

Abstract
Diabetes mellitus (DM) is a major worldwide health problem and its prevalence has been rapidly increasing in the last century. It is caused by defects in insulin secretion or insulin action or both, leading to hyperglycemia. Of the various types of DM, type 2 occurs most frequently. Multiple genes and their interactions are involved in the insulin secretion pathway. Insulin secretion is mediated through the ATP-sensitive potassium (KATP) channel in pancreatic beta cells. This channel is a heteromeric protein, composed of four inward-rectifier potassium ion channel (Kir6.2) tetramers, which form the pore of the KATP channel, as well as sulfonylurea receptor 1 subunits surrounding the pore. Kir6.2 is encoded by the potassium inwardly rectifying channel, subfamily J, member 11 (KCNJ11) gene, a member of the potassium channel genes. Numerous studies have reported the involvement of single nucleotide polymorphisms of the KCNJ11 gene and their interactions in the susceptibility to DM. This review discusses the current evidence for the contribution of common KCNJ11 genetic variants to the development of DM. Future studies should concentrate on understanding the exact role played by these risk variants in the development of DM.
1. Introduction
Diabetes mellitus (DM) is a chronic disease characterized by high blood glucose levels caused by either insufficient insulin production by the pancreas or improper response of the body cells to insulin [1]. Approximately 366 million people were diagnosed with DM worldwide in 2011 and this is expected to increase to 552 million by 2030 [2]. This disease has early- and late-stage complications. Early complications include hyperglycemia, polyphagia, polydipsia, polyuria, and blurred vision, leading to complications manifested later such as vascular disease, heart disease, stroke, peripheral neuropathy, nephropathy, and predisposition to infection [3].
DM is classified into various types, of which type 2 (T2DM) occurs most frequently. Approximately 5%–10% of patients with DM are affected by type 1 (T1DM) and more than 90% by T2DM. T1DM (formerly insulin-dependent diabetes, or juvenile diabetes) results from the autoimmune destruction of the insulin-producing beta cells in the pancreas [4]. T2DM (formerly noninsulin-dependent DM) is a metabolic disorder due to hyperglycemia in the context of insulin resistance and relative lack of insulin. This is in contrast to T1DM, in which there is an absolute lack of insulin due to the breakdown of islet cells in the pancreas [5]. Gestational DM (GDM) affects 3%–10% of pregnancies in various populations. In this disease, insulin receptors do not function properly, which results in high blood glucose levels during pregnancy with subsequent negative effects, such as the baby being large for gestational age, being born stillborn, or dying shortly after birth or the mother having a miscarriage or problems with her eyes and kidneys [6, 7].
DM is a multifactorial disease caused by both genetic and environmental factors and their complex interaction. Environmental risk factors include obesity, physical inactivity, hypertension, abnormal cholesterol levels, age, and smoking [8]. Pathogenic and nonpathogenic mutations in genomic DNA can also contribute to the development of diabetes. Several single nucleotide polymorphisms (SNPs), which are a type of nonpathogenic mutation, have been reported to be associated with susceptibility to different types of DM. The aim of this review is to assess the possible contribution of SNPs of the gene known as the potassium inwardly rectifying channel, subfamily J, member 11 (KCNJ11), which forms a compartment of the ATP-sensitive potassium (KATP) channel, present in beta cells of the islets, in the susceptibility to different types of DM.
2. Role of Genetics in the Development of DM
People with a family history of T1DM and T2DM are six and three times more likely, respectively, to develop these diseases than are unrelated individuals [9]. Multiple genes are involved in DM. Those that have garnered the most attention are the ATP-binding cassette transporter subfamily C member 8 (ABCC8) gene; the KCNJ11 gene; and the peroxisome proliferator-activated receptor-gamma (PPARG) gene. Most of these genes are involved in insulin action/glucose metabolism, pancreatic beta cell function or other metabolic conditions (e.g., energy intake/expenditure, lipid metabolism) [10]. Mutations in genes such as ABCC8 and KCNJ11 can disrupt the potentiation activity of the KATP channel and have thus been associated with permanent neonatal DM [11]. The PPARG gene is implicated in adipogenesis and the development of insulin resistance. Deleterious mutations in this gene impair insulin resistance and can cause lack of response to insulin [12].
From recent genome-wide association studies, more than 60, 500, and 65 loci have been identified for susceptibility to T1DM, T2DM, and GDM, respectively [13]. SNPs are the most common type of genetic variation distributed within or outside a gene region in the human genome. The frequency of SNPs is less than 1% in the genome, and approximately 54% of these variants are not deleterious [14]. SNPs can modify the risk of occurrence of a disease, either alone or in linkage disequilibrium in one gene or in neighborhood genes. For instance, in several studies, the common Pro12Ala polymorphism in the PPARG gene, the Glu23Lys polymorphism in the KCNJ11 gene, or the Ser1369Ala polymorphism in the ABCC8 gene was confirmed to be associated with DM [15].
3. KCNJ11 Gene and Its Product
The KCNJ11 gene, a member of the potassium channel gene family, is located at 11p15.1 and has no intron (Figure 1: 17,385,237–17,389,331). This gene encodes an inward-rectifier potassium ion channel (Kir6.2). The Kir6.2 protein, together with the high-affinity sulfonylurea receptor 1 (SUR1), forms the KATP channel. SUR1 is encoded by the ABCC8 gene located next to the KCNJ11 gene. The Kir6.2 protein is a 390-amino acid protein with two transmembrane domains (M1 and M2) and intracellular N- and C-terminals. Structurally, Kir6.2 tetramers form the pore and four high-affinity SUR1 subunits surround the pore of the KATP channel located at the plasma membrane of pancreatic beta cells. This channel modulates insulin production and secretion through glucose metabolism [16].
Figure 1.

ABCC8 and KNJ11 genes and their encoded proteins and functions. The KCNJ11 and ABCC8 genes are next to each other on chromosome 11p15.1. KCNJ11 is a single exon (dark box) gene encoding the Kir6.2 protein, and ABCC8 has 35 exons (dark boxes) encoding the SUR1 protein; both are subunits of the ATP-sensitive potassium (KATP) channel. Metabolism of glucose can affect ATP levels and thereby the function of the KATP channel. ABCC8: ATP-binding cassette transporter subfamily C member 8; KCNJ11: potassium inwardly-rectifying channel, subfamily J, member 11; Kir6.2: inward-rectifier potassium ion channel; SUR1: sulfonylurea receptor 1; NBD1: nucleotide-binding domain 1; NBD2: nucleotide-binding domain 2; N: NH2 terminal of protein; C: COOH terminal of protein; A: Walker A motif; B: Walker B motif; cAMP: cyclic adenosine monophosphate; ATP: adenosine triphosphate.
