

Abstract
AIM: To investigate the immunogenicity of candidate DNA vaccine against hepatitis C virus (HCV) delivered by two plasmids expressing HCV envelope protein 1 (E1) and envelope protein 2 (E2) antigens respectively and to study the effect of CpG adjuvant on this candidate vaccine.
METHODS: Recombinant plasmids expressing HCV E1 and E2 antigens respectively were used to simultaneously inoculate mice with or without CpG adjuvant. Antisera were then collected and titers of anti-HCV antibodies were analyzed by ELISA. One month after the last injection, animals were sacrificed to prepare single-cell suspension of splenocytes. These cells were subjected to HCV antigen specific proliferation assays and cytokine secretion assays to evaluate the cellular immune responses of the vaccinated animals.
RESULTS: Antibody responses to HCV E1 and E2 antigens were detected in vaccinated animals. Animals receiving CpG adjuvant had slightly lower titers of anti-HCV antibodies in the sera, while the splenocytes from these animals showed higher HCV-antigen specific proliferation. Analysis of cytokine secretion from the splenocytes was consistent with the above results. While no antigen-specific IL-4 secretion was detected for all vaccinated animals, HCV antigen-specific INF-γ secretion was detected for the splenocytes of vaccinated animals. CpG adjuvant enhanced the secretion of INF-γ but did not change the profile of IL-4 secretion.
CONCLUSION: Vaccination of mice with plasmids encoding HCV E1 and E2 antigens induces humoral and cellular immune responses. CpG adjuvant significantly enhances the cellular immune response.
INTRODUCTION
Hepatitis C virus (HCV) infection is a worldwide health problem[1]. Up to now, no effective medical treatment is available for the majority of HCV infected patients[2,3], while new infections are continuously emerging from blood transfusion, needle sharing, unprotected sex, close contact of HCV infected patient and other unidentified sources[4]. Thus to control the spread of HCV by vaccination becomes an urgent task, especially in developing countries including China, where there is a large infected population.
Various routes were taken to develop a vaccine against HCV infection. Recombinant HCV antigens purified from E.coli, yeast or insect cells could not protect the vaccinee from virus challenge[5], possibly due to the fact that post-translational modification of the virus antigens in mammalian cells is greatly different from that in the bacterial or yeast cells. So the study on vaccine development has been focused on mammalian systems. Although many efforts have been made in vaccine development and some encouraging results have been obtained[5-8], no effective vaccine is available.
The RNA genome of HCV has a high mutation rate, which explains the existence of many genotypes, subtypes and quasispecies[9-10]. Although there is still a possibility to develop a universal vaccine against HCV of all genotypes, the use of local HCV strain in vaccine study is preferred. Our laboratory has been focusing on DNA vaccine development[11] using an HCV strain isolated from a patient in northern China[12], since DNA vaccine is effective in eliciting cellular immune responses[13] which play key roles in viral clearance.
Envelope proteins were the first choice in the development of vaccines against virus infection. HCV envelope protein 2 has become the major target in HCV vaccine development not only because it is the putative major envelope protein[14], but also because E2 could mediate the binding of HCV particle to host cells[15] and is the ligand of the possible HCV receptor CD81 on the host cell surface[16]. It makes E2 an attractive choice for vaccine development as it has been shown to be a major target of immune response in HCV infected patients[17,18]. Previous studies also revealed multiple neutralizing epitopes in E2 proteins[19-22]. Another envelope protein of HCV, E1, is also desirable for vaccine development, since its possible role in maintaining the natural conformation of E2 protein (through E1-E2 complex)[23] and the possibility of its cooperation with E2 in mediating host cell binding and entry[24]. However, low expression levels of both E1 and E2 proteins were observed when they were expressed in a single open reading frame (data not shown). In this study, we expressed HCV E1 and E2 proteins separately and analyzed the immune responses of DNA vaccination delivered by two plasmids encoding HCV E1 and E2 proteins respectively.
Recent studies showed that the immune system responded to CpG motifs by activating potent Th1-like immune responses[23]. There have also been several reports demonstrating that CpG could enhance humoral immune responses elicited by DNA vaccination[24,25], possibly due to indirect effect of enhanced cellular immune responses. The effect of CpG adjuvant on the immune response elicited by the candidate DNA vaccine was also investigated in this study.
