

Abstract
Background
Expression of kisspeptin (protein) and Kiss1r (mRNA) was recently documented in the mouse uterus on D4 of pregnancy (the day of embryo implantation) suggesting that the uterine-based kisspeptin (KP)/kisspeptin receptor (KISS1R) signaling system regulates embryo implantation. Despite this important suggestion, it was never demonstrated that the uterus actually exhibits a functional KP/KISS1R signaling system on D4 of pregnancy. Thus, the goal of this study was to determine whether a functional KP/KISS1R signaling system exists in the mouse uterus on D4 of pregnancy.
Findings
Since kisspeptin/KISS1R signaling triggers the phosphorylation of the mitogen-activated protein kinases p38 and ERK1/2, through immunohistochemical analyses, we determined whether exogenously administered kisspeptin could trigger p38 and ERK1/2 phosphorylation in the uterus on D4 of pregnancy. The results clearly demonstrated that kisspeptin could and that its effects were mediated via KISS1R. Additionally, the robust kisspeptin-triggered response was observed in the pregnant uterus only. Finally, it was demonstrated that on D4 of pregnancy the Kiss1 null uterus expresses functional KISS1R molecules capable of mediating the effects of kisspeptin.
Conclusions
These results lead us to conclude that on D4 of pregnancy, the mouse uterus expresses a functional KP/KISS1R signaling system strengthening the possibility that this signaling system regulates embryo implantation. These findings strengthen the rationale for determining whether such a functional system exists in the uterus of the human female and if so, what role it might play in human pregnancy.
Keywords: KISS1R, Kisspeptin, Uterus, Embryo, Implantation, p38, ERK1/2, SAPK/JNK, MAPK
Introduction
It is now well-established that hypothalamic kisspeptins (KPs), signaling via their cognate receptor KISS1R, are potent triggers of gonadotropin-releasing hormone (GnRH) secretion and mice bearing a deletion of the gene encoding KISS1R in the GnRH neuron display hypogonadotropic hypogonadism and infertility [1, 2]. Therefore, the hypothalamic-based KP/KISS1R signaling system is considered a major regulator of reproduction [3]. The KP/KISS1R signaling system is also expressed in the female reproductive tract and roles have been proposed in positively regulating follicular development, ovulation, embryo implantation and endometrial decidualization [4–7]. Therefore, the peripheral KP/KISS1R signaling system is considered a major regulator of reproduction [8].
We have previously reported that KP (protein) and Kiss1r (mRNA) are highly expressed in the mouse uterus on the day of implantation (that is, D4 after mating with the day of mating designated D0) suggesting a role for this signaling system in regulating embryo implantation [4]. To test this idea further, we turned our attention to mice bearing a global deletion of Kiss1. These mice are infertile and it was assumed that their infertility was due to a lack of hypothalamic KP/KISS1R signaling and the resulting reduction in gonadotropins, estradiol and progesterone [9]. However, despite the add-back of these hormones embryos failed to implant in the Kiss1 null uterus leading us to suggest that it was a lack of uterine and not hypothalamic KP/KISS1R signaling that triggered implantation failure. Despite this important suggestion, we did not actually demonstrate the presence of a functional KP/KISS1R signaling system in the uterus on D4 of pregnancy. Such demonstration would have greatly strengthened our suggestion.
Thus, the goal of this study was to determine whether on D4 of pregnancy, the day of embryo implantation, a functional KP/KISS1R signaling system exists in the mouse uterus. It was previously reported that in various cell types, the KP/KISS1R signaling triggers mitogen-activated protein kinase (MAPK) phosphorylation. Specifically, it was demonstrated that KP treatment of KISS1R-expressing cells triggers the phosphorylation of the MAPKs p38 and ERK1/2 [10–12] but not SAPK/JNK [10]. We therefore investigated whether exogenously administered KP could trigger the phosphorylation of MAPKs in the uterus.
Findings
In the following studies 8–12 week old 129S1/SvImJ female mice were used. WT females and estradiol- and gonadotropin-primed Kiss1 null females (as previously described [4]) were mated to WT males (D0) and on the morning of D4 of pregnancy, females which displayed a copulatory plug (evidence of successful mating) were injected with either phosphate-buffered saline (PBS, 100 μl) or KP54 (100 nmol/kg, 100 μl, Tocris Bioscience, Minneapolis, MN, USA) intraperitoneally and 30 min later were rapidly sacrificed. In addition to the use of these pregnant mice, age-matched WT unmated but cycling females were also treated with PBS or KP54 in an identical manner. All experiments were repeated 6 independent times.
