

Abstract
AIM: To select the peptide mimicking the neutralization epitope of hepatitis E virus which bound to non-type-specific and conformational monoclonal antibodies (mAbs) 8C11 and 8H3 fromed 7-peptide phage display library, and expressed the peptide recombinant with HBcAg in E.coli, and to observe whether the recombinant HBcAg could still form virus like particle (VLP) and to test the activation of the recombinant polyprotein and chemo-synthesized peptide that was selected by mAb 8H3.
METHODS: 8C11 and 8H3 were used to screen for binding peptides through a 7-peptide phage display library. After 4 rounds of panning, monoclonal phages were selected and sequenced. The obtained dominant peptide coding sequences was then synthesized and inserted into amino acid 78 to 83 of hepatitis B core antigen (HBcAg), and then expressed in E.coli. Activity of the recombinant proteins was detected by Western blotting, VLPs of the recombinant polyproteins were tested by transmission electron microscopy and binding activity of the chemo-synthesized peptide was confirmed by BIAcore biosensor.
RESULTS: Twenty-one positive monoclonal phages (10 for 8C11, and 11 for 8H3) were selected and the inserted fragments were sequenced. The DNA sequence coding for the obtained dominant peptides 8C11 (N’-His-Pro-Thr-Leu-Leu-Arg-Ile-C’, named 8C11A) and 8H3 (N’-Ser-Ile-Leu-Pro- Tyr-Pro-Tyr-C’, named 8H3A) were then synthesized and cloned to the HBcAg vector, then expressed in E.coli. The recombinant proteins aggregated into homodimer or polymer on SDS-PAGE, and could bind to mAb 8C11 and 8H3 in Western blotting. At the same time, the recombinant polyprotein could form virus like particles (VLPs), which could be visualized on electron micrograph. The dominant peptide 8H3A selected by mAb 8H3 was further chemo-synthesized, and its binding to mAb 8H3 could be detected by BIAcore biosensor.
CONCLUSION: These results implicate that conformational neutralizing epitope can be partially modeled by a short peptide, which provides a feasible route for subunit vaccine development.
INTRODUCTION
Hepatitis E is an acute hepatitis caused by hepatitis E virus (HEV) in developing countries, where it occurs sporadically and in an epidemic form. The causative agent, HEV, is transmitted primarily by the fecal-oral route, and it accounts for about 15%-20% sporadic cases of acute hepatitis in China[1,2]. HEV is an icosahedron non-enveloped virus, and its genome is a single-stranded positive-sense, 3’-polyadenylated RNA about 7.5 kb in length. It contains 3 open reading frames (ORFs). ORF1 codes for a polyprotein of 1693 amino acids and contains domains homologous to a viral methyltransferase, a papainlike cysteine protease, an RNA helicase, and an RNA-dependent RNA polymerase, and the most hypervariable region of HEV genome. ORF2 codes for the viral capsid protein of 660 amino acids, while ORF3 codes for a 123-amino-acid-long polypeptide with unknown function.
It has been proved that the major viral capsid protein encoded by ORF2 contains the protective epitope[3]. We recombined a fragment of HEV ORF2 and expressed the polypeptide named NE2 in E.coli recently[4], and proved it to be a protective antigen[5], which naturally forms homodimer by virtue of its interface domain[6]. Three anti-NE2 neutralization mAbs, 8C11, 8H3 and 13D8 were selected, which could recognize 2 separated neutralization conformational surface epitopes of HEV, 8C11 and 13D8 against one and 8H3 against the other. The 3 mAbs can neutralize HEV in vitro and in vivo test of Rhesus monkey[7-9].
8C11 and 8H3 were used to screen binding peptides from a heptapeptide phage display library, which could mimic the 2 neutralizing epitopes of HEV binding to 8C11 and 8H3 respectively. The obtained dominant peptides’ DNA sequences were then synthesized and cloned into amino acid 78 to 83 of hepatitis B core antigen (HBcAg), then expressed in E.coli. While it has been proven that HBcAg could be expressed in eukaryocytes or prokarocytes, and assembled into VLP automatically. The inserted sequence between amino acids 73 and 94 could be expressed on the HBcAg VLP surface[10]. Because HBcAg is an immunodominant immunogen[11], which results in the immunodominance of the insertion, we expressed the linear epitopes, which could conformationally mimic epitope on the surface of HBcAg, resulting in a VLP antigen, and prove its activity. This assay would not only mimic the neutralizing epitope of HEV but also provide insights into a novel route for subunit vaccine development.