MATERIALS AND METHODS
Plasmids, CpG oligodeoxynucleotide (ODN) and cells
For the construction of pSecTagB/sE2, the fragment of HCV E2 (aa. 384-661) was amplified using plasmid pUC18/E[12] as template with following primers: sense, 5’GGCGTTAAGCTTAACACCTACGTG3’ (HindIII site underlined) ; antisense, 5’CAGGAATTCTCACTCTGATCTATC3’ (EcoRI site underlined). Insertion of the PCR product into pSecTagB (Invitrogen, California) resulted in pSecTagB/sE2, in which a secretion signal was provided at the N-terminal of E2 sequence. The recombinant plasmid was sequenced before further experiments.
HCV E1 encoding plasmid pSec-preS1-E1t340 (pSecTagB/sE1) was constructed as previously described[26]. CpG ODN1826 with phosphorothioate backbone was synthesized in Promega (Shanghai) according to the published sequence 5’TCCATGACGTTCCTGACGTT3[27]. The cell line used for transient expression of the recombinant plasmids, BHK-21, was maintained in DMEM supplemented with 50 mL/L FCS and antibiotics under 50 mL/L CO2 in a humidified 37 °C incubator.
Transient expression
Transfection of BHK-21 cells with HCV-encoding plasmid or empty vector using lipofectAMINE (Invitrogen) was done according to the manuals of the manufacturer. Cells were harvested 48 h after transfection, lysed in SDS-PAGE loading buffer, and subjected to SDS-PAGE (10%). The resolved samples were then transferred onto nitrocellulose membrane and probed with polyclonal anti-E1 (Liu et al[28], unpublished data) or polyclonal anti-E2 antisera. The signals were visualized with SuperSignal West Pico stable peroxide solution (Pierce, USA).
DNA immunization and sera preparation
Four groups of 5 female BALB/c mice (6-8 weeks old) purchased from the Shanghai Laboratory Animal Center were injected 3 times in quadriceps muscles: group 1, with 100 μg pSecTagB in 100 μL PBS; group 2, with 50 μg pSecTagB/sS1E1 and 50 μg pSecTagB/sE2 in a total volume of 100 μL PBS; group 3, with 100 μg pSecTagB together with 10 μg CpG in a total volume of 100 μL PBS; group 4, with 50 μg pSecTagB/sS1E1, 50 μg pSecTagB/sE2 and 10 μg CpG in a total volume of 100 μL PBS. Injections were performed at 0, 4 and 8 wk and blood samples were taken at -2, 2, 6 and 10 wk. Mice were bled under anesthesia through the retro-orbital plexus. Blood was incubated at room temperature for 4 h and centrifuged at 2700 r/min at 4 °C for 10 min. Obtained sera were stored at -20 °C.
Anti-HCV antibody analysis
Enzyme-linked immunosorbent assay (ELISA) was used to determine the presence of antibodies against HCV E1 and E2 antigens in serum samples. E. coli-expressed HCV E1 (aa 192 to 315) or HCV E2 (aa 450 to 565[31]) was used to coat 96-well plates (MaxiSorp Surface, Nunc) at the concentration of 1 μg/mL. The antigens were suspended in PBS with 0.2 g/L sodium azide and incubated overnight at 4 °C. After the plates were washed with PBS plus 0.5 g/L Tween 20 (PBS-T) and blocked in blocking buffer (50 g/L fat-free milk powder in PBS-T), twofold serial dilutions of serum samples in blocking buffer were added and incubated for 2 h at 37 °C. After three wash steps with PBS-T, horseradish peroxidase (HRP) conjugated goat-anti-mouse IgG (Dako, Denmark; 1000-fold diluted in blocking buffer) was added and incubated for 1 h at 37 °C. The plate was then developed with substrate buffer (50 mmol/L Na2HPO4, 25 mmol/L citric acid, 75 μg/mL 3,3,5,5-tetramethylbenzidine, 0.15 mL/L H2O2). After 30 min of incubation at room temperature, the reaction was stopped by adding 0.5 mol/L H2SO4, and absorbance was measured at 450 nm on a microplate reader (Model 450, Bio-Rad Laboratories). Antibody titers were calculated as the highest dilution which gave a positive reading. The cutoff value was set as mean absorbency (A) of sera from the control mice vaccinated with non-recombinant plasmid multiplied by 2.
Preparation of splenocytes
Mice were sacrificed by cervical dislocation. Spleens from these mice were ground on metal mesh to prepare single-cell suspension in grinding media (RPMI1640 supplemented with 100 mL/L FCS, 1 mmol/L Sodium pyruvate and 50 μmol/L β-ME). Red blood cells were lysed by incubating the splenocyte preparations with lysis buffer (0.15 mol/L NH4Cl, 10 mmol/L KHCO2, 0.1 mmol/L EDTA, pH7.2-7.4) briefly and then washed with grinding media. The cells were resuspended in a small volume of grinding media and counted in the presence of Trypan blue. Splenocytes were immediately used for further experiments.