3.1. Role of Kir6.2 in Insulin Secretion
The Kir6.2 protein, coupled with the SUR1 protein in the KATP channel, mediates insulin secretion. This channel is involved in a wide range of physiological responses. Increased glucose induces higher potassium flow into the cell through the KATP channel. ADP in the presence of magnesium (Mg) converts to ATP; the ATP then closes the KATP channel by binding to Kir6.2, increasing the intracellular potassium ion concentration, which depolarizes the cell membrane and subsequently activates calcium ion (Ca2+) channel. Ca2+ is a ubiquitous intracellular second messenger that is critical for cellular functioning. These calcium channels influence the voltage-dependent potassium channels to repolarize the cell membrane, leading to closure of the voltage-dependent calcium channels. Increased intracellular free Ca2+ levels trigger other components of the insulin secretion pathway to release granules at or near the plasma membrane (Figure 2). Mutations in the KCNJ11 gene can cause DM because of the reduced ability of ATP to inhibit the activity of the KATP channel and the enhanced ability of MgATP to simultaneously stimulate the function of this channel. This is associated with defective insulin secretion, ultimately causing DM [17].
Figure 2.
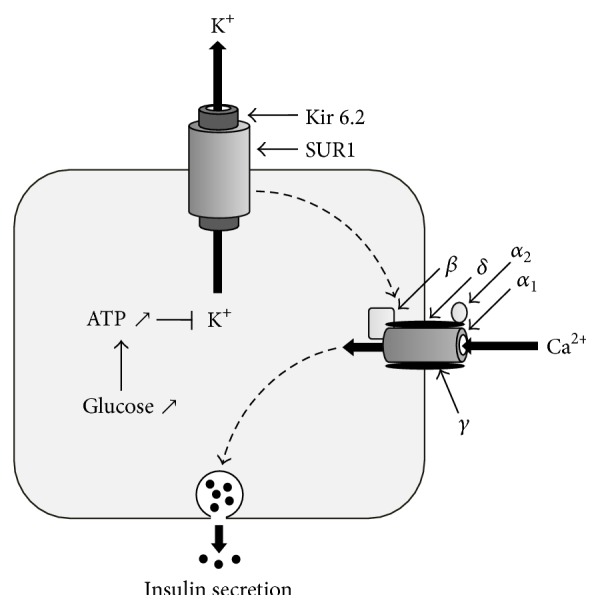
Mechanism of insulin secretion by the KATP channel in pancreatic beta cells. The Kir6.2 and SUR1 proteins in the KATP channel mediate insulin secretion. An increase in glucose levels stimulates the KATP channel to allow the entry of potassium ions. An increase in intracellular potassium ions depolarizes the cell membrane and induces calcium channels to increase intracellular free Ca2+ levels. The calcium ions trigger other components of the insulin secretion pathway to release granules at or near the plasma membrane. KATP: ATP-sensitive potassium channel; Kir6.2: inward-rectifier potassium ion channel; SUR1: sulfonylurea receptor 1; ATP: adenosine triphosphate; K+: potassium ion; Ca2+: calcium ion. The calcium channel is composed of α 1, α 2, β, γ, and δ subunits.
3.2. KCNJ11 Common Polymorphisms Involved in Diabetes
KCNJ11 has 219 SNPs, six of which have been receiving more attention for their association with diabetes. Among these six common SNPs, three are located in the coding regions and three in the noncoding regions (Table 1). These six SNPs include rs5219, rs5215, rs5210, rs5218, rs886288, and rs2285676.
Table 1.
Characteristics of KCNJ11 gene variants in association with diabetes mellitus.
| Number | SNP | Location | MAF | Allele | Amino acid | Diabetes | Association | References | |
|---|---|---|---|---|---|---|---|---|---|
| Chromosome | Gene | ||||||||
| 1 | rs2285676 | 17386478 | 3′ UTR | 0.46 | T>C | — | T2DM | Yes | [30] |
|
| |||||||||
| 2 | rs5210 | 17386704 | 3′ UTR | 0.46 | G>A | — | T2DM | Yes | [30, 36, 76] |
| No | [72, 74] | ||||||||
|
| |||||||||
| 3 | rs5215 | 17387083 | Exon | 0.28 | G>A | Val250Ile | T2DM | Yes | [27, 30, 35, 36] |
| No | [49, 50, 69–72] | ||||||||
| T1DM | No | [73, 74] | |||||||
| GDM | No | [68] | |||||||
|
| |||||||||
| 4 | rs5218 | 17387522 | Exon | 0.27 | C>T | Ala103Ala | T2DM | No | [36] |
|
| |||||||||
| 5 | rs5219 | 17388025 | Exon | 0.27 | G>A | Lys23Gln | T1DM | No | [63, 64] |
| T2DM | Yes | [19–43] | |||||||
| No | [44–64] | ||||||||
| GDM | Yes | [38] | |||||||
| No | [66–68] | ||||||||
|
| |||||||||
| 6 | rs886288 | 17389616 | 5′ near gene | 0.46 | T>C | — | T2DM | Yes | [36] |
MAF: minor allele frequency; T1DM: type 1 diabetes mellitus; T2DM: type 2 diabetes mellitus; GDM: gestational diabetes mellitus; SNP: single nucleotide polymorphism; UTR: untranslated region.
3.2.1. KCNJ11 rs5219
This locus is located in exon 1 of the KCNJ11 gene. Substitution of A to C (AAG→CAG) changes the amino acid from lysine to glutamine (Lys23Gln) at the NH2-terminal tail of Kir6.2. Lysine has a positively charged epsilon-amino group, whereas glutamine is uncharged under all biological conditions. Despite this amino acid substitution, theoretically, it does not make a remarkable change in the structure and function of the KCNJ11 protein [18]. Studies have shown, however, that the rs5219 variant may alter the charge of the ATP-binding region and decrease channel sensitivity to ATP. Twenty-four association studies and a recent meta-analysis showed a strong relationship between the rs5219 polymorphism and susceptibility to T2DM [19–43], whereas 21 studies did not confirm this finding [44–64]. This meta-analysis showed that the rs5219 polymorphism is a risk factor for developing T2DM in Caucasians and in some Asian populations. Populations from East Asia were more prone to this disease, where the A allele frequency in most patients was more common than in controls. Therefore, genetic background can affect susceptibility to T2DM [65].