MATERIALS AND METHODS
Monoclonal antibodies
8C11 and 8H3 mAbs, which could conformationally bind to neutralizing epitopes of HEV were prepared in our laboratory[7,9]. Affinity purified antibody phosphatase labeled goat anti-mouse IgG (H + L) was from Kirkegaard & Perry Laboratory (2 Cessua Court, Gaithersburg Maryland 20879, USA).
Affinity selection and amplification of peptide library
Heptapeptide phage display library and host strain E.coli ER2738 were purchased from New England BioLabs Company. Anti-HEV mAbs binding phages were isolated from the phage display library by successive cycles of selection and amplification. The biopanning procedure was essentially used as described in the phage display library user manual. Briefly, mAbs 8C11 and 8H3 (100 μg/mL) were coated on 96-well microtiter plates. After blocked with 20 g/L BSA, 100 μL diluted phage (10 uL of the original library) was piped onto the coated plate and shaked gently for 1 h at room temperature, the plates were washed for 10 times with PBST (10 g/L Tween 20). The bound phages were eluted with 100 μL 10 mmol/L Gly-Cl (pH 2.2), 1 mg/mL BSA, rocked gently for 10 min. The eluate was piped into a microcentrifuge tube and neutralized with 10 μL 1 mol/L Tris-HCl (pH 9.1). The eluate was added to 20 mL ER2738 culture and incubated at 37 °C with vigorous shaking for 4.5 h. The culture was transferred to a centrifuge tube and spun for 10 min at 10000 g at 4 °C. The supernatant was collected and 1/6 volume of PEG/NaCl (200 g/L PEG8000, 2.5 mol/L NaCl) was added. The phage was allowed to precipitate at 4 °C overnight. PEG precipitation was spun for 15 min at 10000 g at 4 °C. The pellet was re-suspended in 1 mL PBS, and re-precipitated with PEG/NaCl. Finally the pellet was suspended in 200 μL PBS, and stored at 4 °C. The amplified eluate was titered as general methods. The concentration of target mAbs was lowered to 50 μg/mL, at the same time the concentration of Tween-20 was risen to 50 g/L in the washing step, then was panned 3 more times by repeating the above steps. After 4 rounds panning, individual monoclones were sequenced. The single-stranded DNA of M13 phages was purified by using the M13 mini kit purchased from Shanghai Huashun Biotech Ltd, and then sequenced by Shanghai Boya Ltd.
Construction of recombinant expression vector for peptide
The recombinant expression vector named pC149-mut was composed in our laboratory. The peptide of amino acids 1 to 149 of HBcAg expressed in E.coli formed viral like particles (VLPs). The amino acid residues 78 to 83 were exposed at the VLP’s surface. According to this we cloned the HBcAg’s genome of amino acid residues 1 to 149 into the expression vector pTO-T7[12] of E.coli, and designed a linker to replace the immunodominant epitope located at 79 to 80aa of HBc and composed the mutant vector pC149-mut. After exogenous gene was inserted into the amino acid residues 78 to 83, recombinant C antigen was still able to form VLP and expose the exogenous epitope.
We designed the primer (Table 1) according to the dominant sequence of selected monoclonal phages, and the linker sequence of pC149-mut vector. Therefore, we could insert heptapeptide exogenome into the vector and co-express it with HBcAg using 8C11AFP or 8H3AFP as a former primer and 149 mutRP as a lower primer, and using the pC149-mut vector as a template. PCR was carried out after pre-denaturation at 94 °C for 5 min, 25 cycles of denaturation at 94 °C for 50 s, annealing at 56 °C for 50 s, and extension at 72 °C for 25 s, then extension at 72 °C for 10 min. The PCR fragment was cloned into pMD 18-T vector and formed positive clones containing the genome of peptide 8C11A (CATCCTACTCTTTTGCGTATT), which could bind to mAb 8C11 or the genome of peptide 8H3A (TCTATTCTGCCGTATCCTTAT), which could bind to mAb 8H3. Then they were digested with BamH I and EcoR I, and the product fragment was linked into the pC149-mut vector to compose the expression vector named pC149-mut-8C11A or pC149-mut-8H3A (Figure 1).
