

Abstract
Background
New strategies to combat the global scourge of schistosomiasis may be revealed by increased understanding of the mechanisms by which the obligate snail host can resist the schistosome parasite. However, few molecular markers linked to resistance have been identified and characterized in snails.
Methodology/Principal Findings
Here we test six independent genetic loci for their influence on resistance to Schistosoma mansoni strain PR1 in the 13-16-R1 strain of the snail Biomphalaria glabrata. We first identify a genomic region, RADres, showing the highest differentiation between susceptible and resistant inbred lines among 1611 informative restriction-site associated DNA (RAD) markers, and show that it significantly influences resistance in an independent set of 439 outbred snails. The additive effect of each RADres resistance allele is 2-fold, similar to that of the previously identified resistance gene sod1. The data fit a model in which both loci contribute independently and additively to resistance, such that the odds of infection in homozygotes for the resistance alleles at both loci (13% infected) is 16-fold lower than the odds of infection in snails without any resistance alleles (70% infected). Genome-wide linkage disequilibrium is high, with both sod1 and RADres residing on haplotype blocks >2Mb, and with other markers in each block also showing significant effects on resistance; thus the causal genes within these blocks remain to be demonstrated. Other candidate loci had no effect on resistance, including the Guadeloupe Resistance Complex and three genes (aif, infPhox, and prx1) with immunological roles and expression patterns tied to resistance, which must therefore be trans-regulated.
Conclusions/Significance
The loci RADres and sod1 both have strong effects on resistance to S. mansoni. Future approaches to control schistosomiasis may benefit from further efforts to characterize and harness this natural genetic variation.
Author Summary
Aquatic snails transmit schistosome blood flukes, causing a parasitic disease second only to malaria in its global health impact. The mechanisms by which some snails naturally resist infection are poorly understood, but if characterized could enable protocols to interfere with transmission of the disease. Here we identify a region of the snail genome that correlates with resistance to infection, and we examine its effect on immunity jointly with a previously described resistance gene. Variation at other candidate genes has no effect on parasite resistance in this laboratory population. The two resistance regions could serve as targets to block parasite transmission via snails.
Introduction
Approximately one-sixth of the global burden of infectious disease in humans is due to parasites transmitted by invertebrate hosts [1]. A major avenue to combat these diseases lies in understanding the genetic and biochemical basis for natural variation in host resistance [2],[3]. The identification of resistance genes will facilitate genetic manipulation or marker-assisted selective breeding for resistance [4]. An understanding of host-parasite interactions may also suggest molecular therapeutic targets in the parasite. In addition, genes affecting host fitness and host-parasite co-evolution may serve as markers to predict and monitor host population responses to changes in parasite prevalence or virulence.
Although arthropod vectors have been the focus of most work on host resistance genetics, other important hosts include the aquatic snails that carry schistosomes. Measured in terms of healthy years of life lost, schistosomiasis ranks among the most consequential of infectious diseases, costing on the order of 10 million disability-adjusted life years [5]. Specifically, schistosomes infect over 200 million people [6],[7], causing a chronic disease burden that can be lifelong [8],[9]. Furthermore, the disease causes up to 200,000 deaths per year [10]. Treatment is usually based on regular dosing with a single drug, praziquantel [11], which is likely to become less effective as the parasite evolves drug resistance [12],[13]. Resistance of schistosomes to other antihelminthic therapeutics like artemisinin [14] is also possible. Vaccine development is proving elusive [15]. Thus, the development of alternative and complementary strategies, including those targeting the aquatic snails that serve as intermediate hosts, is crucial for controlling this disease.
A major barrier to developing new strategies to interrupt transmission is our limited understanding of the molecular pathways by which snails and parasites interact. Resistance to schistosome infections is highly heritable in snails, and resistance often appears to be due to one or a few major-effect loci [16–19]. However, there are still very few snail loci at which allelic variation is known to associate with resistance. Most genetic work has focused on the New World snail Biomphalaria glabrata. Expression levels of some B. glabrata genes are correlated with resistance [20–23], but it is unknown whether this variation is controlled by cis-regulatory elements, and therefore whether these genes are fundamentally a cause of resistance. The Guadeloupe Resistance Complex (GRC) is a snail genomic region with a strong effect on resistance to Schistosoma mansoni in a Caribbean population of B. glabrata [3], but its importance outside of this population remains to be shown. Alleles of the sod1 gene, encoding superoxide dismutase, affect resistance to S. mansoni in the 13-16-R1 laboratory population of snails, possibly via the role this gene plays in the oxidative burst [24],[25], or perhaps owing to linkage with other immunity genes [26]. A few additional molecular markers are tied to resistance [18],[27], but the functional genes responsible for the phenotype are still unknown. These described loci explain only a minority of phenotypic variation in resistance, suggesting that additional resistance loci remain to be discovered. Not only will such markers represent progress toward the identification of causal resistance genes, they also can provide clues to these genes’ modes of action. That is, even before a functional gene is identified, a linked marker can reveal whether the gene acts dominantly, how it interacts epistatically with other genes, the magnitude of its effect, and which other genes are linked to it.
Strains of mixed geographical origin are ideal for identifying genetic markers with phenotypic effects, because large interpopulation differences will be reflected in the segregating allelic variation, most loci will be polymorphic with intermediate-frequency alleles, and because high linkage disequilibrium (LD) maximizes the power to find markers associated with functional polymorphisms. The B. glabrata strain 13-16-R1 has ancestry in both the Caribbean and Brazil [28–30] and thus shows high sequence diversity and high LD across its genome. Although sod1 is linked to resistance in 13-16-R1 [24],[25], sod1 explains only 4% of phenotypic variation in this lab population, so other genes are likely to be equally or more important. Expression levels at three additional genes (aif, infPhox, and prx1) are correlated with resistance in this population, but their causal roles remain to be demonstrated [23]. Most previous genetic work on this population has targeted candidate genes like these with putative immunological roles [23],[24],[31], but other genes with no prior expectation of immune function could be equally or more important. Here we perform a genome-wide scan to identify other molecular markers tied to resistance. We then test these alongside multiple candidate genes for a correlation with resistance to S. mansoni strain PR1 in the 13-16-R1 strain of B. glabrata.
Methods
Ethics statement
We used mice to maintain the schistosome parasites and to produce miracidia for challenge experiments. Infection is through contact with inoculated water and involves minimal discomfort. Infected rodents are euthanized with CO2 prior to showing clinical signs of disease and are dissected to recover parasitic eggs. Animal numbers were held to the minimum required for the research. The Oregon State University Institutional Animal Care and Use Committee, which adheres to Public Health Service Domestic Assurance for humane care and use of laboratory animals (PHS Animal Welfare Assurance Number A3229-01), approved this research as Animal Care and Use Proposal 4360.