Assay for HCV antigen specific splenocyte proliferation
Splenocytes from vaccinated mice or naïve mice were diluted to 4 × 106/mL with grinding media and plated onto 24-well-plates at 2 mL/well. The cells were in vitro stimulated with 500 ng E2 (aa 450 to 565[31]) or mock stimulated for 3 d. The media were collected for cytokine assay. Fresh grinding medium with 0.5 μCi/mL [3H] thymidine (Amersham Pharmacia Biotech) was added for another 24 h. Cells were then washed with PBS and 100 g/L trichloroacetic acid (TCA), incubated with 100 g/L TCA for 10 min at 37 °C. Afterwards, TCA was removed and cells were lysed with lysis buffer containing 0.33 mol/L NaOH and 10 g/L sodium dodecyl sulfate (SDS). [3H] thymidine incorporation in the cell lysates was measured by liquid scintillation counting. Antigen specific proliferation was presented by stimulation index (SI): SI = cpm of the cells stimulated by antigen/cpm of mock stimulated cells. Splenocytes from naïve mice were used to monitor the specificity of this assay.
Analysis of HCV antigen specific cytokine secretion
Media from the in vitro stimulated splenocytes were collected and precleared by centrifugation at 10000 g and 4 °C for 10 min. Concentration of INF-γ and IL-4 in the media was determined by ELISA using the mouse cytokine assay kits (Jingmei Biotech, Shenzhen) according to the protocols provided by the manufacturer. HCV E2 specific cytokine secretion was represented by stimulation index (SI): SI = cytokine secreted upon E2 stimulation/cytokine secreted upon mock stimulation. Splenocytes from naïve mice were used to monitor the specificity of this assay.
Statistical analysis
Statistical analysis was performed with unpaired 2-sided Student t test. Differences with P values < 0.05 were considered significant.
RESULTS
Transient expression of HCV antigens in mammalian cells
Because the expression level was low when E1 and E2 were expressed in one open reading frame (data not shown), we expressed HCV E1 and E2 genes in separate plasmids in this study. To achieve high-level expression and proper post-translational modification of HCV envelope proteins, the pSecTagB vector with an efficient secretion signal of IgG molecules was chosen. Plasmid expressing HCV E2 was constructed as described in Materials and Methods. C-terminal hydrophobic sequence of E2 was truncated to facilitate the secretion of E2 and to obtain complex-type glycosylation modification, which was presented on the surface of HCV particles[29]. Secreted E2 protein was shown to have better antigenicity, possibly due to its proper modification by Golgi enzymes[30]. Plasmid pSecTagB/sE1 was taken as another component of the candidate DNA vaccine since high-level expression of E1 with this plasmid was observed previously in transiently and stably transfected NIH3T3 cells[26]. Before vaccination experiment, BHK-21 cells were transfected with pSecTagB/sE1 and pSecTagB/sE2, respectively, to check if they would properly express the target HCV proteins in this cell line. By Western blot, both E1 and E2 were detected as glycosylated proteins with MW higher than those of polypeptide backbones, respectively (Figure 1). Secreted products were also detected for E1[26] and E2 (data not shown).
Figure 1.
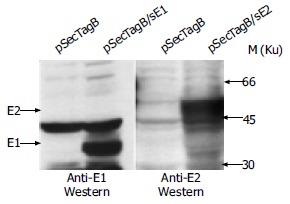
Transient expression with plasmids used in vaccination. Plasmids used for transfection are indicated at the top of each lanes. E1 and E2 products are indicated by arrowheads.
Humoral immune responses after DNA vaccination
DNA vaccination using the above characterized plasmids was carried out as described in Materials and Methods. After two injections, HCV E1 and E2 specific antibodies were detected in the sera of several mice. Without CpG adjuvant, the seroconversion rate was 2/5 for anti-E1 antibody, and 1/5 for anti-E2 antibody. When CpG was included as an adjuvant, the seroconversion rate was 3/4 for anti-E1 antibody and 2/4 for anti-E2 antibody. After the third injection, all animals became seroconverted to both anti-E1 and anti-E2 antibodies. The highest anti-E1 titer was 1:320 after the third injection for mice receiving plasmids only, while the highest titer for mice receiving plasmids with CpG was 80. The highest anti-E2 titers for both groups reached 1280, but the average anti-E2 titer for mice receiving CpG was slightly lower than that for those receiving no CpG (Table 1).
