

Abstract
The radiation is considered as a double edged sword, as its beneficial and detrimental effects have been demonstrated. The potential benefits are being exploited to its maximum by adopting safe handling of radionuclide stipulated by the regulatory agencies. While the occupational workers are monitored by personnel monitoring devices, for general publics, it is not a regular practice. However, it can be achieved by using biomarkers with a potential for the radiation triage and medical management. An ideal biomarker to adopt in those situations should be rapid, specific, sensitive, reproducible, and able to categorize the nature of exposure and could provide a reliable dose estimation irrespective of the time of the exposures. Since cytogenetic markers shown to have many advantages relatively than other markers, the origins of various chromosomal abnormalities induced by ionizing radiations along with dose-response curves generated in the laboratory are presented. Current status of the gold standard dicentric chromosome assay, micronucleus assay, translocation measurement by fluorescence in-situ hybridization and an emerging protein marker the γ-H2AX assay are discussed with our laboratory data. With the wide choice of methods, an appropriate assay can be employed based on the net.
Keywords: Biomarker, Dicentric chromosomes, Micronucleus, Fluorescence in-situ hybridization
Core tip: Of the well-established biomarker, the dicentric chromosome assay remains a gold standard, with sensitivity and specificity to radiation. In contrast, micronucleus is simple, rapid and potential for triage, though the sensitivity is less and not able to differentiate the partial body exposure from that of whole body exposure. The expensive fluorescence in-situ hybridization has the advantage that it can be employed in chronic and retrospective dose estimation. The γ-H2AX assay has a potential for triage despite the fact of limited stability. To conclude none of the assay could fulfil all the criteria of an ideal biomarker.
INTRODUCTION
Ever since the discovery of X-rays and radionuclide, their contribution towards the betterment of humankind is being augmented; thus nuclear technologies, which are finding increasing appliance in almost all walks of human endeavor, be it agriculture, medicine, power generation, research, etc. Similar to any other newer technologies, the nuclear technology is not entirely risk free. An increase in the concern of the accidental hazards linked to the use of ionizing radiation is currently being observed due to: (1) increased demand of radiation sources in several industrial applications, may leads to an higher probability of mishandling; (2) major contribution to the man-made sources of radiation, due to multiple procedures involving more time to treat complex and major disorder; and (3) growing nuclear threat, warfare and natural disaster like a recent events in Fukushima (Japan). At last, recently reported non-DNA targeted effects of ionizing radiations like bystander effects, genomic instability further complicates the risk for stochastic effects, have increased more concerns and fear among the public[1]. The risk associated with a technology can be reduced to satisfactory levels (in terms of cost benefit ratios) by scrupulous observation of practices proven to be safe. Practices of safe handling of radionuclides incurring least radiation exposure have been well established. Regulatory agency, the International Commission on Radiological Protection has laid down the permissible limits of radiation exposure to radiation workers (20 mSv/year) and to the general public (1 mSv/year). The amount of radiation received by a radiation worker is monitored generally by physical dosimeters like thermoluminescence dosimeter and film badge. In contrast, another potential source of exposures to the publics and radiation workers is due to unplanned activities and natural disaster; it is not a routine practice to wear the personnel monitoring devices by the exposed. Estimating the doses received during accidental conditions and management of exposed individuals, in the absence of personnel monitoring devices is an important issue towards medical management; biomarkers are proven to be a reliable tool for the above purpose.
