

Abstract:
Seminiferous tubular atrophy may involve indirectly the disruption of estrogen receptor-α (ESR1) function in efferent ductules of the testis. ESR1 helps to maintain fluid resorption by the ductal epithelium and the inhibition or stimulation of this activity in rodent species will lead to fluid accumulation in the lumen. If not resolved, the abnormal buildup of fluid in the head of the epididymis and efferent ductules becomes a serious problem for the testis, as it leads to an increase in testis weight, tubular dilation and seminiferous epithelial degeneration, as well as testicular atrophy. The same sequence of pathogenesis occurs if the efferent ductule lumen becomes occluded. This review provides an introduction to the role of estrogen in the male reproductive tract but focuses on the various overlapping mechanisms that could induce efferent ductule dysfunction and fluid backpressure histopathology. Although efferent ductules are difficult to find, their inclusion in routine histological evaluations is recommended, as morphological images of these delicate tubules may be essential for understanding the mechanism of testicular injury, especially if dilations are observed in the rete testis and/or seminiferous tubules.
Signature Lesion:
The rete testis and efferent ductules can appear dilated, as if the lumens were greatly expanded with excess fluid or the accumulation of sperm. Because the efferent ductules resorb most of the fluid arriving from the rete testis lumen, one of two mechanisms is likely to be involved: a) reduced fluid uptake, which has been caused by the disruption in estrogen receptor signaling or associated pathways; or b) an increased rate of fluid resorption, which results in luminal occlusion. Both mechanisms can lead to a temporary increase in testicular weight, tubular dilation and atrophy of the seminiferous tubules.
Keywords: Testis, Histopathology, Efferent ductules, Epididymis, Rete testis, Sperm granuloma, Atrophy, Estrogen receptor, Ion and water transport
Introduction
Testicular atrophy is one of the more easily recognized endpoints in male reproductive pathology; however, an interpretation of the mechanism causing seminiferous tubular atrophy is not always easy to uncover. The observation of luminal dilation in the rete testis and/or seminiferous tubules is a signature lesion that could lead one to conclude that testicular atrophy may be a long-term outcome. It has been known since 1924 that occlusion of the efferent ductules near the rete testis will induce increased pressure within the seminiferous tubules and lead to testicular atrophy.1 Yet, the literature is filled with long-term studies showing testicular atrophy, without histopathological evaluation of the efferent ductule region. This is partially due to the difficulty in finding these delicate tubules that are buried in the epididymal fat pad of rodents,2 but also because for years most authors considered these ducts to be nothing more than a conduit from rete testis to the epididymis.3 However, evidence began to reveal that disruption of the kidney-like function of efferent ductules could result in fluid accumulation within the rete testis and seminiferous tubules and eventually testicular atrophy.4,5 One of the disrupting pathways uncovered was estrogen receptor-α (ESR1).6
As early as the 1930's, it was known that developmental exposure to high doses of natural estrogens, as well as diethylstilbestrol (DES) could induce malformation of the male reproductive tract.7-9 However, the prevailing hypothesis to explain these data was that estrogen exposure disrupted testosterone and its metabolite 5α-dihydrotestosterone (DHT), the dominant male sex steroid10 and that estrogen did not have a distinct function in the adult male reproductive tract, but rather played a role in early development during the ambisexual stage and in establishing male behavioral patterns.11 In 1997, examination of the estrogen receptor α knockout mouse (Esr1KO) revealed that ESR1 has a major function in regulating fluid resorption in efferent ductules of the testis,6 which is essential for increasing the concentration of sperm and their maturational development in the head of the epididymis.12-14
Efferent ductules are small, coiled tubules that transport sperm rapidly from rete testis chambers to the epididymal head (Fig. 1). In rodent species, efferent ducts are buried in the epididymal fat pad, beginning as 3-7 individual wide-lumen ducts but merging into a single, highly convoluted tubule with a narrow lumen under the capsule of the initial segment of the epididymis.5 In man and larger mammals, these ductules are more numerous than in rodent species and open independently into the epididymis at multiple sites in the caput epididymis. Most importantly, these ductules form the major portion of the caput region within a densely organized connective tissue that is attached to the tunica albuginea of the testis.5 The discovery that ESR1 is essential for male fertility altered our view of the role that efferent ductules play in the head of the epididymis and provided the basis for testing new hypotheses to explain numerous observed pathologies in the testis and epididymis.3,4,15-18 Several reviews have been written about estrogen's function in the male reproductive tract and should be examined for a more detailed understanding of its molecular interactions and physiological relevance.12-14,19,20 However, histopathological changes in testis and epididymis following ESR1 disruption were found to be similar to those observed after exposures to several environmental compounds and some classes of therapeutic biological products, as well as surgical ligation of the ductules. Therefore, this review will focus on some common histopathological responses of the efferent ductules and head of the epididymis that induce fluid accumulation in the testis, which may contribute to the atrophy of seminiferous tubules.
Figure 1.
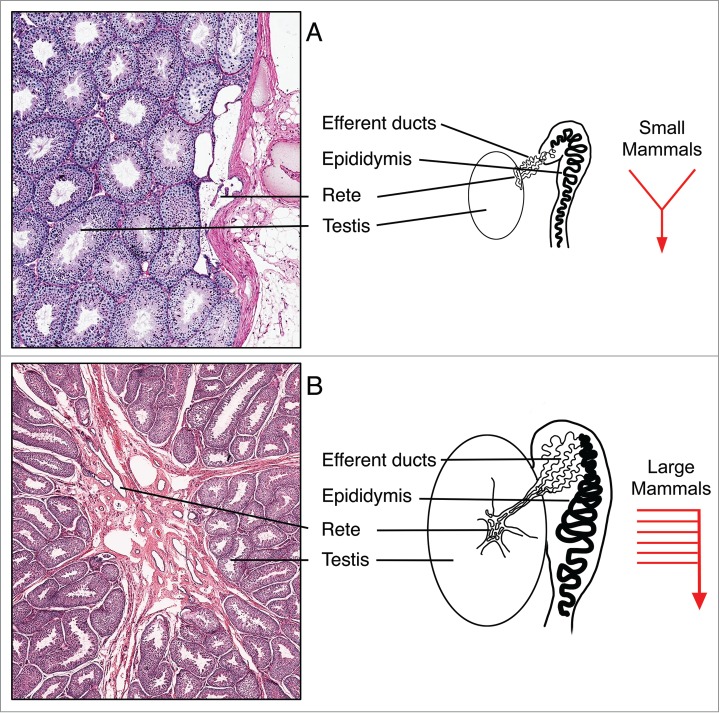
Basic organizational patterns of the rete testis and efferent ductules in small and large mammals. (A) In smaller mammals, such as rats and mice, the rete testis forms flattened chambers adjacent to the tunica albuginea of the testis, where sperm and tubular fluids are released into 3-7 efferent ductules that merge to form a single, highly convoluted common duct that enters the initial segment epididymis. (B) In larger mammals, including dogs and man, the rete testis forms flattened chambers surrounded by dense connective tissue within the mediastinum of the testis, which drains toward the efferent ductules that occupy a major portion of the caput epididymis. Most of the efferent ductules open individually into the caput epididymis.
Source of Estrogen in the Male Reproductive Tract
Estrogen synthesis is controlled by the aromatase enzyme complex of cytochrome P450 (P450arom) encoded by the CYP19 gene and a ubiquitous NADPH cytochrome P450 reductase.21 Testis is a major site for estrogen synthesis in the male and for many years it was assumed that Sertoli cells were the primary source during development, but in the adult only Leydig cells produced estrogen.22 Immunolocalization of P450arom was a major challenge, but in 1993 Nitta et al.23 became the first laboratory to demonstrate its presence in the mammalian spermatid (Fig. 2) and cytoplasmic droplets of sperm traversing the epididymis.24,25 The high concentrations of systemic androgens throughout the body are a blunt force on nearly every tissue in the male, but the unique system of estrogen synthesis in the male reproductive system creates a sequestered androgen/estrogen balance that can be focused specifically on cells expressing the requisite steroid receptors.
Figure 2.
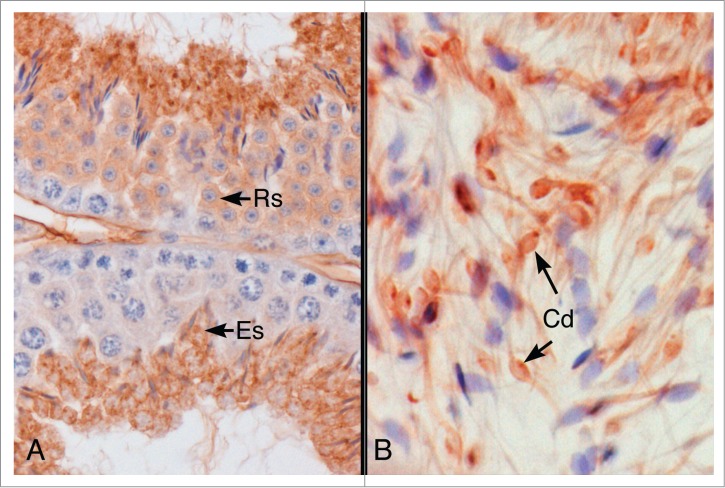
Immunohistochemical localization of P450 aromatase protein in the mouse testis and epididymis. (A) Aromatase protein was localized in the cytoplasm of round (RS) and elongated spermatids (ES) in the mouse seminiferous epithelium. (B) Caput epididymal lumen. Aromatase protein was localized in the cytoplasmic droplet (Cd) and along the thin tails of the spermatozoa.
It was surprising that the P450arom knockout mouse (AromKO) did not show histopathological results26-29 similar to the Esr1KO mouse.6,12 Testicular degeneration in the AromKO male began with ageing and was independent of the efferent ductule abnormalities found in the Esr1KO. Several explanations have been proposed and some have been tested. First, ESR1 expression in the efferent ductule epithelium is constitutive and thus continues to be expressed in the absence of natural ligand29,30 and could be activated in a ligand-independent manner.31-33 It is also possible that an ever-present ESR1, in the absence of estradiol, could bind a metabolite of DHT or other steroids that are present in high concentrations in the male.34 Finally, dietary phytoestrogens have also been shown to be sufficient for activation of Esr1-mediated pathways in the AromKO male28 and to increase the concentration of cauda epididymal sperm in Wild-type and Esr1KO mice35 It has been suggested that dietary phytoestrogens may be ‘agonistic’ in the absence of endogenous estrogen but ‘antagonistic’ when endogenous estrogens are present.12 Treatment with an aromatase inhibitor36-38 also showed no effect on efferent ductule morphology, but did decrease the expression of ESR2 and GPER, while increasing androgen receptor (AR) in the rat. It also delayed the development of the head of the epididymis. Thus, disruption of estrogen pathways in the male can lead to subtle or delayed histopathological results and depend on the presence or absence of its receptors, which are constitutively expressed in efferent ductules.29,30
Estrogen Receptors in the Male
The presence of the female hormone in the male reproductive tract suggested that the target cell and tissue for this luminal estrogen could be the epididymal epithelium, luminal sperm or even the female reproductive tract. Classical mediation of estrogen function is through two estrogen receptors, ERα (ESR1) and ERβ (ESR2), which are members of the nuclear receptor family of transcription factors and bind to estrogen response elements to mediate gene transcription.39-41 It has been known for 35 years that an estrogen receptor-like protein exists in male reproductive tissues42 and that estradiol binding is very strong in efferent ductules and the initial segment epididymis.43 Subsequent studies confirmed this hypothesis, as the efferent ductules were found to express Esr1 mRNA 3.5-fold greater than uterine tissue44 and immunohistochemistry13,14,19 revealed intense co-localization of ESR1 and AR in both ciliated and nonciliated cells of the epithelium (Fig. 3).
Figure 3.
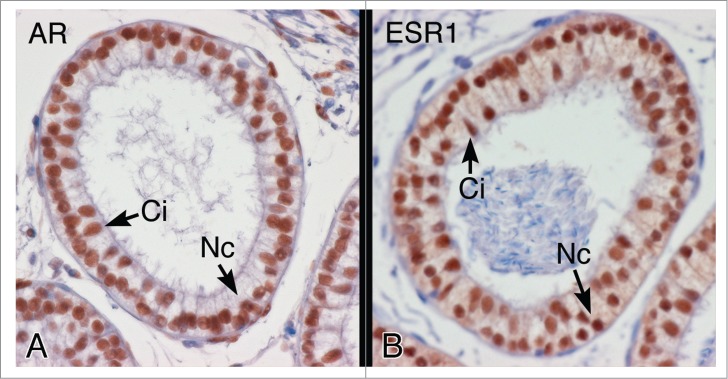
Androgen receptor (AR) and estrogen receptor-α (ESR1) protein in the efferent ductule epithelium of the hamster. (A) AR protein shows intense nuclear staining in both ciliated (Ci) and nonciliated (Nc) cells of the proximal efferent ductule epithelium. (B) ESR1 protein also shows intense nuclear staining in ciliated (Ci) and nonciliated (Nc) cells of the proximal efferent ductule epithelium.
In contrast, localization of ESR1 in the testis and epididymis has been a challenge, as major differences are found between species, as well as between individuals within a species. Results differ between immunohistochemical localization and mRNA analysis of testicular tissues and depend upon antibody source, age of development and experimental design.12-14,16,19,45-52 In general, most studies have concluded that testicular expression of ESR1 is low, but under certain conditions and in some species can be found in germ cells of the testis and sperm.12 On the other hand, ESR2 is expressed nearly ubiquitously throughout the male reproductive system.12,47 Therefore caution must be exercised when studying estrogen action in the testis. ESR1 expression in epididymis is also controversial, due to some studies showing no immunohistochemical staining while others using better fixation and optimal staining have found the protein both in cytoplasm and the nucleus.13
In addition to the genomic effects of estrogen, rapid non-genomic and membrane-associated responses have finally been recognized as indisputable pathways contributing to estrogen's role in specific cellular functions, including the male reproductive system.50,53-59 ESR1 and ESR2 are involved in rapid, non-genomic transduction effects of estradiol, but the G protein-coupled estrogen receptor-1 (GPER-1) also mediates multiple downstream signaling pathways.59-67 However, this area of investigation has become complicated because some studies have shown an ER antagonist inhibiting GPER-1 activity,68 while other studies show activation.69,70
Histopathology of Estrogen Receptor Dysfunction in Efferent ductules and Epididymis
Our acceptance of estrogen and its receptor, ESR1, having a major role in regulating fluid physiology in the male reproductive tract began with the analysis of the Esr1 knockout mice6,29,35, 71-87 and treatment of rodents and other species with the pure antiestrogen ICI 182,780 (ICI).6,37,46,51,58,67-69,75,80,85,88-116 Deletion of Esr1 gene caused male infertility, not only due to a disruption in male sexual behavior,31 but also because the sperm failed to mature properly in the male reproductive tract.117,118 Treatment with ICI induced subfertility at first, but over time complete infertility88,89 and resulted in numerous histopathological changes that were similar to those found in testes, efferent ductules and epididymides of the Esr1KO.