Table 1.
Primers for expression vector of mimic peptide
| Primer ID | Primer sequence |
| 8C11AFP | GGATCCCATCCTACTCTTTTGCGTATTGGTGGTGGAGGTTCAGG |
| 8H3AFP | GGATCCTCTATTCTGCCGTATCCTTATGGTGGTGGAGGTTCAGG |
| 149 mutRP | GAATTCTTAAACAACAGTAGTTT |
The underlined sequences are the insert genes coding the selected peptides.
Figure 1.
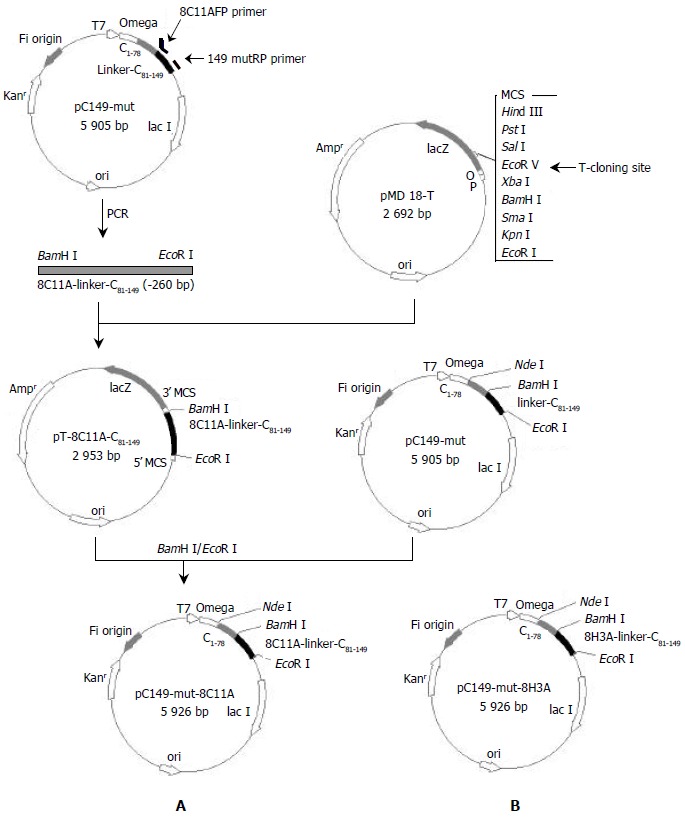
Expression vector construction of pC149-mut-8C11A and pC149-mut-8H3A. A: Construction of pC149-mut-8C11A, B: Map of pC149-mut-8H3A, which was constructed as pC149-mut-8C11A except using the primer 8H3AFP instead of 8C11AFP.
Expression and purification of recombinant polyprotein
Host stain ER2566 was cultured and transformed with the recombinant vector pC149-mut-8C11A or pC149-mut-8H3A in LB culture medium at 37 °C in a shaker, till the A600 reached 0.8-1.0, then the culture was incubated with 0.2 mmol/L IPTG for 6 h at 37 °C. The cells were spun to be collected and were disintegrated by an ultrasonic disintegrator (Uilbra-Cell VGX500 of SONICS & MATERIALS Co.). The insoluble protein was spun and collected, then suspended and deposited 2 times into 10 g/L Triton X-200. The pellet was finally suspended into buffer A (4 mol/L Urea, 20 mmol/L Tris-HCl pH8.5, 5 mmol/L EDTA, 100 mmol/L NaCl), dialysed in PBS overnight. The lysate was spun and collected.
Western blotting
Boiled supernatant of ultrasonic lysate and boiled deposition of ultrasonic lysate and control samples (the expression of pC149-mut could not react with mAbs, and the recombinant polyprotein NE2 was used to select mAbs including 8C11 and 8H3[7,9]) were dissolved in 120 g/L SDS-PAGE, transferred to a nitrocellulose membrance. Each contained one lane of the protein molecular mass mark between 18.3 and 215 ku. The membrane containing proteins with 5 g/L non-fat dry milk as blocking solution was incubated for 90 min and reacted with the test mAb 8H3 or 8C11 diluted in 5 g/L non-fat dry milk. The immunocomplexes were detected by using Ap-labeled goat-anti-mouse IgG, BCIP and NBT as substrates.
