, and identity of crowds, without being able to provide fine details about any individual object in the ensemble (e.g., Williams & Sekuler, 1984; Watamaniuk & Duchon, 1992; Ariely, 2001; Parkes, Lund, Angelucci, Solomon, & Morgan, 2001; Chong & Treisman, 2003; Alvarez & Oliva, 2008; Sweeny, Haroz, & Whitney, 2012; Yamanashi Leib et al., 2014). This dissociation between single-object and object-ensemble processing demonstrates an important and adaptive feature of ensemble processing. Namely, the representation of summary statistics from ensembles of multiple objects complements and guides object-specific processing as it allows the visual system to overcome the capacity limitation inherent in object-based attention (e.g., Pylyshyn & Storm, 1988; Luck & Vogel, 1997; Xu, 2002; Alvarez & Cavanagh, 2004).
Recently, we demonstrated that object ensembles and visual texture patterns are both represented in parahippocampal cortex in a region overlapping with PPA (Cant & Xu, 2012). This finding is consistent with previous reports of texture sensitivity in this region (Peuskens et al., 2004; Cant & Goodale, 2007, 2011). Shared neural mechanisms for ensemble and texture representations likely reflect the fact that both contain repeating elements that can vary in features such as size, orientation, and color (Portilla & Simoncelli, 2000). Interestingly, although previous fMRI studies focusing on single-object perception found that shape and texture were processed in separate brain regions (e.g., Cant & Goodale, 2007, 2011), we found that the region overlapping PPA was sensitive to processing changes in both the texture and the shape of the individual objects within an ensemble (Cant & Xu, 2011). This finding suggests that shape and texture may be processed interactively in ensemble perception and not independently as they are in single-object perception (Cant et al., 2008).
To test this prediction, here in six behavioral experiments, we compare the processing of shape and texture in both single-object and object-ensemble perception. Based on previous behavioral and fMRI experiments, we predicted that shape and texture would always be processed independently in single-object perception but may be processed interactively or independently, depending on how observers are engaged in object-ensemble perception. Specifically, when observers attend to the entire ensemble via a global-processing strategy to extract the summary representation of a particular visual feature, we expect to see interference between shape and texture processing. However, when a global-processing strategy is not employed, for example, when observers attend to the individual objects in an ensemble via a local-processing strategy, we expect to see less interference between shape and texture processing as this is akin to single-object perception.
In our experiments, we used either photographed (Experiments 1, 2, 3, 4, and 5) or computer-generated (Experiment 6) textured stone beads as our stimuli. In all experiments, the stone beads appeared in two different types of textures and two different shapes (see Figure 1 for an example). In Experiments 1, 3, and 4, ensembles of beads were shown, and each ensemble contained beads of the same exact shape with slight variation in surface textures (Experiment 2 focused solely on single-object perception). Thus, the perception of a single bead in an ensemble would allow the observers to determine the exact shape of the beads comprising that ensemble, but the perception of multiple beads from an ensemble would be required to aid in the perception of the texture category of what these beads were made from. Thus, in these experiments, the shape manipulation encourages a local-processing strategy and likely produces independence between shape and texture processing in the ensemble shape discrimination task. In contrast, the texture manipulation encourages a global-processing strategy and likely produces interference between shape and texture processing in the ensemble texture discrimination task.
Figure 1.

Stimuli and results from Experiment 1. In the baseline blocks (blue bars), only the relevant (i.e., attended) feature varied, and in filtering blocks (red bars), both the relevant and irrelevant (i.e., unattended) features varied. Stimuli are shown for illustrative purposes only and thus are not displayed at the actual size that was used in the experiment (this applies to all subsequent figures). Error bars represent within-subject SEM. *p < 0.05.
In Experiment 5, we explored whether inducing a local-processing strategy for ensemble texture would also produce independence between shape and texture as we predicted it would for ensemble shape in Experiments 1, 3, and 4. To this end, the exact same texture pattern repeated across beads so that perception of a single bead was sufficient to determine the texture of all beads in the ensemble (see Figure 5). Furthermore, in Experiment 6, we investigated whether inducing a global-processing strategy for ensemble shape would result in interference between shape and texture as we predicted it would for ensemble texture in Experiments 1, 3, and 4. Thus, in the final experiment, ensembles were comprised of objects with slight variations in both shape and surface texture, encouraging a global-processing strategy for both features (see Figure 6). Finally, we manipulated stimulus duration across experiments (Experiments 1, 2, and 3: 200 ms; Experiments 4, 5, and 6: until response or 2000 ms, whichever came first) to investigate if varying the time course of ensemble perception affected whether or not we would observe interference between shape and texture.
Figure 5.

Stimuli and results from Experiment 5 in which redundant (i.e., homogeneous) ensemble textures were used. In the baseline blocks (blue bars), only the relevant (i.e., attended) feature varied, and in filtering blocks (red bars), both the relevant and irrelevant (i.e., unattended) features varied. Error bars represent within-subject SEM. *p < 0.05.
Figure 6.

Stimuli and results from Experiment 6 in which ensembles contained slight variations in both shape and texture, encouraging a global processing strategy for both features. In the baseline blocks (blue bars), only the relevant (i.e., attended) feature varied, and in filtering blocks (red bars), both the relevant and irrelevant (i.e., unattended) features varied. Error bars represent within-subject SEM. *p < 0.05.
In summary, we predicted an asymmetry in ensemble feature perception, in that ensemble shape and texture would be processed independently when attention is focused locally (similar to the processing of single objects), and, conversely, that ensemble shape and texture would be processed interactively when attention is diffusely deployed over many objects in the ensemble in a global manner. To be clear, our investigation of object-ensemble processing differs from many previous studies in that we did not ask observers to make explicit judgments about the summary statistical representation of a particular ensemble feature and compare it with the same feature in a single object. For example, we did not ask observers to report the mean shape or mean texture of ensembles. Instead, we asked observers to discriminate between two values of an attended ensemble feature and constructed our stimuli and tasks to be able to manipulate how observers processed ensembles (i.e., locally or globally). This allowed us to compare if the processing of visual features differs when attending a single object from an array of multiple redundant objects (i.e., a single object, and not the entire ensemble, is treated as the meaningful perceptual unit) versus when attending to all objects within the array (i.e., the entire ensemble, and not a single object, is treated as the meaningful perceptual unit). The latter situation is akin to more natural object-ensemble perception and likely involves the same cognitive mechanisms underlying explicit judgments of summary statistics from arrays of multiple objects.
We used the well-established Garner speeded-classification task (Garner, 1974) to assess independence. Garner's task measures how efficiently an observer can attend to changes in one feature of an object (e.g., shape) while ignoring changes in a second, unattended feature (e.g., texture). If observers can effectively ignore changes in texture, for example, while classifying differences in shape, then these two features are labeled as separable and thus are processed independently. Examples of separable features include the position of lines and their luminance contrast (Shechter & Hochstein, 1992) and the shape (i.e., aspect ratio) and texture/material of single objects (Cant et al., 2008).If, however, observers cannot ignore changes in texture while attending to differences in shape, then these features are labeled as integral and are processed interactively, resulting in Garner interference. Integral features include the length of lines and their orientation (Dick & Hochstein, 1988) and the length and width of objects (Felfoldy, 1974; Dykes & Cooper, 1978; Ganel & Goodale, 2003; Cant et al., 2008; Cant & Goodale, 2009). In the Garner task, independence and interference are assessed by comparing observers' performance (e.g., response latency) in blocks of trials in which only the attended feature varies (baseline blocks) with blocks in which both the attended and unattended features vary (filtering blocks). If performance does not differ across baseline and filtering blocks, the two features are processed independently. In contrast, slower responses in the filtering compared with the baseline blocks demonstrates Garner interference and indicates that the two features are not processed independently.
Across six behavioral experiments, we report multiple replications of the finding that shape and texture are processed independently in single-object perception, but show that these features may not be processed independently in object-ensemble perception, depending on how observers are engaged in ensemble processing. Specifically, processing ensembles globally leads to interference whereas processing ensembles more locally eliminates such interference. Our results held even when differences in the size, location, color, and luminance of visual stimuli were controlled for (Experiments 2, 3, 4, and 6, respectively). Together, these findings demonstrate that distinct cognitive mechanisms are involved in single-object and object-ensemble perception.
General methods
Observers
Seventy-eight observers participated in this study (Experiment 1: 16 observers, 10 female, six male, all right-handed, mean age = 19.81 years, age range = 18–32 years; Experiment 2: 12 observers, eight female, four male, all right-handed, mean age = 20.83 years, age range = 18–24 years; Experiment 3: 10 observers, eight female, two male, all right-handed, mean age = 24.50 years, age range = 18–31 years; Experiment 4: 13 observers, seven female, six male, 10 right-handed, three left-handed, mean age = 26.31 years, age range = 19–35 years; Experiment 5: 12 observers, six female, six male, all right-handed, mean age = 27.92 years, age range = 20–36 years; Experiment 6: 15 observers, 10 female, five male, 14 right-handed, one left-handed, mean age = 20.67 years, age range = 18–24 years). No observer took part in more than one experiment. The observers were selected from research assistants, undergraduate students, graduate students, and post-doctoral fellows at either Harvard University (Experiments 2, 3, and 4) or The University of Toronto Scarborough (Experiments 1, 5, and 6). Observers had normal or corrected-to-normal visual acuity and reported no history of neurological impairment. They received course credit or $10 for their participation. All observers gave their informed consent in accordance with the Declaration of Helsinki. The experiments in this study were approved by the Committee on the Use of Human Subjects at Harvard University and by the Office of Research Ethics at the University of Toronto.
