

Abstract
Purpose
To report the identification of a novel frameshift mutation and copy number variation (CNV) in PIKFYVE in two probands with fleck corneal dystrophy (FCD).
Methods
Slit-lamp examination was performed to identify characteristic features of FCD. After genomic DNA was collected, PCR amplification and automated sequencing of all 41 exons of PIKFYVE was performed. Using genomic DNA, quantitative PCR (qPCR) was performed to detect CNVs within PIKFYVE.
Results
In the first FCD proband, numerous panstromal punctate opacities were observed in each of the proband’s corneas, consistent with the diagnosis of FCD. Screening of PIKFYVE demonstrated a novel heterozygous frameshift mutation in exon 19, c.3151dupA, which is predicted to encode for a truncated PIKFYVE protein, p.(Asp1052Argfs*18). This variant was identified in an affected sister but not in the proband’s unaffected mother or brother or 200 control chromosomes. The second FCD proband presented with bilateral, discrete, punctate, grayish-white stromal opacities. Exonic screening of PIKFYVE revealed no causative variant. However, CNV analysis demonstrated the hemizygous deletion of exons 15 and 16.
Conclusions
We report a novel heterozygous frameshift mutation (c.3151dupA) and a CNV in PIKFYVE, representing the first CNV and the fifth frameshift mutation associated with FCD.
Introduction
Fleck corneal dystrophy (FCD) is a congenital or early-onset rare autosomal dominant disorder originally described by François and Neetens in 1957 [1,2]. Typically asymptomatic in affected individuals, FCD is associated with punctate corneal stromal opacities, corresponding to dilated keratocytes containing intracytoplasmic vesicles filled with complex lipids and glycosaminoglycans [3]. The epithelium, Bowman’s layer, Descemet’s membrane, and the endothelium are unaffected. Almost 50 years after François and Neetens published the original clinical description of FCD, genetic linkage analysis in four families identified a disease locus on chromosome 2q35 [4]. Two years later, mutations in the phosphoinositide kinase FYVE finger (PIKFYVE; Gene ID: 200576; OMIM: 609414) gene were reported in eight families with FCD [5]. Subsequently, two additional PIKFYVE mutations have been reported, bringing the total to ten (Figure 1 and Table 1) [6,7]. We report a novel PIKFYVE mutation in an individual with FCD, representing only the fifth frameshift mutation associated with FCD. In addition, we report a novel CNV identified in a second proband with FCD. The identification of a CNV in an individual affected with FCD should prompt clinicians to screen for CNVs when exonic screening does not reveal a disease-causing variant.
Figure 1.

Depiction of the PIKFYVE gene with associated functional domains found using PredictProtein. Mutations have been reported in exons 15, 16, 19, 20, and 24. The novel mutations that we report are indicated in red. Predicted binding sites and secondary structures are depicted in relation to the PIKFYVE protein. Functional domains are denoted by letters: A, zinc finger phosphoinositide kinase; B, Disheveld EGL-10, plextrin homology domains; C, β-sheet winged helix DNA/RNA-binding motif; D, cytosolic chaperone CCTγ apical domain-like motif; E, spectrin repeats; and F, C-terminal fragment similar to the common kinase core found in II β phosphatidylinositol-4-phospohate 5-kinase. Binding regions are designated by shapes: Circles represent polynucleotide-binding regions, and diamonds represent protein binding regions. Secondary structures are denoted by blue and red boxes: Blue boxes represent beta strands, and red boxes represent alpha helices.
