

Abstract
AIM: To investigate the expression of gonadotropin-releasing hormone (GnRH) receptor and the effects of GnRH analog (alarelin) on proliferation of cultured gastric smooth muscle cells (GSMC) of rats.
METHODS: Immunohistochemical ABC methods and in situ hybridization methods were used to dectect protein and mRNA expression of GnRH receptor in GSMC, respectively. Techniques of cell culture, OD value of MTT test, measure of 3H-TdR incorporation, average fluorescent values of proliferating cell nuclear antigen (PCNA) and flow cytometric DNA analysis were used in the experiment.
RESULTS: The cultured GSMC of rats showed immunoreactivity for GnRH receptor; positive staining was located in cytoplasm. GnRH receptor mRNA hybridized signals were also detected in cytoplasm. When alarelin (10-9, 10-7, 10-5 mol/L) was administered into the medium and incubated for 24 h, OD value of MTT, 3H-TdR incorporation and average fluorescent values of PCNA all decreased significantly as compared with the control group (P < 0.05). The maximum inhibitory effect on cell proliferation was achieved a concentration of 10-5 mol/L and it acted in a dose-dependent manner. Flow cytometric DNA analysis revealed that alarelin could significantly enhance ratio of G1 phase and decrease ratio of S phase of GSMC of rats (P < 0.05).The maximum inhibitory effect on ratio of S phase was at the concentration of 10-5 mol/L and also acted in a dose-dependent manner.
CONCLUSION: Our data suggest that GnRH receptor can be expressed by GSMC of rats. GnRH analogue can directly inhibit proliferation and DNA synthesis of rat GSMC through GnRH receptors.
INTRODUCTION
Gonadotropin releasing hormone (GnRH), a decapeptide synthesized by the neuroendocrine cells in the pro-optic area of brain, plays an important role in regulation of reproductive function. After released from the hypothalamus, GnRH binds to a specific receptor on the plasma membrane of the pituitary gonadotroph, resulting in stimulation of FSH and LH biosynthesis and release. There is evidence that GnRH and its receptors are expressed in numerous extrapituitary tissues, such as syncytiotrophoblast cells[1], human granulosa-Luteal cells[2], human peripheral lymphocytes[3,4] and so on. They are not only expressed in normal tissues, but also widely exist in various carcinoma that derived from reproductive (ovary cancer[5] and prostate cancer[6]) or non-reproductive tissues (breast cancer[7], lung cancer[8], pancreas cancer[9] and hepatocellular carcinoma, et al[10]). Our previous studies have shown the presence of GnRH and its receptor in rat digestive tract[11], submaxillary gland[12], thyroid glang[13] and pancreas[14] and proposed that GnRH could be classified as a kind of brain-gut hormone. Presently, we first investigated the expression of GnRH receptor in cultured rat gastric smooth muscle cells (GSMC) and investigate the effect of GnRH analogue (alarelin) on the proliferation of cultured GSMC of rats.
MATERIALS AND METHODS
Materials
GnRH anti-idiotypic antibody was prepared by our department[15]. ABC kit was the product of Vector Company (Vector Laboratories Burlingame, CA, U.S.A.). MTT (3-(4, 5-dimethythiazol -zyl-2)-2, 5-diphenyltetrazolium), colleagnase I, PPO (2, 5-diphenyloxazole) and POPOP (1, 4-bis (5-phenyloxazol-2-yl) benzene) were purchased from Sigma Company (Staint Louis, Missouri, U.S.A.). DMEM and Trypsin were the products of Gibco Company (Carlsbad, California, U.S.A.). Alarelin was purchased from Shanghai Lizhu Biochemical and Pharmaceutical Company, Shanghai, China. PCNA (Proliferating cell nuclear antigen) antibody and immunofluorescent kit were purchased from Boster Company (Wuhan, China). 3H-TdR was purchased from Academy of Atomic Energy Sciences of China (Shanghai, China). The 1-3-day-old male Sprague-Dawley rats were purchased from the Center of Laboratory Animal Medicine, the Fourth Military Medical University, (Xi’an, China).
