

miRNA miR528 affects plant development and plant response to salt stress and nitrogen starvation.
Abstract
MicroRNA528 (miR528) is a conserved monocot-specific small RNA that has the potential of mediating multiple stress responses. So far, however, experimental functional studies of miR528 are lacking. Here, we report that overexpression of a rice (Oryza sativa) miR528 (Osa-miR528) in transgenic creeping bentgrass (Agrostis stolonifera) alters plant development and improves plant salt stress and nitrogen (N) deficiency tolerance. Morphologically, miR528-overexpressing transgenic plants display shortened internodes, increased tiller number, and upright growth. Improved salt stress resistance is associated with increased water retention, cell membrane integrity, chlorophyll content, capacity for maintaining potassium homeostasis, CATALASE activity, and reduced ASCORBIC ACID OXIDASE (AAO) activity; while enhanced tolerance to N deficiency is associated with increased biomass, total N accumulation and chlorophyll synthesis, nitrite reductase activity, and reduced AAO activity. In addition, AsAAO and COPPER ION BINDING PROTEIN1 are identified as two putative targets of miR528 in creeping bentgrass. Both of them respond to salinity and N starvation and are significantly down-regulated in miR528-overexpressing transgenics. Our data establish a key role that miR528 plays in modulating plant growth and development and in the plant response to salinity and N deficiency and indicate the potential of manipulating miR528 in improving plant abiotic stress resistance.
Abiotic stresses, especially drought, salt, and nitrogen (N) deficiency, are limiting factors for plant growth, development, and agricultural productivity. To cope with drought and salt stresses, plants have evolved similar strategies of osmotic adjustment (Munns, 2002; Zhu, 2002). Plants also evolved salinity-specific adjustments against ionic disequilibrium, which encompass excluding salt entry into plants and compartmentalizing ions into vacuoles or old leaves (Munns, 1993; Yeo, 1998). Another worldwide limiting factor for crop yields is N deficiency, which triggers reduced leaf growth rate and photosynthetic rate (Chapin et al., 1988). Due to the sessile nature of plants, abiotic stresses are unavoidable. Therefore, it is critical to develop reliable procedures to genetically modify plants for improved performance under environmental stresses, thereby enhancing agricultural productivity to meet the ever-growing demands in food production.
Genetic engineering plays an increasingly important role in agronomic trait modifications in crop species. Currently, many genes encoding functional proteins, transcription factors, and proteins involved in signaling pathways have been identified as abiotic stress-responsive genes (Shinozaki and Yamaguchi-Shinozaki, 2007; Masclaux-Daubresse et al., 2010; Turan et al., 2012). Constitutive expression of some of these genes in transgenic plants has been demonstrated to lead to enhanced salt or drought tolerance (Golldack et al., 2011; Kim et al., 2013; Lu et al., 2013; Li et al., 2014). However, details of the regulatory network in the plant stress response remain elusive. To improve plant performance under N deficiency conditions, substantial efforts have been concentrated on understanding the physiological and molecular processes determining plant nitrogen use efficiency (NUE), including those involved in N uptake, assimilation, translocation, and remobilization. A number of crop plants have been genetically engineered to alter single functional genes in these pathways with limited success, due to posttranscriptional regulation machineries (Ferrario-Mery et al., 1998; Fraisier et al., 2000; Pathak et al., 2008). Abiotic stresses trigger a wide array of plant morphological and physiological responses. Therefore, a comprehensive understanding of plant responses to abiotic stresses and the knowledge of how to manipulate multiple genes concurrently to integrate the responses of plants at various levels under abiotic stresses are keys to genetically improving plants for enhanced performance under adverse environmental conditions.
Since the discovery of microRNAs (miRNAs), the important small regulatory molecules, 2 decades ago, the roles that some of their members play in the complex stress response network have been gradually recognized. Recently, an evolutionarily conserved monocot-specific miRNA, miR528, was identified to respond to multiple stresses, including salt, drought, low-temperature, submergence, nitrate starvation, and arsenite and arsenate stresses in maize (Zea mays), sugarcane (Saccharum officinarum), phalaenopsis orchid (Phalaenopsis aphrodite), and rice (Oryza sativa; Zhang et al., 2008; An et al., 2011; Xu et al., 2011; Ferreira et al., 2012; Nischal et al., 2012; Sharma et al., 2015). In addition, deep sequencing analysis shows that miR528 responds to diazotrophic bacteria in maize and might be involved in cytoplasmic male sterility in rice anther (Thiebaut et al., 2014; Yan et al., 2015). The miR528 in many grass species, such as rice, maize, sugarcane, sorghum (Sorghum bicolor), and Brachypodium distachyon, has been shown to have identical mature sequence (UGGAAGGGGCAUGCAGAGGAG; http://www.mirbase.org/; Kozomara and Griffiths-Jones, 2014), suggesting that miR528 in different monocot species may function similarly in regulating target gene activity. Eleven putative targets of miR528 in rice have been predicted using in silico analysis. Some of them are involved in oxidation-reduction processes (Dai and Zhao, 2011), implying that miR528 may function as a multistress integrator. However, all the stress-responsive data of miR528 expression obtained so far have been from genome-wide large-scale analyses. Experimental evidence is still lacking.
In this study, we analyzed the role of miR528 in regulating the plant response to salinity and N starvation and the underlying physiological and molecular mechanisms using transgenic analysis in an important perennial grass species, creeping bentgrass (Agrostis stolonifera). Using stem-loop reverse transcription-(RT) quantitative PCR (RT-qPCR), we first demonstrated that, like its counterpart in annual species, miR528 in creeping bentgrass is also responsive to salinity stress and water and N deficiency. We then generated transgenic creeping bentgrass plants overexpressing rice primary miR528 (pri-miR528) to evaluate the impact of miR528 on plant development and the response to environmental stress. Our data indicate that transgenic plants exhibit altered growth and development and enhanced tolerance to salt stress and N deficiency. Improved salt stress resistance is associated with increased water retention, cell membrane integrity, chlorophyll content, capacity for maintaining potassium (K) homeostasis, and CATALASE (CAT) activity and with reduced ASCORBIC ACID OXIDASE (AAO) activity; while enhanced tolerance to N starvation is associated with increased biomass, total N accumulation and chlorophyll synthesis, and NITRITE REDUCTASE (NiR) activity and with reduced AAO activity. Our findings highlight a functional role for miR528 in modulating ionic equilibrium, NUE, and the oxidation-reduction process to mediate plant responses to salinity stress and N starvation. Additionally, we also identified two creeping bentgrass miR528 targets, AsAAO and COPPER ION BINDING PROTEIN1 (AsCBP1), which are homologs of rice AAO and CBP1 and implicated in the oxidation-reduction process. We found that both AsAAO and AsCBP1 respond to salinity stress and nitrogen starvation and are significantly down-regulated in miR528-overexpressing transgenic plants. Collectively, our data demonstrate the importance of miR528 in plant development and its key role as a multistress integrator controlling plant responses to salinity and N starvation, pointing to the potential of manipulating miR528 in improving plant abiotic stress resistance.
RESULTS
miR528 Responds to Salt, Drought, and N Deficiency Stresses
miR528 has previously been implicated in plant responses to drought, salt, and N deficiency in annual species (Ding et al., 2009; Xu et al., 2011; Ferreira et al., 2012; Nischal et al., 2012). To study its role in perennial grass species, we first examined its expression in response to various abiotic stresses in creeping bentgrass. Stem-loop RT-qPCR analysis indicates that miR528 is significantly induced by both drought and salt stresses but suppressed under N starvation (Fig. 1). This suggests that miR528 is regulated by abiotic stress, consistent with observations in rice, maize, and sugarcane (Ding et al., 2009; Xu et al., 2011; Ferreira et al., 2012; Nischal et al., 2012).
Figure 1.

Stem-loop RT-qPCR analyses of miR528 expression profiles in response to salt, drought, and N deficiency in wild-type creeping bentgrass leaves. Relative expression levels of the mature miR528 were determined in wild-type plants under salt (A), drought (B), and N deficiency (C) treatment. The relative changes of gene expression were calculated based on the comparative threshold cycle method (Livak and Schmittgen, 2001). UBIQUITIN5 (AsUBQ5) was used as an endogenous control. Data are presented as means of three biological replicates × three technical replicates, and error bars represent se. Asterisks indicate a significant difference of expression levels between untreated and each abiotic-stress treated wild-type plant: **, P < 0.01 and ***, P < 0.001 by Student’s t test.
Generation and Molecular Analysis of Transgenic Creeping Bentgrass Overexpressing Osa-miR528, a Rice miRNA Gene
To further study the role that miR528 plays in plant adaption to abiotic stress, we prepared a miR528 overexpression construct and introduced it into the genome of wild-type creeping bentgrass through Agrobacterium tumefaciens-mediated transformation. The rice complementary DNA (cDNA) containing precursor miR528 (pre-miR528; AK073820; Knowledge-Based Oryza Molecular Biological Encyclopedia; http://cdna01.dna.affrc.go.jp/cDNA/; Kikuchi et al., 2003; Liu et al., 2005; Satoh et al., 2007) was amplified and then cloned into the binary vector pZH01, generating the Osa-miR528 overexpression gene construct, p35S-Osa-miR528/p35S-Hyg. As shown in Figure 2A, the Osa-miR528 gene was under the control of the CaMV 35S promoter and linked to the hygromycin resistance gene, Hyg, driven by the CaMV 35S promoter. To select positive transgenic plants containing miR528 overexpression constructs, we amplified the Hyg gene by PCR with genomic DNA of regenerated plants and identified a total of 13 transgenic lines (Fig. 2B), which were morphologically indistinguishable from each other. Three transgenic lines, TG6, TG8, and TG13, were chosen for further characterization of the aspects of plant development and stress response. To verify the Osa-miR528 expression in transgenic plants, we conducted RT-PCR analysis to amplify the cDNA containing the Osa-miR528 stem-loop structure, comparing the levels of primary Osa-miR528 transcripts between the wild-type control and three transgenic lines. Interestingly, besides a fragment with the expected size of 549 bp, two additional bands of 451 and 240 bp were also detected (Fig. 2C). Cloning and sequencing analysis of these DNA fragments revealed that alternative splicing mechanisms are responsible for the production of the additional two bands of 451 and 240 bp (data not shown). The same splicing pattern of the pri-miR528 observed in rice (Fig. 2C) suggests that molecular mechanisms governing pri-miR528 sequence processing are conserved in different plant species. An additional pair of primers was then designed to amplify a common region of the three alternatively spliced pri-miR528 transcripts in transgenic lines using semiquantitative RT-PCR analysis. As demonstrated in Figure 2D for the five representative transgenic lines, the transcripts of pri-miR528 are significantly elevated in transgenic lines compared with the wild-type controls (Fig. 2D). To determine whether pri-miR528 can be successfully processed into mature miRNA, we conducted stem-loop RT-qPCR analyses and found that the expression levels of mature Osa-miR528 in three representative transgenic lines are significantly higher than that in the wild-type control (Fig. 2E), suggesting that the primary RNA sequence of Osa-miR528 from rice is properly processed into mature miRNA in creeping bentgrass.