Table 1. Reported PIKFIVE gene mutations associated with FCD.
| Reference | Exon | Mutation | Type of mutation | Protein change |
|---|---|---|---|---|
| Current study |
15 and 16 |
c.(2007+1_2008–1)_
(2190+1_2191–1) |
CNV |
p.(Ile671_Glu731)del |
| Li et al. [5] |
16 |
c.2098delA |
frameshift |
p.(Asn701fsThr*7) |
| Li et al. [5] |
16 |
c.2116_2117delCT |
frameshift |
p.(Leu706fsVal*6) |
| Li et al. [5] |
19 |
c.2551C>T |
nonsense |
p.(Arg851*) |
| Kotoulas et al. [7] |
19 |
c.2902_2905delCCTT |
frameshift |
p.(Asp1021fsThr*28) |
| Li et al. [5] |
19 |
c.2962C>T |
nonsense |
p.(Gln988*) |
| Li et al. [5] |
19 |
c.3088G>T |
nonsense |
p.(Glu1030*) |
| Li et al. [5] |
19 |
c.3112C>T |
nonsense |
p.(Arg1038*) |
| Current study |
19 |
c.3151dupA |
frameshift |
p.(Asp1052fsArg*18) |
| Li et al. [5] |
19 |
c.3308A>G |
missense |
p.(Lys1103Arg) |
| Li et al. [5] |
19 |
c.3619–1G>C |
splice-site |
p.(Val1207fsAla*11) |
| Kawasaki et al. [6] | 24 | c.4166_4169delAAGT | frameshift | p.(Glu1389fsAsp*16) |
The novel mutations that we identified are bolded.
Methods
The authors followed the tenets of the Declaration of Helsinki in the treatment of the subjects reported. The study adhered to the ARVO statement on human subjects. Study approval was obtained from the Institutional Review Board at The University of California, Los Angeles (UCLA IRB # 11–000020).
DNA collection and preparation
After informed consent was obtained, saliva samples were collected from the first proband (proband 1) and her mother, sister, and brother. Genomic DNA (gDNA) was extracted from buccal epithelial cells using the Oragene Saliva Collection Kit (DNA Genotek, Inc., Ottawa, Canada) according to the manufacturer’s instructions. A peripheral blood sample was collected into a blood collection tube containing EDTA from the antecubital fossa of the second proband (proband 2) and immediately processed for DNA isolation. Genomic DNA was prepared from peripheral blood leukocytes using the FlexiGene DNA kit (Qiagen, Valencia, CA).
PCR
All 41 exons of the PIKFYVE gene were amplified using previously described primers and conditions [5]. Briefly, each reaction was performed in a 25 μl reaction containing 12.5 μl of pDV 2.1 (2.5 mM MgSO4, 50 mM Tris-HCl pH 9.0, 0.5 Betaine monohydrate, 20 mM NH4Cl), 200 μM dNTPs, 1.5 units of RedTaq Genomic DNA polymerase (Sigma-Aldrich Corp., St. Louis, MO), 80 nM of each primer, and approximately 60 ng of genomic DNA. Samples were denatured at 95 °C for 3 min and then cycled 36 times at 95 °C for 30 s, 50–60 °C for 30 s, and 72 °C for 35 s; a final extension step at 72 °C for 10 min was performed. Thermal cycling was performed in an iCycler Thermal Cycler (Bio-Rad, Hercules, CA).
DNA sequencing
The PCR products were purified with treatment of 15–30 ng of amplicon with 5 units of Exonuclease I and 0.5 units of Shrimp Alkaline Phosphotase (USB Corp., Cleveland, OH) for 15 min at 37 °C and subsequent inactivation at 80 °C for 15 min. Sequencing reactions were performed in 10 μl reactions with Big Dye Terminator Mix v3.1 (Applied Biosystems, Foster City, CA), BDX64 enhancing buffer (MCLAB, San Francisco, CA), and 0.2 μl primer (10 pmoles/μl) following the manufacturer’s instructions. Unincorporated nucleotides were removed using the BigDye Sequencing Clean Up Kit (MCLAB) following the manufacturer’s instructions. Samples were analyzed on an ABI-3100 Genetic Analyzer (Applied Biosystems). The PIKFYVE exon and exon–intron junction sequences were compared with the NCBI Reference sequence for PIKFYVE (GenBank Accession: NM_015040.3).