Inbred line RAD-Seq
From 52 inbred lines of B. glabrata 13-16-R1 (described in [23]), we examined 9 lines with low (<25%) susceptibility (“resistant”) and 10 lines with high (>85%) susceptibility (“susceptible”) (Table 1). The identities of these lines were largely, but not perfectly, concordant with the previously designated S and R lines in Larson et al. [23]. We genotyped a randomly chosen snail from each of these lines using RAD-Seq [32] with SbfI on the Illumina HiSeq 2000 at Oregon State University. We mapped reads to the B. glabrata reference genome version BglaB1 [33] with BWA [34] and converted genotypes to vcf format with SamTools [35]. We considered only codominant single-nucleotide polymorphisms (SNPs) (i.e. both alleles seen rather than presence/absence markers defined by null alleles). For each SNP, we counted the observations of the minor (less frequent) allele (heterozygous lines counted as 1, lines homozygous for the minor allele counted as 2; “minor allele count” = MAC), and we calculated the difference in this MAC between the resistant and susceptible lines. This framework is analogous to an FST outlier approach [3],[36], but because of the small sample sizes and high homozygosity of the inbred lines, we found MAC differences to be more straightforward to interpret than FST. We included only SNPs with a MAC in the full dataset of at least 4, since lower MAC values would provide negligible statistical power to detect a meaningful difference between resistant and susceptible lines. We also excluded all SNPs with missing genotypes. For each “RAD site” (200bp region surrounding a RAD cut site, only retained for analysis if possessing a SNP), we identified the SNP showing the largest difference in MAC, and then we compared these SNPs among all RAD sites to find the RAD sites with the largest MAC differences. Scaffolds with RAD sites showing the largest differences in MAC were considered to be candidate resistance markers and examined further. Specifically, we designed PCR primers for two such candidate resistance markers (S1 Table). We Sanger sequenced one of them in representative snails from 51 of the inbred lines, and we genotyped both of them in an independent phenotype-genotype association test (described below). For all pairs of RAD sites, we calculated linkage disequilibrium (LD) as the correlation coefficient, r, and assessed the extent of high-LD regions by examining pairs with high (r > = 0.75) or perfect (r = 1) LD. LD blocks of interest were visualized with Haploview [37].
Table 1. Inbred lines examined with RAD-Seq.
| Line | Susceptibility (%) | RADres1 Genotype | sod1 Genotype |
|---|---|---|---|
| R4 | 0 | EE | BB |
| R31 | 2 | EE | AA |
| R45 | 22.5 | EE | CC |
| R68 | 0 | EE | AA |
| R84 | 0 | EE | BB |
| R95 | 4 | EE | BB |
| R98 | 0 | EE | AA |
| R113 | 2 | EE | BB |
| R156 | 9 | EE | AA |
| S26 | 100 | FF | CC |
| S36 | 100 | FF | AA |
| S46 | 87.5 | EE | AA |
| S73 | 86.5 | EE | AA |
| S74 | 100 | EF | BD |
| S88 | 95 | FF | BB |
| S90 | 100 | FF | CC |
| S93 | 100 | EE | CC |
| S132 | 98 | FF | AA |
| S163 | 95 | FF | CC |
Outbred snail phenotype-genotype association
We also chose eight candidate loci to test for an association with resistance phenotype in outbred snails (S1 and S2 Tables). Although other genes in B. glabrata have also been tied to immune function, this set of eight represented all genes previously found to show immune relevance in this particular 13-16-R1 lab population, as well as all known coding genes at which allelic variation correlates with S. mansoni resistance in any B. glabrata population. There are four sod1 alleles (A, B, C, and D) segregating in this population [26], but because B is the allele correlated with resistance, for most analyses we grouped A, C, and D together as “nB” (not B). For clarity, we assigned each allele at each locus a distinct letter name without repeating letters among the loci examined in this study (Table 2). The loci aif, infPhox, and prx1 were described in Larson et al. [23], but we designed new primers to target these loci (S1 Table). To represent the GRC [3], we used primers targeting Scaffold1_1732kb (B. glabrata reference genome BglaB1; S1 Table), here designating this locus grc. We designed primers to target the gene encoding biomphalysin (here designated bmplys), which is on the same scaffold as sod1 (S1 Table). This locus is interesting because biomphalysin was recently shown to be a schistosome-killing protein [38], and it is located only ~500 Kb from sod1 (B. glabrata reference genome BglaB1), close enough for substantial LD. Thus, it was of interest to ask if the association with resistance was higher for alleles at bmplys than at sod1. We also included the two scaffolds identified in the inbred line RAD-Seq described above (“RADres1” and “RADres2”). Genotypes at all loci were determined with Sanger sequencing or high resolution melt, except for RADres2, at which we observed PCR products of three different sizes (alleles G, H, and I), and confirmed by sequencing of homozygotes that they differ from each other by ≥ 118bp due to copy number variation. We were consequently able to genotype samples at RADres2 by visual inspection of agarose gels after electrophoresis, without sequencing.
Table 2. Loci examined among 439 outbred snails.
| Locus a | Scaffold b | N c | Alleles (% Frequency) d | Other loci in LD e | Infection Odds Impact f | References g |
|---|---|---|---|---|---|---|
| sod1 | 10 | 439 | A (44), B (33), C (21), D (2) | bmplys | 2.2** | [24] |
| RADres1 | 115 | 439 | E (68), F (32) | RADres2 | 1.8** | this study |
| RADres2 | 332 | 439 | G (73), H (16), I (11) | RADres1 | 1.7** | this study |
| bmplys | 10 | 405 | J (63), K (37) | sod1 | 1.9** | [38] |
| aif | 1380 | 439 | L (59), M (25), N (10), O (6) | none | none | [23] |
| infPhox | 174 | 439 | P (69), Q (23), R (6), S (1) | none | none | [23] |
| prx1 | 838 | 439 | T (61), U (36), V (3) | none | none | [23] |
| grc | 1 | 278 | W (38), X (33), Y (29) | none | none | [3] |
aName of gene or marker
bScaffold number in B. glabrata reference genome version BglaB1
cNumber of genotyped and phenotyped outbred snails
dFrequency of each allele, named alphabetically
eOther loci showing significant linkage disequilibrium with each locus
fMultiplicative decrease in odds of infection conveyed by each copy of a resistance allele at each locus, significance (p < 0.01) conveyed with **
gFirst description of locus
We arbitrarily chose 456 outbred 4mm juvenile snails from the 13-16-R1 population, challenged them each with five S. mansoni miracidia (strain PR1), and classified them as infected or not, following methods described in Bonner et al [25]. Using DNA isolated from these phenotyped outbred snails, we amplified the eight candidate loci with PCR and genotyped them with Sanger sequencing, high resolution melt, and/or agarose gel electrophoresis. We tested for LD among all pairs of loci as described above, applying a Bonferroni correction of 300 (number of pairwise comparisons for 25 distinct alleles). We tested for a correlation with resistance using logistic regression (generalized linear model with a binomial response in R), as is standard for infection genetics (e.g. [39]), applying a Bonferroni correction of 25 (number of distinct alleles tested). We treated infection as a binary response variable, and modeled genotype as either additive (0, 1, or 2 alleles) or dominant (0 or 1). Thus, analyses were performed on the odds of infection, rather than the probability [40], although we also report and display probabilities for ease of interpretation. We ran simple logistic regression for all alleles at all eight loci (“single-allele tests”), and we ran multiple logistic regression with two independent variables for all pairs of alleles (“two-allele tests”, either for alleles at the same locus or across loci). We estimated the impact on odds of infection for each allele as the antilog of the β coefficient. We measured the proportion of variation explained (R2) as (Null deviance-Residual deviance)/(Null deviance). In order to test for a dominance effect, we calculated Clopper-Pearson binomial 95% confidence intervals [41] for the proportion of infected snails of each genotype, and tested whether estimates for heterozygotes and homozygotes overlapped. To test for partial dominance, we calculated the expected heterozygous effect for each locus, under additivity, as the mean between the two homozygous genotypes, based on log of odds, and tested whether this fell within the 95% confidence interval for heterozygotes. We tested for epistatic interaction by adding an interaction term to multiple logistic regression models. We further explored epistatic interaction by testing the effect of an allele at one locus in subsets of individuals with fixed genotypes at another, putatively interacting, locus.