Figure 2.

Generation and molecular analysis of transgenic creeping bentgrass overexpressing Osa-miR528. A, Schematic diagram of the Osa-miR528 gene overexpression construct, p35S-Osa-miR528/p35S-Hyg. Osa-miR528 is under the control of the Cauliflower mosaic virus (CaMV) 35S promoter and linked to the hygromycin resistance gene, Hyg, driven by the CaMV 35S promoter. LB, Left border; NOS, nopaline synthase terminator; RB, right border. B, PCR analysis to amplify the Hyg gene using genomic DNA of wild-type (WT) and transgenic creeping bentgrass to determine the integration of Osa-miR528 in the host genome. C, Pri-miR528 shows alternative splicing in wild-type rice and transgenic creeping bentgrass. The same splicing pattern of pri-miR528 was observed in both rice and creeping bentgrass. Blue, red, and purple lines represent different splicing patterns. Yellow lines are the locations of pre-miR528. Black arrows indicate the position of mature miR528. D, An additional pair of primers was designed to amplify a common region of the three alternatively spliced pri-miR528 transcripts using semiquantitative RT-PCR analysis to compare the expression levels of primary Osa-miR528 in wild-type and transgenic plants. E, Stem-loop RT-qPCR analysis to detect the expression of mature Osa-miR528 in transgenic and wild-type plants. Relative changes in gene expression were calculated based on the comparative threshold cycle method (Livak and Schmittgen, 2001). ACTIN1 (AsACT1) was used as an endogenous control. Data are presented as means of three technical replicates, and error bars represent se. Asterisks indicate a significant difference of expression levels between the wild type and each transgenic line: **, P < 0.01 and ***, P < 0.001 by Student’s t test.
Overexpression of miR528 Alters Plant Development
To determine the involvement of miR528 in plant development, we analyzed wild-type and transgenic plants initiated from a single tiller in pure sand. Transgenic plants produced significantly more, but shorter, tillers than wild-type controls (Fig. 3, A, C, and J; Supplemental Fig. S1, A and B), especially at the later developmental stage (10 weeks old; Fig. 3J). However, no significant difference in the total numbers of shoots, primary and secondary tillers from a crown, and internodes was observed between transgenic and control plants at the later developmental stages (60 and 90 d old; Fig. 3K). The developmental changes observed in transgenic plants were further confirmed by comparing wild-type and Osa-miR528 transgenic plants grown in the same pot filled with soil (Fig. 3B). In addition, transgenic plants exhibited more upright tiller growth than wild-type controls (Fig. 3B; Supplemental Fig. S1C).
Figure 3.

Plant tillering and development. A, Ten-week-old wild-type (WT) and transgenic (TG) plants initiated from a single tiller. Bar = 10 cm. B, Two-month-old wild-type and transgenic plants initiated from the same number of tillers were grown in the same 6-inch pot. Bar = 10 cm. C, Closeup of the longest tillers from wild-type and transgenic plants, respectively. Bar = 5 cm. D, All internodes from the representative longest tiller were sliced from top to bottom and arranged from left to right. Bar = 5 cm. E, Top three fully developed leaves from the representative tillers of wild-type and transgenic plants. Bar = 2 cm. F, Cross-section images of wild-type and transgenic leaves. Bar = 200 μm. G, Cross-section images of wild-type and transgenic stems. Bar = 100 μm. H, Statistical analysis of leaf thickness between representative wild-type and transgenic plants (n = 8). I, Statistical analysis of the number of vascular bundles between representative wild-type and transgenic stems (n = 8). J, Tiller number in wild-type and transgenic plants 5 and 10 weeks after initiation from a single tiller (n = 5). K, Total shoot number including both tillers and lateral shoots in wild-type and transgenic plants at 30, 60, and 90 d after initiation from a single tiller (n = 5). L, Average length of the top eight internodes from wild-type and transgenic tillers (n = 6). Data are presented as means, and error bars represent se. Asterisks indicate a significant difference between the wild type and each transgenic line: *, P < 0.05; **, P < 0.01; and ***, P < 0.001 by Student’s t test. Tillers are grass shoots growing out from the crown at the base of the plants. Shoots include both tillers and lateral shoots growing out from the tillers.
To further study what causes the reduced tiller length in transgenics, we analyzed the average length and number of internodes of the representative tillers from wild-type and transgenic plants (Fig. 3D). We found that the total numbers of internodes in wild-type and transgenic tillers are similar, whereas the average length of the internodes from each tiller in transgenic plants is significantly reduced compared with wild-type controls (Fig. 3L).
Transgenic and wild-type leaves and stems were also compared at the cellular level via histological analysis (Fig. 3, E–G). Transgenic leaves were significantly thicker than wild-type leaves (Fig. 3H), and the number of stem vascular bundles was significantly increased in transgenics compared with that in wild-type controls (Fig. 3I).
The potential impact of miR528 on plant growth was investigated by measuring the shoot and root biomass of 10-week-old wild-type and transgenic plants initiated from a single tiller and the weekly clipping weight thereafter continuously for 4 weeks. Our statistical analyses revealed no significant difference in biomass accumulation between transgenic and control plants (Supplemental Fig. S2).
Overexpression of miR528 Leads to Enhanced Salt Tolerance in Transgenic Plants
To investigate the role that miR528 plays in the plant stress response, we examined transgenic and wild-type control plants under salinity stress using fully developed plants (Fig. 4A). Figure 4, B and C, shows plants recovering for 8 d after a 9-d exposure to 200 mm NaCl. Salt-elicited tissue damage was significantly more pronounced in wild-type controls than in transgenic plants, suggesting an enhanced tolerance to salt stress in miR528-expressing transgenic plants.
Figure 4.

Responses of wild-type controls and transgenics to salinity treatment. A, Wild-type controls (WT) and two transgenic (TG) lines initiated from the same number of tillers were trimmed to the same height before the salt stress test. B, Fully developed wild-type and transgenic plants were subjected to 200 mm NaCl treatment. The performance of wild-type and transgenic plants at 8 d after recovery from a 9-d salt treatment is shown. C, Closeup of representative wild-type and transgenic plants from B. D, Electrolyte leakage values were calculated before and after a 9-d salt treatment. E, Relative water contents were measured before and after a 9-d salt treatment. F, Pro contents of wild-type and transgenic leaves measured before and after a 200 mm NaCl treatment. G to I, Chlorophyll a content (G), chlorophyll b content (H), and total chlorophyll content (I) were measured before and after a 9-d salt treatment. fw, Fresh weight. Data are presented as means (n = 5), and error bars represent se. Asterisks indicate a significant difference between the wild type and each transgenic plant: *, P < 0.05; **, P < 0.01; and ***, P < 0.001 by Student’s t test.
Osa-miR528 Transgenics Exhibit Better Water Retention and Cell Membrane Integrity, and Higher Pro and Chlorophyll Contents, Than Wild-Type Controls under Salt Stress
Salinity stress damages the plant cell membrane and turgidity. Therefore, the maintenance of cell membrane integrity and water status is considered a major component in plant salt stress tolerance. To investigate the degree of cell membrane injury between wild-type and transgenic plants, we measured their electrolyte leakage (EL). Under normal growth conditions, there was no significant difference in EL between the wild type and two transgenic lines. After a 9-d salt stress treatment, the EL value of the wild type was significantly higher than that of transgenic plants (Fig. 4D), indicating that transgenic plants have a better capability of maintaining cell membrane integrity than wild-type controls under salt stress conditions. To compare the water status in wild-type and transgenic plants, we measured the relative water content (RWC). Both transgenics and wild-type controls displayed similar RWC under normal growth conditions (Fig. 4E). However, when subjected to salinity stress for 9 d, transgenics had significantly higher RWC than wild-type controls (Fig. 4E), implying that transgenic plants have improved ability to retain water under salinity stress.
Pro is essential for plant primary metabolism under salt stress. It functions as a molecular chaperone in buffering the pH of the cytosolic redox status within the cell and in reactive oxygen species (ROS) scavenging (Ashraf and Foolad, 2007). Pro contents in both transgenics and wild-type controls were similar before salt treatment (Fig. 4F). However, significantly higher Pro was accumulated in transgenics than in wild-type controls after the salt stress (Fig. 4F), suggesting an enhanced ROS detoxification capacity under osmotic stress in transgenic plants.
Leaf chlorophyll content is also affected under salt stress due to the destruction of the chlorophyll pigment-protein complex, the degradation of the chlorophyll enzyme chlorophyllase, and the interference in the synthesis of chlorophyll structural components (Rao and Rao, 1981; Ali et al., 2004). In this study, there was no significant difference of chlorophyll contents between wild-type and transgenic plants under normal growth conditions, whereas all three transgenic lines showed significantly higher chlorophyll contents than wild-type controls during salt treatment (Fig. 4, G–I), suggesting the possible role the improved photosynthesis of the transgenics plays in contributing to enhanced salt stress resistance.
Osa-miR528 Transgenic Plants Maintain Cellular K+ Homeostasis under Salt Stress
Salt stress imposes ionic imbalance and osmotic stress on plants due to elevated Na+ levels around plant roots. To compare the Na+ uptake in wild-type and Osa-miR528 transgenic plants, Na+ relative contents were measured. While there was no difference in Na+ content in roots between transgenics and wild-type controls, the three transgenic lines accumulated significantly more Na+ than wild-type controls in shoots before the salt stress (Fig. 5A). After the salt treatment, wild-type and transgenic plants had similar Na+ contents in both shoots and roots (Fig. 5B). K plays an essential role in diverse physiological processes, including turgor adjustment, stomata movement, cell elongation, and activation of more than 50 cytoplasmic enzymes (Marschner, 1995; Gambale and Uozumi, 2006; Lebaudy et al., 2007). Salinity also affects K+ homeostasis, because Na+ competes with K+ for binding sites during enzymatic reactions and protein synthesis in the cytoplasm, where K+ functions as a cofactor in these processes (Marschner, 1995). Our result showed that K+ relative contents in wild-type and transgenic shoots were similar or slightly higher in transgenic shoots before salt stress (Fig. 5C). After salinity treatment, interestingly, transgenics maintained their shoot K+ level, whereas the K+ levels in wild-type shoots dropped dramatically, becoming significantly lower than those in transgenic shoots (Fig. 5D). Transgenics also contained higher K+ in roots than wild-type plants, although the difference was insignificant (Fig. 5, C and D). One of the key elements in plant salinity tolerance is the capacity of maintaining a high K+:Na+ ratio. Under normal growth conditions, wild-type shoots had significantly a higher K+:Na+ ratio than transgenics due to their lower Na+ contents than transgenic shoots (Fig. 5E). After salt stress treatment, however, K+:Na+ ratios of shoots and roots were both significantly higher in transgenics than in wild-type controls (Fig. 5F). Figure 5G shows that, under salt stress, transgenics were capable of maintaining similar shoot K+ levels to nonstressed conditions compared with wild-type controls. However, K+ levels in both wild-type and transgenic roots decreased dramatically, although transgenic roots had higher K+ contents than wild-type controls under nonstressed conditions (Fig. 5H).