Protein sequence analysis
To predict the impact of novel PIKFYVE mutations on the encoded protein, mutant PIKFYVE nucleotide sequences were translated using a DNA to protein webtool. The identified sequence variants were described according to the HGVS nomenclature guidelines. Integrated methods from PredictProtein were used to predict protein structural features: PROFphd to predict secondary structures, PHDhtm to predict transmembrane helices, PROFtmb to predict transmembrane beta-barrels, and ISIS to predict protein–protein interaction sites [8].
Copy number analysis with qPCR
Using the KAPA SYBR FAST qPCR Kit (KAPA Biosystems, Woburn, MA), quantitative PCR (qPCR) was performed on proband 2 and the control gDNA according to the manufacturer’s recommended protocol. Primers targeting or flanking PIKFYVE exons 5–7 and 13–19 and targeting intron 11 were used to determine the copy number. Primers targeting RNase P/MRP 30 kDa subunit gene (RPP30; Gene ID: 10556; OMIM: 606115) were used for normalization (Appendix 1) [9,10]. Each primer pair was run in 20 µl reactions in triplicate, with two different concentrations of DNA (4 ng/µl and 0.5 ng/µl) as the internal controls. Reactions were performed using the Lightcycler 480II (Roche, Indianapolis, IN) with the following conditions: 95 °C for 3 min and then cycled 40 times through 95 °C for 3 s, 60 °C for 20 s and 72 °C for 8 s. Subsequently, the copy number was determined as follows: 1) The cycle threshold difference (∆Ct) was calculated between the target gene (PIKFYVE) and the reference gene (RPP30) for the proband and control samples, 2) the ∆∆Ct was determined by subtracting ∆Ct(control) from ∆Ct(proband), and 3) the CNV ratio (RCNV) was calculated by using the formula: CNV ratio=2-∆∆Ct [11].
Results
Clinical findings
Case 1
A 23-year old woman (proband 1; Figure 2A, individual II:2) was referred to one of the authors (AJA) for evaluation of corneal opacities in each eye. Corrected visual acuity measured 20/20 OU, and slit-lamp examination revealed discrete, punctate stromal opacities, diffusely distributed throughout the corneal stroma of each eye (Figure 3A). No other abnormalities were noted in the cornea of either eye. The proband’s mother (Figure 2A, individual I:1) and brother (Figure 2A, individual II:3) did not demonstrate stromal opacities in either cornea. However, the proband’s sister (Figure 2A, individual II:1) demonstrated focal, gray-white stromal opacities diffusely distributed in the mid and posterior stroma of each eye. Although the opacities appeared similar to those observed in the proband, fewer opacities were noted in each cornea compared to the proband.
Figure 2.

Pedigree and PIKFYVE sequencing in a family with Fleck corneal dystrophy. A: Proband 1 is designated with an arrowhead. Filled symbols represent affected individuals, and question marks indicate individuals with undetermined affected status. B: Chromatograms demonstrate the presence of the c.3151dupA variant in proband 1 (individual II:2) and her affected sister (individual II:1); this variant was not present in the unaffected mother (individual I:1) and brother (individual II:3).
Figure 3.
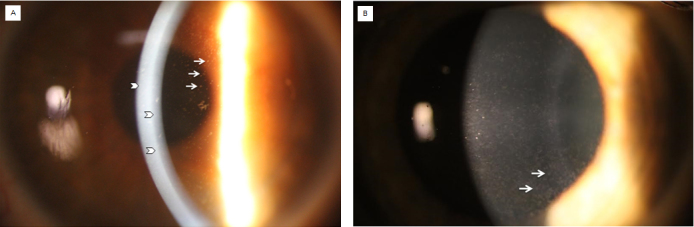
Slit-lamp photomicrograph of two individuals with Fleck corneal dystrophy. A: In proband 1, punctate discrete opacities are visible in retroillumination (arrows), and a few larger opacities are seen in the posterior stroma in direct illumination (arrowheads). B: In proband 2, discrete punctate grayish-white opacities are seen diffusely distributed in the corneal stroma (arrows).