Preparation and culture of GSMC of rats
The culture of GSMC of rats was performed as described previously with modifications[16,17]. 1-3-day-old immature male Sprague-Dawley rats were sterilized by 750 mL/L alchol; removed the stomaches and put them into cool Hanks solution (mmol/L: CaCl2 1.3, KCl 5.4, KH2PO4 0.4, MgCl2·6H2O 0.4, MgSO4·7H2O 0.4, NaCl 137, glucose 5.6) immediately. The mucosas were removed by scrapping with razorblade. The gastric smooth muscle tissues were minced into l.0 mm3 pieces with tissue chopper, then put them into DMEM containing 1 mg/L collagenase I at 37 °C for 30-60 min, and the GSMCs were collected at an interval of 20 min. The cell suspension was centrifuged at 500 r/min for 5 min, washed 2 times in DMEM, then cultured in DMEM containing 100 mL/L calf serum, 100 U/mL pencillin G and 100 U/mL streptomycin under a humidified atmosphere of 50 mL/L CO2 in air. To obtain pure GSMC and remove the fibroblasts, the cells were preattached in flasks with 10% calf serum under DMEM in a humidified atmosphere of 5% CO2 or 90 min at 37 °C. Afterwards, the cells were maintained in fresh DMEM containing 100 mL/L calf serum and removed it per 3 d and subsequently subcultured per 4-5 d by treatment with trypsin (2.5 g/L). The GSMC were identified by “valley and hill appearance”[17] and showed α-actin immunoreactivity, which was located in cytoplasm. The experiments were performed with monolayer of cells from passage 3-5.
Immunocytochemical procedure
The GSMCs (1×105/mL) were seeded in a six-well plate containing the coverglasses inside. The cells of passage 3-5 were fixed in 4% paraformaldehyde solution for 30 min at room temperature. The coverglasses were washed 3 times with phosphate-buffered saline (PBS, pH7.2), and put into methanol-H2O2 for 30 min to remove endogenous peroxidase activity. Then they were stained using immunohistochemical ABC method. They were incubated with first antibody, rabbit GnRH anti-idiotypic antibody (1:200 dilution) at 4 °C for 18-24 h. Then they were incubabated with second antibody, biotin labeled goat anti-rabbit IgG antibody (1:400 dilution), at room temperature for 2 h, followed by incubation with ABC complex (1:400 dilution), at room temperature for 1 h. Finally, DAB (diaminobenzedin) was used for color developed, and then the slides were washed in distilled water to stop the reaction, dehydrated in alcohol, hyalinized in xylene, and mounted on Canada balsam. The first antibody was replaced by normal and diluted rabbit serum for the negative control.
In situ hybridization
The probe was produced according to the cDNA sequence of the rat’s pituitary GnRH receptor[12,18]. The sequence was: 5’-ATGTATGCCCCCAGCCTTCATGATGGTGATTAGCC TGGATC-3’. The digoxigeni was labeled at 5’-end by the TaKaRa Biotechnology Company, (Dalian, China). The in situ hybridization was carried out on the coverglasses after the cells were fixed in 40 g/L paraformaldehyde containing 1 g/L DEPC (Diethyl pyrocarbonate) for 30 min. Then they were processed as follows: The coverglasses were rinsed in 0.01 mol/L PBS (pH7.2) for 5 min×3, in Proteinase K (1 μg/mL) for 2 min at 37 °C. They were post-fixed in 40 g/L paraformaldehyde for 5 min, washed in distilled water for 5 min×3. After being dried in the air, each coverglass was applied with the hybridization buffer containing GnRH receptor oligonuclicotide probe (2.0 μg/mL), and hybridization was carried out in sealed humid box at 42 °C for 20 h. After that the slides were washed with 2×SSC at 37 °C for 5 min×3, 0.1×SSC at 37 °C for 10 min×2, they were incubated with mouse-anti-dig antibody at room temperature for 2 h. Thereafter, the slides were washed with 0.5 mol/L TBS for 5 min×3, followed by incubation with biotin-labeled goat-anti-mouse IgG antibody for 1 h at 37 °C. The slides were washed with 0.5 mol/L TBS for 5 min×3 and incubated with SABC-AP for 30 min at 37 °C. The sections were rinsed in 0.5 mol/L TBS for 5 min×4, and the alkaline phosphatase reaction was conducted by incubation with complex solution of NBT (Nitrio-blue tetrazolium) and BCIP (5-bromo-4-chloro -3-indo-lyl phosphate) for 20-30 min. Finally, the slides were washed in distilled water to stop the reaction, and then dehydrated in alcohol, hyalinized in xylene, and mounted on Canada balsam. The slides were incubated with normal mouse serum instead of the mouse-anti-dig antibody in absence of the labeled probe for the negative controls.