Accession numbers
Illumina data have been deposited at NCBI SRA, Bioproject Accession PRJNA270097.
Results
New candidate resistance region from inbred lines
We observed 4304 RAD-Seq SNPs, of which 692 had a MAC under 4 and were discarded, leaving 3612 SNPs on 1611 informative RAD sites on 1259 scaffolds totaling 209Mb. These represented fewer than 1% of the scaffolds in the reference genome assembly, but because of the bias toward large scaffolds, they accounted for 23% of the genome sequence. The largest observed difference in MAC was 13, seen at 11 RAD sites (top 1%), all on different scaffolds (Fig 1). Of these, 10 had SNPs showing identical segregation patterns, with the minor allele absent in all 9 resistant lines, fixed in 6 of the 10 susceptible lines, and heterozygous in a 7th susceptible line. In other words, these 10 RAD sites showed mutually perfect LD (r = 1; p < 10−5) and seemed to comprise a large haplotype block, here designated the RADres genomic region (Table 3 and S1 Fig). The combined size of all ten scaffolds in the RADres region is 2.4Mb. The one remaining RAD site with a MAC difference of 13, on Scaffold7510, was in high but not perfect LD with RADres (r = 0.79, p < 10−4), with the minor allele homozygous in two additional lines, one resistant and one susceptible.
Fig 1. Distribution of allele frequencies among RAD sites.

We characterized 1611 informative RAD sites in 19 inbred lines by minor allele count (“MAC”) (maximum of 19; i.e. 50% allele frequency) and difference in MAC between susceptible (N = 10) and resistant (N = 9) lines (“MAC difference”, theoretical maximum = 18; i.e. 9 resistant lines fixed for one allele, 10 susceptible lines fixed for another). Circle sizes are proportional to the number of RAD sites showing each pattern. The cumulative percentage of RAD sites, starting with the highest observed MAC difference, is shown on the righthand y-axis. The highest MAC difference was observed for 10 RAD sites in perfect mutual LD, with a MAC of 13 and a MAC difference of 13, which we defined as the RADres region and examined further (red arrow; encompasses scaffolds of subsequently examined markers RADres1 and RADres2). The one remaining RAD site with an equivalent MAC difference was also in high, but not perfect, LD with RADres (pink arrow; scaffold not examined further). The sod1 haplotype block had a MAC difference of 5, which was higher than average but not an outlier (blue arrow).
Table 3. Markers in the RADres region.
| Name a | Type b | Scaffold c | Site d |
|---|---|---|---|
| RADres1 | RAD tag SNP | LG24_random_Scaffold115 | 333090 |
| RADres2 | Indel in candidate gene | LG24_random_Scaffold332 | 87890 |
| Unnamed | RAD tag SNP | LG24_random_Scaffold76 | 382528 |
| Unnamed | RAD tag SNP | LGUN_random_Scaffold201 | 71351 |
| Unnamed | RAD tag SNP | LG24_random_Scaffold332 | 292543 |
| Unnamed | RAD tag SNP | LG24_random_Scaffold592 | 188090 |
| Unnamed | RAD tag SNP | LG24_random_Scaffold788 | 80122 |
| Unnamed | RAD tag SNP | LG24_random_Scaffold2400 | 13516 |
| Unnamed | RAD tag SNP | LG24_random_Scaffold2452 | 30139 |
| Unnamed | RAD tag SNP | LG24_random_Scaffold4819 | 41204 |
| Unnamed | RAD tag SNP | LG24_random_Scaffold7794 | 3150 |
aName of marker. Only named markers were assessed in outbred snails
bType of polymorphism
cScaffold in B. glabrata genome, BglaB1 assembly
dSite in B. glabrata genome, BglaB1 assembly
We developed primers to amplify one of the RADres sites, on Scaffold115_333kb (“RADres1”, Tables 2 and 3), and we obtained genotypes from 51 inbred lines. We observed two alleles at RADres1, defined by a single C/T SNP at position 333090 (allele E = T; allele F = C; S2 Table). Allele E was associated with resistance and allele F with susceptibility, with mean susceptibility in EE lines (38%) half that of FF lines (76%) (t-test, p < 0.001; Fig 2). Among 19 resistant lines (<25% susceptibility), 18 were EE homozygotes and only one line with borderline susceptibility (22%) was an FF homozygote. All 15 highly resistant lines (< 20% susceptibility) were EE homozygotes. In contrast, among the 31 homozygous lines with > 25% susceptibility, 14 were EE and 17 were FF (Fisher’s exact test, p < 0.001). After excluding the 19 resistant lines, there was no relationship between genotype and phenotype among the 31 homozygous lines with > 25% susceptibility (mean EE susceptibility = 77%; mean FF susceptibility = 79%; t-test, p > 0.1).
Fig 2. Influence of RADres1 on susceptibility in 50 inbred lines.
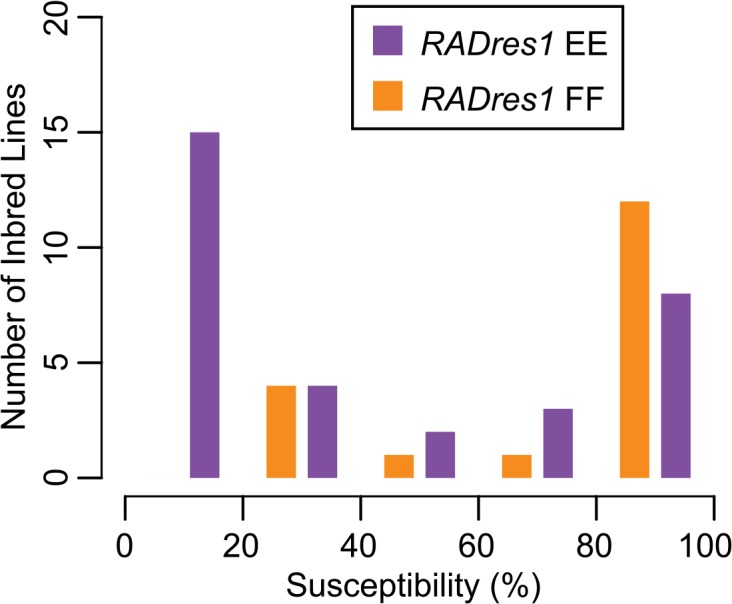
Lines are colored based on genotype (purple = EE, orange = FF) and binned based on susceptibility in windows of 20%. EE lines are more resistant on average. The most striking difference between genotypes occurs in highly resistant lines (susceptibility < 20%), which include no FF lines and 15 EE lines. One additional line (not depicted in figure) was heterozygous with 57% susceptibility.
In addition, we designed PCR primers to amplify a portion of another RADres scaffold, Scaffold332_88kb (“RADres2”), within an intron of a putative glycosyltransferase gene. This gene was of interest because glycosylation could be involved in self vs. non-self recognition. Therefore, we wanted to test whether RADres2 showed a higher association with resistance than our original RAD marker, RADres1. We did not genotype the inbred lines at RADres2, but we used it in our assessment of outbred snails (see below).
Characterization of sod1 region in inbred lines
The sod1 allele that correlates positively with resistance, allele B [24], had a MAC of 11 and a MAC difference of 5, so although it was more prevalent in resistant lines, as expected, it was not an outlier when compared to all RAD sites as was RADres (Fig 1). We observed 80 RAD SNPs at 46 RAD sites on 34 scaffolds showing perfect LD with the sod1 B allele, including a RAD site at Scaffold10_584kb, very near the sod1 position at Scaffold10_612-616kb. The total combined length of these scaffolds was 10.8Mb. Thus, sod1 also resides in a large haplotype block, probably even larger than RADres (S2 Fig).