As a result of low accuracy scores (i.e., less than 70% accuracy whereas the mean was above 93% across all six experiments), one observer was omitted from the analysis of the ensemble condition in Experiment 1, one observer was removed from the analysis of the single-object condition in Experiment 2, one observer was removed from the analysis of both conditions in Experiments 3 and 4, two observers were removed from the analysis of both conditions in Experiment 5, and one observer was removed from the analysis of both conditions in Experiment 6.
Stimuli and apparatus
Stimuli used in this study consisted of images of single objects (150 × 150 pixels for Experiments 1, 3, 4, and 5, 55 × 55 pixels for Experiment 2) and object ensembles (a collection of approximately 60 objects within an image; 378 × 378 pixels in size for Experiments 1, 3, 4, 5, and 6; see Figures 1 through 6). All of the single objects used were also present in all of the corresponding ensemble images. Images in Experiments 1, 2, 3, 4, and 5 were constructed using stone beads made from semiprecious gems and photographed using a desktop studio set up (for more details, see Cant & Xu, 2012, experiment 4, and Cant & Xu, in press). Stimuli were presented in two distinct shapes (a star or a heart for single objects and a collection of stars or hearts for ensembles) and also in two different textures (for single objects: individual surface textures taken from one of two different texture “families” that closely resembled each other; for object ensembles: a heterogeneous mixture of individual surface textures that belonged to one texture “family” and another mixture that belonged to a different, but visually similar, texture “family”). The two texture families were labeled as “texture A” and “texture B” to facilitate responses in the texture-discrimination task in both the single-object and ensemble conditions. For both single objects and ensembles, we used four different exemplars for each shape and texture combination (e.g., four different hearts from texture A, four different stars from texture A, four different hearts from texture B, etc.). The same two shapes and the same two stone textures were used in each experiment (with the exception of Experiments 3 and 4 in which one new texture replaced one of the textures used in Experiments 1, 2, and 5; see Figures 3 and 4). The two shapes were chosen to be distinct to encourage the usage of a local-processing strategy wherein attention to a single object in the ensemble would be sufficient for the observers to successfully perform discriminations of ensemble shape. In contrast, the two types of textures were chosen to be similar to each other to encourage the usage of a global-processing strategy wherein attention to multiple objects in the ensemble would be required for the observers to successfully discriminate one stone texture from the other (with the exception of the textures used in Experiment 5). All stimuli were presented in full color except for those used in Experiments 3 and 4, which were presented in gray scale. Images in Experiment 6 were created using Adobe Photoshop CS6 software (Adobe Systems, San Jose, CA) and MATLAB (Mathworks, Natick, MA). To encourage a global-processing strategy for ensemble shape, 10 unique exemplars of hearts and 10 unique exemplars of stars were created by warping the original heart and star shape contours used in Experiments 1, 2, 3, 4, and 5. Additionally, five unique ambiguous “mixed” shapes were created and added to the ensembles to further create heterogeneity in ensemble shape perception and thus encourage observers to adopt a more global scope of attention when engaged in the ensemble shape discrimination task. The shape exemplars were filled with the same surface texture patterns used in Experiments 1, 2, and 5. Furthermore, in Experiment 6, we matched the hue of the ensembles across textures A and B using Photoshop, and the luminance histograms of the textures were equated using the SHINE toolbox extension for MATLAB (Willenbockel et al., 2010). Luminance histograms were equated in a pair-wise manner in order to hold luminance constant between textures A and B but maintain heterogeneity of luminance within each ensemble image (see Experiment 6 and Figure 6).
Figure 3.

Stimuli and results from Experiment 3 in which images were presented at 10° eccentricity while observers fixated a central cross. Gray scale textures were used to control for differences in color between stimuli. In the baseline blocks (blue bars), only the relevant (i.e., attended) feature varied, and in filtering blocks (red bars), both the relevant and irrelevant (i.e., unattended) features varied. Error bars represent within-subject SEM. *p < 0.05.
Figure 4.

Stimuli and results from Experiment 4 in which stimuli were presented until response (with a limit of 2000 ms). In the baseline blocks (blue bars), only the relevant (i.e., attended) feature varied, and in filtering blocks (red bars), both the relevant and irrelevant (i.e., unattended) features varied. Error bars represent within-subject SEM. *p < 0.05.
Observers sat at a desk in a darkened room with their head mounted in a headrest, and stimuli were presented on an LCD monitor (1920 × 1080 pixels) located directly in front of them. Stimulus presentation and behavioral data collection was controlled by Superlab Pro version 2.0.4 (Cedrus Corporation, Sand Pedro, CA) in Experiments 1, 2, 3, 4, and 5, and MATLAB using the Psychophysics Toolbox extension (Brainard, 1997; Pelli, 1997) in Experiment 6. Stimuli were either presented at central fixation (Experiments 1, 2, 4, 5, and 6) or at 10° eccentricity from central fixation (Experiment 3), and the distance from the observer's eyes to the screen was approximately 40 cm. When classifying the stimuli based on the shapes and textures listed above, observers responded by pressing either the “1” or “3” key on the number pad of the keyboard with their right index or ring finger, respectively. A fixation cross was always present on the computer screen.
Procedure
All observers completed both a single-object stimulus condition and an ensemble stimulus condition in each experiment except for the observers in Experiment 2, who only completed a single-object condition, and the observers in Experiment 6, who only completed an ensemble condition. The order of these stimulus conditions was counterbalanced across observers when they were both included in an experiment. Within each of the single-object or ensemble conditions, there were two feature tasks. In the shape task, observers classified the stimuli based on their outline shape (i.e., star vs. heart for single objects, a collection of all stars vs. all hearts for ensembles); in the single-object texture task, observers classified the stimuli on the basis of their surface texture (i.e., an individual object that belonged to either texture A or texture B). For ensemble texture discriminations, observers were asked to assess whether the collective surface texture of all of the elements comprising an ensemble belonged to the family texture A or the family texture B (there was no mixing of textures A and B elements within a single ensemble image). The order of these tasks was counterbalanced across observers in each experiment. Prior to starting the experiment, observers were shown a display containing examples of the stimuli that they were going to encounter during the testing session.
Before starting each task of each stimulus condition, observers were given 20 practice trials to become familiar with the task. The observer's task upon presentation of the stimulus in any given trial was to classify that stimulus as quickly and accurately as possible. Verbal feedback was provided where necessary. In Experiments 1, 2, and 3, the stimulus was presented for 200 ms. In Experiments 4, 5, and 6, the stimulus was presented until response with a limit of 2000 ms (see Experiment 4 for more detail). Immediately following the response, there was a 2000-ms intertrial interval until the presentation of the next stimulus. Observers were instructed to maintain central fixation throughout the entire practice session (and throughout the entire experiment). For the experiment proper, each stimulus condition (i.e., single object or ensemble) consisted of eight 32-trial blocks with four blocks in each feature task (i.e., shape and texture). Two of these four blocks served as baseline blocks (in which only the relevant feature varied), and the other two blocks served as filtering blocks (in which both the relevant and irrelevant features varied). The order of presentation of the four blocks was counterbalanced across tasks and observers in each experiment. For each block of 32 trials, the two possible responses for the relevant feature (e.g., star or heart in the case of shape) were presented equally often in pseudorandom order, constrained such that a given correct response would repeat no more than three trials in a row. Half of the observers pressed “1” for star and “3” for heart, and the other half of the observers had these button assignments reversed. Thus, the assignment of the response buttons was counterbalanced across observers and tasks in each experiment. An instruction screen separated each block and informed observers that they could take a short break if they desired and reminded them to respond as quickly and accurately as possible in the next block of trials. Each observer completed 512 trials during the entire experimental session (32 trials × 4 blocks × 2 feature tasks × 2 stimulus conditions = 512 trials) except for observers in Experiments 2 and 6, who completed 256 trials (because they only took part in the single-object or ensemble condition, respectively).
Data analysis
Before data were analyzed, two analyses were performed to remove outliers. First, observers who had an overall accuracy of less than 70% in each stimulus condition were excluded from further analyses. As accuracy typically ranged from 93% to 96%, an accuracy below 70% was a good indication that the observer was not properly engaged in the task, making his/her data unreliable and difficult to interpret. Second, for the remainder of observers, following standard practice, response latencies that were 2.5 SD above or below the mean reaction time for each feature task in each stimulus condition were excluded.
Separate analyses of variance (ANOVAs) were conducted for the single-object and the ensemble stimulus conditions. Response latencies (for correct trials only) and the number of errors committed were analyzed in both cases using a 2 × 2 repeated-measures ANOVA with α = 0.05. Throughout this study, we focus more on the analysis of response latencies because, in our experiments, response latency was a much more sensitive measure than error rate as the error rate tended to be low and variable. Main effects of interest included feature task (shape and texture) and interference condition (baseline and filtering blocks). Pair-wise post hoc comparisons were also performed using α = 0.05.