SOURCES OF RADIATION EXPOSURE
Radioactive materials and radiation there from are a part of nature. Each one of us generally tends to associate radiation and radioactive materials not only with nuclear weapons and nuclear reactors alone. Several of the naturally occurring elements are radioactive, e.g., uranium, thorium, radium, and potassium, which are widely distributed as constituents of the earth’s crust. The content of radioactive material in the earth’s crust varies from place to place and on average, radiation exposure due to this natural source of radiation is about 41% of the total[2]. There are certain places in the world where the natural background radiation levels are 5 to 10 times higher than the average levels. In addition to radiation emitted by radioactive materials in the environment, man has also always been exposed to radiation of natural origin from outer space in the form of cosmic rays. Consistent with natural radiation, levels of exposure from both these sources differ from place to place. Cosmic rays are a form of extremely penetrating radiation coming from regions very far away in space. It was estimated that due to cosmic rays alone, the entire population on this earth receives about 16% of the total exposure from natural sources of radiation in a year. Cosmic ray contribution varies with altitude and latitude. While flying in aircraft, the passengers and crew receive about many fold greater exposure than on the ground. In recent times, the use of artificial sources of radiation has grown extensively. Such usage has contributed to human welfare in agriculture, medicine, industry and research. Of which the largest source of human-made radiation exposure are from medical procedures, which is around 0.4 mSv[3]. Among the medical procedures, amount of exposures depends upon procedures and it is as low 0.2 mSv in chest X-ray examination to as high as 450 mSv among interventional procedures like heart catheterization before by-pass surgery. Recently, it has been shown that the annual per capita effective dose from diagnostic medical uses of radiation increased from 0.54 mSv to about 3.0 mSv to US population; the largest contribution and increases have come primarily from CT scanning and nuclear medicine[4]. This has also resulted in a small addition to the already present radiation exposure from natural sources. The estimated worldwide annual per capita effective dose from natural background is 2.4 mSv (Table 1). Radiation exposure in principle has a potential for causing harm to the life. Therefore, excessive and unnecessary exposures to radiation must be avoided. Exposure to radiation of natural origin cannot be kept in line; even so, the exposure due to radiation of artificial origin can be promptly checked. The level and methods of control are matters of scientific and expert judging.
Table 1.
World wide annual per capita effective doses in year 2000 (UNESCAR 2000)
| Source | Worldwide annual per capita effective dose (mSv) |
| Natural background | 2.4 |
| Diagnostic and medical examinations | 0.4 |
| Atmospheric testing | 0.005 |
| Chernobyl | 0.002 |
| Nuclear power production | 0.0002 |
BIOMARKERS OF RADIATION EXPOSURE
Exposure to radiation induces certain changes on the proteins, carbohydrates, lipids, nucleic acids and gene expression in the exposed cell, which are collectively known as biomarkers. In particular, traversal of ionizing radiation in a cellular system can bring about a variety of changes such as base damages, alkylation, intercalation adduct formation, nucleotide modifications, single strand and double strand breaks in the deoxyribonucleic acid (DNA)[5]. Those changes can result either due to direct deposition of energy on the nucleic acids (direct action) or can be mediated by the actions of free radicals released at some point in the interaction with water (indirect action) and membrane (lipid peroxidation) covers the cells[6]. Any measurement reflecting an interaction between a radiation exposures and biological system is defined as biomarkers[7]. The biomarkers are classified based on the changes being looked into like chromosomal aberrations, alterations in cell number, change in an enzyme level and or activity, proteins, or expression of genes, etc[8]. Of late based on the temporal parameters, it has been classified into markers of exposure, marker of susceptibility, markers of late effects and markers of persistent effects[9]. Thus, the manifestations of any of those changes are resulted due to the traversal of ionization track and deposition of energy in exposed cells/tissues. A summary of biomarkers of radiation exposures listed in the literatures is given in Figure 1.
Figure 1.
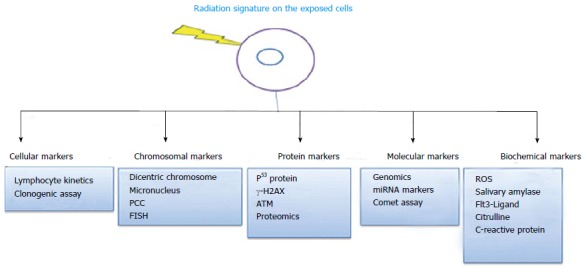
Various biomarkers of ionizing radiation exposure. FISH: Fluorescence in situ hybridization; PCC: Premature chromosome condensation; ROS: Reactive oxygen species; ATM: Ataxia telangiectasia mutated.
