

Abstract
The maintenance of T cell repertoire diversity involves the entry of newly developed T cells, as well as the maintenance of memory T cells generated from previous infections. This balance depends on competition for a limited amount of homeostatic cytokines and interaction with self-peptide MHC-I. In the absence of prior infection, memory-like or memory phenotype (MP) CD8 T cells can arise from homeostatic cytokine exposure during neonatal lymphopenia. Aside from downstream cytokine signaling, little is known about the regulation of the conversion of naïve CD8 T cells to MP CD8 T cells during acute lymphopenia. We have identified a novel negative regulatory role for the adapter protein ADAP in CD8 T cell function. We show that in the absence of ADAP, naïve CD8 T cells exhibit a diminished response to stimulatory Ag, but an enhanced response to weak agonist altered peptide ligands. ADAP-deficient mice exhibit an increased number of MP CD8 T cells that occurs following thymic emigration and is largely T cell intrinsic. Naïve ADAP-deficient CD8 T cells are hyperresponsive to lymphopenia in vivo and exhibit enhanced activation of STAT5 and homeostatic antigen-independent proliferation in response to IL-15. Our results indicate that ADAP dampens naïve CD8 T cell responses to lymphopenia and IL-15, and demonstrates a novel antigen-independent function for ADAP in the suppression of MP CD8 T cell generation.
INTRODUCTION
T cell homeostasis is carefully balanced in a healthy host to maintain a diverse T cell repertoire against potential foreign pathogens. Utilizing both self-peptide MHC-I and IL-7 signaling pathways, naïve CD8 T cells compete for space with each other and a steady emigration of newly developed T cells out of the thymus (1). Early in life, while the T cell pool is developing, increased availability of homeostatic cytokines in the secondary lymphoid organs can induce some recent thymic emigrants (RTEs) to gradually proliferate and differentiate into memory-like T cells, termed memory phenotype (MP) (2). This period of neonatal lymphopenia is the primary generator of MP T cells, which are predominately foreign antigen-inexperienced and are maintained long into adulthood (3). MP T cells have similar functional capabilities as foreign-antigen induced memory cells, but do not require prior antigen experience (4). These cells can also be generated after exposure to an acute lymphopenic environment, which is of clinical relevance, as chemotherapy, late-stage HIV infection and exposure to radiation can render the host lymphopenic (1). Furthermore, allowing naïve T cells to become MP in the absence of an infection is proposed to help protect the neonate from infections, although there is a risk of promoting the survival of self-reactive T cells in this process (1, 4, 5)
The molecular factors that drive naïve T cell homeostasis and permit the generation of MP T cells from the naïve, antigen-inexperienced pool are only partially understood. While joint signaling by IL-7 and self-peptide MHC-I are thought to be the main drivers of naïve T cell homeostasis, optimal survival of naïve T cells is dependent on additional signaling from IL-15 (6). IL-15 signaling in naïve T cells drives the expression of the anti-apoptotic protein Bcl-2, but does not trigger proliferation, except in extreme situations, such as in the absence of CD122 (6,7). Indeed, disruption of IL-15 signaling in mice lacking suppressor of cytokine signaling-1 (SOCS-1) results in altered T cell homeostasis (8). Both naïve and MP CD8 T cells are hyperresponsive to IL-15 in the absence of SOCS-1, leading to robust proliferation, MP generation and neonatal mortality (8). However, while IL-15 can drive MP, additional molecular regulators that control the reactivity to MHC-I and homeostatic cytokines for MP generation have yet to be identified.
ADAP is a multifunctional adaptor protein that coordinates the formation of signaling complexes that promote TCR-mediated activation of integrins, as well as activation of the NF-κB and JNK signaling pathways (9). The expression of ADAP is restricted to cells of hematopoietic origin, including conventional CD4 and CD8 T cells and unconventional thymocytes, but is not expressed in B cell lineage cells after the Pro-B stage (10). ADAP is required for optimal positive and negative selection during conventional CD4 and CD8 T cell development, but dispensable for the development of unconventional thymocytes, including natural killer T (NKT) cells (10, 11). ADAP is localized to the cytosol, where a fraction is constitutively associated with Src kinase-associated phosphoprotein of 55 kDa (SKAP55) (12). The ADAP-SKAP55 signaling module is critical for TCR-mediated activation of integrin-mediated adhesion with APCs (12, 13). A second pool of ADAP is not associated with SKAP55, but activates NF-κB and JNK in a TCR-inducible manner (12, 14–16).
Analysis of ADAP function utilizing primary T cells following Ab-mediated TCR stimulation or mature naïve CD4 T cells following cognate-peptide MHC-II stimulation has demonstrated that ADAP is a positive regulator of T cell signaling that enhances T cell sensitivity to antigen. In contrast, the function of ADAP in CD8 T cells and in self-peptide MHC-I interactions has been largely unexplored. We sought to understand the role of ADAP in the homeostasis of CD8 T cells. Our findings reveal an unexpected negative role for ADAP in the generation of MP CD8 T cells and IL-15 signaling.
MATERIALS AND METHODS
Mice
C57BL/6 (B6) wild-type and ADAP−/− mice were generated as previously described (17). P14 ADAP−/− Rag2−/− mice were generated by crossing ADAP−/− mice with P14 Rag2−/− mice (provided by Dr. S. Jameson, University of Minnesota). OT-I ADAP−/− mice were generated by crossing ADAP−/− mice with OT-I mice (The Jackson Laboratory, Bar Harbor, ME). Nur77-GFP ADAP−/− mice were generated by crossing ADAP−/− mice with Nur77-GFP mice (provided by Dr. K. Hogquist and Dr. M. Farrar, University of Minnesota) (18). All mice were harvested between 8–12 weeks of age, unless otherwise specified. B6 CD45.1, B6 CD45.2 and Rag1−/− CD45.2 recipient mice were purchased from The Jackson Laboratory. IL-15−/− recipient mice were kindly provided by Dr. M. Farrar and Dr. D. Masopust, University of Minnesota. Mice were housed in specific pathogen-free facilities at the University of Minnesota. All experimental protocols involving the use of mice were approved by the Institutional Animal Care and Use Committee at the University of Minnesota.
Flow cytometry and reagents
Cell surface staining for flow cytometry was performed with ice-cold HBSS supplemented with 2% bovine serum (FACS buffer). Single cell suspensions were washed with FACS buffer, stained with surface marker antibodies, then washed twice before multi-parameter flow cytometric detection on a BD LSRFortessa (Becton Dickinson, San Jose, CA, USA). Directly conjugated fluorescent antibodies used include: CD4 (clone GK1.5), CD8α (clone 53–6.7), CD45.1 (clone A20), CD45.2 (clone 104), and B220 (clone RA3-6B2) (Tonbo); CD16/32 (clone 93), CD24 (clone M1/69), CD127 (clone SB/199), I-Ab (clone 25-9-17), Ter-119 (clone TER-119), and Vα2 (clone B20.1) (Biolegend); CD44 (clone IM7), CD122 (clone TM-Beta 1), CD124 (clone MIL4R-M1) and pSTAT5 pY694 (clone 47) (BD Biosciences); TCR-beta (clone H57-597), QA2 (clone 69H1-9-9) and F4/80 (clone BM8) (eBioscience). Cell cultures were maintained in complete T cell media: RPMI 1640 supplemented with 10% FCS, 4 mM L-glutamine, 0.1 mM nonessential amino acids, 1 mM sodium pyruvate, 100 U/ml penicillin and streptomycin, 10 mM HEPES, and 5 mM 2-ME (RP-10).