Figure 5.

Na+ and K+ contents in wild-type and transgenic plants under normal and salt stress conditions. A, Na+ relative contents in shoot and root tissues of wild-type (WT) and transgenic (TG) plants before salinity treatment. B, Na+ relative contents in shoot and root tissues of wild-type and transgenic plants 9 d after salinity treatment. C, K+ relative contents in shoot and root tissues of wild-type and transgenic plants under normal growth conditions. D, K+ relative contents in shoot and root tissues of wild-type and transgenic plants 9 d after salinity treatment. E, K+:Na+ ratio in shoots and roots of wild-type and transgenic plants before a 200 mm NaCl treatment. F, K+:Na+ ratio in shoots and roots of wild-type and transgenic plants 9 d after salt treatment. G, Shoot K+ relative contents in wild-type and transgenic plants before and after salinity stress. H, Root K+ relative contents in wild-type and transgenic plants before and after salinity stress. Data are presented as means (n = 3), and error bars represent se. Asterisks indicate significant differences in K+ content, Na+ content, or K+:Na+ ratio between the wild type and each transgenic line: *, P < 0.05; **, P < 0.01; and ***, P < 0.001 by Student’s t test.
The Impact of miR528 on the Expression of the K Transporter Creeping Bentgrass High Affinity Potassium Transporter5
Differences in Na+ and K+ contents between wild-type and transgenic plants imply that miR528 might mediate the concerted action of ion transport systems. To investigate the underlying mechanism of miR528-mediated ion transport, K transporter genes in creeping bentgrass were identified, and their expression was analyzed in transgenic and wild-type plants. Previous studies indicate that there are mainly seven gene families involved in K+ uptake (Mäser et al., 2001; Véry and Sentenac, 2002, 2003), of which functionally characterized genes encoding K-permeable channels and K transporters were selected for further study. Creeping bentgrass High Affinity Potassium Transporter5 (AsHAK5; KR911825) from the K Uptake Permease/HAK/K Transporter family was successfully amplified in creeping bentgrass and found to be up-regulated in transgenic leaves and roots compared with wild-type controls (Fig. 6), suggesting that constitutive expression of miR528 leads to enhanced K transporter activity and contributes to the increased K+ uptake and enhanced capacity of maintaining K+ homeostasis in transgenic plants.
Figure 6.
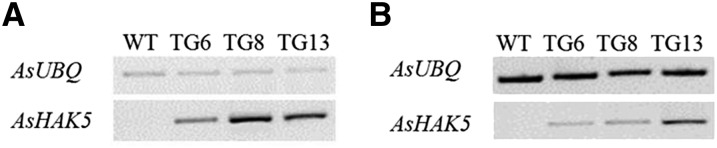
Expression levels of AsHAK5 in wild-type and transgenic plants by semiquantitative RT-PCR analysis. A, Expression levels of AsHAK5 in leaf tissues of wild-type (WT) and transgenic (TG) plants under normal growth conditions. B, Expression levels of AsHAK5 in root tissues of wild-type and transgenic plants under normal growth conditions. AsUBQ5 was used as the endogenous control.
Overexpression of miR528 Results in Enhanced ROS Scavenging Associated with Increased CAT Activity But Decreased AAO Activity under Salt Stress in Transgenics
In addition to the impact on K homeostasis, salt stress also causes the accumulation of ROS in plant cells. Plants evolve stress tolerance mechanism of ROS detoxification via increasing antioxidant enzyme activity. Since the predicted targets of Osa-miR528 contain many antioxidant enzyme-encoding genes, this prompted us to examine how miR528 impacts antioxidant enzyme activity. We first measured the activity of the zinc/copper (Zn/Cu) SUPEROXIDE DISMUTASE (SOD) enzyme encoded by a predicted miR528 target gene, but we observed no significant difference between wild-type and transgenic plants (Supplemental Fig. S3A). SOD catalyzes the dismutation of superoxide radical into oxygen or hydrogen peroxide (H2O2), and the latter is further decomposed to water and oxygen by CAT. Therefore, we conducted a CAT assay and observed that transgenic plants have significantly higher CAT activity than wild-type controls under both normal and salt stress conditions, suggesting an increased capacity of ROS scavenging in miR528 transgenic plants (Supplemental Fig. S3B). Another predicted miR528 target encodes AAO, which catalyzes the reaction of ascorbate (AA) oxidation. Under salt stress, transgenic plants exhibited significantly lower AAO activity than wild-type controls, indicating that transgenics maintain higher levels of AA under redox status, which contributes to a better elimination of ROS than in wild-type controls (Supplemental Fig. S3C).
miR528 Transgenics Exhibit an Enhanced NUE Associated with Up-Regulated Gene Expression and Increased Enzyme Activity of Nitrite Reductase
To examine the responses of wild-type and transgenic plants under different N supplies, we first determined the optimum N concentration for creeping bentgrass by applying Murashige and Skoog (MS) nutrient solution containing 2, 10, or 40 mm N to wild-type and transgenic plants. Four weeks later, wild-type and transgenic plants had the fastest and slowest growth rate, with 10 and 2 mm N solution treatments, respectively (Fig. 7, B, E, and F). Thus, 10 mm is the optimum N concentration in our experiment and was used for further analysis. The result also shows that the excess N level of 40 mm reduces plant growth (Fig. 7B) due to the decreased uptake of other nutrient elements, such as phosphorus and K. When comparing plant growth between transgenics and wild-type controls, no significant difference in shoot biomass was observed under the normal N supply of 10 mm. However, transgenics grew better and produced significantly more shoot biomass than wild-type controls under both N deficiency (2 mm) and overfertilized (40 mm) conditions (Fig. 7, E and F). Moreover, transgenics also exhibited delayed leaf senescence compared with wild-type controls. While wild-type controls exhibited wilting leaf tips under all three N fertilization treatments, transgenic plants barely had this symptom, suggesting a role that miR528 may play in plant leaf senescence and longevity (Supplemental Fig. S4).
Figure 7.

Responses of wild-type and transgenic plants under different concentrations of N solutions. A, Wild-type (WT) and transgenic (TG) plants were trimmed to be uniform before applying N solutions. B, Performance of wild-type controls and three transgenic lines on MS solutions containing 2, 10, or 40 mm N for 4 weeks. C and D, Closeup of wild-type and transgenic shoots under 2 mm (C) and 40 mm (D) N solution treatment for 4 weeks. E and F, Shoot fresh weight (E) and dry weight (F) of wild-type and transgenic plants after 4 weeks of growth with three different N solutions. G, Percentage of shoot total N content of wild-type and transgenic plants measured 4 weeks after applying different N solutions. H, Weight of shoot total N was measured 4 weeks after applying different N solutions. I to K, Plant chlorophyll contents, including chlorophyll a (I), chlorophyll b (J), and total chlorophyll (J), were measured 4 weeks after applying different concentrations of N solutions. fw, Fresh weight. Data are presented as means (n = 4), and error bars represent se. Asterisks indicate significant differences of biomass value, total N weight, or chlorophyll contents between wild-type and transgenic plants: *, P < 0.05; **, P < 0.01; and ***, P < 0.001 by Student’s t test.
To further evaluate the impact of miR528 on the plant response to N nutrients, we measured plant leaf chlorophyll contents in both transgenic and wild-type plants under N starvation and normal conditions. In comparison with N-sufficient plants, plants under N deficiency conditions (0.4 and 2 mm) had reduced chlorophyll contents, especially under 0.4 mm N supply (Fig. 7, I–K). While transgenic plants were similar to wild-type controls in chlorophyll content under N-sufficient conditions, they had significantly higher chlorophyll than wild-type controls under N-starved conditions (Fig. 7, I–K), suggesting a lower degree of chlorophyll degradation and, therefore, likely a higher photosynthetic capability in transgenic plants than in wild-type controls under N deficiency conditions.
N is an essential nutrient for plant biomass production. To investigate how different amounts of N uptake resulted in distinct plant growth rates, we compared the total N contents in wild-type and transgenic plants under N-starved (2 mm), N-sufficient (10 mm), and N-excessive (40 mm) conditions. The result indicates that the higher the concentration of the N solution applied, the more total N that plants accumulated (Fig. 7G). When the total N content was measured as the percentage of the biomass weight, there was no significant difference between wild-type and transgenic plants. However, transgenic shoots accumulated significantly more N than wild-type controls under N-starved and N-excessive conditions due to their higher shoot biomass under both conditions (Fig. 7, G and H; Supplemental Fig. S5), suggesting an enhanced NUE in transgenic plants.
To investigate what causes the enhanced NUE, we examined the transcript levels of the genes encoding key enzymes in the N assimilation pathway in wild-type and transgenic creeping bentgrass plants. The enzymes include NITRATE REDUCTASE (NR), NiR, GLUTAMINE SYNTHETASE (GS), and GLUTAMATE SYNTHASE (GOGAT). As shown in Figure 8A, the expression of AsNiR (KR911829), but not AsNR (KR911828), AsGS (KR911826), or AsGOGAT (KR911827), was significantly up-regulated in transgenic plants in comparison with wild-type controls. Consistently, the enzyme activity of NiR was also significantly higher in transgenic plants than in wild-type controls before and after N starvation treatment (Fig. 8B). It should be noted that the NiR activity increased in both wild-type and transgenic plants in response to N starvation (Fig. 8B).
Figure 8.

AsNiR gene expression analysis and NiR enzyme assay in wild-type and transgenic plants. A, RT-qPCR analysis of AsNiR transcript levels in wild-type plants (WT) and three transgenic (TG) lines under 10 mm N conditions. AsACT1 was used as an endogenous control. Data are presented as means of three technical replicates and three biological replicates. B, NiR assay in wild-type controls and two transgenic lines before and 2 weeks after N starvation. Data are presented as means of three biological replicates. The error bars represent se. Asterisks indicate significant differences of expression levels or enzyme activities between wild-type and transgenic plants: *, P < 0.05; **, P < 0.01; and ***, P < 0.001 by Student’s t test.