Case 2
A 40-year-old woman (proband 2; Figure 4A, individual II:2) was referred to one of the authors (AJA) for evaluation of bilateral corneal stromal opacities. The patient had previously been diagnosed with granular corneal dystrophy, but she denied visual impairment or a family history. Uncorrected distance visual acuity measured 20/20 OU. Slit-lamp examination revealed discrete, punctate, grayish-white stromal opacities, located primarily in the corneal mid-periphery OD and throughout the cornea OS (Figure 3B).
Figure 4.

Pedigree of proband 2 and CNV analysis of PIKFYVE using qPCR. A: Proband 2 is designated with an arrowhead and the filled symbol represents the affected status of the individual. B: The CNV ratios were calculated between proband 2 and the unaffected control. PIKFYVE exons 15 and 16 demonstrated mean (minimum of 3 technical replicates) CNV ratios of approximately 0.5, which is evidence of a hemizygous deletion including these two exons. Error bars represent standard error of the mean.
PIKFYVE screening
Case 1
Sequencing of the PIKFYVE gene in the proband identified a novel heterozygous variant in exon 19, c.3151dupA, that is predicted to result in a truncated protein, p.(Asp1052fsArg*18); (Table 2). In addition, 13 known single nucleotide polymorphisms (SNPs) were identified, with minor allele frequencies varying between 0.0041 and 0.2392. Sequencing of the PIKFYVE gene in the proband’s affected sister demonstrated the same novel heterozygous variant, c. 3151dupA, which was not identified in the proband’s unaffected mother or brother or 200 control chromosomes.
Table 2. Coding region variants identified in individual with Fleck corneal dystrophy (proband 1).
| Region | Nucleotide Change | Zygosity | Type of Polymorphism | Amino Acid Change | SNP | MAF |
|---|---|---|---|---|---|---|
| Exon 16 |
2087G>A |
Homozygous |
Substitution |
Ser696Asn |
rs10932258 |
0.0064 |
| Exon 16 |
2106C>T |
Homozygous |
Substitution |
Pro702Pro |
rs10932259 |
0.062 |
| Exon 19 |
2795T>C |
Homozygous |
Substitution |
Leu932Ser |
rs2363468 |
0.0615 |
| Exon 19 |
2984A>T |
Homozygous |
Substitution |
Gln995Lys |
rs893254 |
0.0849 |
| Exon 19 |
2993C>G |
Homozygous |
Substitution |
Thr998Ser |
rs893253 |
0.0638 |
| Exon 19 |
3097T>G |
Heterozygous |
Substitution |
Ser1033Ala |
rs999890 |
0.092 |
| Exon 19 |
3114A>G |
Heterozygous |
Substitution |
Arg1038Arg |
rs999891 |
0.092 |
| Exon 19 |
3151dupA |
Heterozygous |
Duplication |
Asp1052Argfs*18 |
None |
|
| Exon 19 |
3547C>A |
Homozygous |
Substitution |
Glu1183Lys |
rs36535804
rs1529979 |
0.062 |
| Exon 19 |
3564T>C |
Homozygous |
Substitution |
Asn1188Asn |
rs1529978 |
0.0041 |
| Exon 34 |
5334G>A |
Heterozygous |
Substitution |
Thr1778Thr |
rs2304545 |
0.2392 |
| Exon 35 |
5397A>G |
Homozygous |
Substitution |
Thr1788Thr |
rs2118297 |
0.062 |
| Exon 36 |
5511T>C |
Heterozygous |
Substitution |
Ile1837Ile |
rs13020468 |
0.022 |
| Exon 36 | 5562A>G | Heterozygous | Substitution | Glu1842Glu | rs994697 | 0.062 |
Case 2
Sequencing of the PIKFYVE gene revealed ten known SNPs with minor allele frequencies varying between 0.0041 and 0.2392 but no novel variants. Since all ten of the SNPs appeared to be homozygous, with no heterozygous variants identified, a loss of heterozygosity was suspected, indicating that some, if not all, of the identified variants could be hemizygous (Table 3). To exclude this possibility, CNV analysis was performed using qPCR. Regions within exons 5–7 and 13–19 and intron 11 were analyzed. CNV ratios of approximately 0.5 were obtained for exons 15 and 16, suggestive of a heterozygous deletion in the range of approximately 2,500–9,000 nucleotides (Figure 4B).