Experiment 1: Shape and texture interference in single object and object-ensemble perception
Observers encountered two stimulus conditions in this experiment: a single-object condition and an ensemble condition (see General methods). In both conditions, observers classified stimuli based on differences in their outline shape while ignoring changes in texture and vice versa.
We recorded observers' response latencies and accuracy in each task but placed more emphasis on the analysis of response latencies (see Data analysis). In the single-object condition, we predicted independence in the processing of shape and texture (this prediction applies to all subsequent experiments containing a single-object condition). Specifically, we predicted that response latencies in the baseline and filtering blocks would not differ significantly from each other in both the shape and the texture feature tasks. This would replicate the results of Cant et al. (2008), which demonstrated independent processing of shape and texture in single-object perception. In the ensemble condition, based on previous fMRI results (Cant & Xu, 2011) we predicted the presence of interference between ensemble shape and texture processing when ensembles were processed globally in the texture task (i.e., response latencies significantly faster in the baseline blocks, in which only texture varied, relative to the filtering blocks, in which both texture and shape varied) but reduced or no interference between ensemble shape and texture processing when ensembles were processed locally in the shape task.
Results and discussion
Single-object condition
Mean accuracy for the single-object condition across all participants was high, averaging 95%. There were no differences between the baseline and filtering blocks in either task (ts < 0.50, ps > 0.62). Additionally, there were no significant results for the main effect of feature task (shape vs. texture task), F(1, 15) = 0.03, p = 0.87, partial η2 = 0.002; the main effect of interference condition (filtering vs. baseline condition), F(1, 15) = 0.02, p = 0.88, partial η2 = 0.002; or the feature task-by-interference condition interaction, F(1, 15) = 0.25, p = 0.63, partial η2 = 0.02 (see Table 1 for a list of the mean number of errors committed in each experiment).
Table 1.
Mean number of errors committed in each experiment.
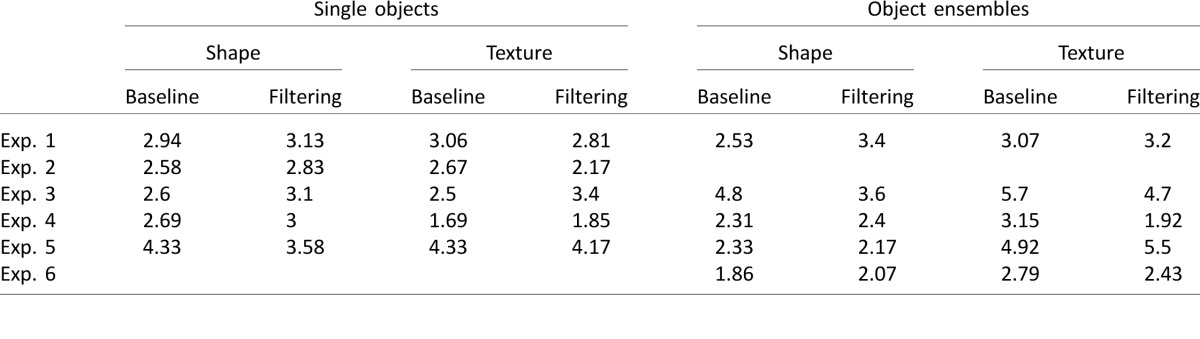
In response latency analyses, there were no differences in response latency between the baseline and filtering blocks in either the shape task, t(15) = 0.38, p = 0.71, or the texture task, t(15) = 0.06, p = 0.96 (see Figure 1). The main effect of feature task (shape vs. texture) was significant, F(1, 15) = 12.63, p = 0.003, partial η2 = 0.48, showing that observers were faster in the shape than in the texture task. Meanwhile, both the main effect of interference condition (baseline vs. filtering), F(1, 15) = 0.03, p = 0.87, partial η2 = 0.002, and the interaction between feature task and interference condition were not significant, F(1, 15) = 0.06, p = 0.80, partial η2 = 0.004. These results thus revealed independent processing of shape and texture in single-object perception.
Overall, these results demonstrated that the shape and texture of single objects could be processed independently. This is consistent with previous behavioral (Cant et al., 2008) and neuroimaging (Cant & Goodale, 2011) studies that together suggest the existence of separate cognitive and neural mechanisms for the processing of shape and texture in single-object perception. Previous studies have investigated shape processing by examining differences in aspect ratio (i.e., differences in the width or length of rectangles) whereas in the present study shape was investigated by examining differences in outline contour (i.e., a star outline or heart outline). Thus the independence between shape and texture processing generalizes across at least two dimensions of shape.
Ensemble condition
Participants performed very well in the ensemble condition, averaging an accuracy of 95%. In the error analysis, there were no significant differences across the baseline and filtering blocks in either task (ts < 1.53, ps > 0.14). Additionally, there were no significant results for the main effect of feature task, F(1, 14) = 0.04, p = 0.84, partial η2 = 0.003; the main effect of interference condition, F(1, 14) = 0.77, p = 0.40, partial η2 = 0.05; or the task-by-interference condition interaction, F(1, 14) = 0.88, p = 0.37, partial η2 = 0.06. (See Table 1 for a list of the mean number of errors committed in each experiment.)
In response latency analyses, we found a significant difference between the baseline and filtering blocks in the texture task, t(14) = 2.23, p = 0.04 (see Figure 1), indicating shape interference in ensemble texture perception. Although we observed a trend of increased reaction times in the filtering compared with the baseline blocks in the shape task, this comparison was not significant, t(14) = 1.46, p = 0.17. However, the main effect of interference condition was significant, F(1, 14) = 10.58, p = 0.006, partial η2 = 0.43, and it did not interact with task, F(1, 14) = 0.07, p = 0.80, partial η2 = 0.005. Additionally, the main effect of feature task was not significant, F(1, 14) = 1.12, p = 0.31, partial η2 = 0.07, indicating that the two tasks were well matched in overall difficulty. Thus, there is an overall interference effect between shape and texture processing. This is different from what we obtained in the single-object condition. Supporting this observation, the two-way interaction between interference condition and stimulus condition (single object vs. ensemble) was significant, F(1, 14) = 9.52, p = 0.008, partial η2 = 0.41. The three-way interaction among stimulus condition, feature task, and interference condition was not significant, F(1, 14) = 0.08, p = 0.78, partial η2 = 0.01, showing that beyond the significant two-way interaction, feature task did not interact further with the other two conditions. Overall these results indicate that shape and texture are processed differently in single objects and in object ensembles, showing independent processing in single-object perception but interference in object-ensemble perception. Additionally, during ensemble perception, interference was present in texture processing, which required a global level of processing, but not in shape processing, which could be accomplished by a local level of processing, providing some support to the notion that manipulating local and global levels of ensemble processing may modulate the amount of interference observed between shape and texture in ensemble perception.
Experiment 2: Controlling for differences in size
In Experiment 1, we observed interference between shape and texture processing in ensemble but not single-object perception. We interpret these data as evidence that shape and texture can be processed independently in single-object perception and are thus mediated by cognitive mechanisms distinct from those involved in ensemble perception. Nevertheless, because the single objects were larger than the objects contained within the ensembles, perhaps interference effects in single object processing are washed out when the size of the stimuli are too large. To test this alternative explanation, in this experiment, observers took part in a version of the single-object task in which the size of the single objects was matched to the size of the objects within the ensemble stimuli presented in the previous experiment. If object size plays an important role, we should see interference between shape and texture processing in single-object processing; however, if size does not matter, then we should still observe independent processing between shape and texture. As the focus of this experiment was on single-object processing, an ensemble task was not conducted.
Results and discussion
Overall, participants performed very well, averaging 96% correct. No significant results were found in the error analysis between the baseline and filtering blocks in both the shape, t(11) = 0.38, p = 0.71, and the texture, t(11) = 0.60, p = 0.56, tasks. Similarly, no significant results were observed for the main effect of task, F(1, 11) = 0.14, p = 0.72, partial η2 = 0.01; the main effect of interference condition, F(1, 11) = 0.05, p = 0.82, partial η2 = 0.01; or the task-by-interference condition interaction, F(1, 11) = 0.52, p = 0.49, partial η2 = 0.05.
For response latencies, there were no significant differences between the baseline and filtering blocks in either task (ts < 0.83, ps > 0.41; see Figure 2). Additionally, there were no significant results in the main effect of task, F(1, 11) = 3.40, p = 0.09, partial η2 = 0.24; the main effect of interference condition, F(1, 11) = 0.46, p = 0.51, partial η2 = 0.04; or the interaction between these two factors, F(1, 11) = 0.89, p = 0.37, partial η2 = 0.07.
Figure 2.

Stimuli and results from Experiment 2 in which the size of the single objects was matched to the size of the objects contained within the ensembles in all other experiments. Observers completed only a single-object task in this experiment (the ensemble stimuli are shown to facilitate a comparison between the size of the single objects and the size of the elements within the ensembles). In the baseline blocks (blue bars), only the relevant (i.e., attended) feature varied, and in filtering blocks (red bars), both the relevant and irrelevant (i.e., unattended) features varied. Error bars represent within-subject SEM.