Linkage disequilibrium analysis
We observed relatively high genome-wide LD in 13-16-R1. Among the 340 RAD site pairs that were on the same scaffold and at least 0.1 Mb apart, 60% showed r > 0.75, and 42% showed perfect LD (r = 1) for at least one SNP. Among all RAD site pairs, including those on separate scaffolds, there were 7757 pairs showing perfect LD for at least one SNP, equivalent to each RAD site occurring in a cluster of 10 mutually identical RAD sites on average. There were 33 haplotype blocks that each contained at least 10 distinct RAD sites showing perfect mutual LD for at least one SNP, encompassing 32% of RAD sites (Fig 3). The single largest haplotype block was the sod1 block described above with 46 RAD sites, and the second-largest block had 34 RAD sites. Thus, multi-scaffold haplotype blocks of high LD extending hundreds of kilobases or more are common in this laboratory population, and the extent of LD observed at RADres is not atypical, although the extent of LD observed at sod1 is.
Fig 3. Extent of linkage disequilibrium (LD) in 13-16-R1.

Among 19 inbred lines, RAD SNPs with identical genotype patterns (LD of 1) were grouped into haplotype blocks. The number of RAD sites represented by each haplotype block is shown. Most (82%) RAD sites show perfect LD with at least one other RAD site, indicating high genome-wide LD. The RADres block shows a typical extent of LD, with 10 RAD sites in perfect mutual LD. RAD sites with SNPs differentiating sod1 B haplotype from the other sod1 haplotypes form the single largest haplotype block in the genome (46 RAD sites). Thus, sod1 appears to be in an unusually large haplotype block of perfect LD.
Outbred snail genotypes
Of the 456 challenged outbred snails, we obtained phenotype and genotype data from 439 (S3 Table), with the remaining failures owing to snail death prior to phenotyping or insufficient extraction of DNA. In all 439 snails we obtained complete genotypes for six loci: sod1, RADres1, RADres2, aif, infPhox, and prx1. We also obtained genotypes for the majority of snails for two additional loci: grc (N = 278), and bmplys (N = 405). All loci were polymorphic with 2–4 alleles, and all had intermediate frequency alleles (25–75% frequency), thus conveying high power to detect both correlations with resistance and linkage disequilibrium among loci (Table 2). All loci were found to be in Hardy-Weinberg equilibrium (p > 0.05) except for aif, which showed a slight deficit of heterozygous genotypes (observe 216, expect 253; p < 0.0001), which could be explained by a rare null allele (~6% frequency) that by chance was never observed as a homozygote.
We observed significant LD between the two pairs of loci for which it was expected (S4 Table). The loci sod1 and bmplys are separated by a physical distance of <500kb on the same scaffold. They are in high but imperfect LD, with the highest LD observed between the sod1 B allele and bmplys (r = 0.91, p < 10−15). There were two bmplys alleles (J and K), with allele K at 37% frequency showing a positive correlation with the resistance allele B at sod1, which occurred at a similar frequency (33%). Almost all (99%) sod1 B alleles co-occurred with bmplys K, and the remaining bmplys K alleles most commonly occurred with sod1 A. Similarly, all three RADres2 alleles showed significant high LD with RADres1, and allele G showed the highest LD with RADres1 (r = 0.71, p < 10−15). Allele G at RADres2 occurred at 71% frequency and was positively correlated with resistance allele E at RADres1, which occurred at a similar frequency (68%). The only other pairwise combination between alleles at different loci showing significant LD was between sod1 D and infPhox S, which was marginally significant after Bonferroni correction (p = 0.04; S4 Table); because these two rare (≤ 2% frequency) alleles co-occur in only three individuals, and because the more common alleles at these loci do not show LD, we dismissed this result as unlikely to be biologically meaningful. No other pairwise combination between alleles at different loci showed significant LD (p > 0.05 for all).
Outbred snail phenotype-genotype association
We observed 174 infected and 265 non-infected outbred snails (40% susceptibility). Our logistic regression analyses repeatedly showed the same main result: two genomic regions, (sod1/bmplys) and RADres, were significantly correlated with resistance, and no other loci were. We describe these results in detail below.
In the single-allele tests, at least one allele of sod1, bmplys, RADres1, and RADres2 showed a significant correlation with resistance (p < 0.05 for each; Fig 4 and S5 Table). For all alleles correlated with resistance at these four loci, an additive model explained more of the phenotypic variation and had a lower Akaike information criterion (AIC) than the dominant model. Comparing within the two pairs of linked loci, sod1 had a non-significantly stronger effect on resistance than bmplys, and RADres1 had a non-significantly stronger effect on resistance than RADres2 (Table 2 and Fig 4). No allele at any of the remaining loci (aif, infPhox, prx1, and grc) showed a significant association with resistance, whether modeled as dominant or additive (p > 0.1 for each; Fig 4). Thus, we found two independent genomic regions correlated with resistance in 13-16-R1, and in both regions the allele most strongly correlated with resistance (sod1 B and RADres1 E) acts additively, reducing the proportion of infected snails by approximately 16% with each additional copy (Fig 5).
Fig 4. Effects of single alleles in outbred snails.

We measured the effects of individual alleles using separate simple logistic regression analyses for each allele at all loci. Each allele is plotted according to its multiplicative effect on the odds of susceptibility (x-axis) and the p-value for its effect (not corrected for multiple tests; y-axis). Standard errors are shown with horizontal lines flanking each point. Alleles are colored based on locus. Only alleles at RADres1, RADres2, sod1, and bmplys are significant (uncorrected threshold of 0.05 indicated by dotted line, threshold after Bonferroni correction of 0.002 indicated by dashed line). Alleles are labeled (A-L, P, W, and T) as in Table 2. For non-significant loci, only the most common allele is shown.
Fig 5. Independent influence of RADres1 and sod1 genotypes on resistance.
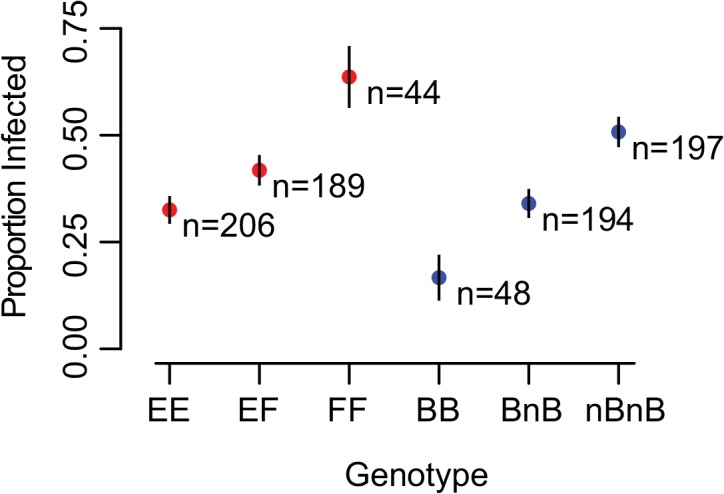
Among 439 snails, both the E allele at RADres1 and the B allele at sod1 are positively correlated with resistance. For each locus (red genotypes = RADres1; blue genotypes = sod1), all three genotypes are significantly different from each other, indicating additive rather than dominant effects. Standard errors of proportions are indicated by vertical bars.