The rs5219 polymorphism can affect the insulin secretion pathway. The A allele of this locus impairs this pathway by reducing ATP sensitivity of the KATP channel, hence resulting in overactivity of the channel and subsequent suppression of insulin secretion. This effect on insulin secretion is more significant in carriers of the AA genotype compared with carriers of the GA genotype [30]. Similar results were observed for fasting plasma glucose and postprandial plasma glucose levels in patients with T2DM. The A allele increased the fasting plasma glucose and postprandial plasma glucose levels in these patients, whereas GA carriers had higher 2 h postprandial plasma glucose levels than did GG carriers with T2DM [32, 38]. This allele was also associated with reduction in serum insulin levels in a postoral glucose tolerance test [39]. In contrast to one study from Scandinavia on GDM risk [37], the remaining studies did not report any association between this locus and T1DM and GDM [66–68].
Hypertension is a main complication of T2DM. The rs5219 polymorphism plays a strong role in HbA1c and blood pressure levels in this disease. The A allele carriers of rs5219 had higher HbA1c levels and blood pressure than did the G allele carriers [33, 35, 36, 40, 41]. In T2DM, a relationship has been suggested between the A allele and increased hepatitis insulin sensitivity [32]. Pharmacogenomics studies demonstrated that A allele carriers of the rs5219 polymorphism who have T2DM have better therapeutic response to gliclazide than do G allele carriers. In the A allele group, HbA1c was also reduced more in patients taking glimepiride and glibenclamide than it was in patients taking gliclazide treatment [40]. The ring-fused pyrrole moiety in these two drugs binds to the A allele, underling the inhibitory potency of these drugs on KATP channels [41]. The rs5219 polymorphism also plays a role in determining the efficacy of repaglinide [33, 39]. Carriers of the C allele were also found to have a reduced response to sulfonylurea therapy [42, 43].
3.2.2. KCNJ11 rs5215
The rs5215 polymorphism is located in exon 1 of the KCNJ11 gene. It is a nonsynonymous variant caused by a substitution of G to A (GTC→ATC), which changes the amino acid from valine to isoleucine at residue 250. Valine is hydrophobic, whereas isoleucine is one of three amino acids having branched hydrocarbon side chains. Isoleucine is usually interchangeable with leucine and occasionally with valine in proteins. Of 13 studies on DM, 3 showed strong associations between this variant and T2DM [27, 30, 35], whereas the remaining studies showed no association with T2DM, T1DM, or GDM [36, 49–51, 69–74]. In another study, the rs5215 polymorphism was associated with blood pressure among subjects with T2DM [36].
3.2.3. KCNJ11 rs5210
The rs5210 polymorphism is located at a highly conserved 3′ untranslated region (UTR) of the KCNJ11 gene. Of four reports relevant to susceptibility to T2DM, two identified a plausible role in development of this disease, whereas the other studies did not confirm this relationship [30, 36, 72, 75]. A study found that this variant improves the clinical efficacy of gliclazide in patients with T2DM [76]. This locus is a target of miR-1910; however, the mechanism of action of this miRNA in the development of DM is unknown. MiRNAs encompass 17 to 25 nucleotides, which posttranscriptionally regulate the expression of thousands of genes in a broad range of organisms in both normal physiological and disease contexts. Appropriate secretion of insulin from pancreatic beta cells is a vital factor in blood glucose homeostasis, and miRNAs have been identified as being involved in the regulation of insulin exocytosis. MiRNAs control insulin synthesis and release it in beta cells. The G allele is a potential target for miR-1910, whereas the A allele abolishes binding of this miRNA to this region [77, 78]. Further studies may reveal the role of miR-1910 in DM.
3.2.4. KCNJ11 rs5218
The rs5218 polymorphism is located in the 3′-UTR of the KCNJ11 gene. It is a synonymous variant with a substitution of G to A (GCC→GCT), which encodes, for alanine at residue 103, a hydrophobic and ambivalent amino acid. There is only one report of this locus in DM, which showed no association with T2DM risk [36].
3.2.5. KCNJ11 rs886288 and rs2285676
The rs886288 polymorphism is located in the 5′ flank near the gene, whereas the rs2285676 polymorphism is located at the 3′-UTR region. Two studies revealed an association of the rs886288 and rs2285676 polymorphisms with T2DM [30, 36].
3.3. Interaction of the KCNJ11 Gene with Other Genes
Insulin secretion from pancreatic beta cells can be modulated by a complex cluster of proteins encoded by related genes, including KCNJ11, ABCC8, voltage-sensitive calcium channels (VSCCs), ABCC9, protein kinase catalytic subunit G (PRKACG), rap guanine nucleotide exchange factor 4 (RAPGEF4), forkhead box A2 (FOXA2), and endosulfine alpha (ENSA). These proteins act at the cell membrane or intracellular level (Figure 3).
Figure 3.
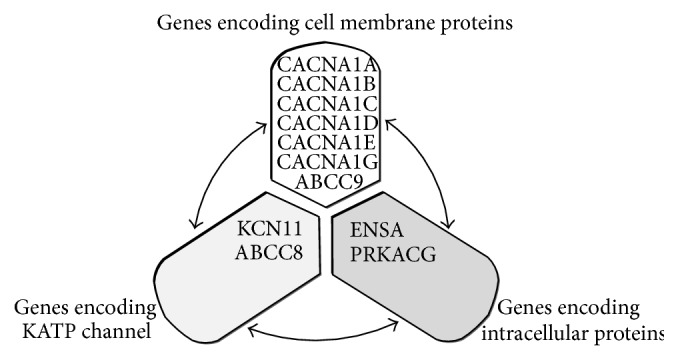
Cross-talk between the KCNJ11 gene and the other genes involved in the regulation of insulin secretion in pancreatic beta cells. KCNJ11 interacts with many genes, of which 10 are most prominent. KCNJ11 interacts with ABCC8 to produce the KATP channel, which transfers potassium ions across the beta cells. The KCNJ11 and ABCC8 genes interact with three groups of gene products at the cell membrane (white) and the intracellular (dark gray) levels. KATP: ATP-sensitive potassium channel; CACNA1A: calcium channel, voltage-dependent, P/Q type, alpha 1A subunit; CACNA1B: calcium channel, voltage-dependent, N type, alpha 1B subunit; CACNA1C: calcium channel, voltage-dependent, L type, alpha 1C subunit; CACNA1D; calcium channel, voltage-dependent, L type, alpha 1D subunit; CACNA1E: calcium channel, voltage-dependent, R type, alpha 1E subunit; CACNA1G: calcium channel, voltage-dependent, T type, alpha 1G subunit; ABCC9: ATP-binding cassette transporter subfamily C member 9; KCNJ11: potassium inwardly rectifying channel, subfamily J, member 11; ABCC8: ATP-binding cassette transporter subfamily C member 8; ENSA: endosulfine alpha; PRKACG; protein kinase catalytic subunit G; RAPGEF4: rap guanine nucleotide exchange factor 4; FOXA2: forkhead box A2.