Overexpression of miR528 Leads to Decreased AAO Activity under N Deficiency
N deprivation triggers redox changes and oxidative stress (Kandlbinder et al., 2004). The predicted miR528 target AAO gene and its role in oxidative stress regulation have been discussed above. Therefore, we conducted a plant AAO assay under normal and N starvation conditions. The result indicates that AAO activity in both wild-type and transgenic plants increased dramatically when subjected to N starvation, but the increase in transgenic plants was significantly less pronounced than that in wild-type controls (Supplemental Fig. S6). The lower level of AAO in transgenics helps maintain relatively high levels of redox AA under N deprivation, thereby keeping the balance between ROS production and its scavenging under oxidative stress.
Osa-miR528 Putative Target Identification and Responses to Stresses
To understand the underlying molecular mechanisms of miR528-mediated plant responses to salinity and N deficiency, we sought to identify putative targets of miR528 in creeping bentgrass. Currently, only SsCBP1, a copper ion-binding domain-containing protein, is experimentally confirmed as the target of miR528 in sugarcane (Zanca et al., 2010). In rice, Os06g37150 encoding AAO is validated as the target of miR528 through a high-throughput degradome sequencing approach (Wu et al., 2009b). To identify its targets in creeping bentgrass, a plant small RNA target analysis tool (psRNA Target) was applied to predict targets in the rice genome (Dai and Zhao, 2011). Eleven putative targets were recognized in rice, including one copper ion-binding protein (Os07g38290), one AAO (Os06g37150), one laccase precursor protein (Os01g44330), one Zn/Cu SOD (Os08g44770), two plastocyanin-like domain-containing proteins (Os06g11310 and Os08g04310), one translation initiation factor (Os01g40150), one F-box domain-containing protein (Os06g06050), two multicopper oxidase domain-containing proteins (Os01g03620 and Os01g03640), and one unknown protein (Os09g33800), of which partial fragments of four genes were successfully amplified in creeping bentgrass based on the sequence similarity to rice (data not shown). Genes encoding AAO and CBP1 showed decreased expression in transgenic plants (Fig. 9, A and B), indicating that they might be targets of miR528 in creeping bentgrass. The miR528 targeting site in AsCBP1 (KR911824) was detected in its open reading frame as described in Figure 9C. Interestingly, the miR528 target site was not found in the coding region of AsAAO; instead, it was identified, by RACE analysis, to be located in the 3′ untranslated region from nucleotides 26 to 45 after the stop codon TGA (Fig. 9C). The descriptions, functions, and corresponding orthologs in rice and Arabidopsis (Arabidopsis thaliana) of AsAAO and AsCBP1 are listed in Figure 9D. AsAAO (KR911823) functions in oxidation reduction, suggesting its potentially important role in the plant abiotic stress response. AsCBP1 encodes a cupredoxin superfamily protein. Proteins from this family function in oxidation homeostasis and electron transfer reactions, which are involved in photosynthesis, respiration, cell signaling, and numerous reactions of oxidases and reductases (Lu et al., 2004; Solomon et al., 2004; Dennison, 2005; Marshall et al., 2009).
Figure 9.

Putative miR528 target identification in creeping bentgrass. A and B, Expression levels of AsAAO (A) and AsCBP1 (B) in wild-type plants (WT) and three transgenic (TG) lines examined via RT-qPCR. AsUBQ5 was used as an endogenous control. Data are presented as means of three technical replicates, and error bars represent se. Asterisks indicate a significant difference of expression levels between the wild type and each transgenic line: *, P < 0.05; **, P < 0.01; and ***, P < 0.001 by Student’s t test. C, Comparison of miR528 target sites in the putative targets AAO and CBP1 between rice and creeping bentgrass. Asterisks indicate identical RNA sequences. D, Information about the orthologs of the two putative miR528 target genes in rice and Arabidopsis.
To investigate whether AsAAO and AsCBP1 respond to salt and N deficiency stress, we conducted real-time RT-PCR analysis to examine their expression profiles under salt and N starvation treatments. Figure 10A shows that the expression of AsAAO was significantly induced in response to salt stress in both leaf (approximately 30-fold at 6 h after treatment) and root (15-fold at 6 h after treatment) tissues. When plants were exposed to N deficiency, the expression of AsAAO was significantly induced (more than 2-fold) in leaves but reduced (approximately 0.5-fold) in roots 5 d after N starvation (Fig. 10B). When plants were exposed to salt stress, AsCBP1 expression in leaf and root tissues was significantly induced (approximately 2.5- and 3-fold, respectively, at 6 h after treatment; Fig. 10A). During N starvation, AsCBP1 was induced approximately 4.5 times at 5 d after treatment and then declined in leaf tissues, while its expression declined 8 d after treatment in root tissues (Fig. 10B).
Figure 10.

Expression patterns of the two miR528 putative targets under salt and N deficiency conditions through real-time RT-PCR analysis. A, Expression profiles of AsAAO and AsCBP1 in wild-type leaf and root tissues under 200 mm NaCl treatment (0–6 h). B, Expression profiles of AsAAO and AsCBP1 in wild-type leaf and root tissues under N starvation (0 mm N) from 0 to 8 d. AsUBQ5 was used as an endogenous control. Data are presented as means of three technical replicates, and error bars represent se. Asterisks indicate significant differences of gene expression levels between untreated and stress-treated leaf or root tissues: *, P < 0.05; **, P < 0.01; and ***, P < 0.001 by Student’s t test.
miR528 Has Cross Talk with Other Abiotic Stress-Related miRNAs and Genes in Perennial Grasses
Although the importance of miRNAs in the complex stress response network is being gradually recognized, the molecular mechanisms of the miRNA-mediated plant stress response are still largely unknown. It would be interesting to know if different miRNAs have interactions in plant responses to abiotic stress. To this end, stem-loop RT-qPCR was conducted to analyze several conserved miRNAs that are implicated in plant abiotic stress responses, including miR396, miR156, and miR172 (Wu et al., 2009a; Gao et al., 2010; Hackenberg et al., 2012; Liang et al., 2012; Bhardwaj et al., 2014; Cui et al., 2014; Gupta et al., 2014; Pandey et al., 2014; Xie et al., 2015). Figure 11, A to C, shows that miR396, miR156, and miR172 were all down-regulated in transgenic plants overexpressing Osa-miR528, suggesting a potential cross talk of miR528 with other miRNAs in the regulatory network to orchestrate the plant response to stress. It should be noted that, since these miRNAs are also predominantly involved in development, for example, miR156 and miR172 control phase change and flowering time (Huijser and Schmid, 2011) and miR396 functions in cell development and proliferation in leaves (Rodriguez et al., 2010), it is possible that changes in these miRNAs in the miR528 transgenic plants may reflect their cross talk with miR528 to contribute to the changes in plant development, leading to the altered plant morphology observed in miR528 transgenic plants.
Figure 11.

Expression levels of the abiotic stress-related miRNAs and AsNAC60 in wild-type controls and Osa-miR528 transgenic lines. A to C, Expression levels of miR396 (A), miRl56 (B), and miR172 (C) in wild-type (WT) and transgenic (TG) plants revealed through stem-loop RT-qPCR analysis. D, Expression levels of AsNAC60 in the wild type and three transgenic lines through RT-qPCR analysis. Three technical replicates were used for the RT-qPCR analysis. AsUBQ5 was used as an endogenous control. The relative changes of gene expression were calculated based on the 2−ΔΔCT method (Livak and Schmittgen, 2001). The error bars represent se (n = 3). Asterisks indicate a significant difference of expression levels between the wild-type control and each transgenic line: *, P < 0.05; **, P < 0.01; and ***, P < 0.001 by Student’s t test.
Besides its cross talk with other miRNAs, miR528 was also examined for its potential impact on the expression of other stress-related genes. NAC (for NAM [No Apical Meristems], ATAF [Arabidopsis Transcription Activation Factor], and CUC [Cup-Shape Cotyledon]) proteins play an important role in plant environmental stress tolerance (Koyama et al., 2010). ONAC60 (Os12g41680), one of the NAC family members in rice, is significantly up-regulated by high salinity (Nuruzzaman et al., 2010). We examined the expression of AsNAC60, an ortholog of ONAC60 in creeping bentgrass (Zhou et al., 2013). Figure 11D shows that AsNAC60 was up-regulated in three transgenic lines, suggesting that miR528 may indirectly regulate AsNAC60 expression, which contributes to the concerted plant response to stress.
DISCUSSION
miR528-Mediated Plant Development
The miRNAs play diverse roles in plant development. However, there has been no report so far demonstrating the involvement of miR528 in plant development at the vegetative stage. In this study, we show that transgenic creeping bentgrass overexpressing Osa-miR528 exhibits thicker leaves, more vascular bundles, more tillers, shorter internodes and tillers, and more upright growth than wild-type controls. Less vertical plant growth due to shortened internodes, resulting in a dwarf phenotype, is one of the desirable traits in turf grass. Increased tiller number that results in denser, more attractive, and uniform communities is another desirable turf trait.
Lignin contributes to the structural rigidity of the cell wall, which is required to keep plants continuously erect. miR528-expressing transgenic creeping bentgrass has more vascular bundles than wild-type controls, suggesting that miR528 transgenics might contain more lignin than wild-type controls. This is supported by the more upright growth of transgenic plants than wild-type controls (Supplemental Fig. S1C). Further study of the difference in lignin contents and cell wall structure between wild-type and transgenic plants should provide evidence for a better understanding of miR528-mediated alteration in plant development.
Lodging largely reduces crop yield and grain quality. Strong and short stems are essential in plant lodging resistance. miR528 transgenic plants with increased vascular bundles and probably higher lignin contents contribute to the stronger stems than wild-type controls. It is likely that the stronger stems of the transgenic plants would lead to enhanced lodging resistance. Additionally, the shorter internodes and shorter tiller length of the transgenics would also account for increased lodging resistance. To our knowledge, there have been no studies showing miRNA-mediated plant lodging responses. Our data demonstrate the possible involvement of miR528 in the plant response to lodging stress via regulating plant architecture.
ROS Scavenging and Abiotic Stress Resistance
Antioxidants such as AA and glutathione, and ROS-scavenging enzymes such as SOD, CAT, and ASCORBATE PEROXIDASE, play important roles in protecting plants against abiotic stresses (Rizhsky et al., 2004; Wang et al., 2005; Leshem et al., 2006; Koussevitzky et al., 2008). Among the predicted targets of miR528 in rice, AAO, laccase precursor protein, and Zn/Cu SOD are involved in ROS detoxification, and CBP1 encodes a protein belonging to the cupredoxin superfamily, which functions in oxidation homeostasis and electron transfer reactions, suggesting the critical role of miR528 in plant abiotic stress tolerance. In zinc-deficient sorghum, miR528 is down-regulated in young leaves, while the gene expression and the enzyme activity of Zn/Cu SOD are increased (Li et al., 2013). In transgenic creeping bentgrass overexpressing miR528, the enzyme activity of SOD does not show significant differences between wild-type and transgenic plants before and after salinity stress. Further studies by cloning and analyzing genes encoding Zn/Cu SOD will provide information for a better understanding of the interaction between SOD and miR528 in creeping bentgrass.