Our results from the single-allele tests were mirrored in the two-allele tests, in that only alleles at sod1, bmplys, RADres1, and RADres2 ever showed a significant correlation with resistance (p < 0.05) (Tables 2 and S6). For the two haplotype blocks with multiple loci and/or alleles (RADres1 and the three alleles of RADres2; bmplys and the four alleles of sod1), we never observed a separate significant effect for a second allele after including the best allele for that block (sod1 B or RADres1 E). Although there is a trend that sod1 D conveys resistance (Fig 4), it has no significant effect either alone or in a model that also includes sod1 B. The single best fitting model included sod1 B and RADres1 E, which were also the two alleles with the strongest individual effects on resistance in their haplotype blocks (Fig 4). A simple additive model, in which alleles B and E contribute to resistance with no dominance and no epistasis, fits the data well (S6 Table and Fig 6), with both terms significant (p < 0.01 for both). This additive model explains 7% of the population variance in resistance, with each sod1 B allele decreasing the odds of infection by 2.2 (95% CI: 1.58–2.99), and each RADres1 E allele decreasing the odds of infection by 1.8 (95% CI: 1.32–2.43). Thus, the odds of infection for a snail with genotype BB/EE (predict 13% infected, odds = 0.15) are 16-fold lower than for a snail without a resistance allele at either locus (predict 70% infected, odds = 2.33). The genotype combinations with the poorest fit to this model (green squares in Fig 6) are BB/FF (predict 33% infected, observe 75% infected, N = 4) and BB/EF (predict 22% infected, observe 13% infected, N = 24). However, predicted values for all genotype combinations fall into the 95% Clopper-Pearson confidence intervals.
Fig 6. Joint influence of RADres1 and sod1 genotypes on resistance.

Genotype combinations are indicated along the x and y axes. Empirical resistance values are plotted as black circles along the z axis. Standard errors of proportions for resistance at each genotype combination are shown with a vertical yellow line. Predicted values from an additive 2-locus multiple regression model with no dominance or epistasis are indicated with green squares. The data are consistent with this simple model, although minor non-additive effects may be responsible for small nonsignificant differences between predicted and empirical values.
We tested for non-additive effects with more complex models, beginning with a test for dominance. If there were complete dominance, we would expect heterozygotes to show an identical impact on resistance as one of the homozygous genotypes. If there were partial dominance, the effects of heterozygotes might be intermediate between both homozygous genotypes, but more similar to one homozygous genotype than the other. For both RADres1 and sod1, all three genotypes are significantly different from each other with respect to proportion of infected snails (Clopper-Pearson 95% confidence intervals do not encompass the proportions of other genotypes; Fig 5). Specifically, heterozygotes are intermediate in resistance; they are significantly more resistant than susceptibility allele homozygotes (FF or nBnB) and significantly less resistant than resistance allele homozygotes (EE or BB). Thus, for both loci we can reject the hypothesis of complete dominance. Observed heterozygous effects were not significantly different than the mean between the homozygous effects (encompassed in Clopper-Pearson 95% confidence intervals), indicating no evidence for partial dominance. Thus, we observe purely additive effects at both loci, rather than complete or partial dominance.
We then tested for epistatic interactions between loci. In a model with an additive effect of both RADres1 and sod1, plus an interaction term, the interaction term was not significant (p > 0.1). To further explore the possibility of an interaction, we tested for an additive effect of each locus in the subset of individuals with each possible genotype at the other locus (Fig 6). The estimated reductions of odds of resistance of the sod1 B allele in specified RADres1 genetic backgrounds were as follows: FF background 1.0-fold (95% CI: 0.39–2.77), EF background 2.5-fold (95% CI: 1.58–4.10), EE background 2.2-fold (95% CI: 1.35–3.66), E- (EE or EF) background 2.4-fold (95% CI: 1.68–3.33). Thus, there is no significant effect of sod1 in a FF background, but this is not significantly lower than the effect of sod1 in an E- background (t-test, p > 0.1). The estimated reductions of odds of resistance of the RADres1 E allele in a specified sod1 genetic background were as follows: nBnB background 1.6-fold (95% CI: 1.06–2.53), BnB background 1.8-fold (95% CI: 1.12–2.78), BB background 4.4-fold (95% CI: 1.06–17.99), B- (BB or BnB) background 1.9-fold (95% CI: 1.23–2.92). Thus, RADres1 has a significant effect in all sod1 genetic backgrounds, with an effect in the BB background estimated to be higher, but not significantly so, than in the other genetic backgrounds. In summary, our data are consistent with an additive model (Fig 6), with no significant evidence for epistatic effects, although we may have low power to detect certain epistatic effects, especially those driven by BB/FF dual homozygotes of which we observed only four individuals.
Discussion
We have identified a genomic region in B. glabrata, RADres, at which each copy of a resistance allele cuts the odds of schistosome infection in half. The impact of this locus is of similar magnitude to the only other known resistance locus in the 13-16-R1 snail strain, sod1, which also conveys an approximately 2-fold effect with each copy of a resistance allele. Thus, combined homozygosity at both loci confers a 16-fold change in the odds of infection (from approximately 2:1 odds to approximately 1:8 odds). Other candidate genes show no association with resistance, including the GRC [3] and three loci with constitutive expression patterns correlated with resistance in this population [23].
The RADres genomic region has not been previously implicated as relevant to snail immunity. Because the RADres region extends over several megabases, and likely includes many scaffolds that were not represented in our RAD-Seq data, the identities of candidate causal genes remain speculative. RADres1 occurs on a scaffold that includes putative genes inferred to encode a methyltransferase, an ATP-dependent zinc metalloprotease, and an F-box/LRR-repeat protein. RADres2 occurs in a putative glycosyltransferase gene, which could have an immune function due to the importance of glycoproteins in snail-trematode interactions [42]. The same scaffold also includes putative genes encoding an autophagy-related protein, a serine carboxypeptidase, and a single-pass transmembrane protein with no sequence similarity to other known proteins. One of the most studied immunity gene families in B. glabrata, the FREPs [43], does not occur among the known RADres scaffolds. Even without identifying the causal gene, we can make inferences about its probable function from the fact that RADres apparently acts additively. As a caveat, we acknowledge that the causal locus could act dominantly, with our observation of additivity being due to imperfect linkage disequilibrium between it and our markers (for example, RADres1 EE homozygotes could be more resistant merely because they have two chances to share a haplotype with a rare dominant causal allele that occurs with only some E alleles). However, both our observation of high genome-wide LD and the admixed history of this laboratory population make this scenario unlikely. Thus, RADres seems to have a quantitative rather than qualitative effect, as resistance scales with the amount of its protein product, with homozygotes being more resistant than heterozygotes. RADres therefore represents a potentially rate-limiting step in the immune response, rather than a step with full functionality at low concentrations such as a recognition molecule that launches a signaling cascade after a single match to its target. If RADres encodes an immune effector like sod1 does, the success of an immune response may depend on the quantity of this protein. Alternatively, the probability that an immune response is mounted at all may scale with the concentration of a signaling molecule produced by RADres, given the importance of signaling in snail defense [44]. In addition, two lines of evidence suggest that RADres contains a fundamental component of the immune system upon which other immune components depend. First, all inbred lines showing high resistance (< 20% susceptibility) are homozygous for the RADres resistance allele. Second, although there is no significant effect of RADres1 genetic background on the effect of sod1, it is striking that the sod1 B allele is not correlated with resistance among RADres1 FF homozygotes (Fig 6). Because we observed only 4 BB/FF dual homozygotes, we have little power to test the role of sod1 in this genetic background. However, the trend is that RADres1 FF homozygotes are highly susceptible regardless of sod1 genotype, suggesting that allele RADres1 E may be required for sod1 to function as an immunity gene. Thus, our data are consistent with the hypothesis that this population includes highly resistant genotypes (i.e. would nearly always resist infection with PR1 strain S. mansoni after indefinite trials) of which RADres1 E is a necessary, though not sufficient, component.