MTT assay
The cells were trypsinized in a solution of 2.5 g/L trypsin and seeded in a 96-well plate at a density of 104 cells/(0.1 mL/well). After the cells were grown for 24 h to approximately 800 g/L subconfluent state, 0.1 mL medium containing 2.5% calf serum and various concentrations (10-9, 10-7, 10-5 mol/ L) of alarelin was added to each well, respectively, and incubated for 24 h in a CO2 incubator. Each concentration was tested in at least 12 wells. The MTT assay was performed as described previously with mondifications[19]. Briefly, 15 μL of MTT solution (5 g/L, PBS, pH7.2) was added to each well and incubated for 4 h. Then, the medium and MTT were removed and 150 μL of DMSO was added to each well and shaken for 10 min to dissolve the crystal. The OD was determined at 490 nm using an ELISA reader.
3H-TdR incorporation
To observe the role of GnRH analogue in growth regulation of GSMC, a 3H-TdR incorporation assay was performed as previously described[20]. The cells were plated into a 96-well plate at a density of 104 cells/(0.1 mL/well). After the cells were grown for 24 h, 0.1 mL medium containing 2.5% calf serum and various concentrations (10-9, 10-7, 10-5 mol/L) of alarelin was added to each well, respectively, and incubated for 16 h in a CO2 incubator. Then the 3H-TdR (2 μCi/mL) was added to each well and continuously incubated for 8 h. Incubation was stopped by adding 1 vol of cold 100 mL/L trichloracetic acid (TCA) which disrupted cells and precipitated the macromolecules. After being washed with a methanol, the precipitate was collected in 9999 fiberglass filter paper, respectively. When the filter paper was dried, 0.5 mL scintillant liquid were mixed and the radioactivity was estimated with a scintillation counter (Backman LS5801). Each concentration was tested in at least 6 wells.
Fluorescent immunocytochemical reaction of PCNA in GSMC
The cells were seeded in six-well plate containing the coverglasses inside. After the cells were grown for 24 h to approximately 800 g/L subconfluent state, 2 mL medium containing 25 mL/L calf serum and various concentrations (10-9, 10 -7, 10-5 mol/L) of alarelin was added to each well, respectively, and incubated for another 24 h in a CO2 incubator. Then the monoplayers were fixed in 40 g/L paraformaldehyde solution for 30 min. The cell coverglasses were washed 3 times with PBS (pH7.2), and then put into methanol-H2O2 for 30 min to remove endogenous peroxidase activity. Then PCNA immunofluorescence reaction was carried out according to the manufacturer’ instructions. The coverglasses were incubated with the first antibody (mouse antibody, 1:200 dilution) at 4 °C overnight, followed by incubation with the second antibody, biotin labeled goat anti-rabbit IgG antibody (1:200 dilution), at room temperature for 2 h. Then the coverglasses were incubated with SABC-fluo Cy3 (1:100 dilution) at room temperature for 1 h. At last, the cells were detected by using the laser confocal microscopy, and scanned the average fluorescent values of PCNA of GSMC. For the negative controls, the first antibody was replaced by normal rabbit serum.
Cell-cycle analysis by flow cytomety
The cells were seeded in 12-well plate at a density of 107 cells/(1.5 mL/well). After the cells were grown for 24 h to approximately 800 g/L subconfluent state, 2 mL medium containing 25 mL/L calf serum and various concentrations (10-9-10-5 mol/L) of alarelin was added to each well, respectively, and incubated for another 48 h in a CO2 incubator. Then the cells were trypsinized in a solution of 2.5 g/L trypsin, fixed in 700 mL/L ethanol and stained by PI (propidium iodide) for DNA analysis. The flow cytometric analyses were performed using EPICS Profile II. DNA content was measured and ratio of each DNA period of cell proliferation was analyzed.
Data analysis
In the proliferation study, the data were expressed as mean ± SD. The data were analyzed by SPLM statistical software. P < 0.05 was considered statistically significant.