CAT, mainly localized in peroxisomes, serves as an efficient ROS scavenger to remove excessive H2O2 to avoid oxidative damage (Mhamdi et al., 2010). In this study, transgenic plants show increased CAT activity in both normal and salinity conditions, indicating that they have an enhanced capacity of balancing between ROS accumulation and scavenging compared with wild-type controls, thus leading to enhanced salinity resistance. Many CATs are regulated by developmental and environmental oxidative stresses (Du et al., 2008; Chen et al., 2012). In Capsicum annuum, a CAT gene is induced under NaCl treatment (Kwon and An, 2001). Among the predicted targets of miR528, however, there is no gene encoding CAT. Presumably, other factors triggered by miR528 overexpression cause the increased CAT activity and contribute to the increased salt tolerance.
Environmental stimuli trigger the oxidative burst mainly caused by changes in the redox state. AA is the major redox buffer in plants, while AAO catalyzes the oxidation of AA to dehydroascorbic acid via dehydroascorbate. Previous studies indicate that the redox state of the AA pool in plants is regulated by AAO (Pignocchi and Foyer, 2003; Pignocchi et al., 2003). Transgenic tobacco (Nicotiana tabacum) overexpressing AAO leads to increased ozone sensitivity due to the oxidized AA pool in the apoplast, the first line of defense against ROS (Pignocchi and Foyer, 2003; Sanmartin et al., 2003). In this study, AAO is predicted to be the direct target of miR528. In addition, transgenic plants exhibit significantly lower AAO activity than wild-type controls under salinity and N deficiency conditions (Fig. 9A), which would presumably result in an enhanced capacity of maintaining the AA redox state in transgenics and thereby enhancing plant oxidative stress resistance.
Leaf senescence is an age-dependent and a highly regulated degenerative process. The accumulation of ROS with age is a signal to trigger leaf senescence. In this study, transgenic plants exhibit less wilting of leaves than wild-type controls under normal growth conditions (Supplemental Fig. S4). Given that miR528 plants show enhanced CAT activity and, consequently, increased capacity of ROS scavenging, it is plausible that the overexpression of miR528 delays the process of leaf senescence in transgenic plants. Although leaf senescence is a genetically programmed and tightly controlled process, it is also influenced by various environmental cues (Becker and Apel, 1993; Buchanan-Wollaston et al., 2005). Under salinity and N deficiency, miR528 plants display less wilting of leaves than wild-type controls, as observed in normal growth conditions, indicating a decreased leaf senescence rate in transgenic plants. It is likely that the increased CAT activity and the decreased AAO activity in transgenics lead to the enhanced ROS scavenging and, consequently, delayed leaf senescence.
K+:Na+ Ratio and High-Salinity Resistance
Elevated cellular Na+ contents result in the inhibition of K+ absorption due to the physiochemical similarity between Na+ and K+ and, consequently, a lack of discrimination between them during cation-transporting and enzyme reactions (Maathuis and Amtmann, 1999). As K+ participates in many important processes during metabolism, growth, and the stress response, it is critical to maintain a high cytosolic K+:Na+ ratio. In this study, the K+:Na+ ratio is significantly increased in three transgenic lines in comparison with wild-type controls under salinity stress, due to the increased capacity of maintaining stable K+ acquisition and distribution (Fig. 5, F and H), which results in the enhanced salt tolerance in miR528 plants. Further gene expression analysis reveals that the stable K+ acquisition in transgenics is associated with the increased expression of HAK5, a high-affinity K+ transporter (Fig. 6). Similar results were also reported in rice. The OsHAK5 knockout mutant exhibits severely impaired K+ influx and transport, decreased K+:Na+ ratio, and sensitivity to salt stress compared with wild-type rice, whereas OsHAK5 overexpression leads to increased K+ uptake and K+:Na+ ratio and enhanced salt stress tolerance in transgenic rice plants (Yang et al., 2014).
Interestingly, miR528 transgenic leaves show significantly higher Na+ contents than wild-type controls under normal growth conditions (Fig. 5A), whereas Na+ contents are similar in wild-type and transgenic plants during salt treatment (Fig. 5B). Similar phenomena have also been observed previously in other plant species. A study based on more than 300 Arabidopsis accessions shows that accessions growing in high-saline soil have higher Na+ accumulation in leaves than others (Baxter et al., 2010). The elevated Na+ in leaves results from a weak allele of Arabidopsis High-Affinity Potassium Transporter1;1 (AtHKT1;1), which promotes the accumulation of Na+ in leaves without Na+ toxicity (Baxter et al., 2010). Similarly, a transgenic barley (Hordeum vulgare) overexpressing HvHKT2;1 exhibits elevated shoot Na+ accumulation and enhanced salt stress tolerance (Mian et al., 2011). We speculate that the elevated Na+ in miR528 transgenic leaves is the result of a salt inclusion mechanism for osmotic adjustment during salt stress. In this study, the similar accumulation of Na+ in wild-type and transgenic plants might account for the saturated Na+ environment under severe salt stress treatment. Further testing under moderate salt conditions and analysis of the main membrane transporters in transgenic plants overexpressing miR528 would allow a better understanding of miR528-mediated Na+ uptake and distribution in plants.
N Assimilation and N Deficiency Tolerance
N is one of the most critical elements for crop productivity. Currently, 67% of N applied to soil is lost into the environment (Abrol and Raghuram, 2007). Although the form and the amount of N available to plants can be improved via optimizing N management, the efficiency of N utilization has to be tackled biologically. NUE mainly includes N uptake and N assimilation. Many attempts have been made so far toward manipulating key genes in the process of these two steps (Ferrario-Mery et al., 1998; Limami et al., 1999; Fei et al., 2003). However, success has been limited. For example, constitutive expression of NiR in Arabidopsis and tobacco leads to increased transcription of NiR but decreased enzyme activity of NiR, due to the posttranslational modification (Crété et al., 1997; Takahashi et al., 2001). In this study, miR528 transgenic plants exhibit increased transcript levels of AsNiR (Fig. 8A) and enzyme activity (Fig. 8B), which might contribute to the increased N assimilation efficiency and better NUE. The molecular mechanisms of miR528-mediated AsNiR regulation remain elusive. It is hypothesized that the expression of AsNiR might be controlled by either the targets of miR528, or some other factors triggered by miR528, or directly by miR528 itself at the transcriptional, posttranscriptional, or posttranslational level. Interestingly, NiR activity in both wild-type and transgenic plants increases under N starvation compared with normal conditions, suggesting the involvement of other factors in the regulation of NiR activity.
Given the improved shoot biomass and total N content in transgenic plants in comparison with wild-type controls during N deficiency (Fig. 7H), it is likely that transgenic plants have increased N uptake or assimilation or total NUE. The expression levels of genes encoding the key enzymes NR, NiR, GS, and GOGAT in the N assimilation pathway have been compared between wild-type and transgenic plants. Further study characterizing important nitrate transporters in wild-type and miR528 transgenic plants will allow a better understanding of the enhanced tolerance to N deficiency.
Excessive N limits the plant uptake of other nutritional elements, such as K and phosphorus, and subsequently results in reduced biomass accumulation compared with N-sufficient conditions, as observed in this study (Fig. 7, B, E, and F). Interestingly, miR528 transgenics display significantly higher shoot biomass and total N contents than wild-type controls under excessive N supplies (Fig. 7, B, D–F, and H). Therefore, it is hypothesized that miR528 might be an important regulator functioning in buffering the imbalanced mineral elements. As discussed above, miR528 promotes AsHAK5, which functions in maintaining K acquisition. It is also likely that other factors impacted by miR528 might be more directly involved in maintaining the homeostasis of different nutritional elements during N-excessive conditions and subsequently leading to the improved biomass in transgenic plants.
Molecular Mechanisms of miR528-Mediated Plant Salt and N Deficiency Resistance
The accumulation of miR528 is elevated during salt stress (Fig. 1A), which might cause reduced transcript levels of its predicted targets AsAAO and AsCBP1 (Fig. 9). Both targets are suggested to mediate oxidation homeostasis and, thus, prevent damage to cellular components. Besides the direct targets of miR528, genes involved in other signaling pathways also contribute to the enhanced salt stress tolerance. A high-affinity K transporter, AsHAK5, induced in transgenic creeping bentgrass overexpressing miR528 (Fig. 6), is critical for maintaining K+ homeostasis during normal and salinity conditions. Moreover, miR528 induces the activity of CAT and, therefore, maintains ROS homeostasis under abiotic stress. In addition to functional proteins, miR528 also positively regulates AsNAC60 (Fig. 11D), which is a creeping bentgrass ortholog of a salt stress-induced transcription factor, suggesting the importance of AsNAC60 in miR528-mediated salt stress tolerance in creeping bentgrass. miR528 is gradually repressed during N deficiency (Fig. 1C), therefore releasing the inhibition of its targets, which contribute to oxidation homeostasis. AsNiR, a key enzyme in the process of N assimilation, is positively regulated by miR528 (Fig. 8B). The enhanced NUE is presumably attributed to the increased AsNiR activity. miRNAs are suggested to serve as master regulators in the complex regulatory network of plant responses to abiotic stress (Zhou and Luo, 2013). The impact of miR528 on the expression of other stress-related miRNAs observed in this study (Fig. 11, A–C) suggests the coordinated interactions of multiple stress regulators, thereby leading to enhanced salt and N deficiency tolerance. The hypothetical model of the miR528-mediated plant abiotic stress response pathway (Fig. 12) provides information allowing the development of novel molecular strategies to genetically engineer crop species for enhanced environmental stress tolerance.
Figure 12.

Hypothetical model of the molecular mechanisms of miR528-mediated plant abiotic stress response in creeping bentgrass. miR528 is induced during salinity stress but down-regulated under N deficiency. miR528 mediates plant abiotic stress responses through directly repressing the expression of its targets AsAAO and AsCBP1, which regulate the oxidation homeostasis during abiotic stresses. In addition, miR528 positively regulates AsNAC60, AsHAK5, and AsNiR and the gene encoding the antioxidant enzyme CAT, which leads to the enhanced tolerance to salinity stress and N deficiency. Furthermore, expression levels of other stress-related miRNAs are negatively regulated by miR528, suggesting that different miRNAs form a regulatory network to coordinately integrate various signals in response to plant abiotic stress.