Transmission electron microscopy of chimeric VLPs
After renatured by the methods described above, the particles were adsorbed to carbon-coated grids, stained with 2 g/L uranyl acetate, and examined by JEM-100CXII electron microscope at about × 100000 magnification.
Active analysis of recombinant proteins by BIAcore biosensor
Heptapeptide named 8H3A (N’-Ser-Ile-Leu-Pro-Tyr-Pro-Tyr-C’) was synthesized and purified by Xi’an Meilian Ltd (Xi’an, China). Then the biosensor chip was coupled with anti-mouse Fc, which was carried out according to the user manual. Briefly, the sensor chip CM5 with CM Dextran on its reaction-surface was activated with EDC and NHS, and then the flow cells were treated with a standard amine-coupling reagent (injecting 10 mm/L NaOAc, pH4.8, instead of protein) or with amine-coupling reagents and anti-mouse Fc polyclonal antibody. The coupling density of the antibody was 900 RU. Experiments were run at 25 °C. After mAb 8H3 was bound to anti-mouse Fc antibody, peptide 8H3A was injected. The data of association and dissociation indicating the reaction between mAb 8H3 and peptide 8H3A were collected. Biacore X biosensor, CM5 sensor chip and reagent of EDC & NHS were from Amersham Biosciences Company (Uppsala, Sweden).
RESULTS
Sequence characteristics of peptide binding to neutralizing mAb 8C11 or 8H3
After 4 rounds of biopanning, 21 monoclonal phages were selected for sequencing (in which 10 phage could bind to mAb 8C11, and the other 11 could bind to mAb 8H3). According to the DNA sequence, the peptide sequence of the phage displayed was made out (Figure 2). Heptapeptide with amino acid sequence HPTLLRI was dominant (50%) in the 10 monoclones bound to mAb 8C11. While in the 11 monoclones bound to mAb 8H3, the preponderant amino acid sequence was SILPYPY (27.3%). Furthermore, in the heptapeptide binding to mAb 8H3, the amino acid sequence of S*LP, S*P, S*L or LP was also more frequent, indicating that S, L and P would be more important to form the domain binding to mAb 8H3. Likewise, amino acids P, T and L were found to be more important in the interaction between peptide and mAb 8C11.
Figure 2.

Sequences of peptides selected by monoclonal antibodies. A: Selected by mAb 8C11, B: Selected by mAb 8H3.
Expression and western blotting of peptide mimic neutralization epitope
Dominant peptides 8C11A (N’-His-Pro-Thr-Leu-Leu-Arg-Ile-C’) and 8H3A (N’-Ser-Ile-Leu-Pro-Tyr-Pro-Tyr-C’) were cloned into the vector pC149-mut named C8C11A and C8H3A respectively, and expressed. The result of expression was shown in Figure 3. Recombinant polyprotein C8C11A was found in the insoluble cell pellet, and formed dimer (about Mr40000 in mass) on the SDS-PAGE mainly while the monomer (about Mr20000) was hard to be seen. As to the Western blotting result, it was the dimer that could interact with mAb 8C11 in the same level as the monomer of NE2 protein (about Mr29000). C8H3A fusions were also found in the insoluble cell pellet. Beside the dimer, a monomer was in the majority, but only the -Mr40000 dimer was able to bind to mAb 8H3 when tested by Western blotting.
Figure 3.
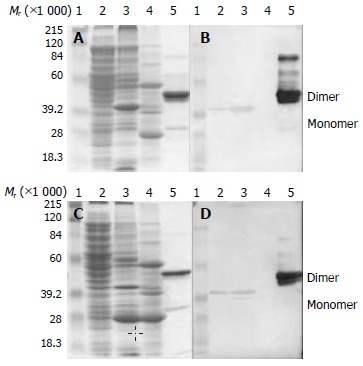
Expression and Western blotting of pC149-mut-8C11A and pC149-mut-8H3A. A, C: SDS-PAGE, B, D: Western blot-ting of mAbs 8C11 and 8H3 respectively, A, B: pC149-mut-8C11A, C, D: pC149-mut-8H3A. 1, Protein molecular weight marker; 2, Supernatant of ultrasonic lysate; 3, Deposition of ultrasonic lysate; 4, C149-mut control; 5, Purified NE2 antigen
Electron micrograph of VLP
The renatured recombinant polyprotein particles were adsorbed to carbon-coated grids, stained with 20 g/L uranyl acetate, and examined with TEM. The recombinant HBcAg with 8C11A or 8H3A on its surface and HBcAg could form VLPs and VLPs’ diameter was about 20 nm (Figure 4).