The sod1 locus was first identified as a candidate gene for S. mansoni resistance due to its crucial role in the oxidative burst [45]. It was subsequently shown that sod1 genotypes correlate with resistance in 13-16-R1 [24] and that the resistance-associated B allele shows higher constitutive expression than the other alleles [46]. In this study we found a 2-fold effect of the B allele, the same magnitude reported by Goodall et al. [24]. Goodall et al. [24] also reported that the C allele is correlated with susceptibility, but they did not tease apart whether C is significantly different than A, or whether the link between C and susceptibility is simply the converse of the B/nB difference. Here, we did not detect an effect of C independent of B, suggesting that C and A are functionally equivalent. Intriguingly, our point estimate of infection odds reduction by sod1 D, after accounting for sod1 B and RADres1 E, was quite high (3.5) but not significant (uncorrected p = 0.06). Our power to assess sod1 D was limited as this allele was rare (2%) and never observed to be homozygous, but it is possible that this allele conveys a resistance impact even greater than sod1 B or RADres1 E. Because sod1 was chosen as an a priori candidate, our working hypothesis is that this gene itself is the causal gene, not merely a linked marker for an unknown causal gene. However, there are several caveats to the putative causality of sod1. First, sod1 resides on the largest high-LD haplotype block in this population, showing perfect LD with markers on many other scaffolds in the 19 inbred lines, suggesting that the region of association with resistance likely extends across many megabases encompassing numerous genes. Second, sod1 is linked to at least two other genes (cat and prx4) that also contribute to the oxidative burst, both of which are in LD with sod1 in the 13-16-R1 population [26]. These genes reside on contigs that were not represented by RAD-Seq sites, underscoring the fact that our RAD-Seq data likely underestimate the true size of the sod1 haplotype block. Third, other potentially immune-relevant genes occur on contigs in this haplotype block, notably the bmplys gene encoding biomphalysin, a β pore-forming toxin which can directly kill S. mansoni sporocysts [38]. We have shown that bmplys is in high LD with sod1 and both are therefore strongly correlated with resistance. If there is a genomic cluster of immunity genes, sod1 is not necessarily the one harboring the functional polymorphism. On the other hand, the correlation with resistance is slightly, though not significantly, lower at bmplys than at sod1, suggesting that sod1 may be closer to the causal center of this region of association. The fact that sod1 was not an MAC difference outlier (Fig 1) underscores the limits of our statistical power with only 19 inbred lines and suggests that other loci of similar effect on host resistance may remain undetected. It is also possible that the effect of sod1 is relatively weaker in highly inbred genetic backgrounds because non-additive genetic variance can play a larger role.
Although expression levels of aif, infPhox, and prx1 are all positively correlated with resistance in this population [23], segregating polymorphisms at these loci show no phenotypic association. These three genes were initially targeted because of their potential immunological role. Specifically, aif encodes allograft inflammatory factor, associated with inflammatory response and activation of hemocytes [23]. Likewise, infPhox, encoding a putative phagocytic oxidase subunit of NADPH oxidase, and prx1, encoding peroxiredoxin, both likely affect the concentration of parasite-killing reactive oxygen species [23]. Our results do not invalidate the potential functional importance of the variation in expression at these loci, but show that it must be controlled in a trans, rather than cis, manner. That is, these genes are upregulated in response to a signal that ultimately depends on genetic variation elsewhere in the genome. For example, resistant individuals may have a transcription factor allele that binds more strongly to the motifs at aif, infPhox, and prx1. It is tempting to speculate that RADres in fact contains such a trans-acting variant. The strong additive effect of the E allele at RADres1 could be due to its influence on expression of a large number of immune-relevant genes. Unfortunately, for the small number of inbred lines in which gene expression was assessed, we do not have the statistical power to dissect an association with RADres from the correlated association with resistance. Furthermore, the majority of phenotypic variation in resistance remains unexplained by the combination of RADres and sod1, so it is likely that other loci also play a role. A promising future direction will be to test for an effect of RADres genotype on genome-wide gene expression patterns.
The GRC genomic region is strongly correlated with S. mansoni resistance in B. glabrata from Guadeloupe [3]. Patterns of polymorphism at several genes in this genomic region in the Guadeloupean snails suggest long-term balancing selection, which would predict that functionally divergent alleles occur through the species range. However, we did not detect a correlation between GRC genotype and resistance in 13-16-R1. This negative result may be due in part to the different parasite strain used in this study (PR1) versus in Tennessen et al. [3] (Guadeloupian S. mansoni), since strain-by-strain (G x G) interactions are known to be highly variable in this host-parasite system [47–51]. In addition, GRC diversity may have been lost in the narrow laboratory bottleneck during the founding of 13-16-R1. That is, 13-16-R1 may have retained only functionally similar GRC alleles by chance, even if all founding populations harbored functional genetic diversity. Intriguingly, no locus in mixed-origin 13-16-R1 conveys as strong an effect on resistance as the GRC in the natural Guadeloupe population (8-fold impact on odds of infection, explains 23% of variation in trait, [3]). As snails likely coevolve with trematodes, it is plausible that many variants affecting immunity will become fixed as populations diverge, resulting in many resistance loci in mixed-origin populations like 13-16-R1, each of which determines a small fraction of phenotypic variation. In contrast, fewer loci will be segregating within a single population in the wild, and thus those that do harbor functional variants, like the GRC in Guadeloupe, will explain a large proportion of the phenotypic variation.
This research represents a step forward toward identifying resistance genes and immune mechanisms in snails that could be targeted in the control of global schistosomiasis. Although no one resistance allele is likely to be a “silver bullet” that could unilaterally block all schistosome infection [52], the host-parasite molecular interactions represented by these resistance loci may be exploited in combination with other strategies for disease control. We have identified in B. glabrata a new genomic region, RADres, with a strong influence on resistance to schistosomes. Genes in this region may be suitable for manipulation of immunity in wild snails, but further work is needed to fine-map the causal gene(s) and to determine their other possible phenotypic effects and their immune roles under different genetic backgrounds, parasite strains, or environmental conditions. We have also further characterized the genomic region harboring sod1, notably showing that linkage disequilibrium is particularly high in this region. Therefore, the causal gene could be any of the other linked genes nearby, not necessarily sod1 itself, although because of its immunological role and expression patterns that vary by genotype [45],[46], sod1 is still the best candidate in this genomic region. Finally, we have shown that several candidate genes known to be correlated with resistance either in other populations (GRC) or via expression patterns in this population (aif, infPhox, and prx1) do not contain genetic variants linked to resistance under the conditions of this study. These other loci could still be useful targets in manipulating the immunity of wild snails, but the RADres region may encode a candidate of similar or greater importance for generating the desirable phenotypic outcome of heightened resistance.
Supporting Information
(XLS)
(XLS)
(XLS)
(XLS)
(XLS)
(XLS)
All SNPs on all scaffolds containing a RADres marker are included. Pairwise LD between SNPs is indicated by color; bright red indicates perfect LD. Sc = scaffold.
(PDF)
All SNPs on all scaffolds showing perfect LD with the sod1 B allele are included. Pairwise LD between SNPs is indicated by color; bright red indicates perfect LD. Sc = scaffold.