Table 1.
Anti-HCV titers1 after the third injection
| Vaccine |
Anti-E1 titers |
||||
| 1 | 2 | 3 | 4 | 5 | |
| pSecTagB/sE1 + pSecTagB/sE22 | 320 | 40 | Dead animal | 320 | 320 |
| pSecTagB/sE1 + pSecTagB/sE2 + CpG3 | 80 | 80 | Dead animal | 80 | 80 |
| Anti-E2 titers | |||||
| 1 | 2 | 3 | 4 | 5 | |
| pSecTagB/sE1 + pSecTagB/sE22 | 160 | 160 | Dead animal | 1280 | 1280 |
| pSecTagB/sE1 + pSecTagB/sE2 + CpG3 | 160 | 160 | Dead animal | 160 | 1280 |
1Titers were determined as described in Materials and Methods. Serial dilution of anti-sera started from 1:40, thus titers lower than 40 were not determined and considered 0.
The cutoff value was established as mean A of sera from the control mice vaccinated with pSecTagB.
The cutoff value was established as mean A of sera from the control mice vaccinated with pSecTagB + CpG.
E2 specific splenocyte proliferation
All the animals were sacrificed 30 d after the last injection to analyze cellular immune responses of the memory phase. Single-cell suspension of splenocytes was prepared for each individual animal. Splenocytes were immediately cultured in the presence of HCV E1 peptide or E2 protein. [3H] thymidine was then added to the cells to measure HCV antigen specific proliferation. In our experiments, HCV E1 specific proliferation was not observed, neither was E1 specific cytokine secretion (data not shown), possibly due to insufficient stimulation. For E2 specific splenocyte proliferation, as shown in Figure 2, animals vaccinated with pSecTagB only or pSecTagB plus CpG had stimulation indexes similar to that of naïve mice. Some of the animals vaccinated with plasmids pSecTagB/sE1 + pSecTagB/sE2 showed E2 specific splenocyte proliferation while the others did not. This gave an average SI slightly higher than that of pSecTagB vaccinated mice. Animals vaccinated with HCV gene encoding plasmids together with CpG all showed E2 specific splenocyte proliferation, which resulted in a significant difference (P = 0.005) of SI between animals injected with pSecTagB/sE1 + pSecTagB/sE2 + CpG and animals injected with pSecTagB + CpG.
Figure 2.
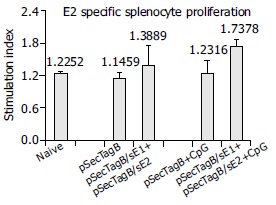
E2-specific splenocyte proliferation. Error bars represent the standard errors. Asterisk indicates the significant difference of the average SI between animals injected with (pSecTagB/sE1 + pSecTagB/sE2 + CpG) and animals injected with (pSecTagB + CpG) (P = 0.005).
E2 specific cytokine secretion
Cellular immune responses were also evaluated by cytokine secretion assay of the in vitro stimulated splenocytes from all vaccinated animals. INF-γ secretion is a marker of Th1 type antigen-specific cellular immune responses and plays key roles in fighting virus infection. Mice injected with pSecTagB/sE1 + pSecTagB/sE2 showed heterogeneous cellular immune responses, some had E2 stimulated INF-γ secretion, while the rest did not. This resulted in a slightly higher average of SI compared to the control group injected with pSecTagB (Figure 3A). Mice injected with pSecTagB/sE1 + pSecTagB/sE2 + CpG all showed highly E2-specific INF-γ secretion, showing a SI higher than 3, while the control group injected with pSecTagB + CpG had an average SI similar to those of naïve mice and mice injected with pSecTagB alone. The difference of SI between the experiment group and its control group was very significant (P = 0.0003).
Figure 3.
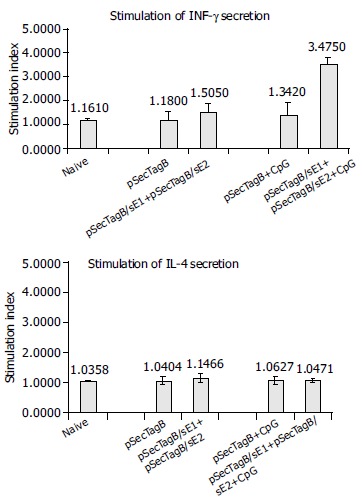
E2-specific cytokine secretion. Error bars represent the standard errors. Asterisk indicates the very significant difference of the average SI between animals injected with (pSecTagB/sE1 + pSecTagB/sE2 + CpG) and animals injected with (pSecTagB + CpG) in terms of INF-γ secretion (P = 0.0003).