MATERIALS AND METHODS
Cloning of the miR528 Gene and Plasmid Construction
The 550-bp rice (Oryza sativa) cDNA (AK073820) containing the Osa-miR528 stem-loop structure was isolated by PCR using the forward and reverse primer set 5′-TCTAGAGATCAGCAGCAGCCACA-3′ and 5′-GTCGACGACCAAATAATGTGTTACTG-3′, which contained XbaI and SalI restriction sites (underlined), respectively. PCR products were cloned into the binary vector pZH01 (Xiao et al., 2003), generating the Osa-miR528 overexpression gene construct p35S-Osa-miR528/p35S-Hyg. The construct contains the CaMV 35S promoter driving Osa-miR528 linked to the CaMV 35S promoter driving the Hyg gene for hygromycin resistance as a selectable marker. For subsequent plant transformation, the construct was transferred into Agrobacterium tumefaciens strain LBA4404.
Plant Materials and Transformation
Creeping bentgrass (Agrostis stolonifera) ‘Penn A-4’ (supplied by HybriGene) was used for plant transformation. Transgenic plants constitutively expressing Osa-miR528 were produced via A. tumefaciens-mediated transformation of embryonic callus induced from mature seeds as described previously (Luo et al., 2004).
Plant Propagation, Maintenance, and Abiotic Stress Treatments
The regenerated transgenic plants overexpressing Osa-miR528 and wild-type controls were clonally propagated from tillers and maintained as described previously (Zhou et al., 2013).
For salt stress treatments, plants grown in cone-tainers were immersed in a 200 mm NaCl solution supplemented with 0.2 g L−1 water-soluble fertilizer. After 9-d salt treatments, shoots and roots were harvested for further physiological analysis. Replicates of the salt-treated plants were recovered from salt stress by watering with 0.2 g L−1 water-soluble fertilizer every other day and were photographed for documentation. For miR528 expression analysis, wild-type leaves were collected at 0, 1.5, 3, and 6 h after salt stress treatment.
To test the performance of wild-type and transgenic plants under different concentrations of N, plants grown in cone-tainers were immersed in modified MS nutrient solution of pH 5.7 containing 3 mm CaCl2∙2H2O, 1.5 mm MgSO4∙7H2O, 1.25 mm KH2PO4, 0.1 mm H3BO3, 0.1 mm MnSO4∙4H2O, 0.1 mm ZnSO4∙H2O, 0.5 µm potassium iodide, 0.56 µm NaMO4∙2H2O, 0.1 µm CuSO4∙5H2O, 0.1 µm CoCl2∙6H2O, 0.1 mm FeSO4∙7H2O, and 0.1 mm Na2EDTA∙2H2O, supplemented with N at different concentrations (0.4, 2, 10, or 40 mm). N supplies include KNO3 and NH4NO3. Equivalent amounts of KCl were added to the nutrient solutions to maintain the osmotic potential and prevent K deficiency caused by low supplies of KNO3. For miR528 expression analysis, wild-type plants were immersed in modified MS nutrient solution without N. Wild-type leaves were collected at 0, 2, 5, 8, and 12 d after N starvation treatment.
The expression profile of miR528 during drought stress was analyzed in wild-type creeping bentgrass leaves. Wild-type plants were grown hydroponically with 0.2 g L−1 water-soluble fertilizer. Right before the drought treatment, the plants were taken out of the nutritional solution and dried with a paper towel. They were then laid on the bench for air drying. The leaf samples were collected at 0, 1, 2, and 4 h after drought treatment.
Plant DNA and RNA Isolation and Expression Analysis
Plant genomic DNA was extracted from 30 mg of fresh leaves following the protocol of Luo et al. (1995). Plant total RNA was isolated from 100 mg of fresh leaves using Trizol reagent (Invitrogen) following the manufacturer’s protocol. First-strand cDNA was synthesized from 2 µg of RNA with SuperScript II Reverse Transcriptase (Invitrogen) and oligo(dT) or gene-specific primers. Semiquantitative RT-PCR was conducted on 24 to 30 cycles based on its exponential phase. PCR products were separated by electrophoresis using a 0.8% (w/v) or 1.5% (w/v) agarose gel, visualized, and photographed with the BioDoc-It imaging system (Ultra-Violet Products).
RT-qPCR was performed with 12.5 µL of iQ SYBR-Green Supermix (Bio-Rad Laboratories) per 25-µL reaction. The green fluorescence signal was monitored on the Bio-Rad iQ5 real-time detection system using the iQ5 Optical System Software version 2.0 (Bio-Rad Laboratories). AsACT1 (JX644005) and AsUBQ5 (JX570760) were used as endogenous controls. The relative changes in gene expression were calculated based on the comparative threshold cycle method (Livak and Schmittgen, 2001).
Stem-loop RT-qPCR was performed according to the protocol of Varkonyi-Gasic et al. (2007). The Osa-miR528 stem-loop RT primer and PCR forward primer are 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCTCCTC-3′ and 5′-GCAGTGGAAGGGGCATGCA-3′, respectively.
Measurement of Mineral Content
For Na+ and K+ content measurements, leaves and roots of wild-type and transgenic plants were collected before and after a 9-d 200 mm NaCl solution treatment. For total N measurement, wild-type and transgenic plant leaves were collected after 4 weeks of treatment with different concentrations of N: 0.4, 2, 10, and 40 mm. A total of 0.2 g of each dried sample for total N measurement and 0.5 g of each dried sample for Na+ and K+ content measurements were analyzed to determine the mineral contents according to previous protocols (Haynes, 1980; Li et al., 2010).
Measurement of Leaf RWC, EL, Chlorophyll, and Pro Content
Plant leaf RWC, EL, chlorophyll a and b, as well as Pro content were measured following previous protocols (Bates et al., 1973; Li et al., 2010).
Antioxidant Enzyme Assay
A total of 0.1 g of leaves was weighed and homogenized immediately in 1 mL of 0.05 m phosphate-buffered saline at pH 7.8 on ice and centrifuged at 14,000g for 30 min at 4°C. Supernatant of each sample was transferred to a new 1.5-mL Eppendorf tube and stored on ice for further enzyme assay. The activity of SOD (EC 1.15.1.1) was determined as described previously (Giannopolitis and Ries, 1977). One unit of SOD activity was defined as the amount of enzyme required to inhibit a 50% (v/v) reduction of nitroblue tetrazolium chloride.
The activity of CAT (EC 1.11.7.6) was determined as described previously (Maehly and Chance, 1954). The decomposition of H2O2 was measured by the decrease of the absorbance at the wavelength of 240 nm for 1 min. An enzyme unit was defined as the amount of enzyme necessary to decompose 1 µm H2O2 at 25°C in 1 min.
NiR Assay
NiR (EC 1.7.2.1) activity was measured by a spectrophotometric assay as described previously (Losada and Paneque, 1971). The nitrite content was calculated from a KNO2 standard curve. NiR activity was determined by the reduction of nitrite that was catalyzed by the enzyme in 1 mg of soluble protein per hour. Protein contents were measured according to the dye-binding method of Bradford (1976).
AAO Assay
The activity of AAO (EC 1.10.3.3) was determined spectrophotometrically at 25°C following the decrease in absorbance of ascorbic acid at 265 nm as described previously (Esaka et al., 1988). The activity of AAO was determined as:
 |
where e = 13.386 mm-1 cm−1 (an extinction coefficient for ascorbic acid at 265 nm), total volume = 2 mL, and enzyme volume = 0.05 mL.
Plant Histology Analysis
The second and third internodes from the top tillers and the fully expanded leaves of wild-type and transgenic plants were collected and immersed in formalin-acetic-alcohol fixation for 48 h at room temperature. After fixation, plant tissues were dehydrated with a series of graded ethanol from 70% to 100% (v/v), followed by paraffin wax infiltration. Tissues were then embedded in paraffin blocks to process sections using a rotary microtome (RM 2165; Leica). Sections were stained using Toluidine Blue and observed with a stereomicroscope (MEIJI EM-5). Photographs were taken using a 35-mm SLR camera body (Canon) connected to a microscope. Scale bars were added to photographs using ImageJ (Abràmoff et al., 2004).
Sequence data from this article can be found in GenBank under accession numbers KR911823, KR911824, KR911825, KR911826, KR911827, KR911828, and KR911829.
Supplemental Data
The following supplemental materials are available.
Supplemental Figure S1. Development of wild-type and transgenic plants.
Supplemental Figure S2. Biomass measurement in wild-type and transgenic plants.
Supplemental Figure S3. Antioxidant enzyme activity assay under normal and salt conditions.
Supplemental Figure S4. Wild-type plants exhibit wilting leaf tips compared with three transgenic lines.
Supplemental Figure S5. Biomass accumulation of wild-type and Osa-miR528 transgenic plants in response to different concentrations of N supplies.
Supplemental Figure S6. AAO activity measurement under normal and N deficiency conditions.
Supplementary Material
Acknowledgments
We thank Dr. Julia Frugoli, Dr. Liangjiang Wang, Dr. William R. Marcotte, Dr. Halina Knap, and members of H.L.’s laboratory for useful discussions and Dr. Lihuang Zhu for providing the binary vector pZH01.
Glossary
- N
nitrogen
- NUE
nitrogen use efficiency
- miRNA
microRNA
- RT-qPCR
reverse transcription-quantitative PCR
- cDNA
complementary DNA
- CaMV
Cauliflower mosaic virus
- RT
reverse transcription
- EL
electrolyte leakage
- RWC
relative water content
- ROS
reactive oxygen species
- K
potassium
- Zn/Cu
zinc/copper
- AA
ascorbate
- MS
Murashige and Skoog
- H2O2
hydrogen peroxide
Footnotes
This work was supported by the U.S. Department of Agriculture (Biotechnology Risk Assessment Grant Program grant no. 2010–33522–21656 from the National Institute of Food and Agriculture and Cooperative State Research, Education, and Extension Service grant no. SC–1700450). This is Technical Contribution no. 6289 of the Clemson University Experiment Station.
Articles can be viewed without a subscription.