There are two basic mechanisms known to cause fluid accumulation and backpressure atrophy of the testis (Fig. 4): a) Inhibition of fluid resorption by the efferent ductule epithelium causing luminal dilution, and b) Compaction of the luminal contents causing occlusion of the efferent ductules. ESR1 disruption (Table 1) involves the first mechanism in rodent species because rodent efferent ductules have essentially a funnel-like design (Fig. 1). When the accumulation of luminal fluids exceeds the capacity of the single common duct exit, fluid pushes back into the testis causing dilation of rete testis and the seminiferous tubules.13,14,19
Figure 4.
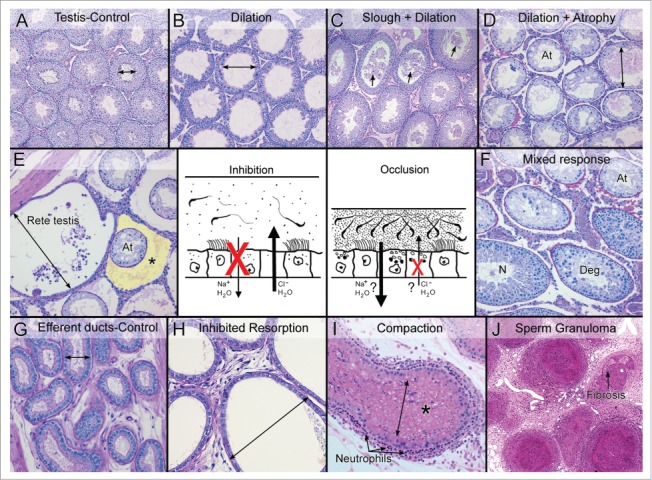
Two mechanisms lead to efferent ductule dysfunction and fluid accumulation in the testis. The central drawing illustrates the two mechanism of efferent ductule dysfunction that will result in the accumulation of luminal fluids and cause backpressure damage to the seminiferous tubules. The ‘inhibition’ mechanism involves the blockage of fluid resorption by inhibiting Na+ and water update and possibly an increase in Cl- and water movement into the lumen, thereby diluting the sperm and exceeding the drainage capacity of the ductules into the epididymis. The ‘occlusion’ mechanism involves excessive resorption and possibly an inhibition of Cl- secretion into the lumen. This mechanism results in a more viscous luminal environment, sperm stasis and eventually the occlusion or blockage of the ductule. (A) Control testis showing normal cross-sectional widths of the seminiferous tubular lumens. (B) Testis showing dilation of the seminiferous tubular lumen caused by the inhibition mechanism. Spermatogenesis appeared normal but there was thinning of the epithelium. (C) Testis showing dilation of the tubules caused by the occlusion mechanism. Sloughing of germ cells into the lumen (arrows) was also involved. (D) Testis showing seminiferous tubular atrophy (At), with some evidence of residual dilation after long-term occlusion of the efferent ductules. (E) Rete testis region showing excessive buildup of fluid and dilation, adjacent to atrophic (At) seminiferous tubules following the inhibition of fluid resorption. The yellow highlighted area (*) illustrates an edematous buildup around the atrophic tubules, which occurs in some cases but not in others. (F) Testis showing a mixed response following long-term occlusion of the efferent ductules. Atrophic tubules (At) are mixed with normal spermatogenesis (N) and degenerative changes (Deg). (G) Control efferent ductules in the conus region showing a normally narrow luminal diameter. (H) Efferent ductules at the proximal/conus junction following the inhibition of fluid resorption show excessive dilation and thinning of the epithelium. (I) Compaction of sperm within the lumen of the efferent ductules leads to dilation of the lumen and occlusion. This response caused the recruitment of polymorphonuclear leukocytes (neutrophils) into the wall lining the epithelium. (J) A long-term consequence of efferent ductule occlusion is the formation of sperm granulomas. The hyaline area shows the beginning of fibrosis.
Table 1.
Causes of efferent ductule dysfunction, with potential for the induction of testicular atrophy
| CAUSE | DESCRIPTION | POTENTIAL TARGETa | REFERENCES |
|---|---|---|---|
| CHEMICAL | |||
| ICI 182,780 | Fulvestrant | Inhibition of fluid resorption; blocks ESR1 and ESR2; similar to Esr1KO | 6,76,78,88-90 |
| GR40370X | 5-hydroxytryptamine receptor agonist; Serotonin-like, monoamine neurotransmitter | Inhibition of fluid resorption; vasoconstriction of venous plexus | 156 |
| PDE4 inhibitor | Phosphodiesterase-4 inhibitor | Inhibition of fluid resorption followed by occlusion; sperm granulomas | 147 |
| Uranyl nitrate hexahydrate | Dietary long-term exposure; proximal convoluted tubules of kidney sensitive | Inhibition of fluid resorption; progressive dilation of seminiferous tubules | 157 |
| LTI-1 | Leukotriene A(4) hydrolase inhibitor | Occlusion; dysregulation in fluid reabsorption; sperm granuloma | 2 |
| 6-chloro-6-deoxysugars | α-chlorohydrin-like chemicals | Occlusion; dysregulation of fluid resorption; sperm granuloma in efferent ductules; initial segment epididymis necrosis; inhibit glyceraldehyde-3-phosphate dehydrogenase | 148,158-165 |
| Isoproterenol | Beta-adrenergic agonist | Potential increase in rate of resorption; upregulates endothelin receptor-A; Et-1 increases Slc9a3 and inflammation | 166-168 |
| Benomylb | Methyl [1-[(butylamino)carbonyl]-1H-benzimidazol-2-yl]carbamate | Occlusion; microtubule disruption; germ cell sloughing; sperm granuloma | 4,130,131,135,136,169,170 |
| 2-Methylimidazole | Polymerization cross-linking and catalytic curing agent for epoxy resins | Occlusion; efferent duct sperm granuloma near caput epididymis | 171 |
| EDS | Ethane-1,2-dimethyl-sulfonate | Occlusion; alkylating agent, cellular toxicity; sperm granuloma | 158,172 |
| Cadmium | Chemical element, Cd | Occlusion; vascular endothelium; sperm granuloma | 173, 174 |
| 1,3-dinitrobenzene | m-Dinitrobenzene | Occlusion; impaired oxygen transport; sperm granuloma | 175, 176 |
| Dibutyl phthalate (DBP) | Di-n-butyl phthalate | Occlusion; prenatal exposure; epididymal malformation | 177 |
| Linuron | N - (3,4-dichlorophenyl)-N’-methoxy-N’-methylurea | Occlusion; herbicide; prenatal exposure; epididymal malformation | 178 |
| DES | Diethylstilbestrol | Neonatal exposure; decreases androgen receptor; sperm granuloma; dilation of lumen | 179-185 |
| Estradiol | β-estradiol 17-cypionate; 17β-estradiol; estradiol benzoate; ethinyl estradiol | Neonatal exposure; sperm granuloma; dilation lumen | 181, 182,186-188 |
| GENE MANIPULATIONd | |||
| Esr1 KO | Estrogen receptor-α | Inhibition of fluid resorption; decreases in SLC9A3, CA2, AQP-1, AQP-9, CAR14, SLC4A4; increases in CFTR, SLC9A1, SLC26A3 | 6,19,35,47,74, 75,77-80,97 |
| AF2ERKI MT | ESR1 AF-2 mutation | Inhibition of fluid resorption; blocks ESR1 AF-2 domain; similar to Esr1 KO | 97 |
| Slc9a3 KO | Sodium/hydrogen exchanger-3 | Inhibition of fluid resorption | 77 |
| Car2 MT | Carbonic anhydrase II | Inhibition of fluid resorption | 77 |
| Gpr64 KO | G protein-coupled receptor 64 (He6) | Inhibition of fluid resorption | 126 |
| He6 KO | GPR64; orphan member of the LNB-7TM (B(2)) subfamily of G-protein-coupled receptors | Inhibition of fluid resorption; proximal efferent ductules; partial sperm stasis | 126 |
| Lgr4 KO or MT | G protein-coupled receptor | Inhibition of fluid resorption; decreased expression of ESR1 and SLC9A3; also occlusion | 123, 124 |
| Prkar1a+/− | Protein kinase A (PKA) type Iα regulatory subunit (RIα) | Inhibition of fluid resorption; inhibition of Slc9a3 by over phosphorylation | 127 |
| Fst OE | Follistatin; inhibitor of activin | Inhibition of fluid resorption or ductule contraction; sperm stasis; decreased expression of ESR1 | 125 |
| Lfng KO | O-fucosylpeptide 3-beta-N-acetylglucosaminyltransferase | cNotch signaling; blocked connection with efferent ducts | 189 |
| Notch1 OE | Notch homolog 1, translocation-associated | cTransmembrane, oncogene, efferent ductule overgrowth | 190 |
| Pkd1 KO | Polycystic kidney disease 1 homolog | Abnormal epididymal development; dilation of efferent ductules | 191 |
| TE rat MT | Outbred Wistar strain | Autoimmune disorder; sperm granuloma | 192 |
| Dax1 KO | Nr0b1; transcription | cOcclusion; overgrowth of Sertoli cell and efferent duct epithelium | 193 |
| ProxE-AR or CEAR KO | Androgen receptor knockout in initial segment or caput epididymis | Occlusion; differentiation failure in caput epididymis; sperm granuloma | 194, 195 |
| Dicer1KO | Endoribonuclease; RNA interference | Occlusion; abnormal growth and blockage | 196 |
| HUMAN DISEASE | |||
| Von Hippel-Lindau disease | Papillary cystadenoma of the epididymis; also cystic kidney | Dysregulation of HIF1α; upregulation of vascular endothelial growth factor (VEGF) | 197-201 |
| Young's syndrome | Chronic sinopulmonary infections; azoospermia | Abnormal secretion or resorption; occlusion of caput and middle epididymis | 202-205 |
| Varicocele | Dilation of veins near rete testis and efferent ductules | Occlusion; compression of excurrent ducts and edema; blockage | 206 |
| Spontaneous granuloma | Caput epididymis efferent ductules | Occlusion; sperm granuloma; fibrosis; recanalization | 207-209 |
| Renal failure | Renal dialysis; renal malformations; renal cysts | Dilation of rete testis and epididymis; can lead to occlusion; intraductal calcium oxalate deposits | 210-216 |
| PHYSICAL | |||
| Ligation of ductules | Surgical blockage | Fluid accumulation; greater testicular effects when occluded closer to the rete testis | 1,129,133,150,151,217-225 |
| Arterial occlusion | Superior epididymal artery | Occlusion; localized ischaemia, sperm granuloma | 151,226,227 |
aPotential target for mechanisms in efferent ductules and rete testis, not necessarily testis or other organs.
bIncluding its metabolite carbendazim.
cOcclusion involves overgrowth of epithelium in rete testis and efferent ductules, but may also involve disruption of fluid reabsorption.
dGene knockout (KO); overexpression (OE); mutation (MT).
Histopathological changes in the male reproductive system following Esr1 disruption were consistent with the inhibition of fluid reabsorption by the efferent ductule epithelium.6 Severe dilation of the lumen (Fig. 5) was observed in the efferent ductules, rete testis and seminiferous tubules.6,29,35,71,74,76,78,79,86 Estrogen action through ESR1 regulates directly a number of major genes or indirectly several proteins involved in ion exchange and water transport in the efferent ductule epithelium. Most notably, ESR1 helps to maintain the activity of sodium/hydrogen exchanger-3 (SLC9A3) and aquaporins 1 and 9 (AQP1, AQP9), which facilitate the resorption of Na+ and water. Also ESR1 provides an inhibitory influence on the Cl- transporters cystic fibrosis transmembrane conductance regulator (CFTR) and Slc26a3 (DAR), as well as Na+/K+ ATPase α1 (Slc9a1), which would decrease the secretion of Cl- and movement of water at the luminal surface, while balancing the removal of cytoplasmic Na+ at the basal plasmalemma. Fluid resorption is further dependent on the endocytic apparatus of the nonciliated cells,119 which was also disorganized after the disruption of ESR1 activity.6,76,88,89,95
Figure 5.
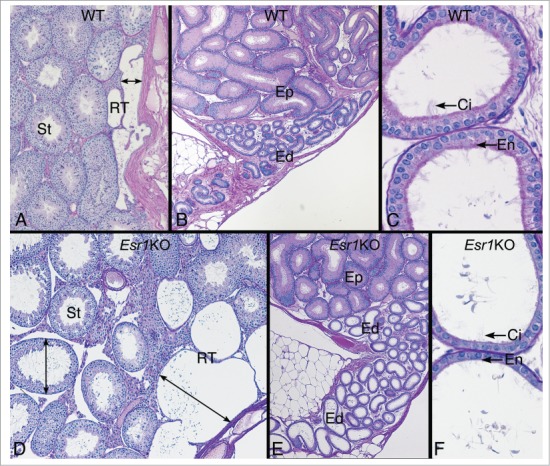
Testis and efferent ductules in the wild type (WT) and Esr1KO mice. (A) WT testis showing the narrow width of the rete testis and normal seminiferous tubules (St). (B) WT head of the epididymis region showing the coiled common efferent ductule (Ed) adjacent to the initial segment epididymis (Ep). (C) WT proximal region of the efferent ductules have a wider lumen than the common duct and show a PAS+ endocytic and brush border of microvilli (En) on the nonciliated cells and long cilia (Ci) protruding in the lumen from the ciliated cells. (D) Esr1KO testis showing dilated rete testis (RT) filled with fluid and causing dilation of seminiferous tubules (St). (E) Head of the epididymis region in the Esr1KO showing dilated efferent ductules (Ed) adjacent to the initial segment epididymis (Ep). (F) Esr1KO showing the dilated proximal region of the efferent ductules. The epithelium is shorter in height and appears to have lost PAS+ endocytic and brush border lining (En) on the nonciliated cells. Cilia (Ci) are noted but they appear to be thinner in density.