Figure 4.
Virus like particles assemblied by recombinant protein C8C11A, C8H3A or C149 mut (Negative staining electron microscopy, × 100000).
Interaction between chemo-synthesized heptapeptide 8H3A and mAb 8H3
The mAb 8H3 was bound to the anti-mouse Fc polyclonal antibody coupled on the cells of CM5 chip. After mAb 8H3 was bound to the anti-mouse Fc antibody, peptide 8H3A (1 mg/mL) was injected and the data of association and dissociation were collected. Chemo-synthesized heptapeptide 8H3A could bind to mAb 8H3, but not stably (Figure 5).
Figure 5.
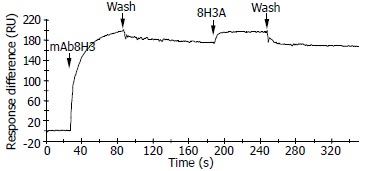
Binding curve of chemo-synthesized seven peptide 8H3A against monoclonal antibody 8H3 in BIAcore biosensor.
DISCUSSION
Phage display technique has been widely used in constructing antibody library or biopanning of random peptides according to their biological activities, and in many other fields or so[13,14]. As a new technique, it makes use of the characteristics of the coat protein of the phage, that is, a peptide is expressed as a fusion with the phage coat protein pIII, resulting in the display of fused protein on the surface of virion. Phage display technique allows rapid identification and amplification of the peptide ligands for target molecules by the affinity of their specifical-binding. Recently, random peptide libraries displayed on phage have been widely used in many fields, such as identification and analysis of a peptide mimic the epitope of HIV[15] and HIV-1-infected cells[16], identification of enzyme inhibitors[17] and immunodepressants[18], and many antigens etc.[19-22]. Phage display technique is specially useful to mimic a variety of half-antigens and non-peptide target molecules’ function, which is a very useful way to produce mimic peptide medication. Many reports have been available on the peptides mimicing epitope, which can neutralize Neisseria meningitidis[23,24] and many other bacteria to produce bacterial vaccines.
There are still no cell models of HEV, or effective methods to produce HEV artificially. Different sections of HEV capsid protein were expressed, and the structures of the recombinant polyproteins were analysed, which can help us to speculate the crude structure of HEV capsid. We cloned a section of HEV ORF2 (named NE2) and expressed this recombinant protein in E.coli. It was found that NE2 formed polymer spontaneously, from dimer to hexamer concretely. This was very similar to the assemble process of HEV capsid protein[4,25,26]. NE2 was used to immunize Rhesus monkey, and induce protective antibodies. The above findings indicate that polyprotein NE2 mimics the crude structure of HEV ORF2[4]. Monoclonal antibodies were selected by immunizing BALB/c mice with NE2[7,9], 3 of which named 8C11, 8H3 and 13D8 were identified to be mAbs which could recognize 2 different epitopes of HEV. These 3 mAbs were proved to be neutralizing mAb by in vivo neutralization test in rhesus. The 2 epitopes recognized by mAbs (1 bound to 8C11 and 13D8, and the other to 8H3) were tested to be conformational epitopes. We panned the random phage display heptapeptide library, which displayed the linear eptiope constructed by heptapeptide, and the peptides could mimic the neutralizing eptiope that could bind to the 2 mAbs (8C11 and 8H3). Furthermore, heptapeptides were recombined on HBcAg for expression in E.coli, and their activities were tested by Western blotting. The recombinant polyprotein expressed by plasmid pC149-mut-8C11A appeared to be the dimer of Mr40000 on SDS-PAGE, which could bind to mAb 8C11 similar to the monomer of NE2. While the production of plasmid pC149-mut-8H3A could form monomer (Mr20000) and dimer (Mr40000) on SDS-PAGE, and only the dimer could react with mAb on Western blotting. Maybe it is because that 7 amino acid peptide is too short, and to display it on the surface of recombinant HBcAg is in need of assistance of special conformation, HBcAg is a large protein while the monomer recombinant HBcAg can not display the exogenesis epitope of heptapeptide, so the activity of the monomer is not obvious. The chemo-synthesized heptapeptide 8H3A could bind to mAb 8H3, but the binding was not stable. Comparing the dominant peptides’ sequence with the primary structure of NE2, no same sequence was found, conforming that the epitopes recognized by mAbs 8C11 and 8H3 are not constructed by continuous amino acid of NE2, but the function of their spatial structure is similar to the mimic peptides.