3.3.1. Interactions at the Cell Membrane Level
KCNJ11 and ABCC8 genes encode Kir6.2 and Sur1, respectively, in pancreatic beta cells. Both proteins form compartments in the KATP channels, which allow potassium to flow into the cell rather than out of it, as mediated by G proteins [17]. The KATP channel interacts with different types of VSCCs, including L (long-lasting), N (neural), P/Q (purkinje), R (residual), and T (transient). Calcium channels are generally composed of four subunits: α 1, α 2-δ, β, and γ. The function of the calcium channel is controlled by the pore-forming α 1 subunit, which blocks the entry of calcium ions into the excitable cells, and by the auxiliary subunits, which modulate trafficking and the biophysical properties of the α 1 subunit. The α 1 subunit isoforms include A, B, C, D, E, and G, encoded by CACNA1A, CACNA1B, CACNA1C, CACNA1D, CACNA1E, and CACNA1G genes, respectively. The A to E forms of the α 1 subunit produce various types of calcium channels, including P/Q, N, L, L, R, and T, respectively. The L, N, P/Q, and R types of these channels belong to the high-voltage activated (HVA) group and the T type belongs to the low-voltage activated (LVA) group. Both HVA and LVA groups are involved in calcium-dependent processes such as neurotransmitter or hormone release, muscle contraction, cell motility, gene expression, cell division, and cell death [78, 79]. Finally, Kir2 and ABCC9 can form another type of KATP channel in cardiac, skeletal, vascular, and nonvascular smooth muscle. The structure of the ABCC9 protein suggests a role as a drug-binding, channel-modulating subunit of the extrapancreatic KATP channels [80].
3.3.2. Interactions at the Intracellular Level
The KATP channels interact with the PRKACG protein encoded by the PRKACG gene. This protein is the gamma catalytic subunit of protein kinase, which is involved in exocytosis through different pathways such as calcium- and hormone-mediated signaling. This protein also activates cellular processes such as intracellular protein kinase A [81]. Kir6.2 interacts with RAPGEF4, FOXA2, and ENSA proteins, encoded by RAPGEF4, FOXA2, ENSA, and ABCC9 genes, respectively. RAPGEF4 is an exchange protein that can be activated by cAMP. FOXA2 functions as a transcription activator for genes such as alpha-fetoprotein, albumin, and tyrosine aminotransferase. ENSA is an endogenous ligand for SUR1, which stimulates insulin secretion [82–84]. Defects in the KCNJ11 gene may also lead to autosomal-dominant T2DM, transient neonatal DM type 3, and permanent neonatal DM [85].
4. Concluding Remarks
DM is one of the most common diseases globally, with high social and economic burdens. Kir6.2 plays a potential role in the function of the KATP channel. Some active mutations in this gene can disrupt Kir6.2 activity and consequently reduce the potential of the KATP channel, leading to DM. It is evident from the literature that several variants of the KCNJ11 gene are associated with different types of DM. This raises the question of which polymorphisms of the KCNJ11 gene and their combinations play more prominent roles in the development of DM.
Most previous studies have focused on six common polymorphisms in DM: rs5210, rs5215, rs5218, rs5219, rs886288, and rs2285676. Of these six loci, rs5219, rs5215, and rs5210 have been given most attention. No evidence yet exists in the literature for the involvement of other SNPs of the KCNJ11 gene. The rs5219 A allele plays an important role in insulin secretion through reduction of ATP sensitivity of the KATP channel and suppression of insulin secretion. However, the mechanism of action of this locus in the insulin secretion pathway is still not completely understood. The rs5210 G allele acts as a potential target for miR-1910, which is implicated in T2DM; however, the mechanism of action of this miRNA in the development of DM is unknown. MiRNAs control insulin synthesis and release from beta cells. Future studies are suggested to reveal the use of miR-1910 as a potential biomarker in the diagnosis of diabetes and its plausible application for treatment of DM.
Regulation of insulin release is mediated by KCNJ11 in concert with different genes such as ABCC8, ABCC9, and CACNA1A-G. Diminished coexpression of these genes may increase the risk of DM. Nevertheless, the exact functional relationship of the network of these genes in the regulation of insulin release remains to be determined. Future studies are suggested to discover the exact role of KCNj11 gene variants and their interaction with other genes in DM for the possible development of suitable therapies and the diagnosis of this common disease.