After controlling for size differences between the elements presented in the single-object and object-ensemble images, we replicated our results from Experiment 1, showing independent processing of shape and texture in single-object perception. Thus, object size cannot account for the absence of shape and texture interference in single object processing. Taken together, the findings thus far, along with the results from previous studies (Cant et al., 2008; Cant & Goodale, 2011) strongly suggest that shape and texture are separable (i.e., independent) dimensions in single-object perception.
Experiment 3: Moving to the periphery
Past research suggests that ensemble processing entails extracting global features from multiple objects, often in the visual periphery (see Alvarez & Oliva, 2008). This predicts that presenting stimuli in the periphery may better promote global processing of ensembles, possibly resulting in interference effects in ensemble shape processing in addition to that seen in ensemble texture processing. To investigate this, in Experiment 3, we moved away from presenting stimuli at the center of the screen and presented both single objects and ensembles at 10° eccentricity while still requiring observers to maintain central fixation. Stimuli appeared randomly on either the left or the right of fixation and were presented for only 200 ms. By performing these manipulations, we ensured that eye movements would not confound our results.
Given that peripheral presentation may further boost global processing of ensembles, we expected to replicate the shape interference effect seen in the ensemble texture task in Experiment 1. For ensemble shape processing, on the one hand, the enhanced global processing of ensembles in the periphery might result in texture interference in the ensemble shape task. On the other hand, because the shape of the elements was still uniform in an ensemble (i.e., all hearts or all stars), observers may be able to covertly attend to a few elements nearest fixation to make their shape discriminations, resulting in a lack of texture interference in the ensemble shape task as was found in Experiment 1.
The two types of textures used thus far had subtle color differences. As such, one could argue that our results are better interpreted as independence or interference between shape and color processing rather than shape and texture processing per se. Although color contributes to texture perception, it would be important to know whether shape and texture interference in ensemble processing still exists after the contribution of color is completely removed. To this end, we gray scaled the stimuli and chose two textures that could reliably be discriminated from each other in the absence of any differences in color (see Figure 3). If observers were using ensemble color rather than texture differences in the texture task in Experiment 1, then controlling for differences in color should eliminate any interference effects. If, however, the interference previously seen in the ensemble texture task was not reliant upon color processing, then controlling for differences in color should have no effect, and we would predict interference in the ensemble texture task, replicating the results of Experiment 1.
Results and discussion
Single-object condition
Mean accuracy for the single-object condition across all participants was high, averaging 95%. No significant results were found in the number of errors committed between the baseline and filtering blocks in both the shape and the texture tasks (ts < 1.38, ps > 0.19). Additionally, there were no significant results for the main effect of task, F(1, 9) = 0.05, p = 0.83, partial η2 = 0.01; the main effect of interference condition, F(1, 9) = 1.72, p = 0.22, partial η2 = 0.16; or the interaction between these two factors, F(1, 9) = 0.44, p = 0.52, partial η2 = 0.05.
Similar to the error analysis, the analysis of response latencies revealed that observers' response latencies in the baseline blocks did not differ from their response latencies in the filtering blocks for both the shape and the texture tasks (ts < 0.73, ps > 0.48; see Figure 3). Additionally, there were nonsignificant main effects of task, F(1, 9) = 0.01, p = 0.93, partial η2 = 0.001, and interference condition, F(1, 9) = 0.69, p = 0.43, partial η2 = 0.07, and a nonsignificant task-by-interference condition interaction, F(1, 9) = 0.22, p = 0.65, partial η2 = 0.02.
Thus, consistent with the results of the first two experiments, these results revealed independence of shape and texture processing in single-object perception. Such lack of interference with single-object stimuli presented in the periphery suggests that either global processing was not effectively engaged with single objects or that global processing of single objects was engaged but is not sufficient to induce interference between shape and texture. We discuss these alternative possibilities in more detail in the General discussion.
Ensemble condition
Mean accuracy for the ensemble condition across all participants was also high, averaging 93%. No significant results were found in the error analysis between the baseline and filtering blocks in either the shape, t(9) = 0.94, p = 0.37, or the texture, t(9) = 0.85, p = 0.42, tasks. Additionally, the main effects of task, F(1, 9) = 0.86, p = 0.38, partial η2 = 0.09; interference condition, F(1, 9) = 1.93, p = 0.20, partial η2 = 0.18; and the interaction between these two factors, F(1, 9) = 0.01, p = 0.92, partial η2 = 0.001, were all nonsignificant.
The analysis of response latencies revealed results consistent with our prediction for texture processing in that we observed significant shape interference in the ensemble texture task as observers were faster at classifying texture in the baseline blocks in which only texture varied, compared with the filtering blocks in which both texture and shape varied, t(9) = 2.59, p = 0.03 (see Figure 3). Although we did observe slightly longer reaction times in the filtering blocks (compared with the baseline blocks) in the ensemble shape task, this trend was not significant, t(9) = 0.75, p = 0.47 (see Figure 3). As a result, it is not surprising that neither the main effect of interference condition, F(1, 9) = 3.03, p = 0.12, partial η2 = 0.25, nor the task-by-interference condition interaction, F(1, 9) = 0.21, p = 0.66, partial η2 = 0.02, were significant. The main effect of task was also not significant, F(1, 9) = 0.07, p = 0.80, partial η2 = 0.01. Finally, the three-way interaction between stimulus condition, task, and interference condition was not significant, F(1, 9) = 1.08, p = 0.33, partial η2 = 0.11, likely because there was a trend of increased reaction times in the filtering compared with the baseline blocks even in the single-object shape task. Nevertheless, across all three experiments, the consistent lack of interference between shape and texture processing in single-object perception and the replication of interference in ensemble texture processing suggests that the perception of single objects and ensembles may be mediated by distinct cognitive mechanisms. Moreover, replicating the effect of interference in ensemble texture processing using gray scale stimuli reveals that our interference effects are not likely explained by appealing to differences in the color of the ensembles.
Moving the ensembles to the periphery led to a replication of the results observed in the ensemble texture task in Experiment 1 but did not increase interference in ensemble shape processing as one would expect given that periphery presentation would presumably promote more global processing of ensembles. We believe this lack of interference can be explained by appealing to a number of different possibilities, all of which relate to the amount of global processing that ensemble shape features received. We discuss these possibilities in detail in the General discussion and make another attempt to induce global processing of ensemble shape in Experiment 6.
Experiment 4: Extending stimulus presentation time
The results of all previous experiments converge on the finding that shape and texture are processed independently in single-object perception. However, for ensemble perception, shape and texture are processed interactively but only when observers attend to texture and not when observers attend to shape. Interestingly, this asymmetric effect holds even when ensembles are presented in the periphery, which should promote global processing of both shape and texture. In Experiment 4, we hypothesized that perhaps the short stimulus presentation time (200 ms) used in previous experiments suppressed interference when observers attended to ensemble shape. Specifically, past research suggests that object shape may be processed earlier in the ventral visual stream (LO; Grill-Spector et al., 1999) than object texture (PPA; Cant & Goodale, 2007, 2011; Cant & Xu, 2012). Thus, with a short presentation time, shape features may receive sufficient processing to interfere with texture, but texture may not yet have received sufficient processing to interfere with shape. To test this idea, in Experiment 4, we presented both single objects and ensembles until response with a limit of 2000 ms. Observers completed both the single-object and ensemble conditions, and all stimuli were presented centrally as in Experiment 1. The stimuli themselves were identical to those in Experiment 3. In line with previous results, we expected to see independence of processing for shape and texture in single-object perception. Moreover, we also expected to replicate interference in ensemble perception when observers attended to texture. Specific to the current experiment, if short stimulus presentation times were indeed suppressing interference when observers attended to ensemble shape, then increasing the presentation times may lead to interference in ensemble perception when observers attend to shape.
Results and discussion
Single-object condition
Overall, participants performed very well in the single-object condition, averaging 96% correct. Error analysis revealed no differences between the baseline and filtering blocks in both the shape and texture tasks (both ts < 0.78, both ps > 0.44). Additionally, there were no significant effects for the main effect of task, F(1, 12) = 2.99, p = 0.11, partial η2 = 0.20; the main effect of interference condition, F(1, 12) = 0.70, p = 0.42, partial η2 = 0.06; and the task-by-interference condition interaction, F(1, 12) = 0.08, p = 0.78, partial η2 = 0.01.
Similar to the results of all previous experiments, the response latency analysis revealed no interference between the processing of shape and texture in single-object perception (see Figure 4). That is, there was no difference in the speed with which observers classified the relevant feature in the baseline blocks, compared with the filtering blocks, in both the shape and the texture tasks (ts < 0.99, ps > 0.34). Moreover, there were no significant results for the main effect of task, F(1, 12) = 0.16, p = 0.70, partial η2 = 0.01; the main effect of interference condition, F(1, 12) = 0.01, p = 0.94, partial η2 = 0.001; and the task-by-interference condition interaction, F(1, 12) = 0.82, p = 0.38, partial η2 = 0.06. The results from Experiment 4 again demonstrate independence in the processing of shape and texture in single-object perception.