Recent studies have shown that estrogen works through the classical activation function (AF) domain, AF-1, but is regulated by the AF-2 domain.97 However, it also maintains a capability for ligand-independent activation in the efferent ductule epithelium, possibly working through phosphorylation of the AF-1 domain,120 or even its membrane receptor.13,50,53-59 Disruption of this ESR1 activity alters the luminal fluid composition, resulting in an alkaline, hypo-osmotic environment that resulted in abnormal sperm morphology.117,121 Treatment of the Esr1KO sperm with cAMP rescued all defective motility parameters.
In addition to the fluid-transport genes, estrogen also regulates several structural proteins responsible for maintenance of the efferent ductule epithelium. Loss of ESR1 activity resulted in significant alterations in epithelial morphology (Fig. 6). There was a 52% reduction in epithelial height, decreases in the endocytic apparatus, a dramatic reduction in the number and size of microvilli and also cilia.6,29,37,47,74-76,89,90,122 Thus, both direct effects (those regulating proteins necessary for ions and water fluxes) and indirect (epithelial morphology) were mediated by ESR1 inactivation in this critical region of the male tract.
Figure 6.
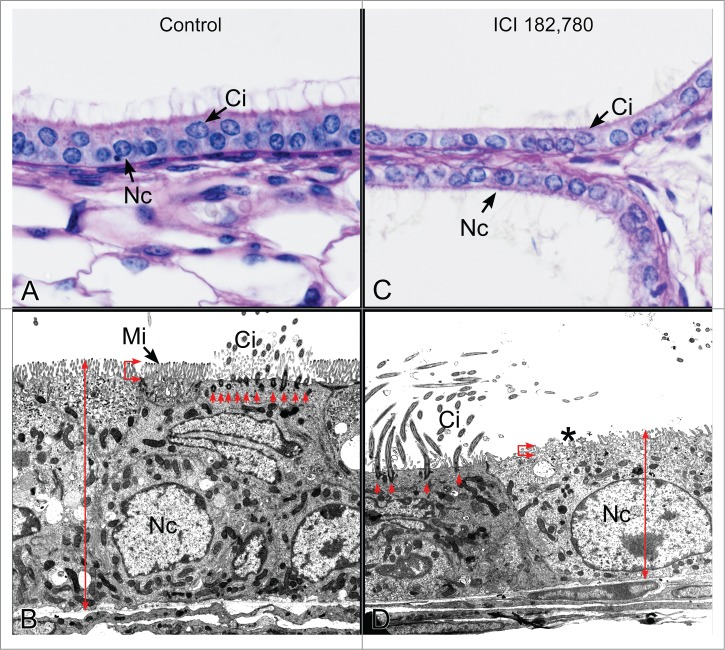
Efferent ductules from control and antiestrogen ICI 182,780 treated mice. (A) Light microscopy of the control proximal efferent ductule epithelium. Nc, nonciliated cell; Ci, ciliated cell. (B) Transmission electron microscopy of the control proximal efferent ductule epithelium. The nonciliated cell (Nc) has a short columnar height (double red arrow) and a prominent brush border of microvilli (Mi). The ciliated cell (Ci) has an abundance of basal bodies (red arrows) supporting the ciliary structures that protrude into the lumen. (C) Light microscopy of the ICI-treated proximal efferent ductule epithelium. The epithelium is shorter than normal and nonciliated cells (Nc) have a scant cytoplasm compared to the control. Ci, ciliated cell. (D) Transmission electron microscopy of the ICI-treated proximal efferent ductule epithelium. The nonciliated cell (Nc) is shorter in height (double red arrow) and is missing the normal finger-like projections of the microvillus border (*). The number of basal bodies (red arrows) supporting cilia (Ci) is greatly reduced.
Several other gene manipulation models and chemical treatments (Table 1) also inhibit fluid resorption in the efferent ductules, resulting in dilation of rete testis and seminiferous tubules. However, many of these appear to either decrease ESR1 activity97,123-125 or inhibit ESR1 associated pathways.77,126,127 Surprisingly, the knockout of two genes regulated by ESR1, Slc9a3 and Car2, produced normal epithelial morphology in the efferent ductules, while exhibiting luminal dilations of the ductules and rete testes that were greater than those observed in the Esr1KO.77 Thus, from a histopathological viewpoint, fluid accumulation with luminal dilation may or may not be associated with altered epithelial morphology. One explanation might be that efferent ductules adapt to the accumulation of fluid in the Slc9a3 and Car2 knockout mice and simply show excessive growth during development. When evaluating global gene knockout mice, it becomes difficult to separate developmental versus adult functions of a gene. This problem was seen in Esr1KO model, as the rete testis and efferent ductules were are already dilated at 10 days of age, prior to puberty.78 Therefore, treatment of the adult male with the antiestrogen ICI was necessary to show ESR1 regulation of both epithelial morphology and physiological function, separate from any developmental influence.
Finally, understanding estrogen activity in the epididymis has been a challenge because androgens have the primary role in its regulation.128 Historically, others have used castration followed by estrogen treatment models to study estrogen function in the epididymis. However, such studies must now be reinterpreted because ESR1 is constitutively expressed in efferent ductules after castration and high dosages of estradiol down-regulated both AR and ESR1.30,91 Thus, an interpretation of the castration model as being representative of estrogen's function in the epididymis appears to be invalid.
Histopathology of Occlusions in Efferent Ductules and the Epididymal Head
Compaction of the luminal contents with occlusion of the efferent ductules is the second basic mechanism known to cause fluid accumulation and backpressure atrophy of seminiferous tubules (Fig. 4). If sperm production and Sertoli cell secretions continue uninhibited following efferent ductule blockage, the following sequence of events will occur: a) proximal efferent ductules dilate and attempt to resorb the excess fluid; b) sperm become more compacted as fluid is resorbed; c) the rete testis begins to dilate and press into the testicular parenchyma; d) dilation of the seminiferous tubular lumens begins in regions proximal to the rete testis junction; e) tubular dilation in all regions of the testis may occur; f) spermatogenesis appears to be normal at first, but over time degenerative changes can appear; g) long term blockage of the proximal efferent ductules leads to cessation of spermatogenesis and tubular atrophy. From a practical point of view, one of the most sensitive indicators of fluid accumulation is the rapid increase in testis weight, which is often unilateral.129-133 However, the increase in testis weight, as well as severity of the tubular dilation and degeneration depends on a number of factors, including: how many efferent ductules were occluded; time elapsed since the onset of the occlusions; dosage of the offending compound; whether the compound also has direct effects on the seminiferous epithelium; if the common duct near the epididymis is involved; and species (mice show more resistance than rats to total atrophy).129-134
There are several potential mechanisms that could lead to the development of efferent ductule occlusions (Table 2). However, microtubule disruption provides one of the best examples of this category of histopathological responses, as illustrated by a single dose of the fungicide benomyl or its metabolite carbendazim.4,130,135 The response begins with a massive sloughing of elongated spermatids due to chemical disruption of microtubule polymerization in the Sertoli cells, which is followed by rapid transport of the sloughed cells into the epididymal lumen. Originally, it was hypothesized that the sloughed cells plugged the common efferent ductule lumen, but microdissection of treated ductules revealed that the occlusions were located primarily in the proximal region near the rete testis.136 Furthermore, several other compounds are known to induce sloughing of germ cells without inducing occlusions;137-141 thus, carbendazim appears to have direct effects on the ductal epithelium, as well as its known effects on the seminiferous epithelium.
Table 2.
Potential mechanisms for inducing occlusions in the head of the epididymis
| Cause | Potential Mechanismsa | References |
|---|---|---|
| Fluid resorption | Increase in the rate of Na+ uptake at the lumen; upregulate endothelin-1 or ET(A); increase in ESR1 expression | 126,136,149,166-168,196,228 |
| Microtubule disruption | Indirect effect on fluid resorption; disruption of epithelial recycle of apical vesicles and membrane proteins associated with ions and water transport | 136,142,144-146,149,229-235 |
| Inflammation | Inhibition of immune tolerance; extravasation of luminal germ cells; influx of macrophages and neutrophils; stretching of ductal epithelium | 2,147,171,236,237 |
| Leakage of fluid | Damage to the tight junctions of the vascular endothelium; leakage at the efferent ductal epithelium | 124,161,173, 174,209 |
| Ischemia | Inhibition of blood flow; dilation of veins; arterial occlusions; also damage to the endothelium | 151, 152,156,173,174,206,226, 227,238 |
| Sperm stasis | Inhibition of peritubular smooth muscle tone, either directly or indirectly through inhibition of sympathetic nerves | 147,239,240 |
| Developmental malformations | Abnormal growth that blocks the lumen | 123, 124,177, 178,194, 195 |
aThese are suggested mechanisms based on collective data and not necessarily direct association with efferent ductules and epididymis.
The potential direct effect of carbendazim on efferent ductules appears to be through the disruption of microtubule-dependent pathways responsible for membrane recycling along the microvillus border of the nonciliated cells. Although this hypothesis has not be tested in efferent ductules, in other tissues the turnover and displacement of ion and water transport proteins was disrupted with microtubule poisons,142-146 which could cause an increased rate of fluid resorption, sperm stasis and luminal compaction. Carbendazim has also been shown to increase the activity of Na+/K+-ATPase along the basolateral border of the nonciliated cells,136 which could be a normal response to an increase in Na+ flux at the luminal surface. However, other potential mechanisms should also be explored. For example, a carbendazim-like sperm granuloma with seminiferous tubular atrophy can be induced by a phosphodiesterase-4 inhibitor, which appears to inhibit fluid resorption rather than increase the rate of uptake.147 The unknown factor in both cases is the stimulus for polymorphonuclear leukocyte recruitment and subsequent formation of sperm granulomas.
Multiple pathways are likely involved in the onset of granuloma formation and ductal blockage and both mechanisms could overlap in some instances. For example, it has been known for many years that α-chlorohydrin inhibits glyceraldehyde-3-phosphate dehydrogenase (G3PDH) activity in spermatozoa but also induces efferent ductule sperm granulomas, similar to those observed with carbendazim. The occlusions were thought to be due to a disruption in blood flow.4,148 However, subsequent studies revealed that G3PDH is a microtubule-associated protein and 24 hours following α-chlorohydrin treatment β-tubulin disappears in the initial segment epithelium.149 If a similar effect is observed in the efferent ductule epithelium, then the mechanism leading to compaction of luminal sperm and formation of sperm granulomas following α-chlorohydrin treatment may overlap with that of carbendazim and indirectly be increasing the rate of fluid resorption.
Complications of histopathological interpretations
The interpretation of histopathological changes in the testis and head of the epididymis will depend on several common factors but also differ depending on which mechanism is causing the accumulation of fluid (Table 3). A major complication occurs if the seminiferous tubules and rete testis are dilated, but histological sections of the efferent ductules and initial segment epididymis have not been preserved. This is a serious problem because partial or total occlusion of the efferent ductules will produce fluid accumulation in the testis similar to the Esr1KO mouse; however, different mechanistic interpretations are required for each condition. Another major problem is time post exposure or post development. Occlusions of the proximal efferent ductules produce rapid increases in testicular weight and dilation of the tubules.150,151 However, when an occlusion or the inhibition of fluid resorption occurs further away from the rete testis, there can be a delay in the onset of increased testicular weight, with the delay taking up to several weeks.6,88,89 The more distal an occlusion occurs, the greater the surface areas of normal efferent ductule epithelium that will remain for continued resorption of luminal fluid, while the ductal wall stretches in diameter to accommodate the continual release of sperm and fluid from the testis.
Table 3.
Complications associated with histopathological interpretations of inhibited fluid resorption and sperm granulomas formation in the head of the epididymis
| INHIBITION OF FLUID RESORPTION | |
|---|---|
| Potential Efferent Ductule Effects | Histopathological Complications |
| Luminal dilation | Dilation may differ depending on region of the ductule; a time-response may be involved; blind ending ducts may confuse the interpretation 3,88,154 |
| Epithelial height decreasea | Can be absent even with large luminal dilation 77 |
| Endocytic apparatus decreasea | Can be absent even with large luminal dilation; could miss with poor fixation 77 |
| Microvillus border decrease in heighta | Can be absent even with large luminal dilation; could miss with poor fixation 77 |
| Potential Testicular Effects | |
| Testis weight increaseb | Species and time dependent; this can be transient; correlated with tubular dilation; must examine over time; may be unilateral 6,88,89 |
| Luminal dilation of rete testis | Species and time dependent; may be induced during development; may be unilateral; could miss observation in histology section 78,88,89,126 |
| Luminal dilation of seminiferous tubulesc | Species and time dependent; not all tubules will show equal effects; must section rete testis region, as this region may be more severe; luminal diameter may be dilated but tubular diameter may not be enlarged; may be unilateral 88, 89,126,147 |
| Seminiferous epithelial degeneration (multinucleated giant cells, vacuolation, sloughing, hypospermatogenesis, apoptosis) | Species and time dependent; correlated with tubular dilation; must examine over time; ranges from normal to mild to severe; rete testis proximity may be more severe; may lead to atrophy 88, 89,124, 125,147,156 |
| Atrophy of seminiferous tubulesb | Must examine after long-term effects; not all tubules will show equal effects; may be unilateral 88, 89,125 |
|
INDUCTION OF SPERM GRANULOMA |
|
| Potential Efferent Ductule Effects | Histopathological Complications |
| Luminal compaction of sperm | Dose and time dependent; not all ductules will show equal effects; proportional to dosage; may be unilateral; could miss observation in histology section 4,130,134, 135,171 |
| Neutrophilic granulocyte inflammation | Dose and time dependent; not all ductules will show equal effects; may subside with the onset of fibrosis 4, 130,134, 135 |
| Fibrosis | Must examine after long-term effects; may require serial sections |
| Recanalization | Must examine after long-term effects; may require serial sections 135,241,242 |
| Potential Testicular Effects | |
| Testis weight increaseb | Species and time dependent; this can be transient; correlated with tubular dilation; may be unilateral; must examine over time 4,130,131 |
| Rete testis lumen dilated | Depends on location of occlusion and species; proportional to dosage; may be unilateral 129,223-225 |
| Seminiferous tubular lumen dilated | Depends on location of occlusion and species; proportional to dosage; may be unilateral 129,223-225 |
| Atrophy of seminiferous tubulesb | Depends on location of occlusion and species; proportional to dosage; may be unilateral 129,171,223-225 |
aAppears to be ESR1 related.
bTransient increase, then decrease following seminiferous epithelial degeneration.
cDepends on the species and age or time post treatment or developmental.