RESULTS
Expression and localization of GnRH receptor
Immunocytochemical reaction showed a dark-brown staining in cytoplasm for GnRH receptor immnoreactivity; background was light yellow or no staining; positive cells were identified easily. The two negative controls showed no staining for GnRH receptor. All cultured GSMCs showed immnoreactivity for GnRH receptor in the cytoplasm, but with variations in degrees of intensity of the immunorecativity (Figure 1).
Figure 1.
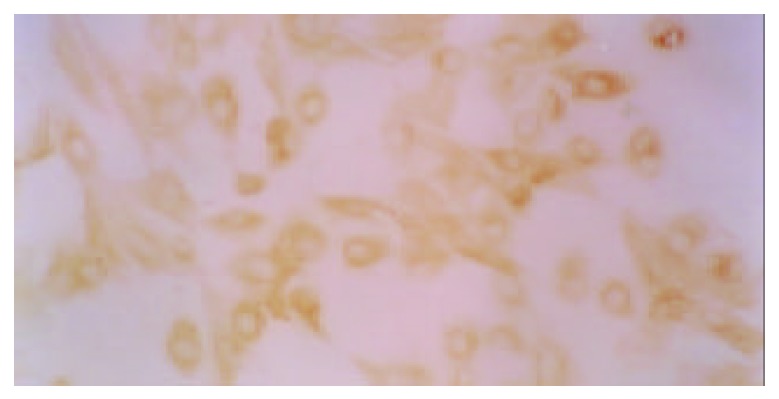
Gastric smooth muscle cells showed GnRH receptor immunoreactivity in cytoplasm. (×200).
In situ hybridization showed GnRH receptor mRNA hybridized signal was blue, and the background was not stained. The GnRH receptor mRNA hybridization signals could be detected in all cultured GSMCs, but with variation in signal strength between them. The signals were only distributed in the cytoplasm with hybridization negative nuclei (Figure 2). The two negative controls did not show GnRH receptor mRNA hybridized signal.
Figure 2.
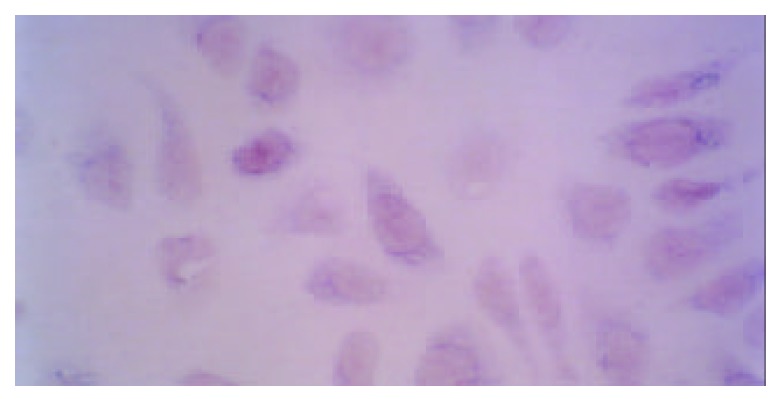
Gastric smooth muscle cells showed GnRH receptor mRNA hybridized signal in cytoplasm. (×400).
Effect of alarelin on GSMC viability
The influence of alarelin on GSMC viability was assessed using the MTT assay. The cell viability in the presence of alarelin was significantly lower than that in the absence of alarelin (F = 59.083, P < 0.01). The maximum stimulatory effect on cell viability was achieved at a concentration of 10-5 mol/L and it acted in a dose-dependent manner (Table 1).
Table 1.
Effects of alarelin on A value of MTT of rat GSMC
| Group | MTT/A value (n = 12) |
| Control | 0.728 ± 0.100 |
| 10-9 mol/L alarelin | 0.533 ± 0.073 |
| 10-7 mol/L alarelin | 0.368 ± 0.029 |
| 10-5 mol/L alarelin | 0.243 ± 0.042 |
3H-TdR Incorporation assay
To evaluate the functional role of GnRH in GSMC, the cells were treated with different concentrations of alarelin and incubated for 16 h, and then 3H-TdR (2 μCi/mL) was added. After 8 h, 3H-TdR incorporation assays were performed. As shown in Table 2, alarelin inhibited the growth of GSMC in a dose-dependent manner. A significant inhibition of proliferation was detected in the different groups, and the antiproliferative effect was the most evident when the concentration of alarelin at 10-5 mol/L (F = 22.33, P < 0.05).
Table 2.