Ensemble condition
Participants performed very well in the ensemble condition, averaging 96% correct across all trials. Pair-wise comparisons revealed no significant differences in the error analysis between the baseline and filtering blocks in both the shape, t(12) = 0.14, p = 0.90, and texture, t(12) = 1.92, p = 0.08, tasks. Moreover, neither the main effects of task and interference condition nor the interaction between the two was significant: task, F(1, 12) = 0.03, p = 0.86, partial η2 = 0.003; interference condition, F(1, 12) = 1.51, p = 0.24, partial η2 = 0.11; interaction, F(1, 12) = 2.88, p = 0.12, partial η2 = 0.19.
In the response-latency analysis, consistent with the results in Experiments 1 and 3, pair-wise comparisons revealed no interference in the shape task (i.e., no difference in response latency between the baseline and filtering blocks), t(12) = 0.89, p = 0.39, but significant interference in the texture task (i.e., observers were faster in the baseline blocks in which only texture varied compared to the filtering blocks in which both texture and shape varied), t(12) = 2.25, p = 0.04 (see Figure 4). Although the main effects of task, F(1, 12) = 0.73, p = 0.41, partial η2 = 0.06, and interference condition, F(1, 12) = 0.46, p = 0.51, partial η2 = 0.04, were not significant, the interaction between these two factors was marginally significant, F(1, 12) = 4.04, p = 0.067, partial η2 = 0.25, suggesting that interference differed between the texture and shape tasks. Finally, response latencies in the baseline and filtering blocks for each visual feature differed across the stimulus types used in this experiment (three-way interaction between stimulus condition, task, and interference condition): F(1, 12) = 7.95, p = 0.02, partial η2 = 0.40, likely driven by the interference effect seen in the ensemble texture task but not the other tasks.
In contrast to previous experiments in which stimuli were presented for short durations (200 ms), here we presented stimuli until response (with a limit of 2000 ms) to investigate if stimulus presentation time could explain the asymmetric interference between ensemble shape and texture observed in Experiments 1 and 3. Despite this change, we replicated our previous results (Experiments 1 and 3) of interference between shape and texture in ensemble perception when observers attended to texture. Specifically, observers could not ignore changes in shape while discriminating the texture of the ensemble stimuli. As in Experiment 3, stimuli were presented in gray scale, which again demonstrates that the interference we observe is based upon attention to ensemble texture features and is not due to attending to differences in the color of the ensembles.
We also replicated the finding in Experiments 1 and 3 of no interference in the ensemble shape condition in that observers were able to ignore changes in texture while attending to ensemble shape. Thus even with extended stimulus presentation time, the interference between shape and texture in ensemble perception remains asymmetric. This suggests that this asymmetric interference is not explained by the possibility that ensemble shape is simply processed faster than ensemble texture and thus has more of an opportunity to interfere with processing when texture is the attended feature (and, using the same logic, that ensemble texture does not have an opportunity to interfere with processing when shape is the attended feature because shape features would be processed before texture processing has a chance to produce interference). Further support for this idea is given by the observation that the main effect of feature task (i.e., shape vs. texture) for response time was not significant in Experiments 1, 3, and 4, demonstrating roughly equivalent processing times for shape and texture in ensemble perception across three independent experiments.
Instead, we believe that our repeated replications of interference when attending to ensemble texture but not ensemble shape may be explained by ensemble feature homogeneity and heterogeneity as this visual cue may be critical to engaging local or global processing, thereby generating independent processing or interference, respectively. Specifically, in all previous experiments (except Experiment 2), ensembles were comprised of objects containing the same shape (i.e., homogeneous: all hearts or all stars) but had subtle variations in texture (i.e., heterogeneous: individual objects belonged to the same texture family but were nonetheless distinct from each other). Thus, the processing of a single element (i.e., a local processing bias) of the ensemble may be sufficient for classifying shape, but diffusely distributing attention over many elements (i.e., a global processing bias) would be optimal for classifying texture. In other words, the scope of attention (i.e., local vs. global) may significantly impact interference in ensemble shape and texture perception, such that global processing would lead to greater levels of interference compared with local processing. This is consistent with the results from Experiments 1, 3, and 4, in which ensemble texture was processed more globally and shape more locally. To provide further support for this idea, in Experiment 5, we presented ensembles with homogeneity in both texture and shape, and in Experiment 6, we presented ensembles with heterogeneity (slight variations) in both shape and texture, predicting a lack of interference in both ensemble shape and texture perception in Experiment 5 but interference in both features in Experiment 6. In both experiments, we again kept the ensemble images on the screen until response (up to a maximum of 2000 ms) to further verify that stimulus presentation time was not responsible for the asymmetric interference observed between ensemble shape and texture in Experiments 1, 3, and 4.
Experiment 5: The effect of texture redundancy on ensemble perception
In Experiments 1, 3, and 4, the interference between ensemble shape and texture processing was present in the texture task but absent in the shape task. We argued that this could be due to the deployment of different processing strategies in the two tasks. Specifically, a global processing strategy would be deployed during texture perception as objects belonging to the same texture family (i.e., texture A or texture B) varied slightly in surface appearance (i.e., ensemble texture was heterogeneous), and thus, attending multiple objects in the ensemble would aid in successful discrimination of the two types of textures. Meanwhile, the deployment of a local processing strategy would be sufficient for successful discrimination of the two shapes as all objects in the ensemble had the exact same shape (i.e., ensemble shape was homogeneous). If this account is correct, then, if we allow all objects in an ensemble to have the exact same texture pattern, just like the shapes of each object, then a global processing strategy would no longer be beneficial in ensemble texture perception. This would, in turn, eliminate the interference between shape and texture processing in the texture task. In Experiment 5, we carried out this manipulation by creating new ensembles that were made out of multiple repetitions of the same element, thereby making the texture of the ensembles redundant and homogeneous (see Figure 5). With this manipulation, observers could easily classify texture by using a local processing strategy in which attention is focused on the single element at the center of fixation. As in Experiments 1, 3, and 4, observers performed both a texture and a shape discrimination task on the ensembles and on single objects (see General methods).
Results and discussion
Single-object condition
Mean accuracy for the single-object condition across all participants was high, averaging 94%. In error analyses, pair-wise comparisons revealed no difference between the baseline and filtering blocks in either task (both ts < 1.22, both ps > 0.25), and the main effects of task, F(1, 11) = 0.06, p = 0.81, partial η2 = 0.01; interference condition, F(1, 11) = 0.63, p = 0.44, partial η2 = 0.06; and the interaction between these two factors, F(1, 11) = 0.20, p = 0.66, partial η2 = 0.02, were all nonsignificant.
In response latency analyses, pair-wise comparisons revealed no difference between the baseline and filtering blocks in the shape task, t(11) = 0.84, p = 0.42 (see Figure 5) but a significantly faster response in the filtering than in the baseline blocks in the texture task, t(11) = 2.83, p = 0.02. Consistent with these results, the main effect of task approached significance, F(1, 11) = 4.80, p = 0.051, partial η2 = 0.30; the main effect of interference condition was not significant, F(1, 11) = 2.88, p = 0.12, partial η2 = 0.21; and the interaction between the two factors did reach significance, F(1, 11) = 5.73, p = 0.04, partial η2 = 0.34. The faster response in the filtering blocks in the texture task, although not indicative of interference, represents an unexpected facilitation effect that was not observed in the single-object condition of any of the four previous experiments.
The lack of interference between shape and texture processing once again showed that the shape and texture of single objects were processed independently. The unexpected finding of observers being significantly faster at classifying texture in the filtering compared with the baseline blocks was puzzling as the same stimuli were used in Experiments 1 and 2. Given that this facilitation effect was not observed in four out of five experiments in the present study that included a single-object condition or in any experiment in previously reported studies (e.g., Cant et al., 2008; Cant & Goodale, 2009), it is reasonable to assume that this effect is not valid and thus should not be interpreted as statistically reliable.
Ensemble condition
Mean accuracy for the ensemble condition across all participants was also high, averaging 94%. In error analyses, pair-wise comparisons revealed no difference between the baseline and the filtering blocks in either task (both ts < 0.75, both ps > 0.46). The main effect of task was significant, F(1, 11) = 7.03, p = 0.02, partial η2 = 0.39, but both the main effect of interference condition, F(1, 11) = 0.21, p = 0.66, partial η2 = 0.02, and the interaction between these two factors, F(1, 11) = 0.32, p = 0.59, partial η2 = 0.03, did not reach significance.
In the analysis of response latencies, just as we observed in the single-object condition, there were no significant interference effects in either the ensemble shape, t(11) = 0.19, p = 0.85, or texture, t(11) = 1.29, p = 0.22, tasks (see Figure 5). The main effect of task, F(1, 11) = 0.45, p = 0.51, partial η2 = 0.04; interference condition, F(1, 11) = 0.90, p = 0.36, partial η2 = 0.08; and the interaction between these two factors were also not significant, F(1, 11) = 0.35, p = 0.57, partial η2 = 0.03. Thus, unlike in Experiments 1, 3, and 4, by allowing all objects in an ensemble to have the exact same texture pattern and encouraging the adoption of a local processing strategy, there was no longer interference between shape and texture processing in the texture task, similar to the results obtained from the shape task.