The immunogenicity of HBcAg is the highest in all kinds of antigens of HBV. The design of vector pC149-mut was to insert foreign genes into amino acids 78 to 83[10], the immunogenicity of that position was better. The inserted amino acid could replace the epitope that reacts with B cells easily, thus reducing their interference with the exogenous epitope in immune response. This feature made HBcAg to be a very useful vector for multi-epitope chimeric vaccine[10,27], so did the pC149-mut vector, which was designed accordingly. In this study, heptapeptide was cloned into pC149-mut between the amino acids 78 to 83 of HBcAg, and the recombinant polyprotein could form VLP, which displayed the foreign heptapeptide on their surface and kept its immunogenicity. The known neutralizing epitopes of viruses are almost conformational epitopes, so the genetically engineered vaccine needs the recombinant protein to be in correct conformation, and be able to preserve the immunity. These 2 qualifications could result in a mass of genetically engineered screen. But if using the peptides which mimic the conformational neutralizing epitopes, using the plasmid pC149-mut as a vector to mimic peptide vaccine expression will get VLP antigens, which remain the activation of neutralizing epitopes, and ensure the immunity of the vaccine, at the same time result in protective immunity. On the one hand, this tactics makes it available to unite more than one kind of virus’s neutralizing epitopes on the VLP antigen. That is to say, it is a way to develop polyvalent vaccines, which shows new direction in the development of new vaccine[28-31]. The preventer of polyvalent vaccine is that the neutralizing epitopes are almost conformational, if in term of the routine method the conformational epitope cannot be exactly formed. On the other hand, if the chemo-synthesized peptide is used, the immunity will be influenced because of the low molecular mass of peptide. In a word, using the recombinant vector to express the protein, which can form VLP, and using the linear functional mimic epitope, which is displayed on the surface of VLP antigen instead of the conformational neutralizing epitope, is a new feasible idea to settle the difficulties in polyvalent vaccines.
Footnotes
Supported by Grant from Science and Technology Projects of Fujian Province, China; No.2002 F013; Excellent Scholar Incubation Plan of the Ministry of Education, China
Edited by Xu FM and Wang XL
References
- 1.Zhuang H, Cao XY, Liu CB, Wang GM. Epidemiology of hepatitis E in China. Gastroenterol Jpn. 1991;26 Suppl 3:135–138. doi: 10.1007/BF02779283. [DOI] [PubMed] [Google Scholar]
- 2.Huang RT, Li XY, Xia XB, Yuan XT, Liu MX, Li DR. Antibody detection and sequence analysis of sporadic HEV in Xiamen region. World J Gastroenterol. 1999;5:270–272. doi: 10.3748/wjg.v5.i3.270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Purcell RH, Emerson SU. Hepatitis E virus In Knipe DM, Howley PM, Griffin DE, eds. Fields virology, 4thed. vol2. Philadelphia: Lippincott Raven Pub; 2001. pp. 3051–3061. [Google Scholar]
- 4.Li SW, Zhang J, He ZQ, Ge SX, Gu Y, Lin J, Liu RS, Xia NS. [The study of aggregate of the ORF2 peptide of hepatitis E virus expressed in Escherichia coli] Shengwu Gongcheng Xuebao. 2002;18:463–467. [PubMed] [Google Scholar]
- 5.Ge S, Zhang J, Huang G, Pang S, Zhou K, Xia N. [The immuno-protect study of a hepatitis E virus ORF2 peptide expressed in E. coli] Weishengwu Xuebao. 2003;43:35–42. [PubMed] [Google Scholar]