Effects of alarelin on 3H-TdR Incorporation of rat GSMC
| Group | 3H-TdR Incorporation (n = 6) |
| Control | 1 936.50 ± 440.99 |
| 10-9 mol/L alarelin | 1 448.17 ± 327.72 |
| 10-7 mol/L alarelin | 945.83 ± 374.32 |
| 10-5 mol/L alarelin | 385.83 ± 184.66 |
The average fluorescent values of PCNA assay
The cells were treated with different concentrations of alarelin (10-9, 10-7, 10-5 mol/L) and incubated for another 24 h, and the average fluorescent values of PCNA assays were performed. As shown in Table 3, alarelin significantly inhibited the average fluorescent value in a dose-dependent manner compared with the control group (F = 15.86, P < 0.01).
Table 3.
Effects of alarelin on average fluorescent values of PCNA in GSMC
| Group | Average fluorescent values of PCNA (n = 5) |
| Control | 13.48 ± 1.31 |
| 10-9 mol/L alarelin | 11.37 ± 1.99 |
| 10-7 mol/L alarelin | 9.35 ± 1.38 |
| 10-5 mol/L alarelin | 7.37 ± 1.06 |
Effects of alarelin on GSMC cell cycles
Table 4 showed the ratio of G1 phase and S phase of DNA period of cell proliferation of each group. The ratio of G1 phase of DNA in group II- VI were markedly higher than that of the control, but did not increase in a dose-dependent manner (F = 74.94, P < 0.05). Moreover, the ratio of S phase of DNA was significantly lower in group II-VI than that in the control and acted in a dose-dependent manner (F = 32.5, P < 0.05, except the Group III, V compared with their border groups).(Figure 3)
Table 4.
Effects of alarelin on GSMC cell cycles (n = 4)
| Group | Concentration of alarelin | G1 phase (%) | S phase (%) |
| Group I | Control | 70.80 ± 11.30 | 17.50 ± 3.40 |
| Group II | 10-9 mol/L alarelin | 80.00 ± 9.42 | 9.93 ± 1.73 |
| Group III | 10-8 mol/L alarelin | 76.60 ± 8.99 | 7.10 ± 1.05 |
| Group IV | 10-7 mol/L alarelin | 79.60 ± 7.43 | 6.60 ± 1.54 |
| Group V | 10-6 mol/L alarelin | 78.50 ± 10.66 | 5.10 ± 1.01 |
| Group VI | 10-5 mol/L alarelin | 80.20 ± 10.03 | 3.20 ± 1.20 |
Figure 3.
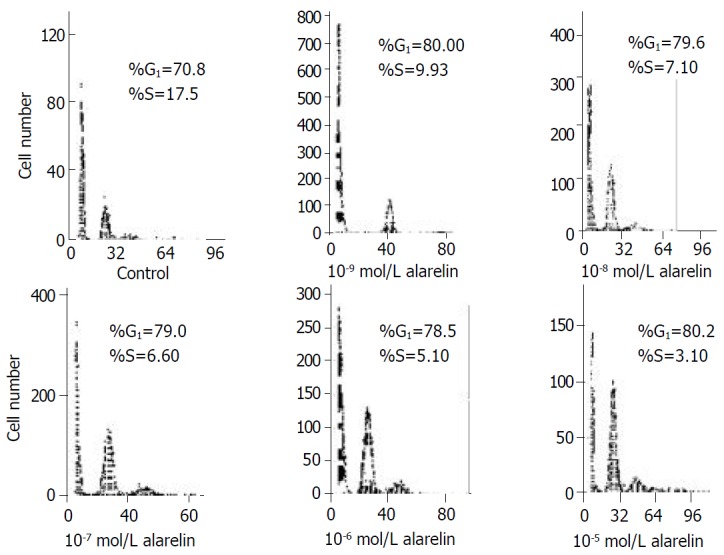
Effects of alarelin on DNA content of GSMCs cell cycles.
DISCUSSION
GnRH anti-idiotypic antibody has been widly applied in immunohistochemical studies of GnRH receptors[11,12,21]. GnRH and its receptor mainly exist in hypothalamus-pituitary axis, and also expressed in extra-hypothalamically, and has extra-pituitary activity. Studies have reported that pancreatic islets have GnRH immunoreactivity[14]. Jan et al[22] reported that GnRH was a possible transmitter in sympathetic ganglion in frog, and our previous studies[4,5] revealed that both GnRH and GnRH receptor were all existed in digestive tract. Our present study revealed, for the first time, that GnRH receptor mRNA was expressed in cultured GSMC of rats. These findings have raised intriguing questions which concern the physiological function of GnRH and its receptor.