IL-4 secretion is a marker of Th2-like cellular immune responses. No significant difference in IL-4 secretion was detected between all groups receiving different inoculums (Figure 3B). IL-4 secretion of the vaccinated animals was similar to that of naïve mice, indicating that Th2-like cellular immune response was decreased to basal level, if there was ever a Th2-like response. In terms of IL-4 secretion, no effect of CpG was detected.
DISCUSSION
In this study, target HCV antigens encoded by the DNA vaccine candidate, E1 and E2, were delivered by two separate plasmids that achieved higher-level expression and appropriate glycosylation of both antigens. DNA immunization results demonstrated that this DNA vaccine candidate could induce HCV specific humoral and cellular immune responses in mice. Anti-E2 titer obtained in this study was similar to our previous study[11], which was also comparable to the results from other laboratories[31-35]. We also detected comparable levels of anti-E1 antibodies in the sera of vaccinated mice, which together with anti-E2 antibodies, might be important in neutralizing the circulating HCV virus and preventing its spread in patients. HCV antigen specific splenocyte proliferation indicated that our candidate DNA vaccine was able to induce cellular immune response, which was critical in viral clearance. Cytokine secretion analysis revealed that the cellular immune response is Th1 type, which could activate CTL to remove cells with HCV antigen expression. Studies of HCV patients showed that protection against HCV infection was positively correlated with cellular immune responses[36,37], indicating that cellular immune responses might be more important in evaluating a candidate vaccine. The ability of our candidate vaccine to induce HCV specific cellular immune responses makes it a favorable choice in the development of prophylactic and therapeutic vaccine against HCV.
CpG motif, that is, CpG dinucleotide in particular base context (XCGY, where X is any base but C, and Y is any base but G), was first found to be immunoactive in activating B cells[38]. These CpG motifs are prevalent in bacterial and many viral DNAs but are heavily methylated and suppressed in vertebrate genomes[38,39]. Recent studies have shown that the immune system could respond to CpG motifs by activating potent Th1-like immune responses[23]. Several reports demonstrated that CpG could enhance humoral immune responses elicited by DNA vaccination[24,25], possibly due to indirect effect of enhanced cellular immune responses. In our vaccination experiment, we observed that after the second injection, the seroconversion rate for mice receiving CpG adjuvant was higher than that for those not receiving CpG. This enhanced humoral immune response at the early stage was likely due to the indirect positive effect of CpG on antigen presentation (via activation of antigen presenting cells by Th1). However, after the third injection, when all animals became seroconverted, those who received CpG as an adjuvant showed lower titers of both anti-E1 and anti-E2. This was consistent with the notion that when Th1-like response is the major immune response, Th2-like response is usually inhibited, followed by decreased antibody production. Further analysis of antigen specific cytokine secretion was consistent with this explanation. E2 specific INF-γ secretion was enhanced in splenocytes from those animals receiving CpG in vaccination, while this adjuvant did not change the E2 specific IL-4 secretion profile. These results indicated that CpG enhanced the Th1-like cellular immune response.
Apart from DNA vaccine, live virus vaccine and peptide vaccine are also efficient in inducing cellular immune response. Live virus vaccine using several different viral vectors has been under investigation[8,28,40], with continuous promising progresses. However, the interest in peptide vaccine is diminishing. The problem lies in the polymorphism of MHC molecules in the population. Certain individuals may have the MHC molecules necessary to bind a specific antigenic peptide, while others may not. Thus, the universality of a vaccine based on a specific peptide becomes a concern. Recently, combinational vaccination strategy with different species of vaccine components has been successful in laboratory in protecting against some elusive infectious reagents[41,42]. Combinational vaccination could reduce host immune responses against the viral vector itself, thus enhancing the efficiency of delivery. More importantly, combinational vaccination with different components could activate different components of the host immune surveillance system and result in enhanced immune responses. Our next effort will be focused on combinational vaccination with DNA vaccine descried above as one component.
ACKNOWLEDGEMENT
We thank Professor Yu Wang (Peking University) for providing HCV cDNA for this study.