Immediately after drug or saline treatment, uteri were removed and simultaneously processed for immunohistochemistry using standard techniques [4] and 5 μm-thick slices were simultaneously immunostained to detect phosphorylated p38, ERK1/2 and SAPK/JNK. Care was taken to expose the experimental and control tissue to identical experimental conditions. Phosphorylated p38, pERK1/2 and SAPK/JNK were assayed using the following rabbit monoclonal antibodies: phospho-p38 (D3F9), phospho-p44/42 (ERK1/2) (D13.14.4E) and phospho-SAPK/JNK (81E11) (Cell Signaling, Boston, MA, USA). Antigen-bound primary antibodies were detected with the ImmunoCruz rabbit ABC Staining System sc-2018 (Santa Cruz Biotechnology, Inc. Dallas, TX, USA). Both the primary and secondary detection systems were used according to the manufacturers’ guidelines without any adaptations.
KP triggers p38 phosphorylation in the uterine luminal and glandular epithelia on D4 of pregnancy
Results showed that, compared to PBS-treated pregnant mice (Fig. 1a and b), uteri from KP54-treated pregnant mice exhibited (approximately 5-10-fold) higher levels of phosphorylated p38 in all cells of both the luminal and glandular epithelia (Fig. 1c, d and e). These findings were recapitulated when uteri were removed from untreated pregnant mice and then treated with either PBS or KP54 in vitro in the absence of any effects that KP54 might have triggered in vivo at the level of the hypothalamus, pituitary and ovaries (data not shown).
Fig. 1.
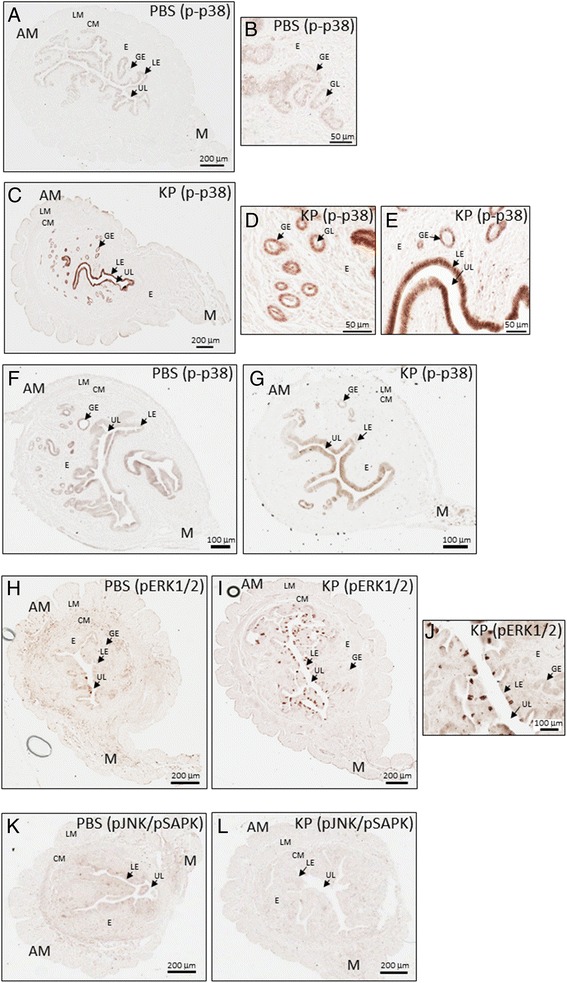
KP54 administration to female WT mice on D4 of pregnancy triggers p38 and ERK1/2 phosphorylation but not SAPK/JNK. Phosphorylated p38 expression in the D4 pregnant uterus following PBS-treatment (a); part of the uterus shown in (a) is magnified to better reveal details on the endometrial glands (b). Posphorylated p38 (p-p38) expression in the D4 pregnant uterus following KP54-treatment (c); parts of the uterus shown in (c) are magnified to better reveal details on the endometrial glands and luminal epithelium (d and e). Phosphorylated p38 expression in the non-pregnant uterus following PBS- (f) or KP54 (g) treatment. Phosphorylated ERK1/2 (pERK1/2) expression in the D4 pregnant uterus following PBS-treatment (h) or KP54-treatment (i); part of the uterus shown in (i) is magnified to better reveal details on the luminal epithelium (j). Phosphorylated SAPK/JNK (pSAPK/pJNK) expression in the D4 pregnant uterus following PBS-treatment (k) or KP54-treatment (l). AM: anti-mesometrial pole; LM: longitudinal muscle of the myometrium; CM: circular muscle of the myometrium; E: endometrium; LE: luminal epithelium; GE: glandular epithelium; M: mesometrial pole; UL: uterine lumen
KP barely triggers p38 phosphorylation in the non-pregnant uterus
When these studies were conducted in non-pregnant mice, relative to PBS-treated mice (Fig. 1f), uteri from KP54-treated mice (Fig. 1g) failed to show a strong increase in phosphorylated p38 in any cell type. However, careful examination of several uteri did suggest there was a very weak (approximately 1.2-fold greater) but almost undetectable response in the uterine epithelium. This suggest that KISS1R is weakly expressed in the non-pregnant uterus and/or poorly coupled to p38 phosphorylation. The weak expression suggests the possibility that basal kisspeptin signaling might also play a role in the non-pregnant uterus or that basal expression keeps the uterus in a primed-stated such that it allows for a more rapid upregulation of signaling when it is required during pregnancy.