Conjugate assay
Flow cytometry-based conjugate assays were performed as previously described (12). Briefly, cells from pLN were harvested from wild-type and ADAP−/− OT-I adult mice and stained with FITC-conjugated anti-CD44. Polyclonal B6 splenocytes were harvested and stained with Cell Tracker Orange (Molecular Probes), then left unpulsed, or pulsed with N4, Q4 or T4 peptides for 30 min. at 37°C at the indicated concentrations. Wild-type or ADAP−/− OT-I T cells were mixed at a 1:1 ratio with labeled, pulsed splenocytes in a 96-well round bottom plate, pelleted and incubated at 37°C for 10 min. The cells were then vigorously mixed in a plate shaker for 20 s., fixed for 30 min. in 1% paraformaldehyde, and stained for flow cytometry. Conjugates were defined as CD8+ Vα2+ T cells, either CD44lo or CD44hi, that co-stain with B22O and Cell Tracker Orange.
Mixed bone marrow chimeras
Mixed bone marrow chimeras were generated by mixing T cell-depleted bone marrow preparations from CD45.1/2 (wild-type) and CD45.2 (ADAP−/−) mice at indicated ratios and injecting 4 × 106 to 10 × 106 total cells into lethally irradiated (1100 rads) CD45.1 animals (19). All chimeras were analyzed 8–12 weeks after transplant.
Naive CD8 T cell transfer
Lymph nodes were harvested from CD45.2 (wild-type) and CD45.1/2 (ADAP−/−) mice and CD8 mature naïve T cells were isolated by negative magnetic bead enrichment similar to previously described methods (20). Briefly, single-cell suspensions were incubated with the following FITC-conjugated antibodies: CD4, B220, F4/80, CD16/32, I-Ab, Ter119, and CD44. After washing, cells were incubated with anti-FITC microbeads and passed over LS columns on magnets according to manufacturer’s instructions (Miltentyi Biotec, Auburn, CA) to capture non-CD8 cells. Purity of the flow-through fraction was > 95% CD8+ CD44lo. Purified cells were co-transferred into CD45.1 recipients at 1:1 ratio. Recipient mice were analyzed 1 and 14 days after transfer. For Rag1−/− recipient experiments, CD45.1 (wild-type) and CD45.1/2 (ADAP−/−) OT-I mice were used as donors. Cells were purified as described above and 2 × 106 naïve CD8 T cells were co-transferred into B6 or Rag1−/− CD45.2 mice at a 1:1 ratio. Recipient mice were analyzed 7 days after transfer.
Naïve CD8 T cell transfer into lymphopenic hosts
Lymph nodes were harvested from CD45.2 (wild-type) and CD45.2 (ADAP−/−) mice and CD8 mature naïve T cells were isolated by negative magnetic bead enrichment as described above. Cells were labeled with the intravital dye CFSE (Invitrogen) for cell proliferation. Briefly, cells were resuspended in PBS with 5% FBS at 2 × 107 cells/mL. CFSE diluted in PBS was added 1:1 to the cells (2.5 uM final concentration) and incubated at 25°C for 5 min. Cells were quenched by washing twice with PBS with 2% FBS. Cells were transferred into separate sub-lethally irradiated (550 rads) CD45.1 animals (21). Recipient mice were analyzed 10 days after transfer.
Detection of intracellular phospho-STAT5
Cytokine stimulation and detection of pSTAT5 were performed as previously described (22). Briefly, splenocytes from polyclonal wild-type and ADAP−/− mice were rested for 2 hours at 37°C in complete T cell medium at 2 × 106 cells/ml. Recombinant mouse IL-7 (Miltenyi Biotech) or IL-15 (R&D Systems) (1 ng/ml, or 0.1–300 ng/ml) was added to culture media for 15 min. Cells were fixed by the addition of paraformaldehyde (to 1.6%) for 10 min at room temperature, washed and then permeabilized with 1 ml of ice-cold methanol added to the cells while vortexing. After incubation for 15 min at 4°C, cells were washed twice and stained for surface and intracellular antigens for 30 min at room temperature.
In vitro cytokine culture
Cytokine culture was performed as previously described (22). Lymph nodes were harvested from CD45.1/2 (wild-type) and CD45.2 (ADAP−/−) OT-I mice and naïve CD8 mature T cells were isolated by negative magnetic bead enrichment as described above. Cells were mixed at a 1:1 ratio and labeled with CFSE, as described above. Cells were then plated at 2 × 105 cells/ml and rested overnight (~16h) at 37°C in complete T cell medium without cytokine. IL-7 or IL-15 (10 ng/ml) was then added to the cultures for 5 days.
Statistics
Graphpad Prism (version 5.03, Graphpad Software, La Jolla, CA) was used to determine statistical significance using Student’s unpaired two-tailed t tests. The p value cutoffs and notation were used as follows: *, p < 0.05; **, p < 0.01; ***, p < 0.001. For pSTAT5 gMFI analysis, statistics were performed using a normalized value = 100. A nonlinear curve was generated for the dose response to IL-15 to determine LogEC50s. For CFSE gMFI analysis, statistics were performed using a normalized value = 1.
RESULTS
ADAP controls sensitivity to antigen
Initially we sought to confirm ADAP as a positive regulator of CD8 T cells interactions with Ag-pulsed APCs, as has been documented for CD4 T cells (9). We assessed the ability of wild-type and ADAP-deficient CD8+ OT-I TCR-Tg T cells, which recognize SIINFEKL (N4) peptide in the context of MHC-I H-2Kb with high affinity (23), to form stable contacts with peptide-pulsed APCs by flow cytometry. Both wild-type and ADAP-deficient naïve OT-I T cells exhibited minimal adhesion to APCs in the absence of peptide Ag (Fig. 1A, left). Addition of Ag resulted in a dose-dependent increase in CD8 T cell-APC conjugates, but we consistently observed reduced CD8 T cell-APC contacts in the absence of ADAP at low antigen doses (Fig. 1A, left). This defect diminished with the addition of higher concentrations of N4 peptide. Thus, similar to our previous observations with CD4 T cells, naïve CD8 T cells require ADAP for maximal T-APC interactions (24).
Figure 1. ADAP modulates T-APC interactions across multiple TCR affinities.

Conjugate assays between WT or ADAP−/− OT-I T cells and B6 splenocytes were performed as described in Materials and Methods. T:APC conjugate formation of naïve CD44lo OT-I T cells (A) and MP CD44hi OT-I T cells (B) with B6 splenocytes pulsed with the indicated concentrations of N4 (left), Q4 (center) or T4 (right) peptides. Results are averages from duplicate or quadruplicate wells from independent experiments (± SEM): seven experiments (N4), four experiments (Q4), and three experiments (T4).
As ADAP positively regulates TCR signaling, we hypothesized that intermediate and weak agonist interactions would be more dependent on ADAP. Altered peptide ligands (APLs) have been developed for the OT-I TCR that trigger less potent responses of mature naïve CD8 T cells, which allows for the analysis of a single TCR over a range of agonist strengths (23). Overall conjugate efficiency of wild-type naïve CD8 OT-I T cells was reduced with intermediate and weak agonist APLs SIIQFEKL (Q4) and SIITFEKL (T4), respectively (23, 25). Unexpectedly, in contrast to our observation with N4, where ADAP was required for maximal conjugate efficiency at a given peptide dose, we found that T-APC interactions with the intermediate strength agonist APL Q4 were equivalent between wild-type and ADAP-deficient OT-I T cells (Fig 1A, center). Even more striking, ADAP-deficient naïve OT-I T cells exhibited enhanced interaction with APCs pulsed with T4 APL when compared to wild-type controls (Fig. 1A, right). Thus, ADAP appears to modulate CD8 T cells interactions with Ag-pulsed APCs, both by boosting strong agonist interactions, and by minimizing weak agonist interactions.
Increased memory phenotype CD8 T cells in the absence of ADAP
Even in the absence of immunization, approximately 10–20% of CD8 peripheral T cells in wild-type C57BL/6 mice housed in SPF conditions express elevated levels of the activation marker CD44 (Fig. 2A) and CD122 (Fig. 2B), a phenotype characteristic of memory CD8 T cells (3). In our analysis of ADAP-deficient mice, we consistently observed a two-fold increase in the frequency of CD44hi CD122hi MP CD8 T cells in ADAP-deficient mice compared to littermate controls (Figs. 2B and 2C). From here on, we define naïve CD8 T cells as CD44lo CD122lo and MP CD8 T cells as CD44hi CD122hi. When compared to wild-type mice, the total number of MP CD8 T cells in the spleen of ADAP-deficient mice was also elevated (Fig. 2D). Thus, ADAP-deficient mice demonstrate an increased CD8 MP T cell population in the steady state.