Table 3. Coding region variants identified in individual with Fleck corneal dystrophy (proband 2).
| Region | Nucleotide Change | Zygosity | Type of Polymorphism | Amino Acid Change | SNP | MAF |
|---|---|---|---|---|---|---|
| Exon 16 |
2087G>A |
Hemizygous |
Substitution |
Ser696Asn |
rs10932258 |
0.0064 |
| Exon 16 |
2106C>T |
Hemizygous |
Substitution |
Pro702Pro |
rs10932259 |
0.062 |
| Exon 19 |
2795T>C |
Homozygous |
Substitution |
Leu932Ser |
rs2363468 |
0.0615 |
| Exon 19 |
2984A>T |
Homozygous |
Substitution |
Gln995Lys |
rs893254 |
0.0849 |
| Exon 19 |
2993C>G |
Homozygous |
Substitution |
Thr998Ser |
rs893253 |
0.0638 |
| Exon 19 |
3547C>A |
Homozygous |
Substitution |
Glu1183Lys |
rs36535804
rs1529979 |
0.062 |
| Exon 19 |
3564T>C |
Homozygous |
Substitution |
Asn1188Asn |
rs1529978 |
0.0041 |
| Exon 34 |
5334G>A |
Homozygous |
Substitution |
Thr1778Thr |
rs2304545 |
0.2392 |
| Exon 35 |
5397A>G |
Homozygous |
Substitution |
Thr1788Thr |
rs2118297 |
0.062 |
| Exon 36 | 5562A>G | Homozygous | Substitution | Glu1842Glu | rs994697 | 0.062 |
PIKFYVE structural analysis
Bioinformatic analysis of PIKFYVE predicted 18 binding sites. Three were polynucleotide-binding regions, and 15 were protein-binding regions. Secondary structures included 42 alpha helices and 44 beta strands (Figure 1). The deletion of exons 15 and 16 caused the loss of one protein binding site at amino acid 692. The frameshift mutation p.(Asp1052fsArg*18) is predicted to result in the loss of 11 binding sites (at amino acids 1072, 1100, 1104, 1158, 1188, 1357, 1655, 1738, 1926, 1957, and 2098) and the loss of 21 of the 42 alpha helices and 25 of the 44 beta strands identified toward the N-terminus of the protein.
Discussion
The PIKFYVE gene, in which mutations have been associated with FCD, spans more than 89 kbp, contains 41 coding exons, and encodes a 2,098 amino acid protein. Interestingly, the ten mutations previously associated with FCD and the two novel mutations that we report are located in only five exons of PIKFYVE (15, 16, 19, 20, and 24; Figure 1 and Table 1). Eight of the ten previously reported mutations are truncating (four frameshift and four nonsense), one is a missense, and one is a splice-site mutation. Thus, the c.3151dupA mutation that we report is only the fifth frameshift mutation associated with FCD.
In two previously reported families, affected individuals were diagnosed with the characteristic FCD phenotype but did not demonstrate a PIKFYVE coding region mutation [5]. Given our identification of a partial PIKFYVE deletion in an individual with FCD, we recommend screening for CNV when sequencing of the PIKFYVE coding region fails to identify a presumed pathogenic variant.