KP triggers ERK1/2 phosphorylation in the uterine luminal epithelium on D4 of pregnancy
When compared to PBS-treated pregnant mice (Fig. 1h), uteri from KP54-treated pregnant mice exhibited visibly higher levels of phosphorylated ERK1/2 in a subset of cells within the luminal epithelium only (Fig. 1i and j). When this study was conducted in non-pregnant mice, relative to PBS-treated mice, uteri from KP54 treated mice failed to show a visible increase in phosphorylated ERK1/2 in any cell type (data not shown) again supporting the idea that KISS1R is very weakly expressed in the non-pregnant uterus and/or uncoupled to ERK1/2 phosphorylation.
KP fails to trigger SAPK/JNK phosphorylation in the uterus
Consistent with data obtained in KISS1R-expressing Chinese hamster ovary cells [10], relative to PBS-treated pregnant mice (Fig. 1k), uteri from KP54-treated pregnant mice (Fig. 1l) failed to show an increase in phosphorylated SAPK/JNK in any cell type. Failure to detect evidence of SAPK/JNK phosphorylation was not due to a loss in antibody quality since an aliquot of this antibody detected phosphorylated SAPK/JNK in UV-treated human embryonic kidney 293 cells (data not shown).
KP triggers p38 phosphorylation in a KISS1R-specific manner in the uterus on D4 of pregnancy
It was reported that in addition to KISS1R, KP can signal via the NPFF1/GPR147 and NPFF2/GPR74 receptors [13]. Therefore, to determine whether the KP-triggered phosphorylation of p38 and ERK1/2 was KISS1R-specific the following experiment was conducted. Eight to 12 week old WT females were mated to WT males and on the morning of D4 of pregnancy, females were injected intraperitoneally with either PBS (100 μl) or the KISS1R antagonist P234 (100 nmol/kg, 100 μl, Tocris Bioscience, Minneapolis, MN, USA) [14]. Thirty minutes later a group of PBS-treated females was injected with either PBS (100 μl) or KP54 (100 nmol/kg, 100 μl) while the P234-treated group was injected with KP54 (100 nmol/kg, 100 μl). Thirty minutes later, mice were sacrificed and uteri were processed to assay for phosphorylated p38.
As expected, based on earlier findings (Fig. 1a-e) results revealed that relative to PBS-treated pregnant mice (Fig. 2a and b), KP-triggered a robust increase in phosphorylated p38 levels in both the luminal and glandular epithelia of pregnant uteri (Fig. 2c and d), however, this KP-triggered increase was completely blocked when mice were pretreated with P234 (Fig. 2e and f). Interestingly, in P234 pretreated mice, phosphorylated p38 levels were visibly lower (by approximately 2-fold) than that of PBS only treated mice (Fig. 2e and f vs. 2a and b) suggesting that on the day of implantation, endogenous KP phosphorylates uterine p38. Based on preliminary data (n = 2), similar observations were made with pERK1/2 levels at the luminal epithelium and again, consistent with earlier experiments, KP54 had no effect on JNK/SAPK phosphorylated levels both in the presence and absence of P234 pre-treatment (n = 2) (data not shown).