TECHNIQUES USED IN RADIATION BIODOSIMETRY
Radiation biodosimetry means, the quantification of the absorbed dose with the help of biological material obtained from an exposed individual. Of the various biomarkers, the extent of which can get expresses varied upon the quantum of exposure, absorbed dose, dose rate, energy of incident photons and radio-sensitivity of the exposed system. Similarly, time needed to express the changes and its stability in the exposed system depends upon those physical factors and the division kinetics of the cells[1]. A large number of protein biomarkers are tested for radiation dosimetry; despite the fact those changes are generally accurate, but cannot be effectively used to quantify the dose, as the level of these changes comes back to normal within short duration after exposure. Alternate to the protein biomarkers, cytogenetic indicators remain stable for a long time and provided a reliable estimate of the dose (Table 2). Dose estimation using the cytogenetic analysis is based on the relationship between chromosome aberration frequency and the amount of absorbed dose. The preferred choice of sample to analyze aberration frequency is the blood lymphocytes as they are easy to collect, culture and processing for biodosimetric studies. Exposed lymphocytes show different types of chromosome aberrations like dicentric chromosome (DC), centric ring, acentrics and translocation, all of which can be related to dose. Low background frequency, specificity to ionizing radiation, a clear dose-effect relationship for high and low linear energy transfer (LET) radiation with different dose and dose rates, reproducibility and comparability of in vitro to in vivo results[10] are several important biological parameters for reliable dose estimation. To keep above views in mind, we have established a laboratory to employ the DC, micronucleus (MN), Translocations and γ-H2AX assay for biodosimetry applications. Two decades experience of those methodology development, improvements and implementation of the assay for regular biodosimetry application is discussed in the present review along with current international status.
Table 2.
A few characteristic features of established bio-markers
| S. No | Parameter |
Technique |
|||
| DC | MN | FISH | γ-H2AX | ||
| 1 | Culture time (h) | 48 | 72 | 48 | Not applicable |
| 2 | Scoring speed (cells/d) | About 150 | About 750 | About 750 | About 100/h |
| 3 | Type of aberrations detectable | Unstable | Unstable | Stable/ unstable | Unstable |
| 4 | Period of detection after exposure (yr) | 2-3 | 2-3 | > 30 | 2-3 d |
| 5 | Cell type/quality | Metaphases/ good | BN cells/good | Metaphases/good | Interphase cells/good |
| 6 | Baseline frequency | 0.001 | 0.015 | 0.001 | 0.042 |
| 7 | Sensitivity (Gy) | 0.1 | 0.25 | 0.1 | 0.05 |
FISH: Fluorescence in situ hybridization; MN: Micronucleus; DC: Dicentric chromosome.
ORIGIN OF CHROMOSOMAL ABERRATIONS
In general, the chromosomes present in a cell are highly vibrant and undergoes extreme morphological changes at different phases within the cell cycle. When an ionization track travels along the cell nucleus, it can induce ionization on the DNA. Alternate, more than one track can pass through in different directions and induce much ionization within the same cell nucleus. Upon the energy deposition, it can induce many changes in the exposed cells and in turn the cells respond to those changes explicitly activation player molecules involved in check points activation, DNA repair and apoptosis[11]. The end result and fate of the cells depends on the many physical parameters of the incident photon as well as the cellular biological machinery. The chromosome aberrations are formed predominantly due to the repair activation that results in perfect rejoining or mis-rejoin to form chromosome aberrations. Thus, the aberration produced depends on the number of breaks, chromatids and chromosomes as well as its proximity of induced breaks involved[12]. The type, complexity and frequency of aberrations induced by radiations are diverse which are traditionally being in use to quantify and relate to the absorbed dose (Figure 2). Among chromosomal changes, they are named based on the methodology employed, or stain used (giemsa or fluorescence) to observe those changes or the end product (micronucleus, translocations)[13].
Figure 2.
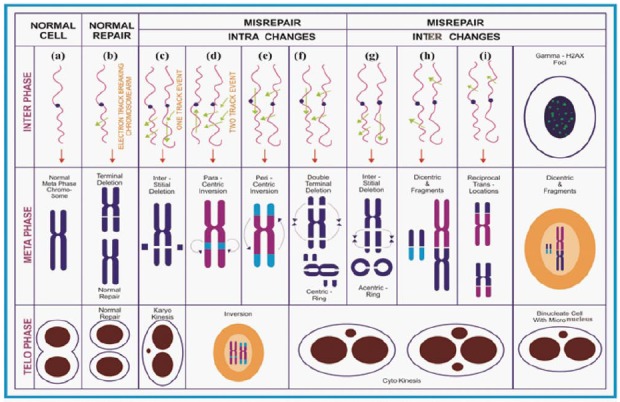
Diagrammatic illustration on the formation of ionizing radiation induced chromosome aberrations.