Prior to seminiferous tubular atrophy, testicular histopathology can show a wide range of responses to fluid accumulation following ductal occlusions, depending on numerous factors already stated. Testicular dilation may be mild to moderate, with normal spermatogenesis or severe dilation with thinning of the seminiferous epithelium and cellular degeneration.131,150,151 Degenerative changes in the seminiferous epithelium may included the formation of multinucleated germ cells, sloughing of immature germ cells, epithelial vacuolation, hypospermatogenesis, and apoptosis.131,147,150-153 However, the testis and head of the epididymis have a remarkable capacity to adapt to the accumulation of fluid, as some testes having only one unobstructed efferent ductule still exhibited normal spermatogenesis in a limited number of seminiferous tubules,134 although an increase in atrophy was noted over a 70-day period.
Species considerations are always complicated, not only from a metabolism and target organ perspective, but also because the histopathology may differ significantly, without an obvious reason. Estrogen receptor studies provide a good example. The Esr1KO mouse testis showed an increase in testis weight and dilation of rete testis and seminiferous tubules over an 80-day period post birth,6 after which testis weight declined until total atrophy was observed. However, the knockout mouse was lacking ESR1 from development, therefore the pure antiestrogen ICI was used to determine if the same response would occur in the adult male. In the rat a similar time response was noted with testis weight and seminiferous tubular dilations, followed by total atrophy of the testis.89 However, the same treatment in the pubertal mouse gave confusing results.88 In the mouse, by day 8 post-treatment the efferent ductule lumen was dilated and epithelial structural integrity was already compromised, but the rete testis did not dilate until day 59. Furthermore, the mouse testis never increased in weight out to day 125 and atrophy was observed in only about 30% of the seminiferous tubules. Thus, the interpretation became complicated and we were never able to determine why backpressure atrophy did not occur with ICI treatment, even though the efferent ductules and rete testis exhibited nearly identical histopathological changes as seen in the Esr1KO mouse.
In the case of inhibited fluid resorption, it is unclear whether tubular atrophy is due to the fluid backpressure or a direct effect of the chemical, such as the antiestrogen ICI, on the seminiferous epithelium? In the Esr1 knock-in mouse (ENERKI), in which a point mutation in the ligand-binding domain of ESR1 allows for ligand-independent signaling,33 the efferent ductules were basically normal but with aging the testes showed focal seminiferous tubular atrophy similar to the ICI-treated mouse. Thus, blockage or physical ligation of the proximal efferent ductules of every species will result in testicular swelling and seminiferous tubular atrophy through rapid pressure-sensitive mechanisms,3 but long-term testicular effects of fluid accumulation following the inhibition of fluid resorption by the efferent ductule epithelium will depend on the species, the response time and other factors not yet uncovered.
Aberrant or blind-ending efferent ductules are an additional complication for histopathologists, as these small tubules are present in about 60% and 40% of the control testes/epididymides in rats and mice, respectively. The lumen of a blind-ending ductule is continuous with that of the male reproductive tract but is connected only at one end, presumably due to a failure in development from the mesonephric system of the embryo.74,154 In rodents, blind-ending tubules are smaller in diameter, have a collapsed lumen with no sperm, stain more intensely but lack the typical number of lysosomes in their cytoplasm. In larger mammals, such as the dog, bull and man, the blind-ending ductules are capable of accumulating stagnant sperm, dilating in size and forming sperm granulomas.155 Thus, the presence of these aberrant tubules must be taken into consideration when interpreting the histopathological responses observed in the head of the epididymis, but appear to be capable of contributing to ductal occlusions only in the larger species.
Conclusion
Disruption of efferent ductule epithelial function results in the accumulation of luminal fluids that is capable of backpressure into the rete testis and seminiferous tubules, causing transient dilation, epithelial degeneration and even testicular atrophy. This histopathological sequence was originally discovered following surgical ligation of the efferent ductules or treatment with chemicals that induced sperm granulomas in the head of the epididymis. However, a similar morphological sequela in the testis was also observed following the disruption of ESR1 function in the efferent ductules, which revealed the importance of preserving these delicate ducts for evaluation, but also brought attention to the role that estrogen plays in maintaining fluid resorption by the efferent ductal epithelium. Although efferent ductules are difficult to preserve for routine histological sectioning,2 their evaluation is essential for determining the mechanism of testicular injury if dilation is observed in the rete testis and/or seminiferous tubules, but also when unexplained seminiferous tubular atrophy is present in a long term study. Backpressure atrophy of the testis can be rapid and once the efferent ductules are occluded the lesion appears to be permanent.4
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- 1. Van Wagenen G. Degeneration of germinal epithelium in the testis of the rat as a result of efferent duct ligation. Anat Rec 1924; 27:189-90. [Google Scholar]
- 2. La DK, Creasy DM, Hess RA, Baxter E, Pereira ME, Johnson CA, Vinken P, Snook SS. Efferent duct toxicity with secondary testicular changes in rats following administration of a novel leukotriene a4 hydrolase inhibitor. Toxicol Pathol 2012; 40:705-14. [DOI] [PubMed] [Google Scholar]
- 3. Ilio KY, Hess RA. Structure and function of the ductuli efferentes: a review. Microsc Res Tech 1994; 29:432-67. [DOI] [PubMed] [Google Scholar]
- 4. Hess RA, Nakai M. Histopathology of the male reproductive system induced by the fungicide benomyl. Histol Histopathol 2000; 15:207-24. [DOI] [PubMed] [Google Scholar]
- 5. Hess RA. The Efferent Ductules: Structure and Functions. In: Robaire B, Hinton B, eds. The Epididymis: from Molecules to Clinical Practice. New York: Kluwer Academic/Plenum Publishers, 2002:49-80. [Google Scholar]
- 6. Hess RA, Bunick D, Lee KH, Bahr J, Taylor JA, Korach KS, Lubahn DB. A role for oestrogens in the male reproductive system. Nature 1997; 390:509-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Burrows H. Carcinoma mammae occurring in a male mouse under continued treatment with oestrin. Am J Cancer 1935; 24:613-6. [PubMed] [Google Scholar]
- 8. McLachlan JA, Newbold RR, Bullock B. Reproductive tract lesions in male mice exposed prenatally to diethylstilbestrol. Science 1975; 190:991-2. [DOI] [PubMed] [Google Scholar]
- 9. Wolff E, Ginglinger A. Sur la transformation des Poulets males en intersexues par injection d’hormone femelle (folliculine) aux embryons. Archs Anat Histol Embryol 1935; 20:219-78. [Google Scholar]
- 10. Pinilla L, Cocconi M, Zoppi S, Martini L. Effect of neonatal estrogenization on testosterone metabolism in the prostate and in the epididymis of the rat. J Steroid Biochem 1989; 32:459-65. [DOI] [PubMed] [Google Scholar]
- 11. Greco TL, Duello TM, Gorski J. Estrogen receptors, estradiol, and diethylstilbestrol in early development: the mouse as a model for the study of estrogen receptors and estrogen sensitivity in embryonic development of male and female reproductive tracts. Endocr Rev 1993; 14:59-71. [DOI] [PubMed] [Google Scholar]
- 12. Carreau S, Hess RA. Oestrogens and spermatogenesis. Philos Trans R Soc Lond B Biol Sci 2010; 365:1517-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Hess RA, Fernandes SA, Gomes GR, Oliveira CA, Lazari MF, Porto CS. Estrogen and its receptors in efferent ductules and epididymis. J Androl 2011; 32:600-13. [DOI] [PubMed] [Google Scholar]
- 14. Joseph A, Shur BD, Hess RA. Estrogen, efferent ductules, and the epididymis. Biol Reprod 2011; 84:207-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Klinefelter GR, Hess RA. Toxicity of the excurrent ducts and accessory organs in the male. In: Korach KS, ed. Reproductive and Developmental Toxicology. New York: Marcel Dekker, Inc, 1998:553-91. [Google Scholar]
- 16. Hess RA, Zhou Q, Nie R, Oliveira C, Cho H, Nakai M, Carnes K. Estrogens and epididymal function. Reprod Fertil Dev 2001; 13:273-83. [DOI] [PubMed] [Google Scholar]
- 17. Jorgensen N, Meyts ER, Main KM, Skakkebaek NE. Testicular dysgenesis syndrome comprises some but not all cases of hypospadias and impaired spermatogenesis. Int J Androl 2010; 33:298-303. [DOI] [PubMed] [Google Scholar]
- 18. Wohlfahrt-Veje C, Main KM, Skakkebaek NE. Testicular dysgenesis syndrome: foetal origin of adult reproductive problems. Clin Endocrinol (Oxf) 2009; 71:459-65. [DOI] [PubMed] [Google Scholar]
- 19. Hess RA, Zhou Q, Nie R. The Role of Estrogens in the Endocrine and Paracrine Regulation of the Efferent Ductules, Epididymis and Vas deferens. In: Robaire B, Hinton BT, eds. The Epididymis: from Molecules to Clinical Practice. New York: Kluwer Academic/Plenum Publishers, 2002:317-38. [Google Scholar]
- 20. Hess RA. Estrogen in the adult male reproductive tract: A review. Reprod Biol Endocrinol 2003; 1:52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Carreau S, Wolczynski S, Galeraud-Denis I. Aromatase, oestrogens and human male reproduction. Philos Trans R Soc Lond B Biol Sci 2010; 365:1571-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Hess RA. Oestrogen in fluid transport and reabsorption in efferent ducts of the male reproductive tract. Rev Reprod 2000; 5:84-92. [DOI] [PubMed] [Google Scholar]
- 23. Nitta H, Bunick D, Hess RA, Janulis L, Newton SC, Millette CF, Osawa Y, Shizuta Y, Toda K, Bahr JM. Germ cells of the mouse testis express P450 aromatase. Endocrinology 1993; 132:1396-401. [DOI] [PubMed] [Google Scholar]
- 24. Janulis L, Hess RA, Bunick D, Nitta H, Janssen S, Osawa Y, Bahr JM. Mouse epididymal sperm contain active P450 aromatase which decreases as sperm traverse the epididymis. J Androl 1996; 17:111-6. [PubMed] [Google Scholar]
- 25. Janulis L, Bahr JM, Hess RA, Janssen S, Osawa Y, Bunick D. Rat testicular germ cells and epididymal sperm contain active P450 aromatase. J Androl 1998; 19:65-71. [PubMed] [Google Scholar]
- 26. O’Donnell L, Robertson KM, Jones ME, Simpson ER. Estrogen and spermatogenesis. Endocr Rev 2001; 22:289-318. [DOI] [PubMed] [Google Scholar]
- 27. Robertson KM, O’Donnell L, Jones ME, Meachem SJ, Boon WC, Fisher CR, Graves KH, McLachlan RI, Simpson ER. Impairment of spermatogenesis in mice lacking a functional aromatase (cyp 19) gene. Proc Natl Acad Sci U S A 1999; 96:7986-91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Robertson KM, O’Donnell L, Simpson ER, Jones ME. The phenotype of the aromatase knockout mouse reveals dietary phytoestrogens impact significantly on testis function. Endocrinology 2002; 143:2913-21. [DOI] [PubMed] [Google Scholar]
- 29. Toda K, Okada T, Hayashi Y, Saibara T. Preserved tissue structure of efferent ductules in aromatase-deficient mice. J Endocrinol 2008; 199:137-46. [DOI] [PubMed] [Google Scholar]
- 30. Oliveira CA, Mahecha GA, Carnes K, Prins GS, Saunders PT, Franca LR, Hess RA. Differential hormonal regulation of estrogen receptors ER alpha and ER beta and androgen receptor expression in rat efferent ductules. Reproduction 2004; 128:73-86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. McDevitt MA, Glidewell-Kenney C, Weiss J, Chambon P, Jameson JL, Levine JE. Estrogen response element-independent estrogen receptor (ER)-alpha signaling does not rescue sexual behavior but restores normal testosterone secretion in male ERalpha knockout mice. Endocrinology 2007; 148:5288-94. [DOI] [PubMed] [Google Scholar]