Figure 2. Increased memory phenotype CD8 T cells in the absence of ADAP.
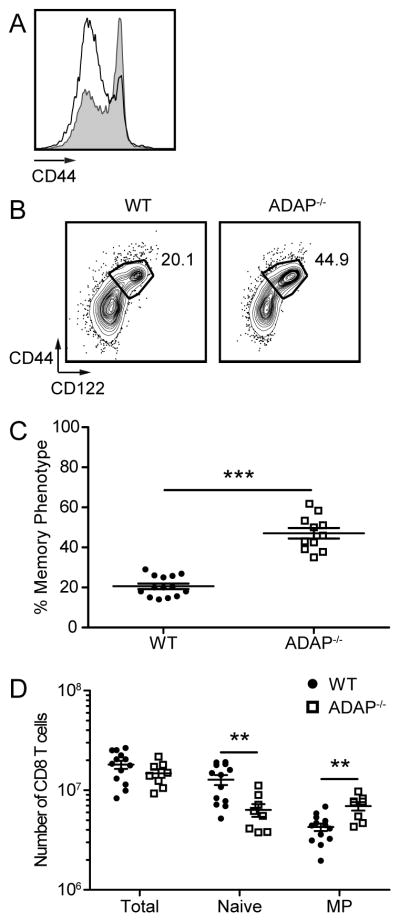
Cells from spleens were obtained from WT and ADAP−/− adult mice (aged 8–12 weeks), and the expression of CD44 and CD122 was analyzed on CD8+ CD4− TCR-β + cells. (A) CD44 staining from WT (black) and ADAP−/− (grey shaded) cells. (B) Contour plots of bulk CD8 T cells. Numbers represent the percentage of cells with the phenotype of memory (CD44hi CD122hi). (C) The percentage of CD44hi CD122hi cells in the spleens of WT (black circles) and ADAP−/− (open squares) mice. (D) Absolute number of total CD8+ CD4− TCR-β+, CD44lo CD122lo, and CD44hi CD122hi cells from the spleen. The results (C and D) are compiled from at least five independent experiments (± SEM), with at least two mice per experiment.
We next assessed the adhesion of wild-type and ADAP-deficient CD44hi MP CD8 OT-I T cells with APCs pulsed with the N4, Q4 or T4 peptides. With all 3 ligands, we observed a higher level of conjugate formation by MP CD8 OT-I T cells compared to naïve CD8 OT-I T cells (Fig. 1). The pattern of promoting strong agonist interactions and suppressing weak agonist interactions was maintained in MP CD8 T cells in the absence of ADAP, although to a lesser degree than we observed with naïve CD8 T cells (Fig. 1B). These results indicate an enhanced ability of MP CD8 T cells to interact with APCs pulsed with agonist peptide ligands compared to naïve CD8 T cells, and a requirement for ADAP in regulating Ag sensitivity of MP CD8 T cells.
Loss of ADAP does not enhance MP CD8 T cells in the thymus
One mechanism for the development of MP CD8 T cells in the periphery involves the response of mature CD8+CD4− thymocytes to IL-4 produced by thymic NKT cells expressing promyelocytic leukemia zinc finger protein (PLZF) (26). When we analyzed ADAP-deficient mice, we did not observe changes in the thymus indicative of ADAP-dependent generation of CD8 MP T cells via PLZF+ NKT cells. First, there were comparable numbers of MP CD8 T cells among mature CD8 SP thymocytes in wild-type and ADAP-deficient mice (Fig. 3A). Total numbers of mature naïve CD8 SP thymocytes were also unaffected by the loss of ADAP (Fig. 3A). Second, increases in MP CD8 thymocytes in other mouse models are associated with an increased percentage of IL-4-producing PLZF+ NKT cells in the thymus and elevated levels of serum IgE (26). We observed a statistically significant increase in the percentage, but not in the number, of thymic CD1d-αGalCer tetramer+ NKT cells and γδ+ NKT cells in ADAP-deficient mice (Supplemental Fig. 1A–C). However, this was not associated with elevated levels of serum IgE in aged ADAP-deficient mice (Supplemental Fig. 1D). Thus, increases in MP CD8 T cells in the periphery of ADAP-deficient mice are not due to elevated generation of MP CD8 T cells in the thymus.
Figure 3. Memory phenotype in the absence of ADAP occurs after CD8 T cells exit the thymus.
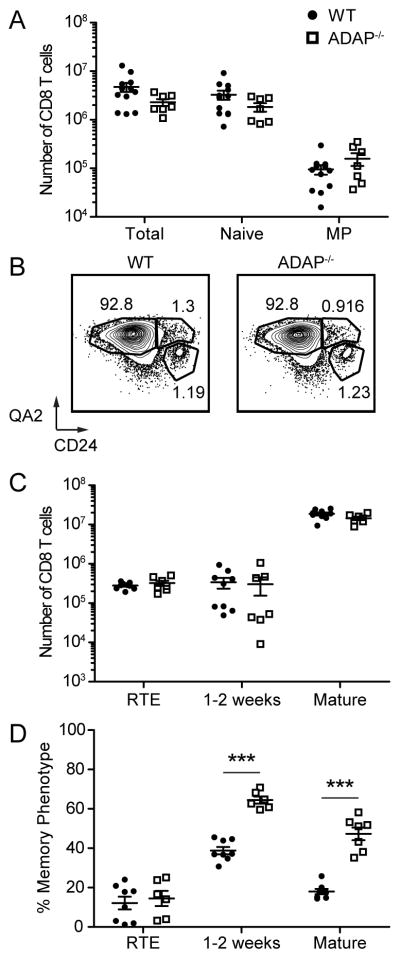
Cells from thymus (A) or spleen (B–D) were obtained from WT and ADAP−/− unimmunized 8–12 week old adult mice and the expression of surface molecules was analyzed on CD8+ CD4− TCR-β+ cells. (A) Absolute number of total CD8SP TCR-βhi, CD44lo CD122lo, and CD44hi CD122hi cells from the thymus. (B) CD24 and QA2 staining from WT and ADAP−/− spleens. Numbers represent the percentage of RTE, 1–2 week post egress and mature CD8 T cells. Recent thymic emigrants (RTEs) are defined as CD24hi Qa2lo. Cells that have emigrated between 1–2 weeks prior are defined as CD24hi Qa2hi. Mature cells are defined as CD24lo Qa2hi. (C) Absolute number of RTE, 1–2 week post egress and mature CD8 T cells. (D) The percentage of MP from RTE, 1–2 week post egress and mature CD8 T cell populations. The results (A) are compiled from at least five independent experiments (± SEM) with at least two mice per experiment, and (C and D) are compiled from at least three independent experiments with at least three mice per experiment.
Higher conversion of recent CD8 thymic emigrants to MP in the absence of ADAP
We next analyzed the maturation of CD8 T cells after emigration from the thymus in adult mice. Recent thymic emigrants (RTEs) and mature CD8 T cells can be differentiated by the expression of QA2 and CD24 (27). RTEs that have emigrated from the thymus within a week are QA2lo CD24hi, while T cells that have emigrated between 1–2 weeks prior have upregulated QA2 while maintaining high levels of CD24 (27). Mature T cells, which emigrated at least 2 weeks prior, are QA2hi CD24lo (27). Comparison of RTE, 1–2 week emigrated, and mature populations from wild-type and ADAP-deficient unimmunized adult mice demonstrate a similar frequency and number of these populations, indicating similar emigration and maturation of RTEs (Fig. 3B and C). We also assessed the frequency of MP CD8 T cells among peripheral CD8 populations. RTEs are predominately CD122lo, while 1–2 week post-egress CD8 T cells exhibit a large population of MP cells. There is a significantly greater frequency of MP CD8 T cells among ADAP-deficient 1–2 week post egress CD8 T cells than wild-type (Fig. 3D). This difference is maintained in the mature population, which constitutes the majority of the CD8 T cells in the periphery of an adult mouse.