CHROMOSOME ABERRATION ASSAY
Studies on chromosomal aberration in Tradescantia microspore with X-rays in the 1930s marked the birth of radiation Cytogenetics[14]. In later years Sax[14] constructed the dose-response curves for both X-ray and neutron and defined chromosome and chromatid type of aberrations. In 1955, Revell[15] proposed the concept of intra and inter chromosomal exchanges and indicated that two lesions are necessary to initiate the exchange followed by forming an exchange type aberration, and failure to complete the exchange will give rise to deletions. The discovery of the clastogenic effect of radiation, gave rise to developmental studies of the dose - effect relationship. In 1962, Lea et al[16] formulated an equation for the dose-response curve obtained with X-ray. He proposed that the pattern of chromosome aberrations follows a Poisson distribution. The pioneering work on cytogenetics, has evolved and come a long way and made possible determination of dose by monitoring the effect and brought into the study of cytogenetic indicators, to estimate radiation absorbed dose. The various biological indicators, which have been reviewed by several authors[7,9,13].
Among the various indicators, DC aberration in the blood lymphocytes of exposed individual is the one which is mainly used for dose measurement[3]. They are chromosomes with two centromeres, which differ from its normal structure with one centromere; complex exchanges even a chromosome with more than two centromeres also is possible. It means, formation of DC is a complex event, because it needs double strand breaks (DSB) in at least two different chromosomes, which should in close proximity to each other so that the probability is high to form abnormal structure[17]. This technique, is well standardized as it is specific, and comparatively sensitive. However, each laboratory should generate its own dose-response curves, this includes the DC of the control population living in that area. It is because the baseline frequency greatly influences the co-efficient in a reference dose response curve[18]. The background DC frequency obtained is 0.002 (0.001, from 8000 metaphases scored), which are comparable to the published values obtained within India[19] and others[3]. In order to estimate the dose during accidental exposures the dose-response should be constructed from the result obtained with blood samples irradiated in less than 15 min with sufficient dose points. Then the exposed lymphocytes are cultured for 48 h under aseptic conditions to prepare a good quality metaphase chromosome and high mitotic index. Later stained slides are used to measure the number of DC at each dose and their frequency is used to construct a reference dose-response curve. Accurate identification of DC from that of twist or artefacts in poorly prepared metaphases are the challenges involved in this assay. The uncertainties can be reduced in combining centromere FISH technique[20]. It has been shown that the number of DC obtained with a given amount of dose is the same when irradiated either in vitro or in vivo condition[21]. Thus the dose-response curve constructed under in vitro condition is applicable for estimating the accidental radiation exposure to plant personnel. The dose response curve follows the equation Y = C + αD or Y = C + αD + βD2 depending upon the nature of radiation (Figure 3). The linear component (αD) often interpreted as the number of aberrations formed due to the traversal of single particle track and is expected to be independent of dose-rate. In alternate the dose squared (βD2) term is formed due to the interaction between two independent particle tracks and its degree determined by the time interval between the two tracks. Thus a delay of time permits repair of damage thereby decreasing the yield of aberration involving interchanges between two chromosomes. In the case of high LET radiation the dose-response curve mostly follows the equation Y = C + αD. Representative images of normal metaphase and a metaphase with DC obtained from a human blood lymphocytes exposed to 60Co-γ-irradiation and the co-efficient for the obtained dose response is given in Figure 4.
Figure 3.
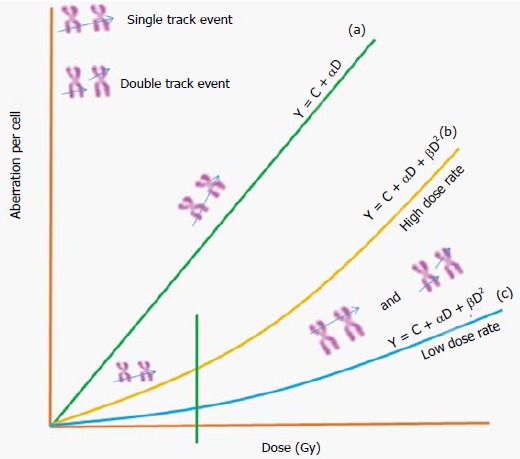
General dose response relationship for chromosome aberrations induced by different types of ionizing radiations.