Fig. 2.
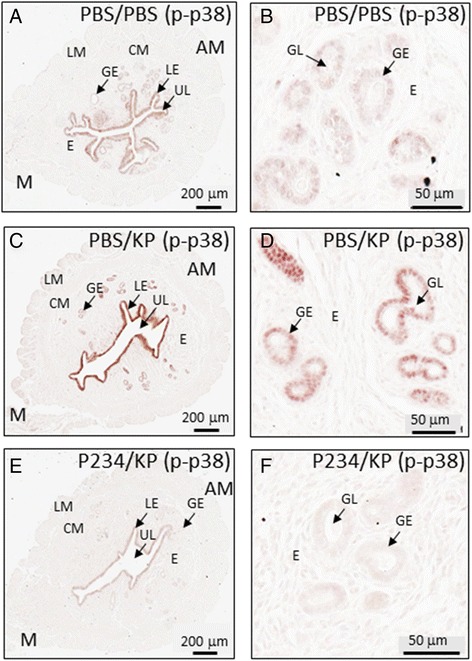
KP signals specifically via KISS1R in the uterus on D4 of pregnancy in WT mice. Phosphorylated p38 expression in the D4 pregnant uterus following 30 min PBS pre-treatment followed by 30 min PBS post-treatment (a); part of the uterus shown in (a) is magnified to better reveal details on the endometrial glands (b). Phosphorylated p38 expression in the D4 pregnant uterus following 30 min PBS pre-treatment followed by 30 min KP54 post-treatment (c); part of the uterus shown in (c) is magnified to better reveal details on the endometrial glands (d). Phosphorylated p38 expression in the D4 pregnant uterus following 30 min P234 pre-treatment followed by 30 min KP54 post-treatment (e); part of the uterus shown in (e) is magnified to better reveal details on the endometrial glands (f). AM: anti-mesometrial pole; LM: longitudinal muscle of the myometrium; CM: circular muscle of the myometrium; E: endometrium; LE: luminal epithelium; GE: glandular epithelium; M: mesometrial pole; UL: uterine lumen
The Kiss1−/− uterus remains responsive to KP
In a previous study we failed to rescue the implantation defect through the add-back of KP to pregnant Kiss1−/− females [4]. We speculated that this could have been due to the inefficient delivery of the drug (which was administered subcutaneously) to the uterus [15]. Another possibility was that although Kiss1r mRNA was still expressed in the Kiss1−/− uterus, this expression did not result in the formation of functional receptor molecules thus precluding the ability of KP to signal and rescue the implantation defect. To test the latter possibility, Kiss1−/− uteri on D4 of pregnancy were assayed for phosphorylated p38 following KP54 treatment. Results showed that relative to PBS-treated pregnant mice (Fig. 3a), uteri from KP54-treated pregnant mice exhibited higher levels of phosphorylated p38 in all cells of both the luminal and glandular epithelia (Fig. 3b). Based on preliminary data (n = 2), similar observations were made with pERK1/2 levels at the luminal epithelium and again, consistent with earlier experiments, KP54 had no effect on JNK/SAPK phosphorylated levels (n = 2) (data not shown). Together, these results confirm the presence of functional KISS1R molecules coupled to p38 and ERK1/2 phosphorylation on D4 of pregnancy in the Kiss1−/− uterus suggesting that failure of exogenous KP to rescue the implantation defect, as reported by Calder et al. [4] was not due to the lack of functional KISS1R molecules in the uterus.
Fig. 3.