References
- Abràmoff MD, Magalhães PJ, Ram SJ (2004) Image processing with ImageJ. Biophoton Int 11: 36–43 [Google Scholar]
- Abrol Y, Raghuram N (2007) Agricultural Nitrogen Use and Its Environmental Implications. I.K. International Publishing House, New Dehli, India [Google Scholar]
- Ali Y, Aslam Z, Ashraf M, Tahir G (2004) Effect of salinity on chlorophyll concentration, leaf area, yield and yield components of rice genotypes grown under saline environment. Int J Environ Sci Technol 1: 221–225 [Google Scholar]
- An FM, Hsiao SR, Chan MT (2011) Sequencing-based approaches reveal low ambient temperature-responsive and tissue-specific microRNAs in Phalaenopsis orchid. PLoS ONE 6: e18937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashraf M, Foolad M (2007) Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ Exp Bot 59: 206–216 [Google Scholar]
- Bates L, Waldren R, Teare I (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39: 205–207 [Google Scholar]
- Baxter I, Brazelton JN, Yu D, Huang YS, Lahner B, Yakubova E, Li Y, Bergelson J, Borevitz JO, Nordborg M, et al. (2010) A coastal cline in sodium accumulation in Arabidopsis thaliana is driven by natural variation of the sodium transporter AtHKT1;1. PLoS Genet 6: e1001193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Becker W, Apel K (1993) Differences in gene expression between natural and artificially induced leaf senescence. Planta 189: 74–79 [Google Scholar]
- Bhardwaj AR, Joshi G, Pandey R, Kukreja B, Goel S, Jagannath A, Kumar A, Katiyar-Agarwal S, Agarwal M (2014) A genome-wide perspective of miRNAome in response to high temperature, salinity and drought stresses in Brassica juncea (Czern) L. PLoS ONE 9: e92456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradford MM. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248–254 [DOI] [PubMed] [Google Scholar]
- Buchanan-Wollaston V, Page T, Harrison E, Breeze E, Lim PO, Nam HG, Lin JF, Wu SH, Swidzinski J, Ishizaki K, et al. (2005) Comparative transcriptome analysis reveals significant differences in gene expression and signalling pathways between developmental and dark/starvation-induced senescence in Arabidopsis. Plant J 42: 567–585 [DOI] [PubMed] [Google Scholar]
- Chapin FS III, Walter CH, Clarkson DT (1988) Growth response of barley and tomato to nitrogen stress and its control by abscisic acid, water relations and photosynthesis. Planta 173: 352–366 [DOI] [PubMed] [Google Scholar]
- Chen HJ, Wu SD, Huang GJ, Shen CY, Afiyanti M, Li WJ, Lin YH (2012) Expression of a cloned sweet potato catalase SPCAT1 alleviates ethephon-mediated leaf senescence and H2O2 elevation. J Plant Physiol 169: 86–97 [DOI] [PubMed] [Google Scholar]
- Crété P, Caboche M, Meyer C (1997) Nitrite reductase expression is regulated at the post-transcriptional level by the nitrogen source in Nicotiana plumbaginifolia and Arabidopsis thaliana. Plant J 11: 625–634 [DOI] [PubMed] [Google Scholar]
- Cui LG, Shan JX, Shi M, Gao JP, Lin HX (2014) The miR156-SPL9-DFR pathway coordinates the relationship between development and abiotic stress tolerance in plants. Plant J 80: 1108–1117 [DOI] [PubMed] [Google Scholar]
- Dai X, Zhao PX (2011) psRNATarget: a plant small RNA target analysis server. Nucleic Acids Res 39: W155–W159 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dennison C. (2005) Investigating the structure and function of cupredoxins. Coord Chem Rev 249: 3025–3054 [Google Scholar]
- Ding D, Zhang L, Wang H, Liu Z, Zhang Z, Zheng Y (2009) Differential expression of miRNAs in response to salt stress in maize roots. Ann Bot (Lond) 103: 29–38 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du YY, Wang PC, Chen J, Song CP (2008) Comprehensive functional analysis of the catalase gene family in Arabidopsis thaliana. J Integr Plant Biol 50: 1318–1326 [DOI] [PubMed] [Google Scholar]
- Esaka M, Imagi J, Suzuki K, Kubota K (1988) Formation of ascorbate oxidase in cultured pumpkin cells. Plant Cell Physiol 29: 231–235 [Google Scholar]
- Fei H, Chaillou S, Hirel B, Mahon JD, Vessey JK (2003) Overexpression of a soybean cytosolic glutamine synthetase gene linked to organ-specific promoters in pea plants grown in different concentrations of nitrate. Planta 216: 467–474 [DOI] [PubMed] [Google Scholar]
- Ferrario-Mery S, Valadier MH, Foyer CH (1998) Overexpression of nitrate reductase in tobacco delays drought-induced decreases in nitrate reductase activity and mRNA. Plant Physiol 117: 293–302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferreira TH, Gentile A, Vilela RD, Costa GGL, Dias LI, Endres L, Menossi M (2012) MicroRNAs associated with drought response in the bioenergy crop sugarcane (Saccharum spp.). PLoS ONE 7: e46703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fraisier V, Gojon A, Tillard P, Daniel-Vedele F (2000) Constitutive expression of a putative high-affinity nitrate transporter in Nicotiana plumbaginifolia: evidence for post-transcriptional regulation by a reduced nitrogen source. Plant J 23: 489–496 [DOI] [PubMed] [Google Scholar]
- Gambale F, Uozumi N (2006) Properties of shaker-type potassium channels in higher plants. J Membr Biol 210: 1–19 [DOI] [PubMed] [Google Scholar]
- Gao P, Bai X, Yang L, Lv D, Li Y, Cai H, Ji W, Guo D, Zhu Y (2010) Over-expression of osa-MIR396c decreases salt and alkali stress tolerance. Planta 231: 991–1001 [DOI] [PubMed] [Google Scholar]
- Giannopolitis CN, Ries SK (1977) Superoxide dismutases. I. Occurrence in higher plants. Plant Physiol 59: 309–314 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golldack D, Lüking I, Yang O (2011) Plant tolerance to drought and salinity: stress regulating transcription factors and their functional significance in the cellular transcriptional network. Plant Cell Rep 30: 1383–1391 [DOI] [PubMed] [Google Scholar]
- Gupta OP, Meena NL, Sharma I, Sharma P (2014) Differential regulation of microRNAs in response to osmotic, salt and cold stresses in wheat. Mol Biol Rep 41: 4623–4629 [DOI] [PubMed] [Google Scholar]
- Hackenberg M, Shi BJ, Gustafson P, Langridge P (2012) A transgenic transcription factor (TaDREB3) in barley affects the expression of microRNAs and other small non-coding RNAs. PLoS ONE 7: e42030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haynes R. (1980) A comparison of two modified Kjeldahl digestion techniques for multi‐element plant analysis with conventional wet and dry ashing methods. Commun Soil Sci Plant Anal 11: 459–467 [Google Scholar]
- Huijser P, Schmid M (2011) The control of developmental phase transitions in plants. Development 138: 4117–4129 [DOI] [PubMed] [Google Scholar]
- Kandlbinder A, Finkemeier I, Wormuth D, Hanitzsch M, Dietz KJ (2004) The antioxidant status of photosynthesizing leaves under nutrient deficiency: redox regulation, gene expression and antioxidant activity in Arabidopsis thaliana. Physiol Plant 120: 63–73 [DOI] [PubMed] [Google Scholar]
- Kikuchi S, Satoh K, Nagata T, Kawagashira N, Doi K, Kishimoto N, Yazaki J, Ishikawa M, Yamada H, Ooka H, et al. (2003) Collection, mapping, and annotation of over 28,000 cDNA clones from japonica rice. Science 301: 376–379 [DOI] [PubMed] [Google Scholar]
- Kim MH, Sato S, Sasaki K, Saburi W, Matsui H, Imai R (2013) COLD SHOCK DOMAIN PROTEIN 3 is involved in salt and drought stress tolerance in Arabidopsis. FEBS Open Bio 3: 438–442 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koussevitzky S, Suzuki N, Huntington S, Armijo L, Sha W, Cortes D, Shulaev V, Mittler R (2008) Ascorbate peroxidase 1 plays a key role in the response of Arabidopsis thaliana to stress combination. J Biol Chem 283: 34197–34203 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koyama T, Mitsuda N, Seki M, Shinozaki K, Ohme-Takagi M (2010) TCP transcription factors regulate the activities of ASYMMETRIC LEAVES1 and miR164, as well as the auxin response, during differentiation of leaves in Arabidopsis. Plant Cell 22: 3574–3588 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kozomara A, Griffiths-Jones S (2014) miRBase: annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res 42: D68–D73 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwon SI, An CS (2001) Molecular cloning, characterization and expression analysis of a catalase cDNA from hot pepper (Capsicum annuum L.). Plant Sci 160: 961–969 [DOI] [PubMed] [Google Scholar]
- Lebaudy A, Véry AA, Sentenac H (2007) K+ channel activity in plants: genes, regulations and functions. FEBS Lett 581: 2357–2366 [DOI] [PubMed] [Google Scholar]
- Leshem Y, Melamed-Book N, Cagnac O, Ronen G, Nishri Y, Solomon M, Cohen G, Levine A (2006) Suppression of Arabidopsis vesicle-SNARE expression inhibited fusion of H2O2-containing vesicles with tonoplast and increased salt tolerance. Proc Natl Acad Sci USA 103: 18008–18013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li C, Chang PP, Ghebremariam KM, Qin L, Liang Y (2014) Overexpression of tomato SpMPK3 gene in Arabidopsis enhances the osmotic tolerance. Biochem Biophys Res Commun 443: 357–362 [DOI] [PubMed] [Google Scholar]
- Li Y, Zhang Y, Shi D, Liu X, Qin J, Ge Q, Xu L, Pan X, Li W, Zhu Y, et al. (2013) Spatial-temporal analysis of zinc homeostasis reveals the response mechanisms to acute zinc deficiency in Sorghum bicolor. New Phytol 200: 1102–1115 [DOI] [PubMed] [Google Scholar]
- Li Z, Baldwin CM, Hu Q, Liu H, Luo H (2010) Heterologous expression of Arabidopsis H+-pyrophosphatase enhances salt tolerance in transgenic creeping bentgrass (Agrostis stolonifera L.). Plant Cell Environ 33: 272–289 [DOI] [PubMed] [Google Scholar]
- Liang G, He H, Yu D (2012) Identification of nitrogen starvation-responsive microRNAs in Arabidopsis thaliana. PloS One 7: e48951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Limami A, Phillipson B, Ameziane R, Pernollet N, Jiang Q, Roy R, Deleens E, Chaumont-Bonnet M, Gresshoff PM, Hirel B (1999) Does root glutamine synthetase control plant biomass production in Lotus japonicus L.? Planta 209: 495–502 [DOI] [PubMed] [Google Scholar]
- Liu B, Li P, Li X, Liu C, Cao S, Chu C, Cao X (2005) Loss of function of OsDCL1 affects microRNA accumulation and causes developmental defects in rice. Plant Physiol 139: 296–305 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25: 402–408 [DOI] [PubMed] [Google Scholar]
- Losada M, Paneque A (1971) Nitrite reductase. Methods Enzymol 23: 487–491 [Google Scholar]