GnRH has been suggested to regulate cell growth and proliferation through its receptor in expituitary tissues. GnRH analogues not only have been used and proven to be efficient in treating GnRH receptor-bearing tumors, including carcinomas of the ovary, breast, and endometrium[23-25], but also inhibited the growth and proliferation of normal cell, such as granulose cells and myometrial smooth muscle cells[26,27]. MTT assays and 3H-TdR assays are considered to be the most reliable techniques for measurement of the cell population. Using MTT assays and 3H-TdR assays, we demonstrated that GnRH analogue inhibited growth of normal GSMC of rats in a dose-dependent manner. This inhibitory effect is consistent with the results of Minaretzis et al[26] in granulosa-lutein cells and Chegini et al[27] in myometrial smooth muscle cells. This inhibitory effect of GnRH analogue appears to be receptor-mediated and its exact mechanism of growth inhibitory effect at the receptor level is unclear.
To examine whether GnRH receptor (GnRHR) mediates direct antiproliferative effects, we attempted to assess the effects of alarelin on average fluorescent values of PCNA and cell cycle of GSMC. PCNA, a protein which functions as a cofactor to DNA polymerase delta, has many characteristics which make it an effective marker for evaluating cell proliferation, particularly in S phase[28]. PCNA is a stable cell-cycle regulated nuclear protein, expressed differentially during the cell cycle[29,30], and it is believed to reflect the different stages of the cell cycles, which begins to increase in terminus of G1, reaches at peak in S phase, and decreases in G2 and M phases, and this is corresponded with the change of DNA content[28]. In the present study, we found that alarelin inhibited the average fluorescent values of PCNA, and acted in a dose-dependent manner. These might cause by the binding of GnRH analogue with its receptor, which induced the activation of phosphatidylinositol kinase activity, known to stimulate mitogenic response in plasma membranes[31]. By flow cytomety, we demonstrated the GnRH analogue improved the ratios of G1/G0 which did not act in a dose-dependent manner, and inhibited the ratios of S phase which acted in a dose-dependent manner. So, the reason of growth inhibition might be that GnRH analogue arrested at G1/S phase, which inhibits DNA synthesis. These results are consistent with the results of MTT assay, 3H-TdR incorporation and fluorescent value of PCNA.
In summary, we have demonstrated for the first time that GnRH receptor is expressed in GSMC and GnRH analogue has a direct growth inhibitory effect on GSMC of rats that is mediated through GnRH receptor. Our findings strongly suggest that GnRH play an important role in digestive tract.
Footnotes
Supported by the Natural Science Foundation of China, No. 39770388
Co-correspondents: Wei-Quan Huang
Edited by Kumar M and Xu FM
References
- 1.Currie WD, Setoyama T, Lee PS, Baimbridge KG, Church J, Yuen BH, Leung PC. Cytosolic free Ca2+ in human syncytiotrophoblast cells increased by gonadotropin-releasing hormone. Endocrinology. 1993;133:2220–2226. doi: 10.1210/endo.133.5.8404673. [DOI] [PubMed] [Google Scholar]
- 2.Nathwani PS, Kang SK, Cheng KW, Choi KC, Leung PC. Regulation of gonadotropin-releasing hormone and its receptor gene expression by 17beta-estradiol in cultured human granulosa-luteal cells. Endocrinology. 2000;141:1754–1763. doi: 10.1210/endo.141.5.7447. [DOI] [PubMed] [Google Scholar]