(PDF)
Acknowledgments
Inbred lines were developed and maintained by Randy Bender and Maureen Larson. Technical assistance was provided by Kelli Lovelace, Laura McMahon and Haley Hutcheson. Coen Adema and Pat Minx facilitated access to earlier versions of the B. glabrata genome assembly.
Data Availability
RAD-Seq Illumina reads have been submitted to SRA (Bioproject Accession PRJNA270097).
Funding Statement
This work was partially supported by National Institutes of Health (NIH; http://www.nih.gov/) grants AI016137 to CJB and AI109134 to MSB. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Townson H, Nathan MB, Zaim M, Guillet P, Manga L, Bos R, et al. Exploiting the potential of vector control for disease prevention. Bull World Health Organ. 2005; 83: 942–947. [PMC free article] [PubMed] [Google Scholar]
- 2. Rottschaefer SM, Riehle MM, Coulibaly B, Sacko M, Niaré O, Morlais I, et al. Exceptional diversity, maintenance of polymorphism, and recent directional selection on the APL1 malaria resistance genes of Anopheles gambiae . PLoS Biol. 2011; 9: e1000600 10.1371/journal.pbio.1000600 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Tennessen JA, Théron A, Marine M, Yeh J-Y, Rognon A, Blouin MS. Hyperdiverse gene cluster in snail host conveys resistance to human schistosome parasites. PLoS Genetics. 2015; 11: e1005067 10.1371/journal.pgen.1005067 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Esvelt KM, Smidler AL, Catteruccia F, Church GM. Concerning RNA-guided gene drives for the alteration of wild populations. eLife. 2014; 3: e03401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. King CH, Sturrock RF, Kariuki HC, Hamburger J. Transmission control for schistosomiasis—why it matters now. Trends Parasitol. 2006; 22: 575–582. [DOI] [PubMed] [Google Scholar]
- 6. King CH, Dickman K, Tisch DJ. Reassessment of the cost of chronic helmintic infection: a meta-analysis of disability-related outcomes in endemic schistosomiasis. Lancet. 2005; 365: 1561–1569. [DOI] [PubMed] [Google Scholar]
- 7. Steinmann P, Keiser J, Bos R, Tanner M, Utzinger J. Schistosomiasis and water resources development: systematic review, meta-analysis, and estimates of people at risk. Lancet Infect Dis. 2006; 6: 411–425. [DOI] [PubMed] [Google Scholar]
- 8. King CH. Parasites and poverty: The case of schistosomiasis. Acta Tropica. 2010; 113: 95–104. 10.1016/j.actatropica.2009.11.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. King CH. Schistosomiasis: challenges and opportunities In Institute of Medicine (US) Forum on Microbial Threats. The Causes and Impacts of Neglected Tropical and Zoonotic Diseases: Opportunities for Integrated Intervention Strategies National Academies Press (US), Washington (DC) 2011. [PubMed] [Google Scholar]
- 10. Thétiot-Laurent SA, Boissier J, Robert A, Meunier B. Schistosomiasis chemotherapy. Angew Chem Int Ed Engl. 2013; 52: 7936–7956. 10.1002/anie.201208390 [DOI] [PubMed] [Google Scholar]
- 11. Doenhoff MJ, Hagan P, Cioli D, Southgate V, Pica-Mattoccia L, Botros S, et al. Praziquantel: its use in control of schistosomiasis in sub-Saharan Africa and current research needs. Parasitology. 2009; 136: 1825–1835. 10.1017/S0031182009000493 [DOI] [PubMed] [Google Scholar]
- 12. Fallon PG, Doenhoff MJ. Drug resistant schistosomiasis: Resistance to praziquantel and oxaminquine induced in Schistosoma mansoni in mice is drug specific. Am J Trop Med Hyg. 1994; 51: 83–88. [DOI] [PubMed] [Google Scholar]
- 13. Melman SD, Steinauer ML, Cunningham C, Kubatko LS, Mwangi IN, Wynn NB, et al. Reduced susceptibility to praziquantel among naturally occurring Kenyan isolates of Schistosoma mansoni . PLoS Negl Trop Dis. 2009; 3: e504 10.1371/journal.pntd.0000504 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Keiser J, Utzinger J. Artemisinins and synthetic trioxolanes in the treatment of helminth infections. Curr Opin Infect Dis. 2007; 20: 605–612. [DOI] [PubMed] [Google Scholar]
- 15. Ricciardi A, Ndao M. Still hope for schistosomiasis vaccine. Hum Vaccin Immunother. 2015; 10.1080/21645515.2015.1059981 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Richards CS, Shade PC. The genetic variation of compatibility in Biomphalaria glabrata and Schistosoma mansoni . J Parasitol. 1987; 73: 1146–1151. [PubMed] [Google Scholar]
- 17. Richards CS, Knight M, Lewis FA. Genetics of Biomphalaria glabrata and its effect on the outcome of Schistosoma mansoni infection. Parasitol Today. 1992; 8: 171–174. [DOI] [PubMed] [Google Scholar]
- 18. Knight M, Miller AN, Patterson CN, Rowe CG, Michaels G, Carr D, et al. The identification of markers segregating with resistance to Schistosoma mansoni infection in the snail Biomphalaria glabrata . Proc Natl Acad Sci USA. 1999; 96: 1510–1515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Webster JP, Gower CM, Blair L. Do hosts and parasites coevolve? Empirical support from the Schistosoma system. Am Nat. 2004; 164: S33–S51. [DOI] [PubMed] [Google Scholar]
- 20. Hanington PC, Forys MA, Dragoo JW, Zhang SM, Adema CM, Loker ES. Role for a somatically diversified lectin in resistance of an invertebrate to parasite infection. Proc Natl Acad Sci USA. 2010; 107: 21087–21092. 10.1073/pnas.1011242107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Hanington PC, Forys MA, Loker ES. A somatically diversified defense factor, FREP3, is a determinant of snail resistance to schistosome infection. PLoS Negl Trop Dis. 2012; 6: e1591 10.1371/journal.pntd.0001591 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Ittiprasert W, Knight M. Reversing the resistance phenotype of the Biomphalaria glabrata snail host Schistosoma mansoni infection by temperature modulation. PLoS Pathog. 2012; 8: e1002677 10.1371/journal.ppat.1002677 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Larson MK, Bender RC, Bayne CJ. Resistance of Biomphalaria glabrata 13-16-R1 snails to Schistosoma mansoni PR1 is a function of haemocyte abundance and constitutive levels of specific transcripts in haemocytes. Int J Parasitol. 2014; 44: 343–353. 10.1016/j.ijpara.2013.11.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Goodall CP, Bender RC, Brooks JK, Bayne CJ. Biomphalaria glabrata cytosolic copper/zinc superoxide dismutase (SOD1) gene: Association of SOD1 alleles with resistance/susceptibility to Schistosoma mansoni . Mol Biochem Parasitol. 2006; 147: 207–210. [DOI] [PubMed] [Google Scholar]