The protein encoded by PIKFYVE has been shown to regulate the vesicle size of early/late endosomes in Golgi-dependent transport [12]. There are six reported functional domains in the PIKFYVE gene, located between amino acids 150–210 and 1791–2085 (Figure 1) [5]. The novel frameshift mutation that we report, p.(Asp1052fsArg*18), is predicted to result in a truncated protein with loss of two important functional domains: spectrin repeats at amino acids 1490–1538 and 1679–1729 and a C-terminal fragment similar to the common kinase core found in II β-phosphatidylinositol-4-phospohate 5-kinase at amino acids 1791–2085. Spectrin repeats binding sites are important for multiprotein interactions and cytoskeleton structure [13], and thus, the loss of this functional domain may affect important protein interactions and the overall structure of PIKFYVE, resulting in a loss of function of the encoded protein. The common kinase core of β-phosphatidylinositol-4-phosphate 5-kinase synthesizes phosphatidylinositol-4,5-bisphosphate, which is involved in vesicle trafficking and cytoskeletal organization [14]. The loss of this domain may affect the ability of PIKFYVE to regulate transport vesicles in the Golgi apparatus, potentially leading to accumulation of the intracytoplasmic vesicles within the dilated keratocytes that characterize FCD.
The heterozygous deletion of a region within PIKFYVE, involving exons 15 and 16, is the first CNV associated with FCD and is predicted to result in the splicing of exons 14 and 17. The cytosolic chaperone CCTγ apical domain-like motif, located at amino acids 677–843 in exons 15–19, is essential for proper folding and stability of cytosolic proteins in eukaryotes [15]. Therefore, the loss of this structural domain will likely affect the proper folding of the PIKFYVE protein, and thus, the overall protein function. In silico analysis of the wild-type PIKFYVE protein sequence identified multiple predicted protein binding sites that would be disrupted or lost secondary to the frameshift mutation and heterozygous deletion that we describe. In addition, the secondary structure of the PIKFYVE protein is predicted to be significantly affected by the p.(Asp1052fsArg*18) frameshift mutation with loss of 21 of the 42 alpha helices and 25 of the 44 beta strands, which are essential to the overall stability of the protein [16].
Acknowledgments
Support provided by National Eye Institute grants R01EY022082 (A.J.A.), P30EY000331 (core grant) and an unrestricted grant from Research to Prevent Blindness The authors have no conflict-of-interest or financial disclosures.
Appendix 1. Primers used for copy number variation analysis by qPCR.
* The numbering of exons and introns starts with the first coding exon. ** Sequences obtained from the Harvard's Harvard PrimerBank. PrimerBank ID: 295789161c3. *** Sequences obtained from Szantai et al. 2009. To access the data, click or select the words “Appendix 1.” This will initiate the download of a compressed (pdf) archive that contains the file.
References
- 1.Francois J, Neetens A. New hereditary-familial dystrophy of the corneal parenchyma (spotted hereditary dystrophy). Bull Soc Belge Ophtalmol. 1957;114:641–6. [PubMed] [Google Scholar]
- 2.Weiss JS, Moller HU, Aldave AJ, Seitz B, Bredrup C, Kivela T, Munier FL, Rapuano CJ, Nischal KK, Kim EK, Sutphin J, Busin M, Labbe A, Kenyon KR, Kinoshita S, Lisch W. IC3D Classification of Corneal Dystrophies-Edition 2. Cornea. 2015;34:117–59. doi: 10.1097/ICO.0000000000000307. [DOI] [PubMed] [Google Scholar]