Finally, the patterns of response latencies in the baseline and filtering blocks for task across stimulus condition (i.e., single objects vs. ensembles) approached significance (three-way interaction between stimulus condition, task, and interference condition), F(1, 11) = 4.27, p = 0.063, partial η2 = 0.28, as did the patterns for interference condition across the different types of stimuli (two-way interaction between stimulus condition and interference condition), F(1, 11) = 3.41, p = 0.092, partial η2 = 0.24, both of which were likely driven by the facilitation effect for texture processing in single-object perception. As discussed previously, this facilitation effect was not replicated in any of the previous experiments and therefore likely lacks statistical reliability and should not be interpreted as reflecting true processing differences between shape and texture in single objects.
Taken together, the results of the first five experiments support the notion that the visual processing strategy utilized in ensemble perception can modulate the interference, or lack thereof, between ensemble shape and texture processing in a Garner paradigm. Specifically, greater global processing tended to produce such interference, and greater local processing tended to remove it, and this effect was observed independent of stimulus presentation time (i.e., whether ensembles were presented for 200 ms or until observer response). Moreover, the consistent lack of interference between shape and texture in single-object perception (all five experiments) and the replications of interference in ensemble texture perception (Experiments 1, 3, and 4) suggest that the processing of single objects and object ensembles may be mediated by distinct cognitive mechanisms (see General discussion for a detailed discussion of this).
Experiment 6: Promoting global processing of shape in ensemble perception
In Experiments 1, 3, and 4, we utilized ensembles comprised of the exact same shapes but slight variations in surface texture. By encouraging global processing of texture and local processing of shape, we observed interference when observers attended to texture but not when observers attended to shape. In Experiment 5, we encouraged a local processing strategy for texture by repeating the exact same surface texture across all elements comprising an ensemble and found that the interference effect was eliminated. Collectively these results suggest that ensemble feature homogeneity may encourage a local processing strategy that leads to a lack of interference between ensemble shape and texture whereas ensemble feature heterogeneity encourages a global processing strategy that produces interference. To provide additional support for this idea, in Experiment 6, we explored whether introducing slight variations in the shape of the elements comprising the ensemble would promote global processing of shape and would thus lead to an interference effect. To this end, we presented ensemble images with heterogeneity in both shape and texture, encouraging a global processing strategy for both features (see Figure 6). If processing the attended ensemble feature in a more global manner leads to interference, then we expect to observe interference effects for both features in Experiment 6 (i.e., symmetric interference).
Furthermore, in all experiments thus far, we have not controlled for differences in luminance across the two types of textures. Although Experiments 3 and 4 controlled for color by presenting gray scale images, it could still be the case that the interference effects we have observed thus far represent interference between shape and luminance rather than shape and texture per se. To address this issue, in Experiment 6, prior to constructing ensemble images, we paired exemplar surface textures across textures A and B and separately equated the luminance histograms for each pair using the SHINE toolbox for MATLAB (Willenbockel et al., 2010). In this way, the mean luminance of ensembles across textures A and B were equal, but the luminance of elements comprising each ensemble image remained heterogeneous. A single-object task was not conducted in this experiment.
Results and discussion
Overall, observers performed well on the ensemble task, averaging 96% correct. In the error analysis, the differences between the baseline and filtering conditions were not significant in both the shape and texture tasks (ts < 1, ps > 0.37). Additionally, we found no significant main effect of task, F(1, 13) = 2.76, p = 0.12, partial η2 = 0.18; no significant main effect of interference condition, F(1, 13) = 0.04, p = 0.85, partial η2 = 0.003; and no interaction between these factors, F(1, 13) = 1.25, p = 0.28, partial η2 = 0.09.
For response latencies, pair-wise comparisons revealed that responses were faster in the baseline condition relative to the filtering condition both when observers attended to shape, t(13) = 2.83, p = 0.01, and when observers attended to texture, t(13) = 2.60, p = 0.02. Consistent with this finding of symmetric interference between ensemble shape and texture, the main effect of interference condition was significant, F(1, 13) = 14.44, p = 0.002, partial η2 = 0.53, and not surprisingly, the interaction between feature task and interference condition was not significant, F(1, 13) = 0.23, p = 0.64, partial η2 = 0.02. Finally, the main effect of task was significant, F(1, 13) = 9.47, p = 0.009, partial η2 = 0.42, revealing that response latencies when attending to shape were faster than when attending to texture.
Consistent with our prediction, when observers were encouraged to adopt a global processing strategy for both ensemble features, we observed interference effects regardless of which feature observers attended to (i.e., symmetric interference between shape and texture). Thus, the data from all six experiments suggest that global processing of an ensemble feature leads to interference whereas local processing abolishes this effect. Importantly, the data from this last experiment reveals that this seems to hold true whether the attended feature is shape or texture and is not specific to situations in which texture is the attended feature. Finally, given that we observed these effects after controlling for differences in luminance across the two types of textures, we can conclude that the results reported here indeed reflect interference between shape and surface texture rather than color or luminance.
General discussion
Across six behavioral experiments, we provide evidence for independent processing of shape and texture in single-object perception but not in object-ensemble perception. These findings are not explained by appealing to differences in the color, size, and luminance of the visual stimuli and replicate after varying their location within the visual field (center vs. periphery). Our findings suggest that interference between shape and texture arises in ensemble perception when the ensemble is processed more globally (i.e., ensemble texture task in Experiments 1, 3, 4, and 6; ensemble shape task in Experiment 6) whereas these same features are processed independently when the ensemble is processed using a local processing strategy, similar to how single objects are perceived (i.e., ensemble shape task in Experiments 1, 3, 4, and 5, and ensemble texture task in Experiment 5). The ensemble interference effect demonstrated in Experiment 1 was independently replicated in different participants in Experiments 3, 4, and 6, showing that the findings of ensemble interference are quite reliable. Moreover, our recent neuroimaging findings that ensemble shape and texture are both processed within the same brain region (anterior-medial ventral visual cortex, overlapping with PPA; Cant & Xu, 2011) is both consistent with and supports the behavioral findings of interference in ensemble processing in the present series of experiments. Taken together, these findings demonstrate that perception of the visual features of single objects and object ensembles may be mediated by distinct cognitive mechanisms.
The perception of single objects
Previous research has demonstrated that the shape and the texture of single objects can be processed independently and thus can be considered separable feature dimensions in single-object perception (Cant et al., 2008). Our findings replicate these results but expand upon them by demonstrating that these features can be processed independently not only when shape is defined as differences in the aspect ratio of rectangles (i.e., Cant et al., 2008) but also when shape is defined as differences in outline contour (i.e., star vs. heart). It is quite likely that both of these aspects of shape are represented in LO, a mid- to high-level object shape processing region (Grill-Spector et al., 1999). Whether both types of shape are equally represented in LO or are processed by similar or different subpopulations of neurons is a question for future research.
Recent neuroimaging results have demonstrated that a third aspect of object shape, surface curvature, is also represented in LO (Cant & Goodale, 2011). The perception of surface curvature is often achieved by processing diagnostic surface features, such as specular highlights, shading gradients, and texture gradients (Fleming, Dror, & Adelson, 2003). Interestingly, when surface features, such as texture, contribute more directly to shape perception, shape and texture are not processed independently (Cant & Goodale, 2009). This contrasts with situations in which surface features contribute more directly to material–property perception, where shape and texture are processed independently (Cant et al., 2008). To further investigate this dichotomy, Cant and Goodale (2011) manipulated observers' attention to different features of the same visual surfaces and found that attending to surface shape (i.e., curvature) via textural cues activates LO whereas attending to material properties (i.e., compliance judgments of hard vs. soft surfaces) via textural cues activates a region of parahippocampal cortex that overlaps with the scene-sensitive PPA. Taken together, these behavioral and neuroimaging results demonstrate that there are different networks of processing for surface properties in visual cortex. Specifically, extracting shape information from surface features activates lateral regions of the visual system whereas extracting material properties from surface features activates more medial and anterior regions. Behavioral independence of shape and texture processing in single-object perception is therefore dictated by the type of information that is extracted from a visual analysis of surface texture (material from texture: independent; shape from texture: not independent).
In Experiment 3, we explored whether presenting stimuli in the periphery promotes greater levels of global processing and if this increased global processing, in turn, leads to interference between shape and texture processing. We did indeed observe interference in ensemble texture perception (see “The perception of object ensembles” below for an account of why this interference was not likely a result of greater levels of global processing for peripheral ensembles) but not in single-object perception. Why then, was interference not observed when single objects were presented in the periphery? One possibility is that the size of the single objects did not promote a global processing strategy at 10° eccentricity. Perhaps interference would be more effectively induced if the single objects were reduced in size so they matched the elements within the ensembles and were then presented at 10° eccentricity (but reducing the size of the single objects on its own does not lead to interference as we observed in Experiment 2). Alternatively, perhaps presenting single objects further in the periphery (without altering their size) would better encourage global processing and would lead to interference between shape and texture. Or perhaps we would still not observe interference in single-object processing with these manipulations, which would provide further evidence that the cognitive mechanisms underlying single-object and object-ensemble processing are quite distinct. Future investigations should focus on whether global processing alone leads to interference in feature-based processing or whether instead it is an interaction between the type of processing (i.e., global) and the type of stimulus (i.e., ensembles) that leads to interference.