Acknowledgments
This study was supported by Malaysian Grants HIR MOHE E000025-20001, UM.C/625/1/HIR/161, RG 520/13HTM, and RG056/09HTM.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Wilkin T. J. The accelerator hypothesis: a review of the evidence for insulin resistance as the basis for type I as well as type II diabetes. International Journal of Obesity. 2009;33(7):716–726. doi: 10.1038/ijo.2009.97. [DOI] [PubMed] [Google Scholar]
- 2.Gardner D. G., Shoback D. M., editors. Greenspan's Basic & Clinical Endocrinology. New York, NY, USA: McGraw-Hill Medical; 2007. [Google Scholar]
- 3.Zimmet P. Z., Magliano D. J., Herman W. H., Shaw J. E. Diabetes: a 21st century challenge. The Lancet Diabetes and Endocrinology. 2014;2(1):56–64. doi: 10.1016/s2213-85871370112-8. [DOI] [PubMed] [Google Scholar]
- 4.Kumar V., Abbas A. K., Fausto N., Aster J. C. Robbins and Cotran Pathologic Basis of Disease. Elsevier Health Sciences; 2009. (Professional Edition: Expert Consult-Online). [Google Scholar]
- 5.Cooke D. W., Plotnick L. Type 1 diabetes mellitus in pediatrics. Pediatrics in Review. 2008;29(11):374–385. doi: 10.1542/pir.29-11-374. [DOI] [PubMed] [Google Scholar]
- 6.Ryan E. A., Enns L. Role of gestational hormones in the induction of insulin resistance. The Journal of Clinical Endocrinology & Metabolism. 1988;67(2):341–347. doi: 10.1210/jcem-67-2-341. [DOI] [PubMed] [Google Scholar]
- 7.Prudente S., Dallapiccola B., Pellegrini F., Doria A., Trischitta V. Genetic prediction of common diseases: still no help for the clinical diabetologist! Nutrition, Metabolism and Cardiovascular Diseases. 2012;22(11):929–936. doi: 10.1016/j.numecd.2012.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sluik D., Boeing H., Li K., et al. Lifestyle factors and mortality risk in individuals with diabetes mellitus: are the associations different from those in individuals without diabetes? Diabetologia. 2014;57(1):63–72. doi: 10.1007/s00125-013-3074-y. [DOI] [PubMed] [Google Scholar]
- 9.Dorman J. S., Bunker C. H. HLA-DQ locus of the human leukocyte antigen complex and type 1 diabetes mellitus: a HuGE review. Epidemiologic Reviews. 2000;22(2):218–227. doi: 10.1093/oxfordjournals.epirev.a018034. [DOI] [PubMed] [Google Scholar]
- 10.Schwenk R. W., Vogel H., Schürmann A. Genetic and epigenetic control of metabolic health. Molecular Metabolism. 2013;2(4):337–347. doi: 10.1016/j.molmet.2013.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Abujbara M. A., Liswi M. I., El-Khateeb M. S., Flanagan S. E., Ellard S., Ajlouni K. M. Permanent neonatal diabetes mellitus in Jordan. Journal of Pediatric Endocrinology and Metabolism. 2014;27(9-10):879–883. doi: 10.1515/jpem-2014-0069. [DOI] [PubMed] [Google Scholar]
- 12.Pattanayak A. K., Bankura B., Balmiki N., Das T. K., Chowdhury S., Das M. Role of peroxisome proliferator-activated receptor gamma gene polymorphisms in type 2 diabetes mellitus patients of West Bengal, India. Journal of Diabetes Investigation. 2014;5(2):188–191. doi: 10.1111/jdi.12130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hivert M.-F., Vassy J. L., Meigs J. B. Susceptibility to type 2 diabetes mellitus—from genes to prevention. Nature Reviews Endocrinology. 2014;10(4):198–205. doi: 10.1038/nrendo.2014.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Mitchell A. A., Chakravarti A., Cutler D. J. On the probability that a novel variant is a disease-causing mutation. Genome Research. 2005;15(7):960–966. doi: 10.1101/gr.3761405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bailey-Wilson J. E., Wilson A. F. Linkage analysis in the next-generation sequencing era. Human Heredity. 2011;72(4):228–236. doi: 10.1159/000334381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.McTaggart J. S., Clark R. H., Ashcroft F. M. The role of the KATP channel in glucose homeostasis in health and disease: more than meets the islet. Journal of Physiology. 2010;588(17):3201–3209. doi: 10.1113/jphysiol.2010.191767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ashcroft F. M. KATP channels and insulin secretion: a key role in health and disease. Biochemical Society Transactions. 2006;34(2):243–246. doi: 10.1042/BST20060243. [DOI] [PubMed] [Google Scholar]
- 18.Harakalova M., Van Harssel J. J. T., Terhal P. A., et al. Dominant missense mutations in ABCC9 cause Cantúsyndrome. Nature Genetics. 2012;44(7):793–796. doi: 10.1038/ng.2324. [DOI] [PubMed] [Google Scholar]
- 19.Abdelhamid I., Lasram K., Meiloud G., et al. E23K variant in KCNJ11 gene is associated with susceptibility to type 2 diabetes in the Mauritanian population. Primary Care Diabetes. 2014;8(2):171–175. doi: 10.1016/j.pcd.2013.10.006. [DOI] [PubMed] [Google Scholar]
- 20.Chen G., Xu Y., Lin Y., et al. Association study of genetic variants of 17 diabetes-related genes/loci and cardiovascular risk and diabetic nephropathy in the Chinese She population. Journal of Diabetes. 2013;5(2):136–145. doi: 10.1111/1753-0407.12025. [DOI] [PubMed] [Google Scholar]
- 21.Hu C., Zhang R., Wang C., et al. PPARG, KCNJ11, CDKAL1, CDKN2A-CDKN2B, IDE-KIF11- HHEX, IGF2BP2 and SLC30A8 are associated with type 2 diabetes in a chinese population. PLoS ONE. 2009;4(10) doi: 10.1371/journal.pone.0007643.e7643 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zhou D., Zhang D., Liu Y., et al. The E23K variation in the KCNJ11 gene is associated with type 2 diabetes in Chinese and East Asian population. Journal of Human Genetics. 2009;54(7):433–435. doi: 10.1038/jhg.2009.54. [DOI] [PubMed] [Google Scholar]
- 23.Tabara Y., Osawa H., Kawamoto R., et al. Replication study of candidate genes associated with type 2 diabetes based on genome-wide screening. Diabetes. 2009;58(2):493–498. doi: 10.2337/db07-1785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Sale M. M., Smith S. G., Mychaleckyj J. C., et al. Variants of the transcription factor 7-like 2 (TCF7L2) gene are associated with type 2 diabetes in an African-American population enriched for nephropathy. Diabetes. 2007;56(10):2638–2642. doi: 10.2337/db07-0012. [DOI] [PubMed] [Google Scholar]