- 25. Bonner KM, Bayne CJ, Larson MK, Blouin MS. Effects of Cu/Zn superoxide dismutase (sod1) genotype and genetic background on growth, reproduction and defense in Biomphalaria glabrata . PLoS Negl Trop Dis. 2012; 6: e1701 10.1371/journal.pntd.0001701 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Blouin MS, Bonner KM, Cooper B, Amarasinghe V, O'Donnell RP, Bayne CJ. Three genes involved in the oxidative burst are closely linked in the genome of the snail, Biomphalaria glabrata . Int J Parasitol. 2013; 43: 51–55. 10.1016/j.ijpara.2012.10.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Ittiprasert W, Miller A, Su XZ, Mu J, Bhusudsawang G, Ukoskit K, et al. Identification and characterisation of functional expressed sequence tags-derived simple sequence repeats (eSSR) markers for genetic linkage mapping of Schistosoma mansoni juvenile resistance and susceptibility loci in Biomphalaria glabrata . Int J Parasitol. 2013; 43: 669–677. 10.1016/j.ijpara.2013.03.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Newton WL. The establishment of a strain of Australorbis glabratus which combines albinism and high susceptibility to infection with Schistosoma mansoni . J Parasitol. 1955; 41: 526–528. [PubMed] [Google Scholar]
- 29. Richards CS, Merritt JW. Genetic factors in susceptibility of juvenile Biomphalaria glabrata to Schistosoma mansoni infection. Am J Trop Med Hyg. 1972; 21: 425–434. [DOI] [PubMed] [Google Scholar]
- 30. DeJong RJ, Morgan JAT, Wilson WD, Al-Jaser MH, Appleton CC, Coulibaly G, et al. Phylogeography of Biomphalaria glabrata and B. pfeifferi, important intermediate hosts of Schistosoma mansoni in the New and Old World tropics. Mol Ecol. 2003; 12: 3041–3056. [DOI] [PubMed] [Google Scholar]
- 31. Bender RC, Broderick EJ, Goodall CP, Bayne CJ. Respiratory burst of Biomphalaria glabrata hemocytes: Schistosoma mansoni-resistant snails produce more extracellular H2O2 than susceptible snails. J Parasitol. 2005; 91: 275–279. [DOI] [PubMed] [Google Scholar]
- 32. Baird NA, Etter PD, Atwood TS, Currey MC, Shiver AL, Lewis ZA et al. Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS ONE. 2008; 3: e3376 10.1371/journal.pone.0003376 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Adema, C. Biomphalaria glabrata genome Assembly BglaB1 on Vectorbase. Available: http://www.vectorbase.org/organisms/biomphalaria-glabrata
- 34. Li H, Durbin R. Fast and accurate short read alignment with Burrows-Wheeler Transform. Bioinformatics. 2009; 25: 1754–1760. 10.1093/bioinformatics/btp324 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, et al. The Sequence alignment/map (SAM) format and SAMtools. Bioinformatics. 2009; 25: 2078–2079. 10.1093/bioinformatics/btp352 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Narum SR, Hess JE. Comparison of FST outlier tests for SNP loci under selection. Mol Ecol Resour. 2011; 11: 184–194. 10.1111/j.1755-0998.2011.02987.x [DOI] [PubMed] [Google Scholar]
- 37. Barrett JC, Fry B, Maller J, Daly MJ. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics. 2005; 21: 263–265. [DOI] [PubMed] [Google Scholar]
- 38. Galinier R, Portela J, Moné Y, Allienne JF, Henri H, Delbecq S, et al. Biomphalysin, a new β pore-forming toxin involved in Biomphalaria glabrata immune defense against Schistosoma mansoni . PLoS Pathogen. 2013; 9: e1003216 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Carr DF, Chaponda M, Jorgensen AL, Castro EC, van Oosterhout JJ, Khoo SH, et al. Association of human leukocyte antigen alleles and nevirapine hypersensitivity in a Malawian HIV-infected population. Clin Infect Dis. 2013; 56: 1330–1339. 10.1093/cid/cit021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. O’Connor AM. Interpretation of odds and risk ratios. J Vet Intern Med. 2013; 27: 600–603. 10.1111/jvim.12057 [DOI] [PubMed] [Google Scholar]
- 41. Clopper C, Pearson ES. The use of confidence or fiducial limits illustrated in the case of the binomial. Biometrika. 1934; 26: 404–413 [Google Scholar]
- 42. Lehr T, Frank S, Natsuka S, Geyer H, Beuerlein K, Doenhoff MJ, et al. N-Glycosylation patterns of hemolymph glycoproteins from Biomphalaria glabrata strains expressing different susceptibility to Schistosoma mansoni infection. Exp Parasitol. 2010; 126: 592–602. 10.1016/j.exppara.2010.06.031 [DOI] [PubMed] [Google Scholar]
- 43. Zhang SM, Loker ES. The FREP gene family in the snail Biomphalaria glabrata: additional members, and evidence consistent with alternative splicing and FREP retrosequences. Dev Comp Immunol. 2003; 27: 175–187. [DOI] [PubMed] [Google Scholar]
- 44. Walker AJ, Lacchini AH, Sealey KL, Mackintosh D, Davies AJ. Spreading by snail (Lymnaea stagnalis) defence cells is regulated through integrated PKC, FAK and Src signalling. Cell Tissue Res. 2010; 341: 131–145. 10.1007/s00441-010-0986-4 [DOI] [PubMed] [Google Scholar]
- 45. Goodall C, Bender R, Broderick E, Bayne C. Constitutive differences in Cu/Zn superoxide dismutase mRNA levels and activity in hemocytes of Biomphalaria glabrata (Mollusca) that are either susceptible or resistant to Schistosoma mansoni (Trematoda). Mol Biochem Parasitol. 2004; 137: 321–328. [DOI] [PubMed] [Google Scholar]
- 46. Bender RC, Goodall CP, Blouin MS, Bayne CJ. Variation in expression of Biomphalaria glabrata SOD1: a potential controlling factor in susceptibility/resistance to Schistosoma mansoni . Dev Comp Immunol. 2007; 31: 874–878. [DOI] [PubMed] [Google Scholar]
- 47. Basch PF. An interpretation of snail-trematode infection rates: specificity based on concordance of compatible phenotypes. Int J Parasitol. 1975; 5: 449–452. [DOI] [PubMed] [Google Scholar]
- 48. Richards CS. Genetic factors in susceptibility of Biomphalaria glabrata for different strains of Schistosoma mansoni . Parasitology 1975; 70: 231–241. [DOI] [PubMed] [Google Scholar]
- 49. Basch PF. Intermediate host specificity in Schistosoma mansoni . Exp Parasitol. 1976; 39: 159–169. [DOI] [PubMed] [Google Scholar]
- 50. Webster JP, Woolhouse MEJ. Selection and strain specificity of compatibility between snail intermediate hosts and their parasitic schistosomes. Evolution. 1998; 52: 1627–1634. [DOI] [PubMed] [Google Scholar]
- 51. Théron A, Rognon A, Gourbal B, Mitta G. Multi-host parasite infectivity and multi-parasite host susceptibility: a new approach of the Biomphalaria glabrata/Schistosoma mansoni compatibility polymorphism. Infect Genet Evol. 2014; 26: 80–88. 10.1016/j.meegid.2014.04.025 [DOI] [PubMed] [Google Scholar]
- 52. Webster JP, Shrivastava J, Johnson PJ, Blair L. Is host-schistosome coevolution going anywhere? BMC Evol Biol. 2007; 7: 91 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(XLS)
(XLS)
(XLS)
(XLS)
(XLS)
(XLS)
All SNPs on all scaffolds containing a RADres marker are included. Pairwise LD between SNPs is indicated by color; bright red indicates perfect LD. Sc = scaffold.
(PDF)
All SNPs on all scaffolds showing perfect LD with the sod1 B allele are included. Pairwise LD between SNPs is indicated by color; bright red indicates perfect LD. Sc = scaffold.
(PDF)
Data Availability Statement
RAD-Seq Illumina reads have been submitted to SRA (Bioproject Accession PRJNA270097).