Alterations in the CD8 T cell repertoire are not driving ADAP-dependent CD8 MP conversion
Previous studies have demonstrated a requirement for ADAP in both positive and negative selection during T cell development in the thymus (11). Altered T cell development could modify the T cell repertoire of ADAP-deficient CD8 T cells, resulting in naïve CD8 T cells with a higher propensity to become MP. To address this possibility, we assessed the frequency of MP CD8 T cells in wild-type and ADAP-deficient, Rag2-deficient, P14 TCR-Tg mice. The use of P14 TCR-Tg mice allowed us to compare wild-type and ADAP-deficient CD8 T cells with the same TCR, eliminating any differences that may have arisen in the T cell repertoire due to differences in positive and negative selection in the absence of ADAP. The frequency of peripheral MP CD8 T cells was increased in ADAP−/− Rag2−/− P14-Tg mice compared to wild-type mice (Fig. 4A and 4B). These results support a TCR-independent role for increased MP CD8 T cells in the absence of ADAP.
Figure 4. Memory phenotype in the absence of ADAP is not due to CD8 T cell self-reactivity, and is specific to ADAP-deficient CD8 T cells.

(A) CD44 and CD122 staining of spleens from WT and ADAP−/− Rag2−/− P14-Tg mice. Numbers represent the percentage of MP cells. Analyzed cells are CD8+ CD4− Vα2+ cells. (B) Percentage of MP WT and ADAP−/− Rag2−/− P14-Tg T cells. (C) Spleens from littermate polyclonal WT and ADAP−/− mice expressing the Nur77-GFP reporter were harvested and the expression of Nur77-GFP was analyzed by flow cytometry in CD8+CD44lo/CD122lo (Naïve) and CD8+CD44hi/CD122hi (MP) cells. Representative histograms depict Nur77-GFP expression for WT (black line) overlaid with ADAP−/− (grey shaded). (D) Quantification of Nur77-GFP fluorescence of naïve and MP CD8+ T cells from 4 separate littermate pairs of WT and ADAP−/− mice analyzed in two independent experiments. Values are normalized to Nur77-GFP expression observed in WT mice. (E–H) WT (CD45.1/2) and ADAP−/− (CD45.2) bone marrow cells at the indicated ratios were transferred into lethally irradiated CD45.1 recipients. Mixed bone marrow chimeras were harvested 8–12 weeks after transfer of donor marrow. (E–F) WT: ADAP−/−, 90:10 ratio, (G–H) WT: ADAP−/−, 10:90 ratio. (E and G) CD44 and CD122 staining of CD8+ CD4− TCR-β + donor cells from recipient spleens. Numbers represent the percentage of MP cells. (F and H) Percentage of CD44hi CD122hi donor cells. The results (B) are compiled from three independent experiments (± SEM) with at least two mice per experiment, (D) is compiled from three independent experiments, each with at least one littermate pair, and (F and G) are compiled from three independent experiments, each with at least 4 mice.
We also utilized the Nur77-GFP reporter mouse to determine if wild-type and ADAP-deficient CD8 T cells had similar tonic signaling (18). In the absence of ADAP, defects in negative selection may allow for the development of CD8 T cells with higher self-reactive TCRs than wild-type. After thymic egress, these self-reactive naïve CD8 T cells could come into contact with self-antigen, proliferate and gain memory-like characteristics. This might explain why more CD8 T cells become MP in the absence of ADAP. The Nur77-GFP reporter mouse allows us to measure the tonic TCR signaling and determine if ADAP-deficient CD8 T cells are more self-reactive. The geometric mean fluorescence intensity (gMFI) of Nur77-GFP was similar in wild-type and ADAP-deficient naïve or MP CD8 T cells isolated from unmanipulated mice (Fig. 4C and D). These results suggest that naïve and MP CD8 T cells in the absence of ADAP are not more self-reactive than wild-type CD8 T cells.
Loss of ADAP induces a CD8 T cell intrinsic increase in MP CD8 T cells
To directly test whether the increased MP observed in ADAP-deficient T cells is cell intrinsic, we created mixed bone marrow chimeras (BMCs). Bone marrow from a wild-type donor (CD45.1/2) was mixed with a minority of ADAP-deficient bone marrow (CD45.2) at a 90:10 ratio and transferred into lethally irradiated wild-type hosts (CD45.1). Spleens and thymi were harvested 8–12 weeks after transfer of donor marrow. In this environment, where the majority of the bone marrow-derived cells are wild-type, we observed that the frequency of ADAP-deficient MP CD8 T cells was 1.5 fold greater than wild-type (Fig. 4E and F), similar to what we observed in unmanipulated ADAP-deficient mice. This result suggests a T cell intrinsic mechanism for ADAP-dependent generation of MP CD8 T cells.
We also generated mixed BMCs with a majority of ADAP-deficient bone marrow to test the effect of having the majority of developing thymocytes lacking ADAP on the minority of wild-type CD8 T cells. In an ADAP-deficient thymocyte majority situation, we observed an increased frequency of wild-type MP CD8 T cells when compared to normal wild-type mice (Fig. 4G and H, Fig 2B and C). Similar to what we observed with mixed BMCs with a majority of wild-type cells, there was a higher percentage of ADAP-deficient MP CD8 T cells when compared to wild-type CD8 T cells although the difference was not as pronounced as what we observed in unmanipulated mice or BMCs with a majority of wild-type cells (Fig. 4G and H). These results suggest that there may also be an additional role for T cell extrinsic factors in ADAP-dependent generation of MP CD8 T cells.
Mature naïve CD8 T cells do not convert to MP in the absence of ADAP
To determine if mature naïve ADAP-deficient CD8 T cells have a higher propensity to convert to MP in the steady state, we isolated CD44lo CD122lo naïve CD8 T cells from wild-type and ADAP-deficient pLNs, and co-transferred these cells into adult wild-type hosts. Two weeks after transfer, we assessed the conversion of donor naïve CD8 T cells to MP, a time frame in which RTEs mature and can convert to MP. The majority of both wild-type and ADAP-deficient CD8 T cells retained a CD44lo CD122lo naïve phenotype (Fig. 5A and B), with minimal conversion to MP. These findings suggest that naïve ADAP-deficient T cells do not have a higher propensity to convert from naïve to MP in a wild-type host.
Figure 5. ADAP-deficient mature naïve CD8 T cells do not more readily convert to memory phenotype in the steady state.
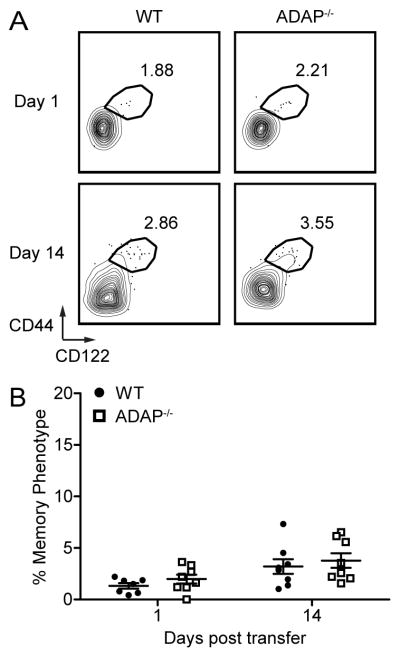
Naïve CD8 T cells from pLNs were isolated from WT (CD45.2) and ADAP−/− (CD45.1/2) adult mice, mixed at a 1:1 ratio and transferred into CD45.1 recipient mice (1 × 105 cells/mice). (A) CD44 and CD122 staining of transferred cells from recipient spleens. Numbers represent the percentage of MP CD8 T cells. (B) Percentage of CD44hi CD122hi donor cells. The results (B) are compiled from two independent experiments, each with four mice (± SEM).