- 25.Weedon M. N., McCarthy M. I., Hitman G., et al. Combining information from common type 2 diabetes risk polymorphisms improves disease prediction. PLoS Medicine. 2006;3(10) doi: 10.1371/journal.pmed.0030374.e374 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Mtiraoui N., Turki A., Nemr R., et al. Contribution of common variants of ENPP1, IGF2BP2, KCNJ11, MLXIPL, PPARγ, SLC30A8 and TCF7L2 to the risk of type 2 diabetes in Lebanese and Tunisian Arabs. Diabetes and Metabolism. 2012;38(5):444–449. doi: 10.1016/j.diabet.2012.05.002. [DOI] [PubMed] [Google Scholar]
- 27.Chavali S., Mahajan A., Tabassum R., et al. Association of variants in genes involved in pancreatic Β-cell development and function with type 2 diabetes in North Indians. Journal of Human Genetics. 2011;56(10):695–700. doi: 10.1038/jhg.2011.83. [DOI] [PubMed] [Google Scholar]
- 28.Wang F., Han X.-Y., Ren Q., et al. Effect of genetic variants in KCNJ11, ABCC8, PPARG and HNF4A loci on the susceptibility of type 2 diabetes in Chinese Han population. Chinese Medical Journal. 2009;122(20):2477–2482. doi: 10.3760/cma.j.issn.0366-6999.2009.20.015. [DOI] [PubMed] [Google Scholar]
- 29.Alsmadi O., Al-Rubeaan K., Wakil S. M., et al. Genetic Study of Saudi Diabetes (GSSD):significant association of the KCNJ11 E23K polymorphism with type 2 diabetes. Diabetes/Metabolism Research and Reviews. 2008;24(2):137–140. doi: 10.1002/dmrr.777. [DOI] [PubMed] [Google Scholar]
- 30.Liu Z., Zhang Y.-W., Feng Q.-P., et al. Association analysis of 30 type 2 diabetes candidate genes in Chinese Han population. Acta Academiae Medicinae Sinicae. 2006;28(2):124–128. [PubMed] [Google Scholar]
- 31.Vangipurapu J., Stančáková A., Pihlajamäki J., et al. Association of indices of liver and adipocyte insulin resistance with 19 confirmed susceptibility loci for type 2 diabetes in 6,733 non-diabetic finnish men. Diabetologia. 2011;54(3):563–571. doi: 10.1007/s00125-010-1977-4. [DOI] [PubMed] [Google Scholar]
- 32.Gonen M. S., Arikoglu H., Erkoc Kaya D., et al. Effects of single nucleotide polymorphisms in KATP channel genes on type 2 diabetes in a Turkish population. Archives of Medical Research. 2012;43(4):317–323. doi: 10.1016/j.arcmed.2012.06.001. [DOI] [PubMed] [Google Scholar]
- 33.He Y.-Y., Zhang R., Shao X.-Y., et al. Association of KCNJ11 and ABCC8 genetic polymorphisms with response to repaglinide in Chinese diabetic patients. Acta Pharmacologica Sinica. 2008;29(8):983–989. doi: 10.1111/j.1745-7254.2008.00840.x. [DOI] [PubMed] [Google Scholar]
- 34.Chistiakov D. A., Potapov V. A., Khodirev D. C., Shamkhalova M. S., Shestakova M. V., Nosikov V. V. Genetic variations in the pancreatic ATP-sensitive potassium channel, β-cell dysfunction, and susceptibility to type 2 diabetes. Acta Diabetologica. 2009;46(1):43–49. doi: 10.1007/s00592-008-0056-5. [DOI] [PubMed] [Google Scholar]
- 35.Sakamoto Y., Inoue H., Keshavarz P., et al. SNPs in the KCNJ11-ABCC8 gene locus are associated with type 2 diabetes and blood pressure levels in the Japanese population. Journal of Human Genetics. 2007;52(10):781–793. doi: 10.1007/s10038-007-0190-x. [DOI] [PubMed] [Google Scholar]
- 36.Koo B. K., Cho Y. M., Park B. L., et al. Polymorphisms of KCNJ11 (Kir6.2 gene) are associated with type 2 diabetes and hypertension in the Korean population. Diabetic Medicine. 2007;24(2):178–186. doi: 10.1111/j.1464-5491.2006.02050.x. [DOI] [PubMed] [Google Scholar]
- 37.Nielsen E.-M. D., Hansen L., Carstensen B., et al. The E23K variant of Kir6.2 associates with impaired post-OGTT serum insulin response and increased risk of type 2 diabetes. Diabetes. 2003;52(2):573–577. doi: 10.2337/diabetes.52.2.573. [DOI] [PubMed] [Google Scholar]
- 38.Shaat N., Ekelund M., Lernmark Å., et al. Association of the E23K polymorphism in the KCNJ11 gene with gestational diabetes mellitus. Diabetologia. 2005;48(12):2544–2551. doi: 10.1007/s00125-005-0035-0. [DOI] [PubMed] [Google Scholar]
- 39.Yu M., Xu X.-J., Yin J.-Y., et al. KCNJ11 Lys23Glu and TCF7L2 rs290487(C/T) polymorphisms affect therapeutic efficacy of repaglinide in Chinese patients with type 2 diabetes. Clinical Pharmacology and Therapeutics. 2010;87(3):330–335. doi: 10.1038/clpt.2009.242. [DOI] [PubMed] [Google Scholar]
- 40.Javorsky M., Klimcakova L., Schroner Z., et al. KCNJ11 gene E23K variant and therapeutic response to sulfonylureas. European Journal of Internal Medicine. 2012;23(3):245–249. doi: 10.1016/j.ejim.2011.10.018. [DOI] [PubMed] [Google Scholar]
- 41.Lang V. Y., Fatehi M., Light P. E. Pharmacogenomic analysis of ATP-sensitive potassium channels coexpressing the common type 2 diabetes risk variants E23K and S1369A. Pharmacogenetics and Genomics. 2012;22(3):206–214. doi: 10.1097/FPC.0b013e32835001e7. [DOI] [PubMed] [Google Scholar]
- 42.Holstein A., Hahn M., Stumvoll M., Kovacs P. The E23K variant of KCNJ11 and the risk for severe sulfonylurea-induced hypoglycemia in patients with type 2 diabetes. Hormone and Metabolic Research. 2009;41(5):387–390. doi: 10.1055/s-0029-1192019. [DOI] [PubMed] [Google Scholar]
- 43.El-sisi A. E., Hegazy S. K., Metwally S. S., Wafa A. M., Dawood N. A. Effect of genetic polymorphisms on the development of secondary failure to sulfonylurea in egyptian patients with type 2 diabetes. Therapeutic Advances in Endocrinology and Metabolism. 2011;2(4):155–164. doi: 10.1177/2042018811415985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Keshavarz P., Habibipour R., Ghasemi M., Kazemnezhad E., Alizadeh M., Omami M. H. Lack of genetic susceptibility of KCNJ11 E23K polymorphism with risk of type 2 diabetes in an Iranian population. Endocrine Research. 2014;39(3):120–125. doi: 10.3109/07435800.2013.860607. [DOI] [PubMed] [Google Scholar]