- 3.Purcell JJ, Jr, Krachmer JH, Weingeist TA. Fleck corneal dystrophy. Arch Ophthalmol. 1977;95:440–4. doi: 10.1001/archopht.1977.04450030082009. [DOI] [PubMed] [Google Scholar]
- 4.Jiao X, Munier FL, Schorderet DF, Zografos L, Smith J, Rubin B, Hejtmancik JF. Genetic linkage of Francois-Neetens fleck (mouchetee) corneal dystrophy to chromosome 2q35. Hum Genet. 2003;112:593–9. doi: 10.1007/s00439-002-0905-1. [DOI] [PubMed] [Google Scholar]
- 5.Li S, Tiab L, Jiao X, Munier FL, Zografos L, Frueh BE, Sergeev Y, Smith J, Rubin B, Meallet MA, Forster RK, Hejtmancik JF, Schorderet DF. Mutations in PIP5K3 are associated with Francois-Neetens mouchetee fleck corneal dystrophy. Am J Hum Genet. 2005;77:54–63. doi: 10.1086/431346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kawasaki S, Yamasaki K, Nakagawa H, Shinomiya K, Nakatsukasa M, Nakai Y, Kinoshita S. A novel mutation (p.Glu1389AspfsX16) of the phosphoinositide kinase, FYVE finger containing gene found in a Japanese patient with fleck corneal dystrophy. Mol Vis. 2012;18:2954–60. [PMC free article] [PubMed] [Google Scholar]
- 7.Kotoulas A, Kokotas H, Kopsidas K, Droutsas K, Grigoriadou M, Bajrami H, Schorderet DF, Petersen MB. A novel PIKFYVE mutation in fleck corneal dystrophy. Mol Vis. 2011;17:2776–81. [PMC free article] [PubMed] [Google Scholar]
- 8.Yachdav G, Kloppmann E, Kajan L, Hecht M, Goldberg T, Hamp T, Honigschmid P, Schafferhans A, Roos M, Bernhofer M, Richter L, Ashkenazy H, Punta M, Schlessinger A, Bromberg Y, Schneider R, Vriend G, Sander C, Ben-Tal N, Rost B. PredictProtein–an open resource for online prediction of protein structural and functional features. Nucleic Acids Res. 2014;42:W337–43. doi: 10.1093/nar/gku366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Spandidos A, Wang X, Wang H, Seed B. PrimerBank: a resource of human and mouse PCR primer pairs for gene expression detection and quantification. Nucleic Acids Res. 2010;38(Database issue):D792–9. doi: 10.1093/nar/gkp1005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Szantai E, Elek Z, Guttman A, Sasvari-Szekely M. Candidate gene copy number analysis by PCR and multicapillary electrophoresis. Electrophoresis. 2009;30:1098–101. doi: 10.1002/elps.200800755. [DOI] [PubMed] [Google Scholar]
- 11.Ji H, Lu J, Wang J, Li H, Lin X. Combined examination of sequence and copy number variations in human deafness genes improves diagnosis for cases of genetic deafness. BMC Ear Nose Throat Disord. 2014;14:9. doi: 10.1186/1472-6815-14-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Nicot AS, Laporte J. Endosomal phosphoinositides and human diseases. Traffic. 2008;9:1240–9. doi: 10.1111/j.1600-0854.2008.00754.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Djinovic-Carugo K, Gautel M, Ylanne J, Young P. The spectrin repeat: a structural platform for cytoskeletal protein assemblies. FEBS Lett. 2002;513:119–23. doi: 10.1016/s0014-5793(01)03304-x. [DOI] [PubMed] [Google Scholar]
- 14.Toker A. The synthesis and cellular roles of phosphatidylinositol 4,5-bisphosphate. Curr Opin Cell Biol. 1998;10:254–61. doi: 10.1016/s0955-0674(98)80148-8. [DOI] [PubMed] [Google Scholar]
- 15.Fares MA, Wolfe KH. Positive selection and subfunctionalization of duplicated CCT chaperonin subunits. Mol Biol Evol. 2003;20:1588–97. doi: 10.1093/molbev/msg160. [DOI] [PubMed] [Google Scholar]
- 16.Dietz H, Rief M. Exploring the energy landscape of GFP by single-molecule mechanical experiments. Proc Natl Acad Sci USA. 2004;101:16192–7. doi: 10.1073/pnas.0404549101. [DOI] [PMC free article] [PubMed] [Google Scholar]