Neonatal lymphopenic generation of MP CD8 T cells is increased in the absence of ADAP
A second mechanism for development of MP CD8 T cells involves the response of naïve CD8 T cells to the lymphopenic environment during the neonatal period (2). The highest frequency of MP CD8 T cells in a wild-type mouse occurs at 3 weeks (approximately 30%) and decreases to adult levels (approximately 20%) by 5 weeks of age (3). When compared to wild-type littermates, the frequency of MP CD8 T cells in 3 week old ADAP-deficient mice was 2 fold higher (Fig. 6A and B). Similar to published accounts, we found that the frequency of wild-type MP CD8 T cells was highest at 3 weeks and reached adult frequencies by 5 weeks (3). At all ages, we observed a greater frequency of MP CD8 T cells in ADAP-deficient mice (data not shown). These results support an early and consistent increased generation of MP CD8 T cells in the absence of ADAP.
Figure 6. Neonatal lymphopenic generation of memory phenotype CD8 T cells is greater in the absence of ADAP.
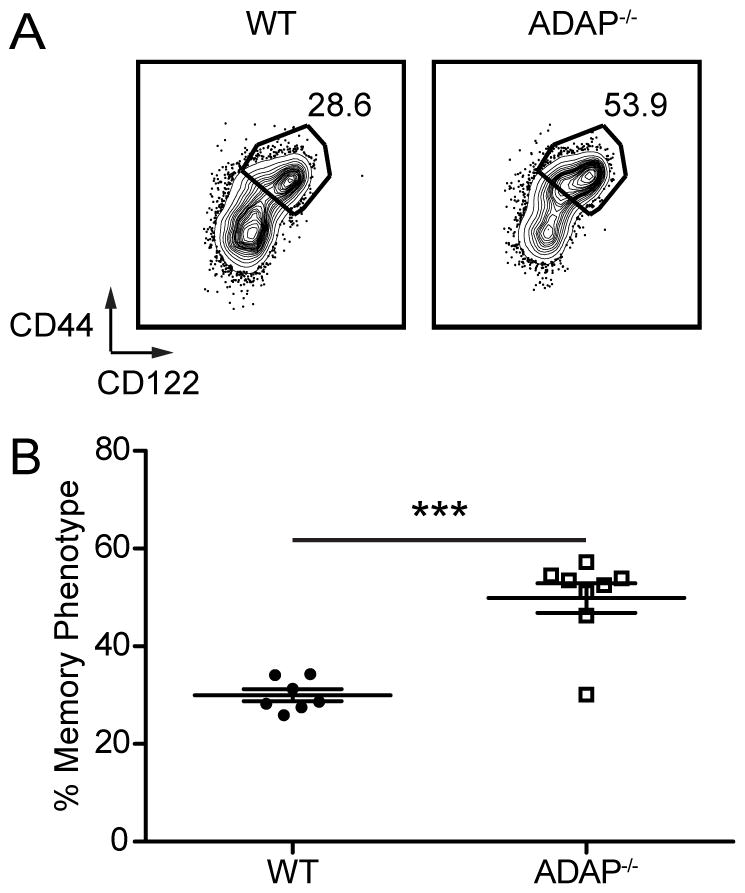
Cells from the spleen of 3 week old WT or ADAP−/− mice were harvested and analyzed for MP. (A) CD44 and CD122 staining. Numbers represent the percentage of MP CD8 T cells. (B) Percentage of CD8+ TCR-beta+ QA2hi CD24lo MP cells. The results (B) are compiled from two independent experiments (± SEM), with at least three mice in each group.
ADAP dampens naïve CD8 T cell responses to lymphopenia
To further investigate the role of lymphopenia in ADAP-deficient conversion of naïve CD8 T cells to MP, CD44lo CD122lo naïve CD8 T cells were labeled with CFSE and transferred into sub-lethally irradiated age-matched recipients. After 10 days, we assessed the conversion of wild-type and ADAP-deficient naïve CD8 T cells to MP, as well as lymphopenia induced proliferation (LIP). We found that a higher percentage of ADAP-deficient naïve CD8 cells converted to MP than their wild-type counterparts in a lymphopenic environment (Fig. 7A and B). In addition, a higher percentage of ADAP-deficient CD8 T cells had undergone greater than 4 divisions compared to wild-type CD8 T cells (Fig. 7C and D). These results suggest that CD44lo CD122lo naïve ADAP-deficient CD8 T cells are more sensitive to lymphopenic signals in vivo.
Figure 7. Mature naïve CD8 T cells convert to memory phenotype in the absence of ADAP in response to lymphopenic signals.

(A–E) Naïve CD8 T cells from pLNs were isolated from WT (CD45.2) and ADAP−/− (CD45.2) adult mice, labeled with CFSE, and separately transferred into normal or sub-lethally irradiated CD45.1 recipients. Recipient spleens were harvested 10 days after transfer. (A) CD44 and CD122 staining of transferred cells from lymphopenic spleens. Numbers represent the percentage of MP CD8 T cells. (B) Percentage of CD44hi CD122hi donor cells from lymphopenic recipient spleens. (C) CFSE staining from WT (black) or ADAP−/− (grey shaded) donor cells. (D) Percentage of donor cells that had undergone >4 cell divisions. (E) Ratio of WT:ADAP−/− CFSE gMFI in normal or lymphopenic recipient spleens. (F–J) Naïve OT-I CD8 T cells from pLNs were isolated from WT (CD45.1) and ADAP−/− (CD45.1/2) adult mice, labeled with CFSE, and co-transferred into normal or Rag1−/− CD45.2 recipients. Recipient spleens were harvested 7 days after transfer. (F) CD44 and CD122 staining of transferred cells from Rag1−/− spleens. Numbers represent the percentage of MP CD8 T cells. (G) Percentage of CD44hi CD122hi donor cells from Rag1−/− recipient spleens. (H) CFSE staining from WT (black) or ADAP−/− (grey shaded) donor cells. (I) Percentage of donor cells that had undergone >4 cell divisions. (J) Ratio of WT:ADAP−/− CFSE gMFI in normal or Rag1−/− recipient spleens. The results (B and D) are representative of three independent experiments (± SEM), with at least four mice of each condition. The results (G and I) are representative of three independent experiments (± SEM), with at least three mice of each condition. The results (E and J) are compiled from three independent experiments (± SEM).
To be sure that ADAP-deficient CD8 T cells were responding to lymphopenic signals and not the cytokine storm that results from sub-lethal irradiation of wild-type recipient mice, we also assessed the conversion of naïve ADAP-deficient CD8 T cells to MP in Rag1−/− hosts. Similar to what we observed with recipient mice that were sub-lethally irradiated, a higher percentage of ADAP-deficient naïve CD8 T cells compared with to wild-type naïve CD8 T cells converted to MP and underwent homeostatic proliferation in Rag1−/− hosts (Fig. 7E–J). These findings suggest that the enhanced response of ADAP-deficient CD8 T cells is due to the lymphopenic environment and not due to a cytokine storm.