- 45.Danquah I., Othmer T., Frank L. K., Bedu-Addo G., Schulze M. B., Mockenhaupt F. P. The TCF7L2 rs7903146 (T) allele is associated with type 2 diabetes in urban Ghana: a hospital-based case-control study. BMC Medical Genetics. 2013;14, article 96 doi: 10.1186/1471-2350-14-96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gamboa-Meléndez M. A., Huerta-Chagoya A., Moreno-Macías H., et al. Contribution of common genetic variation to the risk of type 2 diabetes in the Mexican Mestizo population. Diabetes. 2012;61(12):3314–3321. doi: 10.2337/db11-0550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Iwata M., Maeda S., Kamura Y., et al. Genetic risk score constructed using 14 susceptibility alleles for type 2 diabetes is associated with the early onset of diabetes and may predict the future requirement of insulin injections among Japanese individuals. Diabetes Care. 2012;35(8):1763–1770. doi: 10.2337/dc11-2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ragia G., Tavridou A., Petridis I., Manolopoulos V. G. Association of KCNJ11 E23K gene polymorphism with hypoglycemia in sulfonylurea-treated type 2 diabetic patients. Diabetes Research and Clinical Practice. 2012;98(1):119–124. doi: 10.1016/j.diabres.2012.04.017. [DOI] [PubMed] [Google Scholar]
- 49.Hotta K., Kitamoto A., Kitamoto T., et al. Association between type 2 diabetes genetic susceptibility loci and visceral and subcutaneous fat area as determined by computed tomography. Journal of Human Genetics. 2012;57(5):305–310. doi: 10.1038/jhg.2012.21. [DOI] [PubMed] [Google Scholar]
- 50.Odgerel Z., Lee H. S., Erdenebileg N., et al. Genetic variants in potassium channels are associated with type 2 diabetes in a Mongolian population. Journal of Diabetes. 2012;4(3):238–242. doi: 10.1111/j.1753-0407.2011.00177.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Cho H. J., Lee S. Y., Kim Y. G., et al. Effect of genetic polymorphisms on the pharmacokinetics and efficacy of glimepiride in a Korean population. Clinica Chimica Acta. 2011;412(19):1831–1834. doi: 10.1016/j.cca.2011.06.014. [DOI] [PubMed] [Google Scholar]
- 52.Webster R. J., Warrington N. M., Beilby J. P., Frayling T. M., Palmer L. J. The longitudinal association of common susceptibility variants for type 2 diabetes and obesity with fasting glucose level and BMI. BMC Medical Genetics. 2010;11, article 140 doi: 10.1186/1471-2350-11-140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Meyer T. E., Boerwinkle E., Morrison A. C., et al. Diabetes genes and prostate cancer in the atherosclerosis risk in communities study. Cancer Epidemiology Biomarkers and Prevention. 2010;19(2):558–565. doi: 10.1158/1055-9965.EPI-09-0902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Nikolac N., Simundic A.-M., Katalinic D., Topic E., Cipak A., Zjacic Rotkvic V. Metabolic control in type 2 diabetes is associated with sulfonylurea receptor-1 (SUR-1) but not with KCNJ11 polymorphisms. Archives of Medical Research. 2009;40(5):387–392. doi: 10.1016/j.arcmed.2009.06.006. [DOI] [PubMed] [Google Scholar]
- 55.Ezzidi I., Mtiraoui N., Cauchi S., et al. Contribution of type 2 diabetes associated loci in the Arabic population from Tunisia: a case-control study. BMC Medical Genetics. 2009;10, article 33 doi: 10.1186/1471-2350-10-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Thorsby P. M., Midthjell K., Gjerlaugsen N., et al. Comparison of genetic risk in three candidate genes (TCF7L2, PPARG, KCNJ11) with traditional risk factors for type 2 diabetes in a population-based study—The HUNT study. Scandinavian Journal of Clinical and Laboratory Investigation. 2009;69(2):282–287. doi: 10.1080/00365510802538188. [DOI] [PubMed] [Google Scholar]
- 57.Christopoulos P., Mastorakos G., Gazouli M., et al. Genetic variants in TCF7L2 and KCNJ11 genes in a Greek population with polycystic ovary syndrome. Gynecological Endocrinology. 2008;24(9):486–490. doi: 10.1080/09513590802196379. [DOI] [PubMed] [Google Scholar]
- 58.Sanghera D. K., Ortega L., Han S., et al. Impact of nine common type 2 diabetes risk polymorphisms in Asian Indian Sikhs: PPARG2 (Pro12Ala), IGF2BP2, TCF7L2 and FTO variants confer a significant risk. BMC Medical Genetics. 2008;9, article 59 doi: 10.1186/1471-2350-9-59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Cauchi S., Nead K. T., Choquet H., et al. The genetic susceptibility to type 2 diabetes may be modulated by obesity status: implications for association studies. BMC Medical Genetics. 2008;9, article 45 doi: 10.1186/1471-2350-9-45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Lyssenko V., Almgren P., Anevski D., et al. Genetic prediction of future type 2 diabetes. PLoS Medicine. 2005;2(12) doi: 10.1371/journal.pmed.0020345.e345 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Laukkanen O., Pihlajamäki J., Lindström J., et al. Polymorphisms of the SUR1 (ABCC8) and Kir6.2 (KCNJ11) genes predict the conversion from impaired glucose tolerance to type 2 diabetes. The Finnish Diabetes Prevention Study. Journal of Clinical Endocrinology and Metabolism. 2004;89(12):6286–6290. doi: 10.1210/jc.2004-1204. [DOI] [PubMed] [Google Scholar]
- 62.Neuman R. J., Wasson J., Atzmon G., et al. Gene-gene interactions lead to higher risk for development of type 2 diabetes in an Ashkenazi Jewish population. PLoS ONE. 2010;5(3) doi: 10.1371/journal.pone.0009903.e9903 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ko J. M., Yang S., Kim S. Y., Lee H. S., Hwang J. S., Hwang I. T. E23K polymorphism of the KCNJ11 gene in Korean children with type 1 diabetes. World Journal of Pediatrics. 2012;8(2):169–172. doi: 10.1007/s12519-012-0355-3. [DOI] [PubMed] [Google Scholar]