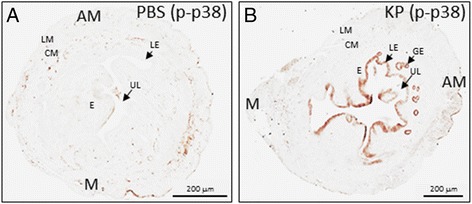
KP54 administration to female Kiss1 −/− mice on D4 of pregnancy triggers p38 phosphorylation. Phosphorylated p38 expression in the D4 pregnant uterus following PBS-treatment (a) or KP54-treatment (b). AM: anti-mesometrial pole; LM: longitudinal muscle of the myometrium; CM: circular muscle of the myometrium; E: endometrium; LE: luminal epithelium; GE: glandular epithelium; M: mesometrial pole; UL: uterine lumen
Conclusions
The goal of this study was to determine whether on D4 of pregnancy, the day of embryo implantation, a functional KP/KISS1R signaling system exists in the mouse uterus. Our data clearly reveal that one exists and this major finding further strengthens the possibility that this signaling system regulates uterine functions in early pregnancy. Because expression, as determined by phosphorylated p38 and ERK1/2 levels was localized to the uterine epithelia, it is possible that KP/KISS1R signaling regulates embryo implantation. For example, through p38 it might regulate the availability (production and/or secretion) of glandular products essential for implantation, or through both p38 and ERK1/2 might regulate embryo implantation at the uterine epithelium. It is also possible that through paracrine signaling, epithelial KP/KISS1R signaling prepares stromal cells for decidualization in the event of successful implantation [7, 16].
Acknowledgements
Dr. Babwah’s research is supported by the Natural Sciences and Engineering Research Council of Canada RGPIN/327334-2011. Ethics approval: Animal studies were approved by the University of Western Ontario Animal Care Committee according to guidelines established by the Canadian Council on Animal Care (Protocol Number: 2008-017-03).
Abbreviations
- KP
Kisspeptin
- KISS1R
Kisspeptin receptor
- GnRH
Gonadotropin-releasing hormone
- MAPK
Mitogen-activated protein kinase
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
MF and MC: conducted the experiments described in the manuscript. MB, GAV, SP and AB: helped draft the manuscript. AB: conceived and designed the study and is the senior corresponding author. All authors read and approved the final manuscript.
Contributor Information
Mehri Fayazi, Email: mehr.fayazi@gmail.com.
Michele Calder, Email: michele.calder@schulich.uwo.ca.
Moshmi Bhattacharya, Email: mbhattac@uwo.ca.
George A. Vilos, Email: george.vilos@lhsc.on.ca
Stephen Power, Email: stephen.power@lhsc.on.ca.
Andy V. Babwah, Phone: 519-685-8500, Email: ababwah@uwo.ca
References
- 1.Kirilov M, Clarkson J, Liu X, Roa J, Campos P, Porteous R, Schutz G, Herbison AE. Dependence of fertility on kisspeptin-Gpr54 signaling at the GnRH neuron. Nat Commun. 2013;4:2492. doi: 10.1038/ncomms3492. [DOI] [PubMed] [Google Scholar]
- 2.Novaira HJ, Momodou LS, Hoffman G, Koo H, Ko C, Wolfe A, Radovick S. Disrupted Kisspeptin Signaling in GnRH Neurons Leads to Hypogonadotrophic Hypogonadism. Mol Endocrinol. 2014;28:225–38. doi: 10.1210/me.2013-1319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Millar RP, Babwah AV. KISS1R: Hallmarks of an Effective Regulator of the Neuroendocrine Axis. Neuroendocrinology. 2015;101:193–210. doi: 10.1159/000381457. [DOI] [PubMed] [Google Scholar]