IL-15 mediated induction of memory phenotype is suppressed by ADAP
LIP of naïve CD8 T cells is driven by IL-7 and self-peptide-MHC complexes, while MP CD8 T cells can utilize IL-7 or IL-15 and are MHC-I independent (1, 27). To determine a mechanism by which ADAP-deficient CD8 T cells are more responsive to lymphopenia, we assessed the ability of naïve CD8 T cells to transduce IL-7 and IL-15 signals. Bulk splenocytes from wild-type or ADAP-deficient mice were stimulated with IL-7 or IL-15 for 15 minutes and stained for intracellular phospho-STAT5. Both wild-type and ADAP-deficient naïve CD8 T cells expressed low levels of pSTAT5 in the absence of cytokine stimulation (Fig. 8A). An increase in pSTAT5 after the addition of IL-7 was comparable between wild-type and ADAP-deficient naïve CD8 T cells (Fig 8A and B). Interestingly, pSTAT5 induction after IL-15 treatment was 20% greater in ADAP-deficient naïve CD8 T cells (Fig. 8A and 8B). Dose response experiments also demonstrate a more robust response of ADAP-deficient naïve and MP CD8 T cells to IL-15 (Fig. 8C). Wild-type naïve CD8 T cells responded to IL-15 with a LogEC50 of 6.3 × 10−10, while ADAP-deficient naïve CD8 T cells responded with a LogEC50 of 2.0 × 10−10. ADAP-deficient MP CD8 T cells also had an enhanced response to IL-15 (LogEC50 at 2.4 × 10−10) compared to wild-type MP CD8 T cells (9.4 × 10−11). These results suggest that in the absence of ADAP, both naïve and MP CD8 T cells exhibit an enhanced response to IL-15 signals.
Figure 8. Mature naïve CD8 T cells are more responsive to IL-15 in the absence of ADAP.

(A–C) Bulk splenocytes were obtained from WT and ADAP−/− adult mice and stained for intracellular pSTAT5. (A) Histograms of WT (black) or ADAP−/− (grey shaded) pSTAT5 staining on CD8+ CD122lo cells after addition of 1 ng/mL of IL-7 or IL-15 for 15 minutes. (B) STAT5 phosphorylation was measured by flow cytometry and quantified as the population gMFI staining normalized to WT IL-7 or IL-15 stimulated samples from CD8+ CD122lo cells. (C) Dose response to IL-15 (0.1–300 ng/mL) for 15 minutes in CD8+ CD122lo cells (top) and CD8+ CD122hi cells (bottom) LogEC50 for CD8+ CD122lo cells was 6.3 × 10−10 (WT) and 2.0 × 10−10 (ADAP−/−), For CD8+ CD122hi cells, the LogEC50 was 2.4 × 10−10 (WT) and 9.4 × 10−11 (ADAP−/−). (D–G) Naïve CD8 T cells were isolated from WT (CD45.1/2) or ADAP−/− (CD45.2) OT-I adult mice, mixed at a 1:1 ratio, and labeled with CFSE. WT and ADAP−/− cells were co-cultured in the presence of 10 ng/mL of IL-7 or IL-15 for 5 days. (D) CFSE staining of WT and ADAP−/− CD8 T cells at day 0 or 5 after addition of cytokine. Gate indicates cells that have undergone 1 or more divisions. (E) Percentage of donor cells that have undergone one or more divisions. (F) CD122 staining of WT and ADAP−/− CD8 T cells at day 5 after addition of cytokine. Gate indicates cells that are CD122hi. (G) Percentage of CD8 T cells with expression of CD122hi. (H) Naïve OT-I CD8 T cells from pLNs were isolated from WT (CD45.1) and ADAP−/− (CD45.1/2) adult mice, labeled with CFSE, and co-transferred into untreated or sub-lethally irradiated CD45.2 or IL-15−/− CD45.2 recipients. Recipient spleens were harvested 7 days after transfer. Ratio of WT:ADAP−/− CFSE gMFI in untreated or sub-lethally irradiated wild-type or IL-15−/− recipient spleens. The results (B, C, E, G and H) are compiled from three independent experiments (± SEM).
To determine if this difference in IL-15 signaling is associated with functional differences in the response of ADAP-deficient CD8 T cells to IL-15, we cultured CFSE-labeled wild-type and ADAP-deficient mature naïve OT-I CD8 T cells in IL-7 or IL-15. After 5 days in culture, we assessed cytokine induced proliferation and MP conversion. The addition of IL-7 induced minimal proliferation of ADAP-deficient CD8 T cells, which was not significantly altered when compared to wild-type (Fig. 8D and E). In contrast, IL-15 stimulation induced proliferation of ADAP-deficient CD8 T cells, but not wild-type CD8 T cells. In addition, stimulation with IL-7 increased CD122 expression in both wild-type and ADAP-deficient CD8 T cells, but to a greater degree in the absence of ADAP (Fig. 8F and G). This difference in CD122 upregulation was even more striking with the addition of IL-15. To determine if IL-15 signaling in vivo is necessary for greater proliferation of ADAP-deficient naïve CD8 T cells, we co-transferred CFSE-labeled wild-type and ADAP-deficient naïve CD8 T cells into untreated or sublethally irradiated wild-type or IL-15−/− hosts. After 7 days, we assessed the CFSE gMFI of wild-type and ADAP-deficient CD8 T cells. As observed in previous experiments, ADAP-deficient CD8 T cells had undergone more divisions than their wild-type counterparts in a lymphopenic host. In contrast, in the absence of IL-15 signals, wild-type and ADAP-deficient CD8 T cells proliferated equally in a lymphopenic environment. We conclude that expression of ADAP dampens the ability of naïve CD8 T cells to respond to IL-15 signals.
DISCUSSION
In this study, we have revealed a novel negative regulatory role for the adapter protein ADAP in naïve CD8 responses to weak agonist peptides and IL-15. This work focused on the thymic and extra-thymic pathways that contribute to the generation of antigen-independent MP CD8 T cells (3, 26). Thymic-based MP CD8 T cell generation pathways can be traced to alterations in unconventional thymocyte development that result in an increase in NKT cells expressing IL-4, a cytokine that can drive the conversion of mature CD8 SP thymocytes to MP at high concentrations (26). In the periphery, the prime drivers of peripheral mature naïve CD8 T cell conversion to MP are: 1) self-reactivity; 2) exposure to the homeostatic cytokines IL-7 and IL-15; and 3) absence of negative signals (4). Our findings support an extra-thymic generation of MP CD8 T cells in the absence of ADAP, due to an enhancement of IL-15 signaling.
After our initial observation that ADAP-deficient CD8 T cells had greater frequencies of MP CD8 T cells, we investigated thymic-based MP CD8 T cell generation. Both inducible T cell kinase (Itk)-deficient mice and mice with a mutation in Src homology 2 domain-containing leukocyte phosphoprotein of 76 kDa (SLP-76:Y145F) demonstrate IL-4+ NKT cell-mediated CD8 MP generation (26). Itk inducibly binds SLP76-pY145 after TCR signaling, which may explain why the SLP-76:Y145F mutation phenocopies Itk-deficient animals (26). Although ADAP also inducibly associates with SLP-76 (9), we did not find evidence that the increased frequency of MP CD8 T cells in ADAP-deficient mice was due to an IL-4-dependent mechanism in the thymus. Our results are also consistent with the findings that the significant developmental defects in conventional T cells and NKT cells in SLP-76:Y145F and Itk-deficient mice are not observed in ADAP-deficient mice. Our findings do not support a thymic pathway generating MP CD8 T cells in the absence of ADAP.
A role for ADAP in regulating T cell responses to self peptides might be predicted, since ADAP is required for the optimal response of naïve CD4 T cells to Ag both in vitro and in vivo (13, 24), and TCR signaling has been proposed to be moderately attenuated in ADAP-deficient thymocytes (11). Similar to naïve CD4 T cells, we have observed that ADAP positively regulates the interaction of naïve CD8 OT-I T cells with APCs pulsed with the strong agonist N4 peptide. In contrast, the loss of ADAP enhanced the weak agonist interaction of OT-I T cells to APCs pulsed with the T4 APL. The enhanced adhesion of ADAP-deficient CD8 T cells to APCs pulsed with the weak agonist T4 APL is unusual and indicates that ADAP can play a negative regulatory role in TCR-dependent responses under certain conditions. This result suggested the possibility that the loss of ADAP might enhance the self-reactivity of naïve CD8 T cells, leading to increased MP CD8 T cell generation. The enhanced LIP of ADAP-deficient naïve CD8 T cells in vivo is consistent with this possibility. However, similar to what was observed in polyclonal mice, we found an increased percentage of MP CD8 T cells in P14 TCR-Tg mice. The P14 TCR-Tg is on Rag2−/− background, which prevents endogenous receptor rearrangement, ensuring TCR specificity. This result suggests that ADAP-deficient CD8 T cells do not convert to MP because of an altered TCR-repertoire with self-reactive T cells. Additionally, measuring tonic TCR signaling in naïve or MP CD8 T cells with the Nur77-GFP reporter system indicates similar self-reactivity between wild-type and ADAP-deficient CD8 T cells. Thus, ADAP negatively regulates the response of naïve CD8 T cells to weak agonist APLs, but the enhanced frequency of MP CD8 T cells in ADAP-deficient mice does not appear to be due to altered self-reactivity of ADAP-deficient naïve CD8 T cells.