- 3.Azad N, La Paglia N, Jurgens KA, Kirsteins L, Emanuele NV, Kelley MR, Lawrence AM, Mohagheghpour N. Immunoactivation enhances the concentration of luteinizing hormone-releasing hormone peptide and its gene expression in human peripheral T-lymphocytes. Endocrinology. 1993;133:215–223. doi: 10.1210/endo.133.1.8319570. [DOI] [PubMed] [Google Scholar]
- 4.Weesner GD, Becker BA, Matteri RL. Expression of luteinizing hormone-releasing hormone and its receptor in porcine immune tissues. Life Sci. 1997;61:1643–1649. doi: 10.1016/s0024-3205(97)00769-8. [DOI] [PubMed] [Google Scholar]
- 5.Furui T, Imai A, Tamaya T. Intratumoral level of gonadotropin-releasing hormone in ovarian and endometrial cancers. Oncol Rep. 2002;9:349–352. [PubMed] [Google Scholar]
- 6.Leuschner C, Enright FM, Gawronska-Kozak B, Hansel W. Human prostate cancer cells and xenografts are targeted and destroyed through luteinizing hormone releasing hormone receptors. Prostate. 2003;56:239–249. doi: 10.1002/pros.10259. [DOI] [PubMed] [Google Scholar]
- 7.Chen A, Kaganovsky E, Rahimipour S, Ben-Aroya N, Okon E, Koch Y. Two forms of gonadotropin-releasing hormone (GnRH) are expressed in human breast tissue and overexpressed in breast cancer: a putative mechanism for the antiproliferative effect of GnRH by down-regulation of acidic ribosomal phosphoproteins P1 and P2. Cancer Res. 2002;62:1036–1044. [PubMed] [Google Scholar]
- 8.Nechushtan A, Yarkoni S, Marianovsky I, Lorberboum-Galski H. Adenocarcinoma cells are targeted by the new GnRH-PE66 chimeric toxin through specific gonadotropin-releasing hormone binding sites. J Biol Chem. 1997;272:11597–11603. doi: 10.1074/jbc.272.17.11597. [DOI] [PubMed] [Google Scholar]
- 9.Wang L, Xie LP, Zhang RQ. Gene expression of gonadotropin-releasing hormone and its receptor in rat pancreatic cancer cell lines. Endocrine. 2001;14:325–328. doi: 10.1385/ENDO:14:3:325. [DOI] [PubMed] [Google Scholar]
- 10.Yin H, Cheng KW, Hwa HL, Peng C, Auersperg N, Leung PC. Expression of the messenger RNA for gonadotropin-releasing hormone and its receptor in human cancer cell lines. Life Sci. 1998;62:2015–2023. doi: 10.1016/s0024-3205(98)00173-8. [DOI] [PubMed] [Google Scholar]
- 11.Huang W, Yao B, Sun L, Pu R, Wang L, Zhang R. Immunohistochemical and in situ hybridization studies of gonadotropin releasing hormone (GnRH) and its receptor in rat digestive tract. Life Sci. 2001;68:1727–1734. doi: 10.1016/s0024-3205(01)00968-7. [DOI] [PubMed] [Google Scholar]
- 12.Yao B, Huang W, Huang Y, Chui Y, Wang Y, Li H, Pu R, Wan L, Zhang R. A study on the localization and distribution of GnRH and its receptor in rat submaxillary glands by immunohistochemical, in situ hybridization and RT-PCR. Life Sci. 2003;72:2895–2904. doi: 10.1016/s0024-3205(03)00194-2. [DOI] [PubMed] [Google Scholar]
- 13.Zhou J, Huang WQ, Ji QH. The expression of gonadotropin-re-leasing hormone and its receptor in human thyroid gland. Acta Anatomica Sinica. 2002;33:511–515. [Google Scholar]
- 14.Wang L, Xie LP, Huang WQ, Yao B, Pu RL, Zhang RQ. Presence of gonadotropin-releasing hormone (GnRH) and its mRNA in rat pancreas. Mol Cell Endocrinol. 2001;172:185–191. doi: 10.1016/s0303-7207(00)00369-5. [DOI] [PubMed] [Google Scholar]
- 15.Huang WQ, Sun L, Zhong CL. Preparation and identification of GnRH anti-idiotypic antibody. Chin J Anatom. 1994;17(Suppl):353–356. [Google Scholar]
- 16.Fatigati V, Murphy RA. Actin and tropomyosin variants in smooth muscles. Dependence on tissue type. J Biol Chem. 1984;259:14383–14388. [PubMed] [Google Scholar]
- 17.Chamley-Campbell J, Campbell GR, Ross R. The smooth muscle cell in culture. Physiol Rev. 1979;59:1–61. doi: 10.1152/physrev.1979.59.1.1. [DOI] [PubMed] [Google Scholar]