- 4.Calder M, Chan YM, Raj R, Pampillo M, Elbert A, Noonan M, Gillio-Meina C, Caligioni C, Berube NG, Bhattacharya M, Watson AJ, Seminara SB, Babwah AV. Implantation failure in female Kiss1−/− mice is independent of their hypogonadic state and can be partially rescued by leukemia inhibitory factor. Endocrinology. 2014;155:3065–78. doi: 10.1210/en.2013-1916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Dorfman MD, Garcia-Rudaz C, Alderman Z, Kerr B, Lomniczi A, Dissen GA, Castellano JM, Garcia-Galiano D, Gaytan F, Xu B, Tena-Sempere M, Ojeda SR. Loss of Ntrk2/Kiss1r signaling in oocytes causes premature ovarian failure. Endocrinology. 2014;155:3098–11. doi: 10.1210/en.2014-1111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gaytan F, Garcia-Galiano D, Dorfman MD, Manfredi-Lozano M, Castellano JM, Dissen GA, Ojeda SR, Tena-Sempere M. Kisspeptin receptor haplo-insufficiency causes premature ovarian failure despite preserved gonadotropin secretion. Endocrinology. 2014;155:3088–97. doi: 10.1210/en.2014-1110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zhang P, Tang M, Zhong T, Lin Y, Zong T, Zhong C, Zhang B, Ren M, Kuang H. Expression and function of kisspeptin during mouse decidualization. PloS ONE. 2014;9:e97647. doi: 10.1371/journal.pone.0097647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bhattacharya M, Babwah AV. Kisspeptin: Beyond the brain. Endocrinology. 2015;156:1218–27. doi: 10.1210/en.2014-1915. [DOI] [PubMed] [Google Scholar]
- 9.Lapatto R, Pallais JC, Zhang D, Chan Y-M, Mahan A, Cerrato F, Le WW, Hoffman GE, Seminara SB. Kiss1−/− mice exhibit more variable hypogonadism than Gpr54−/− mice. Endocrinology. 2007;148:4927–36. doi: 10.1210/en.2007-0078. [DOI] [PubMed] [Google Scholar]
- 10.Kotani M, Detheux M, Vandenbogaerde A, Communi D, Vanderwinden JM, Le Poul E, Brézillon S, Tyldesley R, Suarez-Huerta N, Vandeput F, Blanpain C, Schiffmann SN, Vassart G, Parmentier M. The metastasis suppressor gene KiSS-1 encodes kisspeptins, the natural ligands of the orphan G protein-coupled receptor KISS1R. J Biol Chem. 2001;276:34631–36. doi: 10.1074/jbc.M104847200. [DOI] [PubMed] [Google Scholar]
- 11.Szereszewski JM, Pampillo M, Ahow MR, Offermanns S, Bhattacharya M, Babwah AV. GPR54 regulates ERK1/2 activity and hypothalamic gene expression in a Gαq/11 and β-arrestin-dependent manner. PloS One. 2010;5:e12964. doi: 10.1371/journal.pone.0012964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ahow M, Min L, Pampillo M, Nash C, Wen J, Soltis K, Carroll RS, Glidewell-Kenney CA, Mellon PL, Bhattacharya M, Tobet SA, Kaiser UB, Babwah AV. Kiss1r signals independently of Gαq/11 and triggers LH secretion via the β-arrestin pathway in the male mouse. Endocrinology. 2014;155:4433–46. doi: 10.1210/en.2014-1304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.ELHabazi K, Humbert JP, Bertin I, Schmitt M, Bihel F, Bourguignon JJ, Bucher B, Becker JA, Sorg T, Meziane H, Petit-Demouliere B, Ilien B, Simonin F. Endogenous mammalian RF-amide peptides, including PrRP, kisspeptin and 26RFa, modulate nociception and morphine analgesia via NPFF receptors. Neuropharmacology. 2013;75:164–71. doi: 10.1016/j.neuropharm.2013.07.012. [DOI] [PubMed] [Google Scholar]
- 14.Roseweir AK, Kauffman AS, Smith JT, Guerriero KA, Morgan K, Pielecka-Fortuna J, Pineda R, Gottsch ML, Tena-Sempere M, Moenter SM, Terasawa E, Clarke IJ, Steiner RA, Millar RP. Discovery of potent kisspeptin antagonists delineate physiological mechanisms of gonadotropin regulation. J Neurosci. 2009;29:3920–9. doi: 10.1523/JNEUROSCI.5740-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Liu Z, Ren C, Jones W, Chen P, Seminara SB, Chan YM, Smith NF, Covey JM, Wang J, Chan KK. LC-MS/MS quantification of a neuropeptide fragment kisspeptin-10 (NSC 741805) and characterization of its decomposition product and pharmacokinetics in rats. J Chromatogr B Analyt Technol Biomed Life. 2013;926:1–8. doi: 10.1016/j.jchromb.2013.02.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hantak AM, Bagchi IC, Bagchi MK. Role of uterine stromal-epithelial crosstalk in embryo implantation. Int J Dev Biol. 2014;58:139–46. doi: 10.1387/ijdb.130348mb. [DOI] [PMC free article] [PubMed] [Google Scholar]