Footnotes
Supported by the National High Technology R&D Program of China, No. 2001AA215171
Edited by Zhang JZ and Wang XL Proofread by Xu FM
References
- 1.Cohen J. The scientific challenge of hepatitis C. Science. 1999;285:26–30. doi: 10.1126/science.285.5424.26. [DOI] [PubMed] [Google Scholar]
- 2.McHutchison JG, Gordon SC, Schiff ER, Shiffman ML, Lee WM, Rustgi VK, Goodman ZD, Ling MH, Cort S, Albrecht JK. Interferon alfa-2b alone or in combination with ribavirin as initial treatment for chronic hepatitis C. Hepatitis Interventional Therapy Group. N Engl J Med. 1998;339:1485–1492. doi: 10.1056/NEJM199811193392101. [DOI] [PubMed] [Google Scholar]
- 3.Zein CO, Zein NN. Advances in therapy for hepatitis C infection. Microbes Infect. 2002;4:1237–1246. doi: 10.1016/s1286-4579(02)01651-9. [DOI] [PubMed] [Google Scholar]
- 4.Williams I. Epidemiology of hepatitis C in the United States. Am J Med. 1999;107:2S–9S. doi: 10.1016/s0002-9343(99)00373-3. [DOI] [PubMed] [Google Scholar]
- 5.Choo QL, Kuo G, Ralston R, Weiner A, Chien D, Van Nest G, Han J, Berger K, Thudium K, Kuo C. Vaccination of chimpanzees against infection by the hepatitis C virus. Proc Natl Acad Sci USA. 1994;91:1294–1298. doi: 10.1073/pnas.91.4.1294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Fournillier A, Depla E, Karayiannis P, Vidalin O, Maertens G, Trépo C, Inchauspé G. Expression of noncovalent hepatitis C virus envelope E1-E2 complexes is not required for the induction of antibodies with neutralizing properties following DNA immunization. J Virol. 1999;73:7497–7504. doi: 10.1128/jvi.73.9.7497-7504.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Gordon EJ, Bhat R, Liu Q, Wang YF, Tackney C, Prince AM. Immune responses to hepatitis C virus structural and nonstructural proteins induced by plasmid DNA immunizations. J Infect Dis. 2000;181:42–50. doi: 10.1086/315162. [DOI] [PubMed] [Google Scholar]
- 8.Makimura M, Miyake S, Akino N, Takamori K, Matsuura Y, Miyamura T, Saito I. Induction of antibodies against structural proteins of hepatitis C virus in mice using recombinant adenovirus. Vaccine. 1996;14:28–36. doi: 10.1016/0264-410x(95)00161-s. [DOI] [PubMed] [Google Scholar]
- 9.Davis GL. Hepatitis C virus genotypes and quasispecies. Am J Med. 1999;107:21S–26S. doi: 10.1016/s0002-9343(99)00376-9. [DOI] [PubMed] [Google Scholar]
- 10.Chen S, Wang YM. Genetic evolution of structural region of hepatitis C virus in primary infection. World J Gastroenterol. 2002;8:686–693. doi: 10.3748/wjg.v8.i4.686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhu J, Wang CL, Zhu LX, Kong YY, Wang Y, Li GD. DNA immunization with recombinant HCV E2 expression plasmids. Sheng Wu Hua Xue Yu Sheng Wu Wu Li Xue Bao (Shanghai) 2002;34:445–451. [PubMed] [Google Scholar]
- 12.Wang Y, Okamoto H, Tsuda F, Nagayama R, Tao QM, Mishiro S. Prevalence, genotypes, and an isolate (HC-C2) of hepatitis C virus in Chinese patients with liver disease. J Med Virol. 1993;40:254–260. doi: 10.1002/jmv.1890400316. [DOI] [PubMed] [Google Scholar]
- 13.Gurunathan S, Wu CY, Freidag BL, Seder RA. DNA vaccines: a key for inducing long-term cellular immunity. Curr Opin Immunol. 2000;12:442–447. doi: 10.1016/s0952-7915(00)00118-7. [DOI] [PubMed] [Google Scholar]