- 18.Kaiser UB, Zhao D, Cardona GR, Chin WW. Isolation and characterization of cDNAs encoding the rat pituitary gonadotropin-releasing hormone receptor. Biochem Biophys Res Commun. 1992;189:1645–1652. doi: 10.1016/0006-291x(92)90266-n. [DOI] [PubMed] [Google Scholar]
- 19.Chiba K, Kawakami K, Tohyama K. Simultaneous evaluation of cell viability by neutral red, MTT and crystal violet staining assays of the same cells. Toxicol In Vitro. 1998;12:251–258. doi: 10.1016/s0887-2333(97)00107-0. [DOI] [PubMed] [Google Scholar]
- 20.Kang SK, Choi KC, Cheng KW, Nathwani PS, Auersperg N, Leung PC. Role of gonadotropin-releasing hormone as an autocrine growth factor in human ovarian surface epithelium. Endocrinology. 2000;141:72–80. doi: 10.1210/endo.141.1.7250. [DOI] [PubMed] [Google Scholar]
- 21.Ji QH, Huang WQ, Sun L, Zhao BQ. Double-labeling immunohis-tochemical location of glucagon, gonadorelin (GnRH) and GnRH receptor within pancreas of the guinea pig. J Fourth Milit Med Univ. 1998;19:34–36. [Google Scholar]
- 22.Jan YN, Jan LY, Kuffler SW. A peptide as a possible transmitter in sympathetic ganglia of the frog. Proc Natl Acad Sci U S A. 1979;76:1501–1505. doi: 10.1073/pnas.76.3.1501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gründker C, Emons G. Role of gonadotropin-releasing hormone (GnRH) in ovarian cancer. Reprod Biol Endocrinol. 2003;1:65. doi: 10.1186/1477-7827-1-65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Sayer HG, Höffken K. [Hormone therapy, chemotherapy and immunotherapy in breast carcinoma. The best strategy for your patient] MMW Fortschr Med. 2003;145:40–42. [PubMed] [Google Scholar]
- 25.Krauss T, Huschmand H, Hinney B, Viereck V, Emons G. [Hormonal therapy and chemotherapy of endometrial cancer] Zentralbl Gynakol. 2002;124:45–50. doi: 10.1055/s-2002-20309. [DOI] [PubMed] [Google Scholar]
- 26.Minaretzis D, Jakubowski M, Mortola JF, Pavlou SN. Gonadotropin-releasing hormone receptor gene expression in human ovary and granulosa-lutein cells. J Clin Endocrinol Metab. 1995;80:430–434. doi: 10.1210/jcem.80.2.7852501. [DOI] [PubMed] [Google Scholar]
- 27.Chegini N, Rong H, Dou Q, Kipersztok S, Williams RS. Gonadotropin-releasing hormone (GnRH) and GnRH receptor gene expression in human myometrium and leiomyomata and the direct action of GnRH analogs on myometrial smooth muscle cells and interaction with ovarian steroids in vitro. J Clin Endocrinol Metab. 1996;81:3215–3221. doi: 10.1210/jcem.81.9.8784072. [DOI] [PubMed] [Google Scholar]
- 28.Galand P, Del Bino G, Morret M, Capel P, Degraef C, Fokan D, Feremans W. PCNA immunopositivity index as a substitute to 3H-thymidine pulse-labeling index (TLI) in methanol-fixed human lymphocytes. Leukemia. 1995;9:1075–1084. [PubMed] [Google Scholar]
- 29.Kurki P, Vanderlaan M, Dolbeare F, Gray J, Tan EM. Expression of proliferating cell nuclear antigen (PCNA)/cyclin during the cell cycle. Exp Cell Res. 1986;166:209–219. doi: 10.1016/0014-4827(86)90520-3. [DOI] [PubMed] [Google Scholar]
- 30.Mathews MB, Bernstein RM, Franza BR, Garrels JI. Identity of the proliferating cell nuclear antigen and cyclin. Nature. 1984;309:374–376. doi: 10.1038/309374a0. [DOI] [PubMed] [Google Scholar]
- 31.Takagi H, Imai A, Furui T, Horibe S, Fuseya T, Tamaya T. Evidence for tight coupling of gonadotropin-releasing hormone receptors to phosphatidylinositol kinase in plasma membrane from ovarian carcinomas. Gynecol Oncol. 1995;58:110–115. doi: 10.1006/gyno.1995.1192. [DOI] [PubMed] [Google Scholar]