- 32. Sinkevicius KW, Burdette JE, Woloszyn K, Hewitt SC, Hamilton K, Sugg SL, Temple KA, Wondisford FE, Korach KS, Woodruff TK, et al. . An estrogen receptor-alpha knock-in mutation provides evidence of ligand-independent signaling and allows modulation of ligand-induced pathways in vivo. Endocrinology 2008; 149:2970-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Sinkevicius KW, Laine M, Lotan TL, Woloszyn K, Richburg JH, Greene GL. Estrogen-dependent and -independent estrogen receptor-alpha signaling separately regulate male fertility. Endocrinology 2009; 150:2898-905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Picciarelli-Lima P, Oliveira AG, Reis AM, Kalapothakis E, Mahecha GA, Hess RA, Oliveira CA. Effects of 3-beta-diol, an androgen metabolite with intrinsic estrogen-like effects, in modulating the aquaporin-9 expression in the rat efferent ductules. Reprod Biol Endocrinol 2006; 4:51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Ruz R, Gregory M, Smith CE, Cyr DG, Lubahn DB, Hess RA, Hermo L. Expression of aquaporins in the efferent ductules, sperm counts, and sperm motility in estrogen receptor-alpha deficient mice fed lab chow versus casein. Mol Reprod Dev 2006; 73:226-37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Gomes GR, Yasuhara F, Siu ER, Fernandes SA, Avellar MC, Lazari MF, Porto CS. In Vivo Treatments with Fulvestrant and Anastrozole Differentially Affect Gene Expression in the Rat Efferent Ductules. Biol Reprod 2011; 84:52-61. [DOI] [PubMed] [Google Scholar]
- 37. Lazari MF, Lucas TF, Yasuhara F, Gomes GR, Siu ER, Royer C, Fernandes SA, Porto CS. Estrogen receptors and function in the male reproductive system. Arq Bras Endocrinol Metabol 2009; 53:923-33. [DOI] [PubMed] [Google Scholar]
- 38. Pearl CA, At-Taras E, Berger T, Roser JF. Reduced endogenous estrogen delays epididymal development but has no effect on efferent duct morphology in boars. Reproduction 2007; 134:593-604. [DOI] [PubMed] [Google Scholar]
- 39. Chang EC, Charn TH, Park SH, Helferich WG, Komm B, Katzenellenbogen JA, Katzenellenbogen BS. Estrogen Receptors alpha and beta as determinants of gene expression: influence of ligand, dose, and chromatin binding. Mol Endocrinol 2008; 22:1032-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Ellmann S, Sticht H, Thiel F, Beckmann MW, Strick R, Strissel PL. Estrogen and progesterone receptors: from molecular structures to clinical targets. Cell Mol Life Sci 2009; 66:2405-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Nilsson S, Gustafsson JA. Estrogen receptors: therapies targeted to receptor subtypes. Clinical pharmacology and therapeutics 2011; 89:44-55. [DOI] [PubMed] [Google Scholar]
- 42. Danzo BJ, Eller BC. The presence of a cytoplasmic estrogen receptor in sexually mature rabbit epididymides: comparison with the estrogen receptor in immature rabbit epididymal cytosol. Endocrinol 1979; 105:1128-34. [DOI] [PubMed] [Google Scholar]
- 43. Schleicher G, Drews U, Stumpf WE, Sar M. Differential distribution of dihydrotestosterone and estradiol binding sites in the epididymis of the mouse. An autoradiographic study. Histochemistry 1984; 81:139-47. [DOI] [PubMed] [Google Scholar]
- 44. Hess RA, Gist DH, Bunick D, Lubahn DB, Farrell A, Bahr J, Cooke PS, Greene GL. Estrogen receptor (alpha and beta) expression in the excurrent ducts of the adult male rat reproductive tract. J Androl 1997; 18:602-11. [PubMed] [Google Scholar]
- 45. Ramesh R, Pearl CA, At-Taras E, Roser JF, Berger T. Ontogeny of androgen and estrogen receptor expression in porcine testis: effect of reducing testicular estrogen synthesis. Anim Reprod Sci 2007; 102:286-99. [DOI] [PubMed] [Google Scholar]
- 46. Berger T, Conley AJ, Van Klompenberg M, Roser JF, Hovey RC. Increased testicular Sertoli cell population induced by an estrogen receptor antagonist. Mol Cell Endocrinol 2013; 366:53-8. [DOI] [PubMed] [Google Scholar]
- 47. Hess RA, Carnes K. The role of estrogen in testis and the male reproductive tract: a review and species comparison. Anim Reprod 2004; 1:5-30. [Google Scholar]
- 48. Saunders PT, Sharpe RM, Williams K, Macpherson S, Urquart H, Irvine DS, Millar MR. Differential expression of oestrogen receptor alpha and beta proteins in the testes and male reproductive system of human and non-human primates. Mol Hum Reprod 2001; 7:227-36. [DOI] [PubMed] [Google Scholar]
- 49. Lucas TF, Lazari MF, Porto CS. Differential role of the estrogen receptors ESR1 and ESR2 on the regulation of proteins involved with proliferation and differentiation of Sertoli cells from 15-day-old rats. Mol Cell Endocrinol 2014; 382:84-96. [DOI] [PubMed] [Google Scholar]
- 50. Lucas TF, Pimenta MT, Pisolato R, Lazari MF, Porto CS. 17beta-estradiol signaling and regulation of Sertoli cell function. Spermatogenesis 2011; 1:318-24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Lucas TF, Siu ER, Esteves CA, Monteiro HP, Oliveira CA, Porto CS, Lazari MF. 17beta-estradiol induces the translocation of the estrogen receptors ESR1 and ESR2 to the cell membrane, MAPK3/1 phosphorylation and proliferation of cultured immature rat Sertoli cells. Biol Reprod 2008; 78:101-14. [DOI] [PubMed] [Google Scholar]
- 52. Lombardi AP, Royer C, Pisolato R, Cavalcanti FN, Lucas TF, Lazari MF, Porto CS. Physiopathological aspects of the Wnt/beta-catenin signaling pathway in the male reproductive system. Spermatogenesis 2013; 3:e23181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Revankar CM, Cimino DF, Sklar LA, Arterburn JB, Prossnitz ER. A transmembrane intracellular estrogen receptor mediates rapid cell signaling. Science 2005; 307:1625-30. [DOI] [PubMed] [Google Scholar]
- 54. Levin ER. Membrane oestrogen receptor alpha signalling to cell functions. J Physiol 2009; 587:5019-23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Sanden C, Broselid S, Cornmark L, Andersson K, Daszkiewicz-Nilsson J, Martensson UE, Olde B, Leeb-Lundberg LM. G protein-coupled estrogen receptor 1/G protein-coupled receptor 30 localizes in the plasma membrane and traffics intracellularly on cytokeratin intermediate filaments. Mol Pharmacol 2011; 79:400-10. [DOI] [PubMed] [Google Scholar]
- 56. Fietz D, Bakhaus K, Wapelhorst B, Grosser G, Gunther S, Alber J, Doring B, Kliesch S, Weidner W, Galuska CE, et al. . Membrane transporters for sulfated steroids in the human testis - cellular localization, expression pattern and functional analysis. PLoS One 2013; 8:e62638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Chimento A, Sirianni R, Casaburi I, Pezzi V. Role of Estrogen Receptors and G Protein-Coupled Estrogen Receptor in Regulation of Hypothalamus-Pituitary-Testis Axis and Spermatogenesis. Frontiers in endocrinology 2014; 5:1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Ge LC, Chen ZJ, Liu HY, Zhang KS, Liu H, Huang HB, Zhang G, Wong CK, Giesy JP, Du J, et al. . Involvement of activating ERK1/2 through G protein coupled receptor 30 and estrogen receptor alpha/beta in low doses of bisphenol A promoting growth of Sertoli TM4 cells. Toxicol Lett 2014. [DOI] [PubMed] [Google Scholar]
- 59. Prossnitz ER, Barton M. Estrogen Biology: New Insights into GPER Function and Clinical Opportunities. Mol Cell Endocrinol 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Han G, Li F, Yu X, White RE. GPER: a novel target for non-genomic estrogen action in the cardiovascular system. Pharmacol Res 2013; 71:53-60. [DOI] [PubMed] [Google Scholar]
- 61. Hammes SR, Levin ER. Minireview: Recent advances in extranuclear steroid receptor actions. Endocrinology 2011; 152:4489-95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Levin ER. Plasma membrane estrogen receptors. Trends Endocrinol Metab 2009; 20:477-82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Hammes SR, Levin ER. Extranuclear steroid receptors: nature and actions. Endocr Rev 2007; 28:726-41. [DOI] [PubMed] [Google Scholar]
- 64. Saint-Criq V, Kim SH, Katzenellenbogen JA, Harvey BJ. Non-genomic estrogen regulation of ion transport and airway surface liquid dynamics in cystic fibrosis bronchial epithelium. PLoS One 2013; 8:e78593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Filardo EJ, Thomas P. Minireview: G protein-coupled estrogen receptor-1, GPER-1: its mechanism of action and role in female reproductive cancer, renal and vascular physiology. Endocrinology 2012; 153:2953-62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Sirianni R, Chimento A, Ruggiero C, De Luca A, Lappano R, Ando S, Maggiolini M, Pezzi V. The novel estrogen receptor, G protein-coupled receptor 30, mediates the proliferative effects induced by 17beta-estradiol on mouse spermatogonial GC-1 cell line. Endocrinology 2008; 149:5043-51. [DOI] [PubMed] [Google Scholar]
- 67. Lucas TF, Royer C, Siu ER, Lazari MF, Porto CS. Expression and signaling of G protein-coupled estrogen receptor 1 (GPER) in rat sertoli cells. Biol Reprod 2010; 83:307-17. [DOI] [PubMed] [Google Scholar]
- 68. Bois C, Delalande C, Bouraima-Lelong H, Durand P, Carreau S. 17beta-Estradiol regulates cyclin A1 and cyclin B1 gene expression in adult rat seminiferous tubules. J Mol Endocrinol 2012; 48:89-97. [DOI] [PubMed] [Google Scholar]
- 69. Filardo EJ, Quinn JA, Frackelton AR, Jr., Bland KI. Estrogen action via the G protein-coupled receptor, GPR30: stimulation of adenylyl cyclase and cAMP-mediated attenuation of the epidermal growth factor receptor-to-MAPK signaling axis. Mol Endocrinol 2002; 16:70-84. [DOI] [PubMed] [Google Scholar]
- 70. Prossnitz ER, Maggiolini M. Mechanisms of estrogen signaling and gene expression via GPR30. Mol Cell Endocrinol 2009; 308:32-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Eddy EM, Washburn TF, Bunch DO, Goulding EH, Gladen BC, Lubahn DB, Korach KS. Targeted disruption of the estrogen receptor gene in male mice causes alteration of spermatogenesis and infertility. Endocrinol 1996; 137:4796-805. [DOI] [PubMed] [Google Scholar]
- 72. Dupont S, Krust A, Gansmuller A, Dierich A, Chambon P, Mark M. Effect of single and compound knockouts of estrogen receptors alpha (ERalpha) and beta (ERbeta) on mouse reproductive phenotypes. Development 2000; 127:4277-91. [DOI] [PubMed] [Google Scholar]
- 73. Lubahn DB, Moyer JS, Golding TS, Couse JF, Korach KS, Smithies O. Alteration of reproductive function but not prenatal sexual development after insertional disruption of the mouse estrogen receptor gene. Proc Natl Acad Sci USA 1993; 90:11162-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Hess RA, Bunick D, Lubahn DB, Zhou Q, Bouma J. Morphologic changes in efferent ductules and epididymis in estrogen receptor-alpha knockout mice. J Androl 2000; 21:107-21. [PubMed] [Google Scholar]
- 75. Lee KH, Hess RA, Bahr JM, Lubahn DB, Taylor J, Bunick D. Estrogen receptor alpha has a functional role in the mouse rete testis and efferent ductules. Biol Reprod 2000; 63:1873-80. [DOI] [PubMed] [Google Scholar]
- 76. Nakai M, Bouma J, Nie R, Zhou Q, Carnes K, Jassim E, Lubahn DB, Hess RA. Morphological analysis of endocytosis in efferent ductules of estrogen receptor-alpha knockout male mouse. Anat Rec 2001; 263:10-8. [DOI] [PubMed] [Google Scholar]
- 77. Zhou Q, Clarke L, Nie R, Carnes K, Lai LW, Lien YH, Verkman A, Lubahn D, Fisher JS, Katzenellenbogen BS, et al. . Estrogen action and male fertility: Roles of the sodium/hydrogen exchanger-3 and fluid reabsorption in reproductive tract function. Proc Natl Acad Sci U S A 2001; 98:14132-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Lee KH, Park JH, Bunick D, Lubahn DB, Bahr JM. Morphological comparison of the testis and efferent ductules between wild-type and estrogen receptor alpha knockout mice during postnatal development. J Anat 2009; 214:916-25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Weiss J, Bernhardt ML, Laronda MM, Hurley LA, Glidewell-Kenney C, Pillai S, Tong M, Korach KS, Jameson JL. Estrogen actions in the male reproductive system involve estrogen response element-independent pathways. Endocrinology 2008; 149:6198-206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Lee KH, Finnigan-Bunick C, Bahr J, Bunick D. Estrogen Regulation of Ion Transporter Messenger RNA Levels in Mouse Efferent Ductules Are Mediated Differentially Through Estrogen Receptor (ER) alpha and ERbeta. Biol Reprod 2001; 65:1534-41. [DOI] [PubMed] [Google Scholar]
- 81. Mahato D, Goulding EH, Korach KS, Eddy EM. Spermatogenic cells do not require estrogen receptor-alpha for development or function [see comments]. Endocrinology 2000; 141:1273-6. [DOI] [PubMed] [Google Scholar]
- 82. Rosenfeld CS, Ganjam VK, Taylor JA, Yuan X, Stiehr JR, Hardy MP, Lubahn DB. Transcription and translation of estrogen receptor-beta in the male reproductive tract of estrogen receptor-alpha knock-out and wild-type mice. Endocrinology 1998; 139:2982-7. [DOI] [PubMed] [Google Scholar]