- Lu Y. (2003) Cupredoxins. In J McCleverty, TJ Meyer, eds, Comprehensive Coordination Chemistry II: From Biology to Nanotechnology, Vol 8. Elsevier, Oxford, pp 91–122 [Google Scholar]
- Lu Y, Li Y, Zhang J, Xiao Y, Yue Y, Duan L, Zhang M, Li Z (2013) Overexpression of Arabidopsis molybdenum cofactor sulfurase gene confers drought tolerance in maize (Zea mays L.). PLoS One 8: e52126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo H, Hu Q, Nelson K, Longo C, Kausch AP, Chandlee JM, Wipff JK, Fricker CR (2004) Agrobacterium tumefaciens-mediated creeping bentgrass (Agrostis stolonifera L.) transformation using phosphinothricin selection results in a high frequency of single-copy transgene integration. Plant Cell Rep 22: 645–652 [DOI] [PubMed] [Google Scholar]
- Luo H, Van Coppenolle B, Seguin M, Boutry M (1995) Mitochondrial DNA polymorphism and phylogenetic relationships in Hevea brasiliensis. Mol Breed 1: 51–63 [Google Scholar]
- Maathuis FJ, Amtmann A (1999) K nutrition and Na toxicity: the basis of cellular K/Na ratios. Ann Bot (Lond) 84: 123–133 [Google Scholar]
- Maehly AC, Chance B (1954) The assay of catalases and peroxidases. Methods Biochem Anal 1: 357–424 [DOI] [PubMed] [Google Scholar]
- Marschner H. (1995) Functions of mineral nutrients: macronutrients. In H Marschner, ed, Mineral Nutrition of Higher Plants. Academic Press, New York, pp 299–312 [Google Scholar]
- Marshall NM, Garner DK, Wilson TD, Gao YG, Robinson H, Nilges MJ, Lu Y (2009) Rationally tuning the reduction potential of a single cupredoxin beyond the natural range. Nature 462: 113–116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masclaux-Daubresse C, Daniel-Vedele F, Dechorgnat J, Chardon F, Gaufichon L, Suzuki A (2010) Nitrogen uptake, assimilation and remobilization in plants: challenges for sustainable and productive agriculture. Ann Bot (Lond) 105: 1141–1157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mäser P, Thomine S, Schroeder JI, Ward JM, Hirschi K, Sze H, Talke IN, Amtmann A, Maathuis FJ, Sanders D, et al. (2001) Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol 126: 1646–1667 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mhamdi A, Queval G, Chaouch S, Vanderauwera S, Van Breusegem F, Noctor G (2010) Catalase function in plants: a focus on Arabidopsis mutants as stress-mimic models. J Exp Bot 61: 4197–4220 [DOI] [PubMed] [Google Scholar]
- Mian A, Oomen RJ, Isayenkov S, Sentenac H, Maathuis FJ, Véry AA (2011) Over-expression of an Na+-and K+-permeable HKT transporter in barley improves salt tolerance. Plant J 68: 468–479 [DOI] [PubMed] [Google Scholar]
- Munns R. (1993) Physiological processes limiting plant growth in saline soils: some dogmas and hypotheses. Plant Cell Environ 16: 15–24 [Google Scholar]
- Munns R. (2002) Comparative physiology of salt and water stress. Plant Cell Environ 25: 239–250 [DOI] [PubMed] [Google Scholar]
- Nischal L, Mohsin M, Khan I, Kardam H, Wadhwa A, Abrol YP, Iqbal M, Ahmad A (2012) Identification and comparative analysis of microRNAs associated with low-N tolerance in rice genotypes. PLoS ONE 7: e50261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nuruzzaman M, Manimekalai R, Sharoni AM, Satoh K, Kondoh H, Ooka H, Kikuchi S (2010) Genome-wide analysis of NAC transcription factor family in rice. Gene 465: 30–44 [DOI] [PubMed] [Google Scholar]
- Pandey R, Joshi G, Bhardwaj AR, Agarwal M, Katiyar-Agarwal S (2014) A comprehensive genome-wide study on tissue-specific and abiotic stress-specific miRNAs in Triticum aestivum. PLoS ONE 9: e95800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pathak RR, Ahmad A, Lochab S, Raghuram N (2008) Molecular physiology of plant nitrogen use efficiency and biotechnological options for its enhancement. Current Science Bangalore 94: 1394 [Google Scholar]
- Pignocchi C, Fletcher JM, Wilkinson JE, Barnes JD, Foyer CH (2003) The function of ascorbate oxidase in tobacco. Plant Physiol 132: 1631–1641 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pignocchi C, Foyer CH (2003) Apoplastic ascorbate metabolism and its role in the regulation of cell signalling. Curr Opin Plant Biol 6: 379–389 [DOI] [PubMed] [Google Scholar]
- Rao G, Rao GR (1981) Pigment composition and chlorophyllase activity in pigeon pea (Cajanus indicus Spreng) and gingelley (Sesamum indicum L.) under NaCl salinity. Indian J Exp Biol 19: 768–770 [Google Scholar]
- Rizhsky L, Liang H, Shuman J, Shulaev V, Davletova S, Mittler R (2004) When defense pathways collide: the response of Arabidopsis to a combination of drought and heat stress. Plant Physiol 134: 1683–1696 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez RE, Mecchia MA, Debernardi JM, Schommer C, Weigel D, Palatnik JF (2010) Control of cell proliferation in Arabidopsis thaliana by microRNA miR396. Development 137: 103–112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanmartin M, Drogoudi PA, Lyons T, Pateraki I, Barnes J, Kanellis AK (2003) Over-expression of ascorbate oxidase in the apoplast of transgenic tobacco results in altered ascorbate and glutathione redox states and increased sensitivity to ozone. Planta 216: 918–928 [DOI] [PubMed] [Google Scholar]
- Satoh K, Doi K, Nagata T, Kishimoto N, Suzuki K, Otomo Y, Kawai J, Nakamura M, Hirozane-Kishikawa T, Kanagawa S, et al. (2007) Gene organization in rice revealed by full-length cDNA mapping and gene expression analysis through microarray. PLoS ONE 2: e1235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma D, Tiwari M, Lakhwani D, Tripathi RD, Trivedi PK (2015) Differential expression of microRNAs by arsenate and arsenite stress in natural accessions of rice. Metallomics 7: 174–187 [DOI] [PubMed] [Google Scholar]
- Shinozaki K, Yamaguchi-Shinozaki K (2007) Gene networks involved in drought stress response and tolerance. J Exp Bot 58: 221–227 [DOI] [PubMed] [Google Scholar]
- Solomon EI, Szilagyi RK, DeBeer George S, Basumallick L (2004) Electronic structures of metal sites in proteins and models: contributions to function in blue copper proteins. Chem Rev 104: 419–458 [DOI] [PubMed] [Google Scholar]
- Takahashi M, Sasaki Y, Ida S, Morikawa H (2001) Nitrite reductase gene enrichment improves assimilation of NO2 in Arabidopsis. Plant Physiol 126: 731–741 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thiebaut F, Rojas CA, Grativol C, Motta MR, Vieira T, Regulski M, Martienssen RA, Farinelli L, Hemerly AS, Ferreira PC (2014) Genome-wide identification of microRNA and siRNA responsive to endophytic beneficial diazotrophic bacteria in maize. BMC Genomics 15: 766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turan S, Cornish K, Kumar S (2012) Salinity tolerance in plants: breeding and genetic engineering. Aust J Crop Sci 6: 1337–1348 [Google Scholar]
- Varkonyi-Gasic E, Wu R, Wood M, Walton EF, Hellens RP (2007) Protocol: a highly sensitive RT-PCR method for detection and quantification of microRNAs. Plant Methods 3: 12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Véry AA, Sentenac H (2002) Cation channels in the Arabidopsis plasma membrane. Trends Plant Sci 7: 168–175 [DOI] [PubMed] [Google Scholar]
- Véry AA, Sentenac H (2003) Molecular mechanisms and regulation of K+ transport in higher plants. Annu Rev Plant Biol 54: 575–603 [DOI] [PubMed] [Google Scholar]
- Wang FZ, Wang QB, Kwon SY, Kwak SS, Su WA (2005) Enhanced drought tolerance of transgenic rice plants expressing a pea manganese superoxide dismutase. J Plant Physiol 162: 465–472 [DOI] [PubMed] [Google Scholar]
- Wu G, Park MY, Conway SR, Wang JW, Weigel D, Poethig RS (2009a) The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell 138: 750–759 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu L, Zhang Q, Zhou H, Ni F, Wu X, Qi Y (2009b) Rice microRNA effector complexes and targets. Plant Cell 21: 3421–3435 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiao H, Wang Y, Liu D, Wang W, Li X, Zhao X, Xu J, Zhai W, Zhu L (2003) Functional analysis of the rice AP3 homologue OsMADS16 by RNA interference. Plant Mol Biol 52: 957–966 [DOI] [PubMed] [Google Scholar]
- Xie F, Wang Q, Sun R, Zhang B (2015) Deep sequencing reveals important roles of microRNAs in response to drought and salinity stress in cotton. J Exp Bot 66: 789–804 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu Z, Zhong S, Li X, Li W, Rothstein SJ, Zhang S, Bi Y, Xie C (2011) Genome-wide identification of microRNAs in response to low nitrate availability in maize leaves and roots. PLoS ONE 6: e28009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan J, Zhang H, Zheng Y, Ding Y (2015) Comparative expression profiling of miRNAs between the cytoplasmic male sterile line MeixiangA and its maintainer line MeixiangB during rice anther development. Planta 241: 109–123 [DOI] [PubMed] [Google Scholar]
- Yang T, Zhang S, Hu Y, Wu F, Hu Q, Chen G, Cai J, Wu T, Moran N, Yu L, et al. (2014) The role of a potassium transporter OsHAK5 in potassium acquisition and transport from roots to shoots in rice at low potassium supply levels. Plant Physiol 166: 945–959 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeo A. (1998) Molecular biology of salt tolerance in the context of whole-plant physiology. J Exp Bot 49: 915–929 [Google Scholar]
- Zanca AS, Vicentini R, Ortiz-Morea FA, Del Bem LE, da Silva MJ, Vincentz M, Nogueira FT (2010) Identification and expression analysis of microRNAs and targets in the biofuel crop sugarcane. BMC Plant Biol 10: 260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Z, Wei L, Zou X, Tao Y, Liu Z, Zheng Y (2008) Submergence-responsive microRNAs are potentially involved in the regulation of morphological and metabolic adaptations in maize root cells. Ann Bot (Lond) 102: 509–519 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou M, Li D, Li Z, Hu Q, Yang C, Zhu L, Luo H (2013) Constitutive expression of a miR319 gene alters plant development and enhances salt and drought tolerance in transgenic creeping bentgrass. Plant Physiol 161: 1375–1391 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou M, Luo H (2013) MicroRNA-mediated gene regulation: potential applications for plant genetic engineering. Plant Mol Biol 83: 59–75 [DOI] [PubMed] [Google Scholar]
- Zhu JK. (2002) Salt and drought stress signal transduction in plants. Annu Rev Plant Biol 53: 247–273 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.