- 64.Raj S. M., Howson J. M., Walker N. M., et al. No association of multiple type 2 diabetes loci with type 1 diabetes. Diabetologia. 2009;52(10):2109–2116. doi: 10.1007/s00125-009-1391-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Qiu L., Na R., Xu R., et al. Quantitative assessment of the effect of KCNJ11 gene polymorphism on the risk of type 2 diabetes. PLoS ONE. 2014;9(4) doi: 10.1371/journal.pone.0093961.e93961 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Pappa K. I., Gazouli M., Economou K., et al. Gestational diabetes mellitus shares polymorphisms of genes associated with insulin resistance and type 2 diabetes in the Greek population. Gynecological Endocrinology. 2011;27(4):267–272. doi: 10.3109/09513590.2010.490609. [DOI] [PubMed] [Google Scholar]
- 67.Ekelund M., Shaat N., Almgren P., et al. Genetic prediction of postpartum diabetes in women with gestational diabetes mellitus. Diabetes Research and Clinical Practice. 2012;97(3):394–398. doi: 10.1016/j.diabres.2012.04.020. [DOI] [PubMed] [Google Scholar]
- 68.Cho Y. M., Kim T. H., Lim S., et al. Type 2 diabetes-associated genetic variants discovered in the recent genome-wide association studies are related to gestational diabetes mellitus in the Korean population. Diabetologia. 2009;52(2):253–261. doi: 10.1007/s00125-008-1196-4. [DOI] [PubMed] [Google Scholar]
- 69.Kurzawski M., Dziewanowski K., Łapczuk J., Wajda A., Droździk M. Analysis of common type 2 diabetes mellitus genetic risk factors in new-onset diabetes after transplantation in kidney transplant patients medicated with tacrolimus. European Journal of Clinical Pharmacology. 2012;68(12):1587–1594. doi: 10.1007/s00228-012-1292-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Heni M., Ketterer C., Thamer C., et al. Glycemia determines the effect of type 2 diabetes risk genes on insulin secretion. Diabetes. 2010;59(12):3247–3252. doi: 10.2337/db10-0674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Lin Y., Li P., Cai L., et al. Association study of genetic variants in eight genes/loci with type 2 diabetes in a Han Chinese population. BMC Medical Genetics. 2010;11, article 97 doi: 10.1186/1471-2350-11-97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Cruz M., Valladares-Salgado A., Garcia-Mena J., et al. Candidate gene association study conditioning on individual ancestry in patients with type 2 diabetes and metabolic syndrome from Mexico city. Diabetes/Metabolism Research and Reviews. 2010;26(4):261–270. doi: 10.1002/dmrr.1082. [DOI] [PubMed] [Google Scholar]
- 73.Winkler C., Raab J., Grallert H., Ziegler A.-G. Lack of association of type 2 diabetes susceptibility genotypes and body weight on the development of islet autoimmunity and type 1 diabetes. PloS ONE. 2012;7(4) doi: 10.1371/journal.pone.0035410.e35410 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Sanda S., Wei S., Rue T., Shilling H., Greenbaum C. A SNP in G6PC2 predicts insulin secretion in type 1 diabetes. Acta Diabetologica. 2013;50(3):459–462. doi: 10.1007/s00592-012-0389-y. [DOI] [PubMed] [Google Scholar]
- 75.Feng Y., Mao G., Ren X., et al. Ser 1369Ala variant in sulfonylurea receptor gene ABCC8 is associated with antidiabetic efficacy of gliclazide in Chinese type 2 diabetic patients. Diabetes Care. 2008;31(10):1939–1944. doi: 10.2337/dc07-2248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Xu H., Murray M., McLachlan A. J. Influence of genetic polymorphisms on the pharmacokinetics and pharmacodynamics of sulfonylurea drugs. Current Drug Metabolism. 2009;10(6):643–658. doi: 10.2174/138920009789375388. [DOI] [PubMed] [Google Scholar]
- 77.Dehwah M. A. S., Xu A., Huang Q. MicroRNAs and type 2 diabetes/obesity. Journal of Genetics and Genomics. 2012;39(1):11–18. doi: 10.1016/j.jgg.2011.11.007. [DOI] [PubMed] [Google Scholar]
- 78.Alvarez M. L., DiStefano J. K. The role of non-coding RNAs in diabetic nephropathy: potential applications as biomarkers for disease development and progression. Diabetes Research and Clinical Practice. 2013;99(1):1–11. doi: 10.1016/j.diabres.2012.10.010. [DOI] [PubMed] [Google Scholar]
- 79.Dolphin A. C. A short history of voltage-gated calcium channels. British Journal of Pharmacology. 2006;147(1):S56–S62. doi: 10.1038/sj.bjp.0706442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Bryan J., Muñoz A., Zhang X., et al. ABCC8 and ABCC9: ABC transporters that regulate K+ channels. Pflugers Archiv European Journal of Physiology. 2007;453(5):703–718. doi: 10.1007/s00424-006-0116-z. [DOI] [PubMed] [Google Scholar]
- 81.Søberg K., Jahnsen T., Rognes T., Skålhegg B. S., Laerdahl J. K. Evolutionary paths of the cAMP-dependent protein kinase (PKA) catalytic subunits. PLoS ONE. 2013;8(4) doi: 10.1371/journal.pone.0060935.e60935 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Olson T. M., Alekseev A. E., Moreau C., et al. KATP channel mutation confers risk for vein of Marshall adrenergic atrial fibrillation. Nature Clinical Practice Cardiovascular Medicine. 2007;4(2):110–116. doi: 10.1038/ncpcardio0792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Guttula S. V., Rao A., Sridhar G., Chakravarthy M., Nageshwararo K., Rao P. Cluster analysis and phylogenetic relationship in biomarker identification of type 2 diabetes and nephropathy. International Journal of Diabetes in Developing Countries. 2010;30(1):52–56. doi: 10.4103/0973-3930.60003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Fogarty M. P., Cannon M. E., Vadlamudi S., Gaulton K. J. Identification of a regulatory variant that binds FOXA1 and FOXA2 at the CDC123/CAMK1D type 2 diabetes GWAS locus. PLoS Genetics. 2014;10(9) doi: 10.1371/journal.pgen.1004633.e1004633 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Alkorta-Aranburu G., Carmody D., Cheng Y. W., Nelakuditi V. Phenotypic heterogeneity in monogenic diabetes: the clinical and diagnostic utility of a gene panel-based next-generation sequencing approach. Molecular Genetics and Metabolism. 2014 doi: 10.1016/j.ymgme.2014.09.007. [DOI] [PMC free article] [PubMed] [Google Scholar]