- 14.Hijikata M, Kato N, Ootsuyama Y, Nakagawa M, Shimotohno K. Gene mapping of the putative structural region of the hepatitis C virus genome by in vitro processing analysis. Proc Natl Acad Sci USA. 1991;88:5547–5551. doi: 10.1073/pnas.88.13.5547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rosa D, Campagnoli S, Moretto C, Guenzi E, Cousens L, Chin M, Dong C, Weiner AJ, Lau JY, Choo QL, et al. A quantitative test to estimate neutralizing antibodies to the hepatitis C virus: cytofluorimetric assessment of envelope glycoprotein 2 binding to target cells. Proc Natl Acad Sci USA. 1996;93:1759–1763. doi: 10.1073/pnas.93.5.1759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Pileri P, Uematsu Y, Campagnoli S, Galli G, Falugi F, Petracca R, Weiner AJ, Houghton M, Rosa D, Grandi G, et al. Binding of hepatitis C virus to CD81. Science. 1998;282:938–941. doi: 10.1126/science.282.5390.938. [DOI] [PubMed] [Google Scholar]
- 17.Zonaro A, Ravaggi A, Puoti M, Kremsdorf D, Albertini A, Cariani E. Differential pattern of sequence heterogeneity in the hepatitis C virus E1 and E2/NS1 proteins. J Hepatol. 1994;21:858–865. doi: 10.1016/s0168-8278(94)80250-5. [DOI] [PubMed] [Google Scholar]
- 18.Shimizu YK, Hijikata M, Iwamoto A, Alter HJ, Purcell RH, Yoshikura H. Neutralizing antibodies against hepatitis C virus and the emergence of neutralization escape mutant viruses. J Virol. 1994;68:1494–1500. doi: 10.1128/jvi.68.3.1494-1500.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zibert A, Schreier E, Roggendorf M. Antibodies in human sera specific to hypervariable region 1 of hepatitis C virus can block viral attachment. Virology. 1995;208:653–661. doi: 10.1006/viro.1995.1196. [DOI] [PubMed] [Google Scholar]
- 20.Lechner S, Rispeter K, Meisel H, Kraas W, Jung G, Roggendorf M, Zibert A. Antibodies directed to envelope proteins of hepatitis C virus outside of hypervariable region 1. Virology. 1998;243:313–321. doi: 10.1006/viro.1998.9069. [DOI] [PubMed] [Google Scholar]
- 21.Meunier JC, Fournillier A, Choukhi A, Cahour A, Cocquerel L, Dubuisson J, Wychowski C. Analysis of the glycosylation sites of hepatitis C virus (HCV) glycoprotein E1 and the influence of E1 glycans on the formation of the HCV glycoprotein complex. J Gen Virol. 1999;80(Pt 4):887–896. doi: 10.1099/0022-1317-80-4-887. [DOI] [PubMed] [Google Scholar]
- 22.Meyer K, Basu A, Ray R. Functional features of hepatitis C virus glycoproteins for pseudotype virus entry into mammalian cells. Virology. 2000;276:214–226. doi: 10.1006/viro.2000.0547. [DOI] [PubMed] [Google Scholar]
- 23.Krieg AM. CpG motifs in bacterial DNA and their immune effects. Annu Rev Immunol. 2002;20:709–760. doi: 10.1146/annurev.immunol.20.100301.064842. [DOI] [PubMed] [Google Scholar]
- 24.Moldoveanu Z, Love-Homan L, Huang WQ, Krieg AM. CpG DNA, a novel immune enhancer for systemic and mucosal immunization with influenza virus. Vaccine. 1998;16:1216–1224. doi: 10.1016/s0264-410x(98)80122-9. [DOI] [PubMed] [Google Scholar]
- 25.Encke J, zu Putlitz J, Stremmel W, Wands JR. CpG immuno-stimulatory motifs enhance humoral immune responses against hepatitis C virus core protein after DNA-based immunization. Arch Virol. 2003;148:435–448. doi: 10.1007/s00705-002-0935-y. [DOI] [PubMed] [Google Scholar]
- 26.Zhu J, Kong YY, Liu J, Zhang ZC, Wang Y, Li GD. Secretory Expression of Different C-terminal Truncated HCV E1 Proteins in Mammalian Cells and Characterization of the Expressed Products. Sheng Wu Hua Xue Yu Sheng Wu Wu Li Xue Bao (Shanghai) 2001;33:634–640. [PubMed] [Google Scholar]
- 27.Chu RS, Targoni OS, Krieg AM, Lehmann PV, Harding CV. CpG oligodeoxynucleotides act as adjuvants that switch on T helper 1 (Th1) immunity. J Exp Med. 1997;186:1623–1631. doi: 10.1084/jem.186.10.1623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Liu J, Zhu L, Zhang X, Lu M, Kong Y, Wang Y, Li G. Expression, purification, immunological characterization and application of Escherichia coli-derived hepatitis C virus E2 proteins. Biotechnol Appl Biochem. 2001;34:109–119. doi: 10.1042/ba20010036. [DOI] [PubMed] [Google Scholar]