- 6.Li SW, He ZQ, Wang YB, Chen YX, Liu RS, Lin J, Gu Y, Zhang J, Xia NS. [Interface domain of hepatitis E virus capsid protein homodimer] Shengwu Gongcheng Xuebao. 2004;20:90–98. [PubMed] [Google Scholar]
- 7.Gu Y, Zhang J, Li SW, Ge SX, He ZQ, Zhu ZH, Xian YL, Li YM, Xia NS. Characterazation of the anti-HEV ORF2 monoclonal antibodies by biosensor. Xibao Yu Fenzi Mianyixue Zazhi. 2002;18:617–620. [Google Scholar]
- 8.Ge SX, Zhang J, Peng G, Huang GY, He ZQ, Gu Y, Zhu ZH, Ng MH, Xia NS. Development and evaluation of ELISAs for antihepatitis E virus IgM and IgG detection based on polmerized recombinant antigen. Bingdu Xuebao. 2003;19:78–86. [Google Scholar]
- 9.Gu Y, Ge SX, Huang GY, Li SW, Zhu ZH, He ZQ, Chen YX, Wang YB, Zhang J, Xia NS. Identification of neutralizing monoclonal antibodies to the hepatitis E virus. Bingdu Xuebao. 2003;19:217–223. [Google Scholar]
- 10.Böttcher B, Wynne SA, Crowther RA. Determination of the fold of the core protein of hepatitis B virus by electron cryomicroscopy. Nature. 1997;386:88–91. doi: 10.1038/386088a0. [DOI] [PubMed] [Google Scholar]
- 11.Ding CL, Yao K, Zhang TT, Zhou F, Xu L, Xu JY. Generation of cytotoxic T cell against HBcAg using retrovirally transduced dendritic cells. World J Gastroenterol. 2003;9:1512–1515. doi: 10.3748/wjg.v9.i7.1512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Luo WX, Zhang J, Yang HJ, Li SW, Xie XY, Pang SQ, Li SJ, Xia NS. [Construction and application of an Escherichia coli high effective expression vector with an enhancer] Shengwu Gongcheng Xuebao. 2000;16:578–581. [PubMed] [Google Scholar]
- 13.Yu ZC, Ding J, Nie YZ, Fan DM, Zhang XY. Preparation of single chain variable fragment of MG(7) mAb by phage display technology. World J Gastroenterol. 2001;7:510–514. doi: 10.3748/wjg.v7.i4.510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wu BP, Xiao B, Wan TM, Zhang YL, Zhang ZS, Zhou DY, Lai ZS, Gao CF. Construction and selection of the natural immune Fab antibody phage display library from patients with colorectal cancer. World J Gastroenterol. 2001;7:811–815. doi: 10.3748/wjg.v7.i6.811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ferrer M, Sullivan BJ, Godbout KL, Burke E, Stump HS, Godoy J, Golden A, Profy AT, van Schravendijk MR. Structural and functional characterization of an epitope in the conserved C-terminal region of HIV-1 gp120. J Pept Res. 1999;54:32–42. doi: 10.1034/j.1399-3011.1999.00082.x. [DOI] [PubMed] [Google Scholar]
- 16.MacDonald NJ, Shivers WY, Narum DL, Plum SM, Wingard JN, Fuhrmann SR, Liang H, Holland-Linn J, Chen DH, Sim BK. Endostatin binds tropomyosin. A potential modulator of the antitumor activity of endostatin. J Biol Chem. 2001;276:25190–25196. doi: 10.1074/jbc.M100743200. [DOI] [PubMed] [Google Scholar]
- 17.Kiczak L, Kasztura M, Koscielska-Kasprzak K, Dadlez M, Otlewski J. Selection of potent chymotrypsin and elastase inhibitors from M13 phage library of basic pancreatic trypsin inhibitor (BPTI) Biochim Biophys Acta. 2001;1550:153–163. doi: 10.1016/s0167-4838(01)00282-5. [DOI] [PubMed] [Google Scholar]
- 18.Aramburu J, Yaffe MB, López-Rodríguez C, Cantley LC, Hogan PG, Rao A. Affinity-driven peptide selection of an NFAT inhibitor more selective than cyclosporin A. Science. 1999;285:2129–2133. doi: 10.1126/science.285.5436.2129. [DOI] [PubMed] [Google Scholar]
- 19.Shchelkunov SN, Nesterov AE, Ryazankin IA, Ignat'ev GM, Sandakhchiev LS. Development of a candidate polyvalent live vaccine against human immunodeficiency, hepatitis B, and orthopox viruses. Dokl Biochem Biophys. 2003;390:180–183. doi: 10.1023/a:1024476626759. [DOI] [PubMed] [Google Scholar]