Altered thymocyte development could result in decreased thymic output, thus enhancing MP conversion of peripheral mature naïve CD8 T cells by increasing the exposure of these T cells to homeostatic cytokines during neonatal development (4). We failed to detect defects in the development of mature CD8 SP T cells or in CD8 thymic output in the absence of ADAP, which has been reported in a previous study (11). We are uncertain of the reasons for this discrepancy, although we note that our experiments analyzed older mice (8–12 weeks) than the previous work, which assessed mice at 6 weeks of age (11). We do recognize that by qualifying RTEs only by QA2loCD24hi cells, we are excluding a large percentage of cells that could be identified in other systems (28). When we assessed mature naïve CD8 T cell conversion to MP in neonatal mice, we observed a greater frequency of MP CD8 T cells in the absence of ADAP. Together these results suggest that the enhanced conversion of neonatal ADAP-deficient CD8 T cells to MP is not likely due to thymic development or export defects. We performed mixed BMC experiments to rule out the possibility that the enhanced frequency of ADAP-deficient MP CD8 T cells is due to lymphopenic conditions in ADAP-deficient mice that might result in greater exposure of naïve CD8 T cells to homeostatic cytokines such as IL-7 and IL-15. These experiments support a T cell intrinsic response of ADAP-deficient CD8 T cells to lymphopenia, as 1.5 fold greater frequency of MP CD8 T cells was observed in ADAP-deficient CD8 T cells, compared to wild-type CD8 T cells, regardless of the majority population. Taken together, these results suggest that the conversion of naïve mature CD8 T cells to MP CD8 T cells in the absence of ADAP is not likely due to increased exposure to homeostatic cytokines.
We were surprised to discover our results support a unique negative regulatory role for ADAP in naïve CD8 T cell responses to the homeostatic cytokine IL-15. This is a previously unreported function for ADAP independent of TCR signaling. It is interesting to note that while ADAP-deficient CD8 T cells exhibit enhanced responsiveness to IL-15, the response of ADAP-deficient CD8 T cells to IL-7 is similar to what we observed with wild-type CD8 T cells. This suggests the possibility that ADAP plays a particularly important role in regulating CD8 T cell survival, since the optimal survival of naïve T cells is dependent on an IL-15 signal (6). The SOCS-1 protein, which binds directly to JAKs to inhibit the tyrosine-kinase activity and promotes ubiquitin-mediated degradation (29), has also been implicated in negatively regulating T cell homeostasis. SOCS-1-deficient mice have severe neonatal lethality due to overproduction of type II IFN (30). Similar to ADAP-deficient mice, SOCS-1-deficient mice have increased LIP and MP CD8 T cell conversion that is not linked to TCR affinity, but associated with an enhancement of IL-15 signaling (8, 30). As SOCS-1-deficient mice succumb to neonatal mortality (29), a phenotype that is not observed in ADAP-deficient mice, the molecular link between the enhanced responsiveness of SOCS-1-deficient CD8 T cells and ADAP-deficient CD8 T cells will require further investigation.
Our studies have revealed a novel negative regulatory role for ADAP in the conversion of naïve CD8 T cells to MP in the steady state and the response of naïve CD8 T cells to IL-15. ADAP is therefore a signaling protein involved in both the positive regulation of TCR responses to foreign antigens, and the negative regulation of naive T cell responses to homeostatic cytokines. Our results indicate that ADAP is a critical mediator of the body’s need to balance the maintenance of potentially useful T cells against genuine memory T cells from prior infections. We propose that ADAP functions to license pathogen-reactive T cells for activation during an immune response, while limiting self-reactive cells from attaining space in the T cell pool. By promoting strong agonist peptide MHC-I interactions, ADAP enhances activation of naïve T cells with foreign-peptide MHC-I specificity. In contrast, ADAP dampens weak agonist interactions and prevents generation of self-reactive cells. In addition, suppressing naïve IL-15 signaling could decrease the generation of MP CD8 T cells during instances of acute lymphopenia, which can occur after an infection (1). These findings reinforce the importance of future studies to understand how T cell homeostasis is maintained throughout life, from birth, during infections and into old age.
Supplementary Material
Acknowledgments
We thank F. Shoyama, T. Rivard, S. Jin and K. Benson for mouse genotyping and colony maintenance. Dr. Sarah Hamilton provided assistance with LIP and pSTAT5 staining. Dr. Yu-Jung Lee provided assistance with mixed bone marrow chimeras and unconventional thymocyte staining. Dr. Kristin Hogquist provided CD1d-αGalCer multimers. Drs. Mike Farrar and Kristin Hogquist provided Nur77-GFP mice, Dr. Stephen Jameson provided P14 Rag2−/− mice and Drs. Mike Farrar and David Masopust provided IL-15−/− mice. We thank Dr. Stephen Jameson and Dr. Erik Peterson for helpful discussion and critical review of the manuscript.
Abbreviations used in this article
- ADAP
adhesion and degranulation promoting adapter protein
- BMC
bone marrow chimera
- gMFI
geometric mean fluorescence intensity
- Itk
inducible T cell kinase
- LIP
lymphopenia induced proliferation
- MP
memory phenotype
- N4
SIINFEKL peptide
- NKT cells
natural killer T cells
- PLZF
promyelocytic leukemia zinc finger protein
- pLN
peripheral lymph node
- Q4
SIIQFEKL peptide
- RTE
recent thymic emigrant
- SKAP55
Src kinase-associated phosphoprotein of 55 kDa
- SLP-76
Src homology 2 domain-containing leukocyte phosphoprotein of 76 kDa
- SOCS-1
suppressor of cytokine signaling-1
- T4
SIITFEKL peptide
Footnotes
This work was supported by National Institutes of Health grant R01 AI038474 (to Y.S.). Y.S. is also supported by the Harry Kay Chair in Biomedical Research at the University of Minnesota.