- 83. Couse JF, Curtis Hewitt S, Korach KS. Receptor null mice reveal contrasting roles for estrogen receptor alpha and beta in reproductive tissues. J Steroid Biochem Mol Biol 2000; 74:287-96. [DOI] [PubMed] [Google Scholar]
- 84. Prins GS, Birch L, Couse JF, Choi I, Katzenellenbogen B, Korach KS. Estrogen imprinting of the developing prostate gland is mediated through stromal estrogen receptor alpha: studies with alphaERKO and betaERKO mice. Cancer Res 2001; 61:6089-97. [PubMed] [Google Scholar]
- 85. Akingbemi BT, Ge R, Rosenfeld CS, Newton LG, Hardy DO, Catterall JF, Lubahn DB, Korach KS, Hardy MP. Estrogen receptor-alpha gene deficiency enhances androgen biosynthesis in the mouse Leydig cell. Endocrinology 2003; 144:84-93. [DOI] [PubMed] [Google Scholar]
- 86. Gould ML, Hurst PR, Nicholson HD. The effects of oestrogen receptors {alpha} and {beta} on testicular cell number and steroidogenesis in mice. Reproduction 2007; 134:271-9. [DOI] [PubMed] [Google Scholar]
- 87. Karim Rumi MA, Dhakal P, Kubota K, Chakraborty D, Lei T, Larson MA, Wolfe MW, Roby KF, Vivian JL, Soares MJ. Generation of Esr1 knockout rats using zinc finger nuclease-mediated genome editing. Endocrinology 2014:en20132150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Cho HW, Nie R, Carnes K, Zhou Q, Sharief NA, Hess RA. The antiestrogen ICI 182,780 induces early effects on the adult male mouse reproductive tract and long-term decreased fertility without testicular atrophy. Reprod Biol Endocrinol 2003; 1:57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Oliveira CA, Carnes K, Franca LR, Hess RA. Infertility and testicular atrophy in the antiestrogen-treated adult male rat. Biol Reprod 2001; 65:913-20. [DOI] [PubMed] [Google Scholar]
- 90. Oliveira CA, Zhou Q, Carnes K, Nie R, Kuehl DE, Jackson GL, Franca LR, Nakai M, Hess RA. ER Function in the Adult Male Rat: Short- and Long-Term Effects of the Antiestrogen ICI 182,780 on the Testis and Efferent Ductules, without Changes in Testosterone. Endocrinology 2002; 143:2399-409. [DOI] [PubMed] [Google Scholar]
- 91. Oliveira CA, Nie R, Carnes K, Franca LR, Prins GS, Saunders PT, Hess RA. The antiestrogen ICI 182,780 decreases the expression of estrogen receptor-alpha but has no effect on estrogen receptor-beta and androgen receptor in rat efferent ductules. Reprod Biol Endocrinol 2003; 1:75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Oliveira CA, Carnes K, Franca LR, Hermo L, Hess RA. Aquaporin-1 and -9 are differentially regulated by estrogen in the efferent ductule epithelium and initial segment of the epididymis. Biol Cell 2005; 97:385-95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93. Shayu D, Kesava CC, Soundarajan R, Rao AJ. Effects of ICI 182780 on estrogen receptor expression, fluid absorption and sperm motility in the epididymis of the bonnet monkey. Reprod Biol Endocrinol 2005; 3:10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Anahara R, Toyama Y, Maekawa M, Yoshida M, Kai M, Ishino F, Toshimori K, Mori C. Anti-estrogen ICI 182,780 and anti-androgen flutamide induce tyrosine phosphorylation of cortactin in the ectoplasmic specialization between the Sertoli cell and spermatids in the mouse testis. Biochem Biophys Res Commun 2006; 346:276-80. [DOI] [PubMed] [Google Scholar]
- 95. Choi H, Seong CN, Park MS, Cho HW. Effects of alternate treatment of estrogen receptor antagonist and agonist on morphology of male reproductive organs of adult mice. Appl Microscopy 2013; 43:103-9. [Google Scholar]
- 96. Berger T, Conley A, Klompenberg MV, Roser JF, Hovey RC. Increased Testicular Sertoli Cell Population Induced by an Estrogen Receptor Antagonist. Mol Cell Endocrinol 2012. [DOI] [PubMed] [Google Scholar]
- 97. Arao Y, Hamilton KJ, Goulding EH, Janardhan KS, Eddy EM, Korach KS. Transactivating function (AF) 2-mediated AF-1 activity of estrogen receptor alpha is crucial to maintain male reproductive tract function. Proc Natl Acad Sci U S A 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Fernandes SA, Gomes GR, Siu ER, Damas-Souza DM, Bruni-Cardoso A, Augusto TM, Lazari MF, Carvalho HF, Porto CS. The anti-oestrogen fulvestrant (ICI 182,780) reduces the androgen receptor expression, ERK1/2 phosphorylation and cell proliferation in the rat ventral prostate. Int J Androl 2011; 34:486-500. [DOI] [PubMed] [Google Scholar]
- 99. Thuillier R, Mazer M, Manku G, Boisvert A, Wang Y, Culty M. Interdependence of platelet-derived growth factor and estrogen-signaling pathways in inducing neonatal rat testicular gonocytes proliferation. Biol Reprod 2010; 82:825-36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100. Chimento A, Sirianni R, Delalande C, Silandre D, Bois C, Ando S, Maggiolini M, Carreau S, Pezzi V. 17beta-estradiol activates rapid signaling pathways involved in rat pachytene spermatocytes apoptosis through GPR30 and ERalpha. Mol Cell Endocrinol 2010; 320:136-44. [DOI] [PubMed] [Google Scholar]
- 101. Oliveira CA, Victor-Costa AB, Hess RA. Cellular and regional distributions of ubiquitin-proteasome and endocytotic pathway components in the epithelium of rat efferent ductules and initial segment of the epididymis. J Androl 2009; 30:590-601. [DOI] [PubMed] [Google Scholar]
- 102. Deshpande SN, Vijayakumar G, Rao AJ. Oestrogenic regulation and differential expression of WNT4 in the bonnet monkey and rodent epididymis. Reprod Biomed Online 2009; 18:555-61. [DOI] [PubMed] [Google Scholar]
- 103. Alam S, Ohsako S, Matsuwaki T, Bo Zhu X, Tsunekawa N, Kanai Y, Sone H, Tohyama C, Kurohmaru M. Induction of spermatogenic cell apoptosis in prepubertal rat testes irrespective of testicular steroidogenesis: A possible estrogenic effect of di(n-butyl) phthalate. Reproduction 2009. [DOI] [PubMed] [Google Scholar]
- 104. Yasuhara F, Gomes GR, Siu ER, Suenaga CI, Marostica E, Porto CS, Lazari MF. Effects of the antiestrogen fulvestrant (ICI 182,780) on gene expression of the rat efferent ductules. Biol Reprod 2008; 79:432-41. [DOI] [PubMed] [Google Scholar]
- 105. Cobellis G, Cacciola G, Chioccarelli T, Izzo G, Meccariello R, Pierantoni R, Fasano S. Estrogen regulation of the male reproductive tract in the frog, Rana esculenta: a role in Fra-1 activation in peritubular myoid cells and in sperm release. Gen Comp Endocrinol 2008; 155:838-46. [DOI] [PubMed] [Google Scholar]
- 106. Shayu D, Hardy MP, Rao AJ. Delineating the role of estrogen in regulating epididymal gene expression. Soc Reprod Fertil Suppl 2007; 63:31-43. [PubMed] [Google Scholar]
- 107. Pinto PI, Singh PB, Condeca JB, Teodosio HR, Power DM, Canario AV. ICI 182,780 has agonistic effects and synergizes with estradiol-17 beta in fish liver, but not in testis. Reprod Biol Endocrinol 2006; 4:67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108. Ebling FJ, Nwagwu MO, Baines H, Myers M, Kerr JB. The hypogonadal (hpg) mouse as a model to investigate the estrogenic regulation of spermatogenesis. Hum Fertil (Camb) 2006; 9:127-35. [DOI] [PubMed] [Google Scholar]
- 109. Aquila S, Sisci D, Gentile M, Middea E, Catalano S, Carpino A, Rago V, Ando S. Estrogen Receptor (ER)alpha and ERbeta Are Both Expressed in Human Ejaculated Spermatozoa: Evidence of Their Direct Interaction with Phosphatidylinositol-3-OH Kinase/Akt Pathway. J Clin Endocrinol Metab 2004; 89:1443-51. [DOI] [PubMed] [Google Scholar]
- 110. Lassurguere J, Livera G, Habert R, Jegou B. Time- and dose-related effects of estradiol and diethylstilbestrol on the morphology and function of the fetal rat testis in culture. Toxicol Sci 2003; 73:160-9. [DOI] [PubMed] [Google Scholar]
- 111. Huynh H, Alpert L, Alaoui-Jamali MA, Ng CY, Chan TW. Co-administration of finasteride and the pure anti-oestrogen ICI 182,780 act synergistically in modulating the IGF system in rat prostate. J Endocrinol 2001; 171:109-18. [DOI] [PubMed] [Google Scholar]
- 112. Turner KJ, Morley M, MacPherson S, Millar MR, Wilson JA, Sharpe RM, Saunders PT. Modulation of gene expression by androgen and oestrogens in the testis and prostate of the adult rat following androgen withdrawal. Mol Cell Endocrinol 2001; 178:73-87. [DOI] [PubMed] [Google Scholar]
- 113. Gancarczyk M, Paziewska-Hejmej A, Carreau S, Tabarowski Z, Bilinska B. Dose- and photoperiod-dependent effects of 17beta-estradiol and the anti-estrogen ICI 182,780 on testicular structure, acceleration of spermatogenesis, and aromatase immunoexpression in immature bank voles. Acta Histochem 2004; 106:269-78. [DOI] [PubMed] [Google Scholar]
- 114. Villa R, Bonetti E, Penza ML, Iacobello C, Bugari G, Bailo M, Parolini O, Apostoli P, Caimi L, Ciana P, et al. . Target-specific action of organochlorine compounds in reproductive and nonreproductive tissues of estrogen-reporter male mice. Toxicol Appl Pharmacol 2004; 201:137-48. [DOI] [PubMed] [Google Scholar]
- 115. Russo M, Troncone G, Guarino FM, Angelini F, Chieffi P. Estrogen-induced Akt-1 activity in the lizard (Podarcis s. sicula) testis. Mol Reprod Dev 2005; 71:52-7. [DOI] [PubMed] [Google Scholar]
- 116. Cardoso L, Nascimento A, Royer C, Porto C, Lazari M. Locally produced relaxin may affect testis and vas deferens function in rats. Reproduction 2009. [DOI] [PubMed] [Google Scholar]
- 117. Joseph A, Shur BD, Ko C, Chambon P, Hess RA. Epididymal Hypo-Osmolality Induces Abnormal Sperm Morphology and Function in the Estrogen Receptor Alpha Knockout Mouse. Biol Reprod 2010; 82:958-67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118. Mahato D, Goulding EH, Korach KS, Eddy EM. Estrogen receptor-alpha is required by the supporting somatic cells for spermatogenesis. Mol Cell Endocrinol 2001; 178:57-63. [DOI] [PubMed] [Google Scholar]
- 119. Hermo L, Clermont Y, Morales C. Fluid-phase and adsorptive endocytosis in ciliated epithelial cells of the rat ductuli efferentes. Anat Rec 1985; 211:285-94. [DOI] [PubMed] [Google Scholar]
- 120. Rochette-Egly C. Nuclear receptors: integration of multiple signalling pathways through phosphorylation. Cellular signalling 2003; 15:355-66. [DOI] [PubMed] [Google Scholar]
- 121. Joseph A, Hess RA, Schaeffer DJ, Ko C, Hudgin-Spivey S, Chambon P, Shur BD. Absence of Estrogen Receptor Alpha Leads to Physiological Alterations in the Mouse Epididymis and Consequent Defects in Sperm Function. Biol Reprod 2010; 82:948-57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122. Hess RA. Estrogen in the Adult Male: From a Curiosity to Absolute Necessity. Ann Rev Biomed Sci 2004; 6:1-12. [Google Scholar]
- 123. Hoshii T, Takeo T, Nakagata N, Takeya M, Araki K, Yamamura K. LGR4 Regulates the Postnatal Development and Integrity of Male Reproductive Tracts in Mice. Biol Reprod 2007; 76:303-13. [DOI] [PubMed] [Google Scholar]
- 124. Mendive F, Laurent P, Van Schoore G, Skarnes W, Pochet R, Vassart G. Defective postnatal development of the male reproductive tract in LGR4 knockout mice. Dev Biol 2006; 290:421-34. [DOI] [PubMed] [Google Scholar]
- 125. Seachrist DD, Johnson E, Magee C, Clay CM, Graham JK, Veeramachaneni DN, Keri RA. Overexpression of follistatin in the mouse epididymis disrupts fluid resorption and sperm transit in testicular excurrent ducts. Biol Reprod 2012; 87:41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126. Davies B, Baumann C, Kirchhoff C, Ivell R, Nubbemeyer R, Habenicht UF, Theuring F, Gottwald U. Targeted deletion of the epididymal receptor HE6 results in fluid dysregulation and male infertility. Mol Cell Biol 2004; 24:8642-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Burton KA, McDermott DA, Wilkes D, Poulsen MN, Nolan MA, Goldstein M, Basson CT, McKnight GS. Haploinsufficiency at the Protein Kinase A RI? gene locus leads to fertility defects in male mice and men. Mol Endocrinol 2006; 20:2504-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128. Robaire B, Hamzeh M. Androgen action in the epididymis. J Androl 2011; 32:592-9. [DOI] [PubMed] [Google Scholar]
- 129. Oslund RM. Ligation of vasa efferentia in rats. Am J Physiol 1926; 77:83-90. [Google Scholar]
- 130. Nakai M, Hess RA, Moore BJ, Guttroff RF, Strader LF, Linder RE. Acute and long-term effects of a single dose of the fungicide carbendazim (methyl 2-benzimidazole carbamate) on the male reproductive system in the rat. J Androl 1992; 13:507-18. [PubMed] [Google Scholar]