Finally, although we argue our results provide evidence that distinct cognitive mechanisms are involved in the processing of single objects and object ensembles, could it be that the processing of single objects and ensembles may not be distinct in that the processing of the latter stimulus class may simply be an extension of the former, in which perceptual pooling of multiple visual features follows a stage of processing features from individual objects? In this situation, interference may arise when the pooling of multiple visual features is necessary but not when the same visual features are processed and contained within a single bounded object. If this were indeed the case, then the processing of single objects and ensembles would likely be mediated by shared, rather than distinct, cognitive mechanisms. We believe this is unlikely. Behavioral investigations have demonstrated that observers can extract summary statistical information from ensembles of objects, such as their mean size, direction of motion, speed, orientation, and center location, without being able to provide fine details about any individual object in the ensemble (e.g., Williams & Sekuler, 1984; Watamaniuk & Duchon, 1992; Ariely, 2001; Parkes et al., 2001; Chong & Treisman, 2003; Alvarez & Oliva, 2008). Thus, single object processing does not seem to be capable of supporting ensemble processing. Moreover, if ensemble processing is to play an important and adaptive role to complement and guide object-specific processing to allow the visual system to overcome the capacity limitation inherent in object-based attention, then it cannot simply be an extension of single-object processing. Lastly, our recent neuroimaging results reveal that separate neural regions are involved in the processing of single objects and object ensembles (single objects in LO and ensembles in PPA; Cant & Goodale, 2011; Cant & Xu, 2012). This collection of evidence, together with results from the present investigation, supports the notion that single object and ensemble processing are mediated by different cognitive mechanisms.
The perception of object ensembles
In contrast to the findings in single-object perception, we found that shape and texture are not processed independently in object-ensemble perception. Specifically, we found that if ensembles were processed more globally, observers could not ignore changes in shape when attending to ensemble texture (Experiments 1, 3, 4, and 6), and similarly, could not ignore changes in texture when attending to ensemble shape (Experiment 6). In contrast, processing ensemble features using more of a local processing strategy eliminates interference effects (Experiments 1, 3, 4, and 5: ensemble shape; Experiment 5: ensemble texture). These findings demonstrate that interference effects can be explained by the processing strategy directed at the attended feature. These findings not only hold important implications for future investigations of Garner interference but also make an important contribution to the study of human visual perception. Namely, our results demonstrate that the perception of single objects and object ensembles may be mediated by distinct cognitive mechanisms and that the scope of visual attention can shape the underlying cognitive mechanisms mediating object-ensemble perception.
We do not believe that the saliency of ensemble shape differences (as the unattended feature) accounts for the interference effects observed in ensemble texture processing. The differences in ensemble shape were quite salient and never changed across the first five experiments that contained an ensemble task, yet we observed interference in the texture condition in some experiments (Experiments 1, 3, and 4) but not in others (Experiment 5). We again observed interference in the texture condition in Experiment 6 despite using differences in shape (the unattended feature) that were more subtle than those used in previous experiments. Moreover, we observed interference in the shape condition in Experiment 6 despite using the same subtle surface texture differences (the unattended feature) that did not produce interference in Experiment 1.
We also do not believe that stimulus presentation time was the key variable in explaining ensemble interference because the presence or absence of interference replicated across experiments containing variations in stimulus presentation time (Experiments 1, 2, and 3: 200 ms; Experiments 4, 5, and 6: until response).
In experiments in which ensembles were comprised of the same exact shape or texture, it is quite likely that observers were using a local processing strategy to discriminate differences in ensemble features simply because the single element at fixation was entirely predictive of the correct response in either task. In other words, global processing was not necessary when redundancy in the attended feature was high, and this may explain the lack of interference. In Experiment 3, the ensembles were presented in the periphery while observers fixated centrally. Despite this encouragement of the global processing of ensemble features, we still failed to obtain interference in ensemble shape processing. It is unlikely that observers relied on a single object to make shape discriminations in this experiment because at 10° eccentricity the elements closest to the fovea would be prone to perceptual crowding effects. But it is possible that due to the uniform and homogeneous nature of ensemble shape (i.e., all hearts or all stars), observers may have been able to extract shape information from a few crowded elements that were closest to fixation, a strategy that would indeed invoke some level of ensemble processing but may not have been indicative of efficient global processing.
Thus, in Experiment 6, we constructed ensembles that had more subtle within-family variations in shape (i.e., several distinct star exemplars within the star family and several distinct heart exemplars within the heart family) that made the shape discriminations more perceptually similar to the texture discriminations used in previous experiments (i.e., both shape and texture now contained heterogeneity within a family and subtle, but distinct, differences across families). We also used the subtle texture variations that produced interference in Experiment 1 when texture was the attended feature, and we reasoned that using these stimuli would promote global processing of both shape and texture. Unlike previous experiments, we did indeed observe interference when shape was the attended feature. We also replicated the interference effect (for the third time) when texture was the attended feature. Together these results suggest that using heterogeneous ensemble elements promotes a global processing strategy for both shape and texture, which leads to interference between these ensemble features. The fact that we found symmetric interference between shape and texture in Experiment 6 demonstrates that these ensemble interference effects are not idiosyncratic to situations in which only texture is the attended dimension.
Moreover, our interference effects (both symmetric and asymmetric) are not likely explained by differences in the difficulty of ensemble shape and texture discriminations. Indeed, in Experiments 1, 3, and 4, in which we observed interference in ensemble texture processing, the main effect of feature task (shape vs. texture) for accuracy and reaction time were not significant, demonstrating that the ensemble shape and texture tasks were matched in difficulty. In fact, the only experiment in which we observed a significant main effect of feature task for accuracy was Experiment 5 (accuracy for texture was slightly lower than accuracy for shape, but texture accuracy was still near ceiling at around 95%), which is the only experiment in which we did not observe any ensemble interference effects. In contrast, in Experiment 6, when there was indeed a difference in reaction time such that ensemble shape processing was faster than ensemble texture processing, we still found equally strong interference effects in both types of ensemble feature processing.
Instead, this interference (and the instances in which interference was abolished) may speak to a more general global and local processing mechanism that applies to how we perceive at least two ensemble features (shape and texture), and how attention can be used to vary the meaningful perceptual unit of a visual stimulus (a single object vs. an entire array of multiple objects). Future research can extend these findings by investigating whether global processing of other ensemble features (e.g., motion and orientation) produces symmetric interference, which will provide further details on the cognitive mechanisms underlying object-ensemble perception.
Finally, the finding that shape and texture are not processed independently in ensemble perception is entirely consistent with our recent fMRI results that anterior-medial ventral visual cortex (overlapping with PPA) is sensitive to processing changes in both the shape and the texture of the elements that make up an ensemble (Cant & Xu, 2011). Indeed, the findings from our previous fMRI study partially motivated the present series of behavioral experiments.
Conclusions
Our findings provide evidence that feature-based processing may differ in single-object perception compared with object-ensemble perception. Specifically, in single-object perception shape and texture are separable dimensions (i.e., processed independently), but in object-ensemble perception, they are integral dimensions (i.e., not processed independently). This suggests that there may be distinct cognitive mechanisms for these two types of perceptual processes. Additionally, our results show that even within object ensembles, the scope of visual attention plays a critical role in determining which type of object processing (single object or ensemble) is engaged by the visual system, such that local processing leads to independence and global processing leads to interactions between ensemble features. Together, these results reveal how visual attention can shape the underlying cognitive mechanisms mediating object-ensemble perception.
Supplementary Material
{kind=link}
Acknowledgments
This research was supported by grants from the National Science Foundation (0855112) and the National Institutes of Health (1R01EY022355) to Y. X., an Ontario Graduate Scholarship to S. Z. S., and a Natural Sciences and Engineering Research Council Postdoctoral Fellowship and Discovery Grant (435647-2013) to J. S. C.
Commercial relationships: none.
Corresponding author: Jonathan S. Cant.
Contributor Information
Jonathan S. Cant, jonathan.cant@utoronto.ca, http://www.utsc.utoronto.ca/~jcant/.
Sol Z. Sun, Email: sol.sun@mail.utoronto.ca.
Yaoda Xu, yaodaxu@wjh.harvard.edu, http://visionlab.harvard.edu/Members/Yaoda/Yaoda_Xu.html.