References
- 1.Takada K, Jameson SC. Naive T cell homeostasis: from awareness of space to a sense of place. Nat Rev Immunol. 2009;9:823–832. doi: 10.1038/nri2657. [DOI] [PubMed] [Google Scholar]
- 2.Min B, McHugh R, Sempowski GD, Mackall C, Foucras G, Paul WE. Neonates support lymphopenia-induced proliferation. Immunity. 2003;18:131–140. doi: 10.1016/s1074-7613(02)00508-3. [DOI] [PubMed] [Google Scholar]
- 3.Akue AD, Lee JY, Jameson SC. Derivation and maintenance of virtual memory CD8 T cells. J Immunol. 2012;188:2516–2523. doi: 10.4049/jimmunol.1102213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Sprent J, Surh CD. Normal T cell homeostasis: the conversion of naive cells into memory-phenotype cells. Nat Immunol. 2011;12:478–484. doi: 10.1038/ni.2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Surh CD, Sprent J. Homeostasis of naive and memory T cells. Immunity. 2008;29:848–862. doi: 10.1016/j.immuni.2008.11.002. [DOI] [PubMed] [Google Scholar]
- 6.Berard M, Brandt K, Bulfone-Paus S, Tough DF. IL-15 promotes the survival of naive and memory phenotype CD8+ T cells. J Immunol. 2003;170:5018–5026. doi: 10.4049/jimmunol.170.10.5018. [DOI] [PubMed] [Google Scholar]
- 7.Cho JH, Boyman O, Kim HO, Hahm B, Rubinstein MP, Ramsey C, Kim DM, Surh CD, Sprent J. An intense form of homeostatic proliferation of naive CD8+ cells driven by IL-2. J Exp Med. 2007;204:1787–1801. doi: 10.1084/jem.20070740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Davey GM, Starr R, Cornish AL, Burghardt JT, Alexander WS, Carbone FR, Surh CD, Heath WR. SOCS-1 regulates IL-15-driven homeostatic proliferation of antigen-naive CD8 T cells, limiting their autoimmune potential. J Exp Med. 2005;202:1099–1108. doi: 10.1084/jem.20050003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Witte A, DJ, Baumgart K, Waldt N, Kuropka B, Freund C, Schraven B, Kliche S. Emerging roles of ADAP, SKAP55, and SKAP-HOM for integrin and NF-κB Signaling in T cells. Clin Cell Immunol. 2012;S12:002. doi: 10.4172/2155-9899.S12-002. [DOI] [Google Scholar]
- 10.Dluzniewska J, Zou L, Harmon IR, Ellingson MT, Peterson EJ. Immature hematopoietic cells display selective requirements for adhesion- and degranulation-promoting adaptor protein in development and homeostatsis. Eur J Immunol. 2007;37:3208–3219. doi: 10.1002/eji.200737094. [DOI] [PubMed] [Google Scholar]
- 11.Wu JN, Gheith S, Bezman NA, Liu QH, Fostel LV, Swanson AM, Freedman BD, Koretzky GA, Peterson EJ. Adhesion- and degranulation-promoting adapter protein is required for efficient thymocyte development and selection. J Immunol. 2006;176:6681–6689. doi: 10.4049/jimmunol.176.11.6681. [DOI] [PubMed] [Google Scholar]
- 12.Burbach BJ, Srivastava R, Ingram MA, Mitchell JS, Shimizu Y. The pleckstrin homology domain in the SKAP55 adapter protein defines the ability of the adapter protein ADAP to regulate integrin function and NF-κB activation. J Immunol. 2011;186:6227–6237. doi: 10.4049/jimmunol.1002950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mitchell JS, Burbach BJ, Srivastava R, Fife BT, Shimizu Y. Multistage T cell-dendritic cell interactions control optimal CD4 T cell activation through the ADAP-SKAP55-signaling module. J Immunol. 2013;191:2372–2383. doi: 10.4049/jimmunol.1300107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Srivastava R, Burbach BJ, Mitchell JS, Pagan AJ, Shimizu Y. ADAP regulates cell cycle progression of T cells via control of cyclin E and Cdk2 expression through two distinct CARMA1-dependent signaling pathways. Mol Cell Biol. 2012;32:1908–1917. doi: 10.1128/MCB.06541-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Srivastava R, Burbach BJ, Shimizu Y. NF-κB activation in T cells requires discrete control of IκB kinase α/β (IKKα/β) phosphorylation and IKKγ ubiquitination by the ADAP adapter protein. J Biol Chem. 2010;285:11100–11105. doi: 10.1074/jbc.M109.068999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Medeiros RB, Burbach BJ, Mueller KL, Srivastava R, Moon JJ, Highfill S, Peterson EJ, Shimizu Y. Regulation of NF-κB activation in T cells via association of the adapter proteins ADAP and CARMA1. Science. 2007;316:754–758. doi: 10.1126/science.1137895. [DOI] [PubMed] [Google Scholar]
- 17.Peterson EJ, Woods ML, Dmowski SA, Derimanov G, Jordan MS, Wu JN, Myung PS, Liu QH, Pribila JT, Freedman BD, Shimizu Y, Koretzky GA. Coupling of the TCR to integrin activation by Slap-130/Fyb. Science. 2001;293:2263–2265. doi: 10.1126/science.1063486. [DOI] [PubMed] [Google Scholar]
- 18.Moran AE, Holzapfel KL, Xing Y, Cunningham NR, Maltzman JS, Punt J, Hogquist KA. T cell receptor signal strength in Treg and iNKT cell development demonstrated by a novel fluorescent reporter mouse. J Exp Med. 2011;208:1279–1289. doi: 10.1084/jem.20110308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Weinreich MA, Takada K, Skon C, Reiner SL, Jameson SC, Hogquist KA. KLF2 transcription-factor deficiency in T cells results in unrestrained cytokine production and upregulation of bystander chemokine receptors. Immunity. 2009;31:122–130. doi: 10.1016/j.immuni.2009.05.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zumwalde NA, Domae E, Mescher MF, Shimizu Y. ICAM-1-dependent homotypic aggregates regulate CD8 T cell effector function and differentiation during T cell activation. J Immunol. 2013;191:3681–3693. doi: 10.4049/jimmunol.1201954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Tan JT, Ernst B, Kieper WC, LeRoy E, Sprent J, Surh CD. Interleukin (IL)-15 and IL-7 jointly regulate homeostatic proliferation of memory phenotype CD8+ cells but are not required for memory phenotype CD4+ cells. J Exp Med. 2002;195:1523–1532. doi: 10.1084/jem.20020066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Palmer MJ, V, Mahajan S, Chen J, Irvine DJ, Lauffenburger DA. Signaling thresholds govern heterogeneity in IL-7-receptor-mediated responses of naive CD8+ T cells. Immunol Cell Biol. 2011;89:581–594. doi: 10.1038/icb.2011.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Daniels MA, Teixeiro E, Gill J, Hausmann B, Roubaty D, Holmberg K, Werlen G, Hollander GA, Gascoigne NR, Palmer E. Thymic selection threshold defined by compartmentalization of Ras/MAPK signalling. Nature. 2006;444:724–729. doi: 10.1038/nature05269. [DOI] [PubMed] [Google Scholar]
- 24.Mueller KL, Thomas MS, Burbach BJ, Peterson EJ, Shimizu Y. Adhesion and degranulation-promoting adapter protein (ADAP) positively regulates T cell sensitivity to antigen and T cell survival. J Immunol. 2007;179:3559–3569. doi: 10.4049/jimmunol.179.6.3559. [DOI] [PubMed] [Google Scholar]
- 25.Yachi PP, Ampudia J, Zal T, Gascoigne NR. Altered peptide ligands induce delayed CD8-T cell receptor interaction-a role for CD8 in distinguishing antigen quality. Immunity. 2006;25:203–211. doi: 10.1016/j.immuni.2006.05.015. [DOI] [PubMed] [Google Scholar]
- 26.Lee YJ, Jameson SC, Hogquist KA. Alternative memory in the CD8 T cell lineage. Trends Immunol. 2011;32:50–56. doi: 10.1016/j.it.2010.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Boursalian TE, Golob J, Soper DM, Cooper CJ, Fink PJ. Continued maturation of thymic emigrants in the periphery. Nat Immunol. 2004;5:418–425. doi: 10.1038/ni1049. [DOI] [PubMed] [Google Scholar]
- 28.Fink PJ. The biology of recent thymic emigrants. Annu Rev Immunol. 2013;31:31–50. doi: 10.1146/annurev-immunol-032712-100010. [DOI] [PubMed] [Google Scholar]
- 29.Alexander WS, Hilton DJ. The role of suppressors of cytokine signaling (SOCS) proteins in regulation of the immune response. Annu Rev Immunol. 2004;22:503–529. doi: 10.1146/annurev.immunol.22.091003.090312. [DOI] [PubMed] [Google Scholar]
- 30.Ramanathan S, Gagnon J, Leblanc C, Rottapel R, Ilangumaran S. Suppressor of cytokine signaling 1 stringently regulates distinct functions of IL-7 and IL-15 in vivo during T lymphocyte development and homeostasis. J Immunol. 2006;176:4029–4041. doi: 10.4049/jimmunol.176.7.4029. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.