- 131. Hess RA, Moore BJ, Forrer J, Linder RE, Abuel-Atta AA. The fungicide benomyl (methyl 1-(butylcarbamoyl)-2- benzimidazolecarbamate) causes testicular dysfunction by inducing the sloughing of germ cells and occlusion of efferent ductules. Fund Appl Toxicol 1991; 17:733-45. [DOI] [PubMed] [Google Scholar]
- 132. Creasy DM. Evaluation of testicular toxicology: a synopsis and discussion of the recommendations proposed by the Society of Toxicologic Pathology. Birth Defects Res Part B Dev Reprod Toxicol 2003; 68:408-15. [DOI] [PubMed] [Google Scholar]
- 133. Baillie AH. Observations comparing the effects of epididymal obstruction at various levels on the mouse testis with those of ischaemia. J Anat 1962; 96:335-54. [PMC free article] [PubMed] [Google Scholar]
- 134. Gotoh Y, Netsu J, Nakai M, Nasu T. Testicular damage after exposure to carbendazim depends on the number of patent efferent ductules. J Vet Med Sci 1999; 61:755-60. [DOI] [PubMed] [Google Scholar]
- 135. Nakai M, Moore BJ, Hess RA. Epithelial reorganization and irregular growth following carbendazim-induced injury of the efferent ductules of the rat testis. Anat Rec 1993; 235:51-60. [DOI] [PubMed] [Google Scholar]
- 136. Hess RA. Effects of environmental toxicants on the efferent ducts, epididymis and fertility. J Reprod Fertil Suppl 1998; 53:247-59. [PubMed] [Google Scholar]
- 137. Quintar AA, Nishioka S, Torres AI, Maldonado CA, Aoki A. The initial segment of the rat epididymis is able to uptake immature germ cells shed by testicular damage. Cells Tissues Organs 2011; 193:170-83. [DOI] [PubMed] [Google Scholar]
- 138. Kluwe WM. Acute toxicity of 1,2-dibromo-3-chloropropane in the F344 male rat. II. Development and repair of the renal, epididymal, testicular, and hepatic lesions. Toxicol Appl Pharmacol 1981; 59:84-95. [DOI] [PubMed] [Google Scholar]
- 139. Funabashi H, Fujioka M, Kohchi M, Tateishi Y, Matsuoka N. Collaborative work to evaluate toxicity on male reproductive organs by repeated dose studies in rats 22). Effects of 2- and 4-week administration of theobromine on the testis. J Toxicol Sci 2000; 25 Spec No:211-21. [DOI] [PubMed] [Google Scholar]
- 140. Chapin RE, Dutton SL, Ross MD, Sumrell BM, Lamb JCt. Development of reproductive tract lesions in male F344 rats after treatment with dimethyl methylphosphonate. Exp Mol Pathol 1984; 41:126-40. [DOI] [PubMed] [Google Scholar]
- 141. Chapin RE, Dutton SL, Ross MD, Sumrell BM, Lamb JCt. The effects of ethylene glycol monomethyl ether on testicular histology in F344 rats. J Androl 1984; 5:369-80. [DOI] [PubMed] [Google Scholar]
- 142. Elkjaer ML, Birn H, Agre P, Christensen EI, Nielsen S. Effects of microtubule disruption on endocytosis, membrane recycling and polarized distribution of Aquaporin-1 and gp330 in proximal tubule cells. Eur J Cell Biol 1995; 67:57-72. [PubMed] [Google Scholar]
- 143. Breton S, Nsumu NN, Galli T, Sabolic I, Smith PJ, Brown D. Tetanus toxin-mediated cleavage of cellubrevin inhibits proton secretion in the male reproductive tract. Am J Physiol Renal Physiol 2000; 278:F717-25. [DOI] [PubMed] [Google Scholar]
- 144. Pastor-Soler N, Bagnis C, Sabolic I, Tyszkowski R, McKee M, Van Hoek A, Breton S, Brown D. Aquaporin 9 expression along the male reproductive tract. Biol Reprod 2001; 65:384-93. [DOI] [PubMed] [Google Scholar]
- 145. Sabolic I, Herak-Kramberger CM, Ljubojevic M, Biemesderfer D, Brown D. NHE3 and NHERF are targeted to the basolateral membrane in proximal tubules of colchicine-treated rats. Kidney Int 2002; 61:1351-64. [DOI] [PubMed] [Google Scholar]
- 146. Maldonado-Baez L, Cole NB, Kramer H, Donaldson JG. Microtubule-dependent endosomal sorting of clathrin-independent cargo by Hook1. J Cell Biol 2013; 201:233-47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147. Heuser A, Mecklenburg L, Ockert D, Kohler M, Kemkowski J. Selective inhibition of PDE4 in Wistar rats can lead to dilatation in testis, efferent ducts, and epididymis and subsequent formation of sperm granulomas. Toxicol Pathol 2013; 41:615-27. [DOI] [PubMed] [Google Scholar]
- 148. Cooper ER, Jones AR, Jackson H. Effects of alpha-chlorohydrin and related compounds on the reproductive organs and fertility of the male rat. J Reprod Fertil 1974; 38:379-86. [DOI] [PubMed] [Google Scholar]
- 149. Jelks KB, Miller MG. alpha-Chlorohydrin inhibits glyceraldehyde-3-phosphate dehydrogenase in multiple organs as well as in sperm. Toxicol Sci 2001; 62:115-23. [DOI] [PubMed] [Google Scholar]
- 150. Harrison RG. The effect of ligation of the vasa efferentia on the rat testis. Proc Soc Study Fertil 1953; 7:97-100. [Google Scholar]
- 151. Macmillan EW. Higher epididymal obstructions in male infertility; etiology and treatment. Fertil Steril 1953; 4:101-27. [DOI] [PubMed] [Google Scholar]
- 152. Creasy DM. Pathogenesis of male reproductive toxicity. Toxicol Pathol 2001; 29:64-76. [DOI] [PubMed] [Google Scholar]
- 153. Creasy DM. Histopathology of the male reproductive system II: interpretation. Curr Protoc Toxicol 2002; Chapter 16:Unit16 4. [DOI] [PubMed] [Google Scholar]
- 154. Guttroff RF, Cooke PS, Hess RA. Blind-ending tubules and branching patterns of the rat ductuli efferentes. Anat Rec 1992; 232:423-31. [DOI] [PubMed] [Google Scholar]
- 155. Foley GL, Bassily N, Hess RA. Intratubular spermatic granulomas of the canine efferent ductules. Toxicol Pathol 1995; 23:731-4. [DOI] [PubMed] [Google Scholar]
- 156. Piner J, Sutherland M, Millar M, Turner K, Newall D, Sharpe RM. Changes in vascular dynamics of the adult rat testis leading to transient accumulation of seminiferous tubule fluid after administration of a novel 5-hydroxytryptamine (5-HT) agonist. Reprod Toxicol{Piner, 2002 #3901} 2002; 16:141-50. [DOI] [PubMed] [Google Scholar]
- 157. Mason KE, Shaver SL. Some functions of the caput epididymis. Ann N Y Acad Sci 1952; 55:585-93. [DOI] [PubMed] [Google Scholar]
- 158. Cooper ER, Jackson H. The vasa efferentia in the rat and mouse. J Reprod Fert 1972; 28:317-9. [DOI] [PubMed] [Google Scholar]
- 159. Hoffer AP, Hamilton DW, Fawcett DW. The ultrastructural pathology of the rat epididymis after administration of a-chlorhydrin (U-5897). I. Effects of a single high dose. Anat Rec 1973; 175:203-29. [DOI] [PubMed] [Google Scholar]
- 160. Ericsson RJ. Male antifertility compounds: U-5897 as a rat chemosterilant. J Reprod Fert 1970; 22:213-22. [DOI] [PubMed] [Google Scholar]
- 161. Reijonen K, Kormano M, Ericsson RJ. Studies on the rat epididymal blood vessels following alpha-chlorohydrin administration. Biol Reprod 1975; 12:483-90. [DOI] [PubMed] [Google Scholar]
- 162. Wong PYD, Yeung CH. Absorptive and secretory functions of the perfused rat cauda epididymis. J Physiol 1978; 275:13-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163. Wong PY, Yeung CH. Inhibition by alpha-chlorohydrin of fluid reabsorption in the rat cauda epididymidis. J Reprod Fertil 1977; 51:469-71. [DOI] [PubMed] [Google Scholar]
- 164. Ford WC, Waites GM. The effect of high doses of 6-chloro-6-deoxyglucose on the rat. Contraception 1981; 24:577-88. [DOI] [PubMed] [Google Scholar]
- 165. Ford WC, Waites GM. Activities of various 6-chloro-6-deoxysugars and (S) alpha-chlorohydrin in producing spermatocoeles in rats and paralysis in mice and in inhibiting glucose metabolism in bull spermatozoa in vitro. J Reprod Fertil 1982; 65:177-83. [DOI] [PubMed] [Google Scholar]
- 166. Filippi S, Vannelli GB, Granchi S, Luconi M, Crescioli C, Mancina R, Natali A, Brocchi S, Vignozzi L, Bencini E, et al. . Identification, localization and functional activity of oxytocin receptors in epididymis. Mol Cell Endocrinol 2002; 193:89-100. [DOI] [PubMed] [Google Scholar]
- 167. Fibbi B, Filippi S, Morelli A, Vignozzi L, Silvestrini E, Chavalmane A, De Vita G, Marini M, Gacci M, Manieri C, et al. . Estrogens regulate humans and rabbit epididymal contractility through the RhoA/Rho-kinase pathway. J Sex Med 2009; 6:2173-86. [DOI] [PubMed] [Google Scholar]
- 168. Cheng YS, Dai DZ, Dai Y. Testis dysfunction by isoproterenol is mediated by upregulating endothelin receptor A, leptin and protein kinase Cvarepsilon and is attenuated by an endothelin receptor antagonist CPU0213. Reprod Toxicol 2010; 29:421-6. [DOI] [PubMed] [Google Scholar]
- 169. Nakai M, Hess RA. Effects of carbendazim (methyl 2-benzimidazole carbamate; MBC) on meiotic spermatocytes and subsequent spermiogenesis in the rat testis. Anat Rec 1997; 247:379-87. [DOI] [PubMed] [Google Scholar]
- 170. Markelewicz RJ, Jr., Hall SJ, Boekelheide K. 2,5-hexanedione and Carbendazim coexposure synergistically disrupts rat spermatogenesis despite opposing molecular effects on microtubules. Toxicol Sci 2004; 80:92-100. [DOI] [PubMed] [Google Scholar]
- 171. Tani Y, Foster PM, Sills RC, Chan PC, Peddada SD, Nyska A. Epididymal sperm granuloma induced by chronic administration of 2-methylimidazole in B6C3F1 mice. Toxicol Pathol 2005; 33:313-9. [DOI] [PubMed] [Google Scholar]
- 172. Klinefelter GR, Laskey JW, Roberts NR, Slott V, Suarez JD. Multiple effects of ethane dimethanesulfonate on the epididymis of adult rats. Toxicol Appl Pharmacol 1990; 105:271-87. [DOI] [PubMed] [Google Scholar]
- 173. Waites GM, Setchell BP. Changes in blood flow and vascular permeability of the testis, epididymis and accessory reproductive organs of the rat after the administration of cadmium chloride. J Endocrinol 1966; 34:329-42. [DOI] [PubMed] [Google Scholar]
- 174. Mason KE, Young JO. Effects of cadmium upon the excurrent duct system of the rat testis. Anat Rec 1967; 159:311-24. [DOI] [PubMed] [Google Scholar]
- 175. Linder RE, Hess RA, Perreault SD, Strader LF, Barbee RR. Acute effects and long-term sequelae of 1,3-dinitrobenzene on male reproduction in the rat. I. Sperm quality, quantity, and fertilizing ability. J Androl 1988; 9:317-26. [DOI] [PubMed] [Google Scholar]
- 176. Linder RE, Hess RA, Strader LF. Testicular toxicity and infertility in male rats treated with 1,3-dinitrobenzene. J Toxicol Environ Health 1986; 19:477-89. [DOI] [PubMed] [Google Scholar]
- 177. Barlow NJ, Foster PM. Pathogenesis of male reproductive tract lesions from gestation through adulthood following in utero exposure to Di(n-butyl) phthalate. Toxicol Pathol 2003; 31:397-410. [DOI] [PubMed] [Google Scholar]
- 178. McIntyre BS, Barlow NJ, Wallace DG, Maness SC, Gaido KW, Foster PM. Effects of in utero exposure to linuron on androgen-dependent reproductive development in the male Crl:CD(SD)BR rat. Toxicol Appl Pharmacol 2000; 167:87-99. [DOI] [PubMed] [Google Scholar]
- 179. Miyaso H, Naito M, Hirai S, Matsuno Y, Komiyama M, Itoh M, Mori C. Neonatal exposure to diethylstilbestrol causes granulomatous orchitis via epididymal inflammation. Anat Sci Int 2014. [DOI] [PubMed] [Google Scholar]
- 180. Atanassova N, McKinnell C, Fisher J, Sharpe RM. Neonatal treatment of rats with diethylstilboestrol (DES) induces stromal-epithelial abnormalities of the vas deferens and cauda epididymis in adulthood following delayed basal cell development. Reproduction 2005; 129:589-601. [DOI] [PubMed] [Google Scholar]
- 181. Fisher JS, Pastor-Soler N, Sharpe RM, Breton S. Modulation of the onset of postnatal development of H(+)-ATPase-rich cells by steroid hormones in rat epididymis. Biol Reprod 2002; 67:1106-14. [DOI] [PubMed] [Google Scholar]
- 182. Fisher JS, Turner KJ, Brown D, Sharpe RM. Effect of neonatal exposure to estrogenic compounds on development of the excurrent ducts of the rat testis through puberty to adulthood. Environ Health Perspect 1999; 107:397-405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183. Fisher JS, Turner KJ, Fraser HM, Saunders PT, Brown D, Sharpe RM. Immunoexpression of aquaporin-1 in the efferent ducts of the rat and marmoset monkey during development, its modulation by estrogens, and its possible role in fluid resorption. Endocrinol 1998; 139:3935-45. [DOI] [PubMed] [Google Scholar]
- 184. Pastor-Soler NM, Fisher JS, Sharpe R, Hill E, Van Hoek A, Brown D, Breton S. Aquaporin 9 expression in the developing rat epididymis is modulated by steroid hormones. Reproduction 2010; 139:613-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185. McKinnell C, Atanassova N, Williams K, Fisher JS, Walker M, Turner KJ, Saunders TK, Sharpe RM. Suppression of androgen action and the induction of gross abnormalities of the reproductive tract in male rats treated neonatally with diethylstilbestrol. J Androl 2001; 22:323-38. [PubMed] [Google Scholar]