References
- Adelson, E. H. (2001). On seeing stuff: The perception of materials by humans and machines. Rogowitz B. E., Pappas T. N. (Eds.) Proceedings of the SPIE, 4299, Human Vision and Electronic Imaging, VI (pp. 1–12). San Jose, CA: SPIE. [Google Scholar]
- Adelson, E. H. (2008). Image statistics and surface perception. In B. E. Rogowitz, & T. N. Pappas (Eds.), Proceedings of the SPIE, 6806, Human Vision and Electronic Imaging, XIII (pp. 680602-1–680602-9). San Jose, CA: SPIE. [Google Scholar]
- Alvarez G. A.,, Cavanagh P. (2004). The capacity of visual short-term memory is set both by visual information load and by number of objects. Psychological Science, 15, 106–111. [DOI] [PubMed] [Google Scholar]
- Alvarez G. A.,, Oliva A. (2008). The representation of simple ensemble visual features outside the focus of attention. Psychological Science, 19, 392–398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ariely D. (2001). Seeing sets: Representation by statistical properties. Psychological Science, 12, 157–162. [DOI] [PubMed] [Google Scholar]
- Biederman I. (1987). Recognition-by-components: A theory of human image understanding. Psychological Review, 94, 115–147. [DOI] [PubMed] [Google Scholar]
- Biederman I.,, Cooper E. E. (1991). Priming contour-deleted images: Evidence for intermediate representations in visual object recognition. Cognitive Psychology, 23, 393–419. [DOI] [PubMed] [Google Scholar]
- Brainard D. H. (1997). The Psychophysics Toolbox. Spatial Vision, 10, 433–436. [PubMed] [Google Scholar]
- Cant J. S.,, Arnott S. R.,, Goodale M. A. (2009). fMR-adaptation reveals separate processing regions for the perception of form and texture in the human ventral stream. Experimental Brain Research, 192, 391–405. [DOI] [PubMed] [Google Scholar]
- Cant J. S.,, Goodale M. A. (2007). Attention to form or surface properties modulates different regions of human occipitotemporal cortex. Cerebral Cortex, 17, 713–731. [DOI] [PubMed] [Google Scholar]
- Cant J. S.,, Goodale M. A. (2009). Asymmetric interference between the perception of shape and the perception of surface properties. Journal of Vision, 9 (5): 12 1–20, doi:10.1167/9.5.13. [PubMed] [Article] [DOI] [PubMed] [Google Scholar]
- Cant J. S.,, Goodale M. A. (2011). Scratching beneath the surface: New insights into the functional properties of the lateral occipital area and parahippocampal place area. The Journal of Neuroscience, 31, 8248–8258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cant J. S.,, Large M. E.,, McCall L.,, Goodale M. A. (2008). Independent processing of form, colour, and texture in object perception. Perception, 37, 57–78. [DOI] [PubMed] [Google Scholar]
- Cant J. S.,, Xu Y. (2011). Object ensemble coding is distinct from texture processing in the parahippocampal place area. Journal of Vision, 11 (11): 877, doi:10.1167/11.11.877. [Abstract] [Google Scholar]
- Cant J. S.,, Xu Y. (2012). Object ensemble processing in human anterior-medial ventral visual cortex. The Journal of Neuroscience, 32, 7685–7700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cant J. S.,, Xu Y. (in press). The impact of density and ratio on object-ensemble representation in human anterior-medial ventral visual vortex. Cerebral Cortex, doi:10.1093/cercor/bhu145. [DOI] [PMC free article] [PubMed]
- Cavina-Pratesi C.,, Kentridge R. W.,, Heywood C. A.,, Milner A. D. (2010a). Separate channels for processing form, texture, and color: Evidence from fMRI adaptation and visual object agnosia. Cerebral Cortex, 20, 2319–2332. [DOI] [PubMed] [Google Scholar]
- Cavina-Pratesi C.,, Kentridge R. W.,, Heywood C. A.,, Milner A. D. (2010b). Separate processing of texture and form in the ventral stream: Evidence from fMRI and visual agnosia. Cerebral Cortex, 20, 433–446. [DOI] [PubMed] [Google Scholar]
- Chong S. C.,, Treisman A. (2003). Representation of statistical properties. Vision Research, 43, 393–404. [DOI] [PubMed] [Google Scholar]
- Dick M.,, Hochstein S. (1988). Interactions in the dimensions and absolute judgments of orientation and length. Perception, 17, 177–189. [DOI] [PubMed] [Google Scholar]
- Dykes J. R.,, Cooper R. G. (1978). An investigation of the perceptual basis of redundancy gain and orthogonal interference for integral dimensions. Perception & Psychophysics, 23, 36–42. [DOI] [PubMed] [Google Scholar]
- Epstein R.,, Kanwisher N. (April 9, 1998). A cortical representation of the local visual environment. Nature, 392, 598–601. [DOI] [PubMed] [Google Scholar]
- Felfoldy G. L. (1974). Repetition effects in choice reaction time to multidimensional stimuli. Perception & Psychophysics, 15, 453–459. [Google Scholar]
- Fleming R. W.,, Dror R. O.,, Adelson E. H. (2003). Real-world illumination and the perception of surface reflectance properties. Journal of Vision, 3 (5): 12, 347–368, doi:10.1167/3.5.3. [PubMed] [Article] [DOI] [PubMed] [Google Scholar]
- Ganel T.,, Goodale M. A. (Dec. 11, 2003). Visual control of action but not perception requires analytical processing of object shape. Nature, 426, 664–667. [DOI] [PubMed] [Google Scholar]
- Garner W. R. (1974). The processing of information and structure. Potomac, MD: Lawrence Erlbaum Associates. [Google Scholar]
- Grill-Spector K.,, Kushnir T.,, Edelman S.,, Avidan G.,, Itzchak Y.,, Malach R. (1999). Differential processing of objects under various viewing conditions in the human lateral occipital complex. Neuron, 24, 187–203. [DOI] [PubMed] [Google Scholar]
- Grill-Spector K.,, Kushnir T.,, Hendler T.,, Edelman S.,, Itzchak Y.,, Malach R. (1998). A sequence of object-processing stages revealed by fMRI in the human occipital lobe. Human Brain Mapping, 6, 316–328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- James T. W.,, Culham J.,, Humphrey G. K.,, Milner A. D.,, Goodale M. A. (2003). Ventral occipital lesions impair object recognition but not object-directed grasping: An fMRI study. Brain, 126, 2463–2475. [DOI] [PubMed] [Google Scholar]
- Kourtzi Z.,, Kanwisher N. (2000). Cortical regions involved in processing object shape. The Journal of Neuroscience, 20 (9), 3310–3318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luck S. J.,, Vogel E. K. (1997). The capacity of visual working memory for features and conjunctions. Nature, 390, 279–281. [DOI] [PubMed] [Google Scholar]
- Marr D.,, Nishihara H. K. (1978). Representation and recognition of the spatial organization of three-dimensional shapes. Proceeding of the Royal Society B, Biological Sciences, 200 (1140), 269–294. [DOI] [PubMed] [Google Scholar]
- Parkes L.,, Lund J.,, Angelucci A.,, Solomon J. A.,, Morgan M. (2001). Compulsory averaging of crowded orientation signals in human vision. Nature Neuroscience, 4, 739–744. [DOI] [PubMed] [Google Scholar]
- Pelli D. G. (1997). The VideoToolbox software for visual psychophysics: Transforming numbers into movies. Spatial Vision, 10, 437–442. [PubMed] [Google Scholar]
- Peuskens H.,, Claeys K. G.,, Todd J. T.,, Norman J. F.,, Hecke P. V.,, Orban G. A. (2004). Attention to 3-D shape, 3-D motion, and texture in 3-D structure from motion displays. Journal of Cognitive Neuroscience, 16, 665–682. [DOI] [PubMed] [Google Scholar]
- Portilla J.,, Simoncelli E. P. (2000). A parametric texture model based on joint statistics of complex wavelet coefficients. International Journal of Computer Vision, 40, 49–71. [Google Scholar]
- Pylyshyn Z. W.,, Storm R. W. (1988). Tracking multiple independent targets: Evidence for a parallel tracking mechanism. Spatial Vision, 3, 179–197. [DOI] [PubMed] [Google Scholar]
- Rossion B.,, Pourtois G. (2004). Revisiting Snodgrass and Vanderwart's object pictorial set: The role of surface detail in basic-level object recognition. Perception, 33, 217–236. [DOI] [PubMed] [Google Scholar]
- Shechter S.,, Hochstein S. (1992). Asymmetric interactions in the processing of the visual dimensions of position, width, and contrast bar stimuli. Perception, 21, 297–312. [DOI] [PubMed] [Google Scholar]
- Sweeny T. D.,, Haroz S.,, Whitney D. (2012). Perceiving group behavior: Sensitive ensemble coding mechanisms for biological motion of human crowds. Journal of Experimental Psychology: Human Perception and Performance, 39 (2), 329–337. [DOI] [PubMed] [Google Scholar]
- Watamaniuk S. N.,, Duchon A. (1992). The human visual system averages speed information. Vision Research, 32, 931–941. [DOI] [PubMed] [Google Scholar]
- Willenbockel V.,, Sadr J.,, Fiset D.,, Horne G. O.,, Gosselin F.,, Tanaka J. W. (2010). Controlling low-level image properties: The SHINE toolbox. Behavior Research Methods, 42 (3), 671–684. [DOI] [PubMed] [Google Scholar]
- Williams D. W.,, Sekuler R. (1984). Coherent global motion percepts from stochastic local motions. Vision Research, 24, 55–62. [DOI] [PubMed] [Google Scholar]
- Wilson H. R.,, Wilkinson F. (1998). Detection of global structure in glass patterns: Implications for form vision. Vision Research, 38, 2933–2947. [DOI] [PubMed] [Google Scholar]
- Xu Y. (2002). Limitations in object-based feature encoding in visual short-term memory. Journal of Experimental Psychology: Human Perception and Performance, 28, 458–468. [DOI] [PubMed] [Google Scholar]
- Yamanashi Leib A.,, Fischer J. T.,, Liu Y.,, Qiu S.,, Robertson L.,, Whitney D. (2014). Ensemble crowd perception: A viewpoint-invariant mechanism to represent average crowd identity. Journal of Vision, 14 (8): 12 1–13, doi:10.1167/14.8.26. [PubMed] [Article] [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.