- 29.Sato K, Okamoto H, Aihara S, Hoshi Y, Tanaka T, Mishiro S. Demonstration of sugar moiety on the surface of hepatitis C virions recovered from the circulation of infected humans. Virology. 1993;196:354–357. doi: 10.1006/viro.1993.1488. [DOI] [PubMed] [Google Scholar]
- 30.Inudoh M, Nyunoya H, Tanaka T, Hijikata M, Kato N, Shimotohno K. Antigenicity of hepatitis C virus envelope proteins expressed in Chinese hamster ovary cells. Vaccine. 1996;14:1590–1596. doi: 10.1016/s0264-410x(96)00154-5. [DOI] [PubMed] [Google Scholar]
- 31.Zucchelli S, Capone S, Fattori E, Folgori A, Di Marco A, Casimiro D, Simon AJ, Laufer R, La Monica N, Cortese R, et al. Enhancing B- and T-cell immune response to a hepatitis C virus E2 DNA vaccine by intramuscular electrical gene transfer. J Virol. 2000;74:11598–11607. doi: 10.1128/jvi.74.24.11598-11607.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Tedeschi V, Akatsuka T, Shih JW, Battegay M, Feinstone SM. A specific antibody response to HCV E2 elicited in mice by intramuscular inoculation of plasmid DNA containing coding sequences for E2. Hepatology. 1997;25:459–462. doi: 10.1002/hep.510250234. [DOI] [PubMed] [Google Scholar]
- 33.Ou-Yang P, Hwang LH, Tao MH, Chiang BL, Chen DS. Co-delivery of GM-CSF gene enhances the immune responses of hepatitis C viral core protein-expressing DNA vaccine: role of dendritic cells. J Med Virol. 2002;66:320–328. doi: 10.1002/jmv.2148. [DOI] [PubMed] [Google Scholar]
- 34.Encke J, zu Putlitz J, Geissler M, Wands JR. Genetic immunization generates cellular and humoral immune responses against the nonstructural proteins of the hepatitis C virus in a murine model. J Immunol. 1998;161:4917–4923. [PubMed] [Google Scholar]
- 35.Dou J, Liu K, Chen Z, Wo J, He N, Liu Y, Zhang M, Wang X, Xu C. Effect of immunization in mice with recombinant DNA encoding the hepatitis C virus structural protein. Chin Med J (Engl) 1999;112:1036–1039. [PubMed] [Google Scholar]
- 36.Cooper S, Erickson AL, Adams EJ, Kansopon J, Weiner AJ, Chien DY, Houghton M, Parham P, Walker CM. Analysis of a successful immune response against hepatitis C virus. Immunity. 1999;10:439–449. doi: 10.1016/s1074-7613(00)80044-8. [DOI] [PubMed] [Google Scholar]
- 37.Lechner F, Wong DK, Dunbar PR, Chapman R, Chung RT, Dohrenwend P, Robbins G, Phillips R, Klenerman P, Walker BD. Analysis of successful immune responses in persons infected with hepatitis C virus. J Exp Med. 2000;191:1499–1512. doi: 10.1084/jem.191.9.1499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Krieg AM, Yi AK, Matson S, Waldschmidt TJ, Bishop GA, Teasdale R, Koretzky GA, Klinman DM. CpG motifs in bacterial DNA trigger direct B-cell activation. Nature. 1995;374:546–549. doi: 10.1038/374546a0. [DOI] [PubMed] [Google Scholar]
- 39.Jones PA. The DNA methylation paradox. Trends Genet. 1999;15:34–37. doi: 10.1016/s0168-9525(98)01636-9. [DOI] [PubMed] [Google Scholar]
- 40.Ezelle HJ, Markovic D, Barber GN. Generation of hepatitis C virus-like particles by use of a recombinant vesicular stomatitis virus vector. J Virol. 2002;76:12325–12334. doi: 10.1128/JVI.76.23.12325-12334.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Amara RR, Villinger F, Altman JD, Lydy SL, O'Neil SP, Staprans SI, Montefiori DC, Xu Y, Herndon JG, Wyatt LS, et al. Control of a mucosal challenge and prevention of AIDS by a multiprotein DNA/MVA vaccine. Science. 2001;292:69–74. doi: 10.1126/science.1058915. [DOI] [PubMed] [Google Scholar]
- 42.Haddad D, Liljeqvist S, Ståhl S, Hansson M, Perlmann P, Ahlborg N, Berzins K. Characterization of antibody responses to a Plasmodium falciparum blood-stage antigen induced by a DNA prime/protein boost immunization protocol. Scand J Immunol. 1999;49:506–514. doi: 10.1046/j.1365-3083.1999.00543.x. [DOI] [PubMed] [Google Scholar]