- 186. Naito M, Hirai S, Terayama H, Qu N, Hayashi S, Hatayama N, Kawamura H, Nakano T, Itoh M. Neonatal estrogen treatment with beta-estradiol 17-cypionate induces in post-pubertal mice inflammation in the ductuli efferentes, epididymis, and vas deferens, but not in the testis, provoking obstructive azoospermia. Med Mol Morphol 2013. [DOI] [PubMed] [Google Scholar]
- 187. Yamashita R, Oshima A, Hasegawa-Baba Y, Wada M, Shibuya K. Endocrine disrupting effects of low dose 17 beta-estradiol (E(2)) on the Japanese quail (Coturnix japonica) were detected by modified one-generation reproduction study. J Toxicol Sci 2011; 36:43-54. [DOI] [PubMed] [Google Scholar]
- 188. Aceitero J, Llanero M, Parrado R, Pena E, Lopez-Beltran A. Neonatal exposure of male rats to estradiol benzoate causes rete testis dilation and backflow impairment of spermatogenesis. Anat Rec 1998; 252:17-33. [DOI] [PubMed] [Google Scholar]
- 189. Hahn KL, Beres B, Rowton MJ, Skinner MK, Chang Y, Rawls A, Wilson-Rawls J. A deficiency of lunatic fringe is associated with cystic dilation of the rete testis. Reproduction 2009; 137:79-93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190. Lupien M, Dievart A, Morales CR, Hermo L, Calvo E, Kay DG, Hu C, Jolicoeur P. Expression of constitutively active Notch1 in male genital tracts results in ectopic growth and blockage of efferent ducts, epididymal hyperplasia and sterility. Dev Biol 2006. [DOI] [PubMed] [Google Scholar]
- 191. Nie X, Arend LJ. Pkd1 is required for male reproductive tract development. Mech Dev 2013; 130:567-76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192. Ikadai H, Fujita H, Ajisawa C, Imamichi T. Sperm retention cysts in the efferent duct and testicular atrophy of TE inbred rats. J Reprod Fertil 1987; 81:635-41. [DOI] [PubMed] [Google Scholar]
- 193. Jeffs B, Meeks JJ, Ito M, Martinson FA, Matzuk MM, Jameson JL, Russell LD. Blockage of the rete testis and efferent ductules by ectopic sertoli and leydig cells causes infertility in dax1-deficient male mice. Endocrinology 2001; 142:4486-95. [DOI] [PubMed] [Google Scholar]
- 194. O’Hara L, Welsh M, Saunders PT, Smith LB. Androgen receptor expression in the caput epididymal epithelium is essential for development of the initial segment and epididymal spermatozoa transit. Endocrinology 2011; 152:718-29. [DOI] [PubMed] [Google Scholar]
- 195. Krutskikh A, De Gendt K, Sharp V, Verhoeven G, Poutanen M, Huhtaniemi I. Targeted inactivation of the androgen receptor gene in murine proximal epididymis causes epithelial hypotrophy and obstructive azoospermia. Endocrinology 2011; 152:689-96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196. Bjorkgren I, Saastamoinen L, Krutskikh A, Huhtaniemi I, Poutanen M, Sipila P. Dicer1 ablation in the mouse epididymis causes dedifferentiation of the epithelium and imbalance in sex steroid signaling. PLoS ONE 2012; 7:e38457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197. Choyke PL, Glenn GM, Wagner JP, Lubensky IA, Thakore K, Zbar B, Linehan WM, Walther MM. Epididymal cystadenomas in von Hippel-Lindau disease. Urology 1997; 49:926-31. [DOI] [PubMed] [Google Scholar]
- 198. Pozza D, Masci P, Amodeo S, Marchionni L. Papillary cystadenoma of the epididymis as a cause of obstructive azoospermia. Urol Int 1994; 53:222-4. [DOI] [PubMed] [Google Scholar]
- 199. Jones MA, Young RH, Scully RE. Adenocarcinoma of the epididymis: a report of four cases and review of the literature. Am J Surg Pathol 1997; 21:1474-80. [DOI] [PubMed] [Google Scholar]
- 200. Jaakkola P, Mole DR, Tian YM, Wilson MI, Gielbert J, Gaskell SJ, von Kriegsheim A, Hebestreit HF, Mukherji M, Schofield CJ, et al. . Targeting of HIF-alpha to the von Hippel-Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science 2001; 292:468-72. [DOI] [PubMed] [Google Scholar]
- 201. Leung SY, Chan AS, Wong MP, Yuen ST, Fan YW, Chung LP. Expression of vascular endothelial growth factor in von Hippel-Lindau syndrome-associated papillary cystadenoma of the epididymis. Hum Pathol 1998; 29:1322-4. [DOI] [PubMed] [Google Scholar]
- 202. Hendry WF, A’Hern RP, Cole PJ. Was Young's syndrome caused by exposure to mercury in childhood? Bmj 1993; 307:1579-82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203. Hendry WF, Levison DA, Parkinson MC, Parslow JM, Royle MG. Testicular obstruction: clinicopathological studies. Ann R Coll Surg Engl 1990; 72:396-407. [PMC free article] [PubMed] [Google Scholar]
- 204. Nistal M, Riestra ML, Galmes-Belmonte I, Paniagua R. Testicular biopsy in patients with obstructive azoospermia. Am J Surg Pathol 1999; 23:1546-54. [DOI] [PubMed] [Google Scholar]
- 205. Handelsman DJ, Conway AJ, Boylan LM, Turtle JR. Young's syndrome. Obstructive azoospermia and chronic sinopulmonary infections. N Engl J Med 1984; 310:3-9. [DOI] [PubMed] [Google Scholar]
- 206. Nistal M, Paniagua R, Regadera J, Santamaria L. Obstruction of the tubuli recti and ductuli efferentes by dilated veins in the testes of men with varicocele and its possible role in causing atrophy of the seminiferous tubules. Int J Androl 1984; 7:309-23. [DOI] [PubMed] [Google Scholar]
- 207. Ball RY, Mitchinson MJ. Obstructive lesions of the genital tract in men. J Reprod Fertil 1984; 70:667-73. [DOI] [PubMed] [Google Scholar]
- 208. Eigenmann J, Bandhauer K, Tomamichel G. Seminal carnitine concentration in obstructive azoospermia. Eur Urol 1994; 26:134-6. [DOI] [PubMed] [Google Scholar]
- 209. Nistal M, Mate A, Paniagua R. Granulomatous epididymal lesion of possible ischemic origin. Am J Surg Pathol 1997; 21:951-6. [DOI] [PubMed] [Google Scholar]
- 210. Robson WL, Thomason MA, Minette LJ. Cystic dysplasia of the testis associated with multicystic dysplasia of the kidney. Urology 1998; 51:477-9. [DOI] [PubMed] [Google Scholar]
- 211. Nistal M, Gonzalez-Peramato P, Sousa G, Garcia-Cabezas MA, Rodriguez JI, Cajaiba MM. Cystic dysplasia of the epididymis: a disorder of mesonephric differentiation associated with renal maldevelopment. Virchows Arch 2010; 456:695-702. [DOI] [PubMed] [Google Scholar]
- 212. Nistal M, Santamaria L, Paniagua R. Acquired cystic transformation of the rete testis secondary to renal failure. Hum Pathol 1989; 20:1065-70. [DOI] [PubMed] [Google Scholar]
- 213. Belet U, Danaci M, Sarikaya S, Odabas F, Utas C, Tokgoz B, Sezer T, Turgut T, Erdogan N, Akpolat T. Prevalence of epididymal, seminal vesicle, prostate, and testicular cysts in autosomal dominant polycystic kidney disease. Urology 2002; 60:138-41. [DOI] [PubMed] [Google Scholar]
- 214. Nistal M, Jimenez-Heffernan JA, Garcia-Viera M, Paniagua R. Cystic transformation and calcium oxalate deposits in rete testis and efferent ducts in dialysis patients. Hum Pathol 1996; 27:336-41. [DOI] [PubMed] [Google Scholar]
- 215. Guvel S, Pourbagher MA, Torun D, Egilmez T, Pourbagher A, Ozkardes H. Calcification of the epididymis and the tunica albuginea of the corpora cavernosa in patients on maintenance hemodialysis. J Androl 2004; 25:752-6. [DOI] [PubMed] [Google Scholar]
- 216. Bozzini G, Lunelli L, Berlingheri M, Groppali E, Carmignani L. Epididymis microlithiasis and semen abnormalities in young adult kidney transplant recipients. Andrologia 2012. [DOI] [PubMed] [Google Scholar]
- 217. Barack BM. Transport of spermatozoa from seminiferous tubules to epididymis in the mouse: a histological and quantitative study. J Reprod Fertil 1968; 16:35-48. [DOI] [PubMed] [Google Scholar]
- 218. Harrison RG, Macmillan EW. The effects of high epididymal obstruction upon the Leyding cell volume of the rat testis. J Endocrinol 1954; 11:89-96. [DOI] [PubMed] [Google Scholar]
- 219. Van Wagenen G. Changes in the testis of the rat following ligation of the ductuli efferentis. Anat Rec 1925; 29:399. [Google Scholar]
- 220. White WE. The duration of fertility and the histological changes in the reproductive organs after ligation of the vas efferentia in the rat. Proc Royal Soc (London) B 1933; 133:544-50. [Google Scholar]
- 221. Smith G. The effects of ligation of the vasa efferentia and vasectomy on testicular function in the adult rat. J Endocrinol 1962; 23:285-99. [DOI] [PubMed] [Google Scholar]
- 222. Le Lannou D, Chambon Y. [Effects of ligation of the ductuli efferentes or the epididymis on testicular function and secretion of gonadotropins in rats]. C R Seances Soc Biol Fil 1977; 171:197-200. [PubMed] [Google Scholar]
- 223. Nykänen M, Kormano M. Early Effects of Efferent Duct Ligation on the Rat Rete Testis. Int J Androl 1978; 1:225-34. [Google Scholar]
- 224. Soler C, Blazquez C, Pertusa J, Nunez M, Nunez J, Nunez A. A comparison of the effects of bilateral efferent duct ligation and of partial epididymectomy on the testes of rats. Reprod Fertil Dev 1990; 2:321-6. [DOI] [PubMed] [Google Scholar]
- 225. Young WC. Die resorption in den ductuli efferentes der maus und ihre bedeutung fur das problem der unterbindung im hoden-nebenhodensystem. Z Zellforsch Mikrosk anat Abt Histochem 1933; 17:727-59. [Google Scholar]
- 226. Macmillan EW. The immediate effects of occlusion of superior epididymal arteries in the rat as demonstrated by the use of radiopaque medium. Proc Soc Study Fertil 1956; 8:67-75. [PubMed] [Google Scholar]
- 227. Markey CM, Jequier AM, Meyer GT. Effects of ischaemia on the caput epididymis and its relationship to higher epididymal obstruction: a qualitative study in the ram. Int J Androl 1995; 18:185-96. [DOI] [PubMed] [Google Scholar]
- 228. Sabolic I, Ljubojevic M, Herak-Kramberger CM, Brown D. Cd-MT causes endocytosis of brush-border transporters in rat renal proximal tubules. Am J Physiol Renal Physiol 2002; 283:F1389-402. [DOI] [PubMed] [Google Scholar]
- 229. Morris RG, Tousson A, Benos DJ, Schafer JA. Microtubule disruption inhibits AVT-stimulated Cl- secretion but not Na+ reabsorption in A6 cells [In Process Citation]. Am J Physiol 1998; 274:F300-14. [DOI] [PubMed] [Google Scholar]
- 230. Mirela B, Mila MP, Brown D, Ivan S. In colchicine-treated rats, cellular distribution of AQP-1 in convoluted and straight proximal tubule segments is differently affected. Pflugers Arch 2000; 439:321-30. [PubMed] [Google Scholar]
- 231. Rogers SL, Gelfand VI. Membrane trafficking, organelle transport, and the cytoskeleton. Curr Opin Cell Biol 2000; 12:57-62. [DOI] [PubMed] [Google Scholar]
- 232. Romano G, Favret G, Catone B, Bartoli E. The Effect of Colchicine On Proximal Tubular Reabsorption. Pharmacol Res 2000; 41:305-11. [DOI] [PubMed] [Google Scholar]
- 233. Chalumeau C, du Cheyron D, Defontaine N, Kellermann O, Paillard M, Poggioli J. NHE3 activity and trafficking depend on the state of actin organization in proximal tubule. Am J Physiol Renal Physiol 2001; 280:F283-90. [DOI] [PubMed] [Google Scholar]
- 234. Li XC, Hopfer U, Zhuo JL. AT1 receptor-mediated uptake of angiotensin II and NHE-3 expression in proximal tubule cells through a microtubule-dependent endocytic pathway. Am J Physiol Renal Physiol 2009; 297:F1342-52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235. Lapierre LA, Ducharme NA, Drake KR, Goldenring JR, Kenworthy AK. Coordinated regulation of caveolin-1 and Rab11a in apical recycling compartments of polarized epithelial cells. Exp Cell Res 2012; 318:103-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236. Tung KSK, Primakoff P, Woolman-Gamer L, Myles DG. Mechanism of Infertility in Male Guinea Pigs Immunized with Sperm PH-20. Biology of Reproduction 1997; 56:1133-41. [DOI] [PubMed] [Google Scholar]
- 237. Collin O, Damber JE, Bergh A. Effects of endothelin-1 on the rat testicular vasculature. J Androl 1996; 17:360-6. [PubMed] [Google Scholar]
- 238. Povysil C. Spermatic granulomas of ischemic origin in the rat epididymis. Invest Urol 1976; 14:219-24. [PubMed] [Google Scholar]
- 239. Billups KL, Tillman SL, Chang TS. Reduction of epididymal sperm motility after ablation of the inferior mesenteric plexus in the rat. Fertil Steril 1990; 53:1076-82. [PubMed] [Google Scholar]
- 240. Billups KL, Tillman S, Chang TS. Ablation of the inferior mesenteric plexus in the rat: alteration of sperm storage in the epididymis and vas deferens. J Urol 1990; 143:625-9. [DOI] [PubMed] [Google Scholar]
- 241. Aruldhas MM, Subramanian S, Sekhar P, Hasan GC, Govindarajulu P, Akbarsha MA. Microcanalization in the epididymis to overcome ductal obstruction caused by chronic exposure to chromium - a study in the mature bonnet monkey (Macaca radiata Geoffroy). Reproduction 2004; 128:127-37. [DOI] [PubMed] [Google Scholar]
- 242. Labrecque M, Hays M, Chen-Mok M, Barone MA, Sokal D. Frequency and patterns of early recanalization after vasectomy. BMC Urol 2006; 6:25. [DOI] [PMC free article] [PubMed] [Google Scholar]