- 20.Ragupathi G, Livingston P. The case for polyvalent cancer vaccines that induce antibodies. Expert Rev Vaccines. 2002;1:193–206. doi: 10.1586/14760584.1.2.193. [DOI] [PubMed] [Google Scholar]
- 21.Cardó-Vila M, Arap W, Pasqualini R. Alpha v beta 5 integrin-dependent programmed cell death triggered by a peptide mimic of annexin V. Mol Cell. 2003;11:1151–1162. doi: 10.1016/s1097-2765(03)00138-2. [DOI] [PubMed] [Google Scholar]
- 22.Xu L, Jin BQ, Fan DM. Selection and identification of mimic epitopes for gastric cancer-associated antigen MG7 Ag. Mol Cancer Ther. 2003;2:301–306. doi: 10.4161/cbt.2.3.386. [DOI] [PubMed] [Google Scholar]
- 23.Grothaus MC, Srivastava N, Smithson SL, Kieber-Emmons T, Williams DB, Carlone GM, Westerink MA. Selection of an immunogenic peptide mimic of the capsular polysaccharide of Neisseria meningitidis serogroup A using a peptide display library. Vaccine. 2000;18:1253–1263. doi: 10.1016/s0264-410x(99)00390-4. [DOI] [PubMed] [Google Scholar]
- 24.Prinz DM, Smithson SL, Westerink MA. Two different methods result in the selection of peptides that induce a protective antibody response to Neisseria meningitidis serogroup C. J Immunol Methods. 2004;285:1–14. doi: 10.1016/j.jim.2003.08.005. [DOI] [PubMed] [Google Scholar]
- 25.Zhang JZ, Ng MH, Xia NS, Lau SH, Che XY, Chau TN, Lai ST, Im SW. Conformational antigenic determinants generated by interactions between a bacterially expressed recombinant peptide of the hepatitis E virus structural protein. J Med Virol. 2001;64:125–132. doi: 10.1002/jmv.1027. [DOI] [PubMed] [Google Scholar]
- 26.Im SW, Zhang JZ, Zhuang H, Che XY, Zhu WF, Xu GM, Li K, Xia NS, Ng MH. A bacterially expressed peptide prevents experimental infection of primates by the hepatitis E virus. Vaccine. 2001;19:3726–3732. doi: 10.1016/s0264-410x(01)00100-1. [DOI] [PubMed] [Google Scholar]
- 27.Pumpens P, Razanskas R, Pushko P, Renhof R, Gusars I, Skrastina D, Ose V, Borisova G, Sominskaya I, Petrovskis I, et al. Evaluation of HBs, HBc, and frCP virus-like particles for expression of human papillomavirus 16 E7 oncoprotein epitopes. Intervirology. 2002;45:24–32. doi: 10.1159/000050084. [DOI] [PubMed] [Google Scholar]
- 28.Sompuram SR, Kodela V, Ramanathan H, Wescott C, Radcliffe G, Bogen SA. Synthetic peptides identified from phage-displayed combinatorial libraries as immunodiagnostic assay surrogate quality-control targets. Clin Chem. 2002;48:410–420. [PubMed] [Google Scholar]
- 29.Poloni F, Puddu P, Moretti F, Flego M, Romagnoli G, Tombesi M, Capone I, Chersi A, Felici F, Cianfriglia M. Identification of a LFA-1 region involved in the HIV-1-induced syncytia formation through phage-display technology. Eur J Immunol. 2001;31:57–63. doi: 10.1002/1521-4141(200101)31:1<57::aid-immu57>3.0.co;2-9. [DOI] [PubMed] [Google Scholar]
- 30.Li BW, Rush A, Zhang SR, Curtis KC, Weil GJ. Antibody responses to Brugia malayi antigens induced by DNA vaccination. Filaria J. 2004;3:1. doi: 10.1186/1475-2883-3-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Combredet C, Labrousse V, Mollet L, Lorin C, Delebecque F, Hurtrel B, McClure H, Feinberg MB, Brahic M, Tangy F. A molecularly cloned Schwarz strain of measles virus vaccine induces strong immune responses in macaques and transgenic mice. J Virol. 2003;77:11546–11554. doi: 10.1128/JVI.77.21.11546-11554.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]