Figure 4.
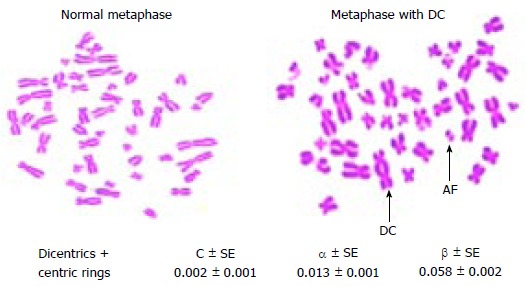
Metaphase chromosomes with (or) without dicentric chromosomes and dose response curve coefficients obtained from peripheral blood lymphocytes. AF: Acentric fragment; DC: Dicentric chromosome.
MN ASSAY
The chromosome fragments or whole chromosomes, which are failing to incorporate in the nuclei of daughter cells are known as micronuclei. Generally they are regular in shape with a similar staining intensity to that of daughter nuclei and within the cytoplasm of the daughter cells are called as micronuclei (Figure 5)[22]. MN reflects chromosomal damage and is a useful index for monitoring environmental effects on genetic material in human cells[23]. Due to the simplicity and the rapidity of scoring, this assay has shown promising potential in the triage medical management. However, due to background frequency of spontaneous MN frequency (0.002 to 0.036/cells) the sensitivity is 0.25 Gy[3]. Matter et al[24] coined the term MN based on its size and appearance. Fenech et al[22] developed a simple, most effective and reliable methodology to select cells between first and second mitosis division using cytochlasin-B; it inhibits cell division at cytokinesis in a cycling cell and, results in the binucleated cells and named as cytokinesis blocked micronucleus (CBMN) assay. The CBMN assay in addition to measuring the MN, it can also be used to measure nuclear-plasmic bridges, nuclear buds, necrotic cells, apoptotic cell and nuclear division rate collectively known as cytome assay[25]. Several studies have been carried out using the MN analysis in vitro and in vivo, for the purposes of biological dosimetry. A good correlation between the doses estimated from the MN frequency was observed in radiation workers[26] and in thyroid cancer patients undergoing radioiodine treatment[27]. A large volume of published reports for in vitro dose response curves is available[19,28-30]. An important caution is that many factors like age, genetic makeup and storage of blood samples could influence the dose estimation using the MN assay[31]. Similar to DC many laboratories has established dose response curve to estimate the dose; it follows linear-quadratic pattern despite the fact that there are differences in the obtained co-efficients among the established laboratories.
Figure 5.
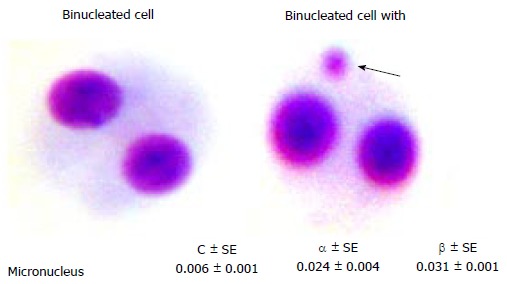
Binucleated cell with (or) without micronucleus and dose response curve coefficients obtained from peripheral blood lymphocytes.
FISH ASSAY
Despite the fact that scoring DC and MN is cost effective and well established assays for biodosimetry, both DC and MN are of unstable type aberrations and can get eliminated in a cycling cell over a period of time. Whereas, stable aberrations like reciprocal translocation (RT), induced by radiation has been shown to remain in circulation for longer periods. Provided if the progenitor cells are also exposed[3]. As it has been suggested that measurement of such RT may provide cumulative radiation exposure, we standardized FISH technique to score translocations (TL). This technique is based on the higher affinity among nucleotide bases in homologous sequences compared to non-homologous sequences. By using fluorescent labelled DNA probes, one could selectively paint a chromosome/set of chromosomes, which can be seen easily under fluorescence microscope. During hybridization the fluorescent labelled DNA probes bind to its complementary strand which helps in the detection of rearrangement, if any, which has taken place in these labelled chromosomes. The chromosomes which are not painted with fluorescent material are stained with different colors. The fluorescent labelled chromosome if undergone translocations will exhibit a bicolour and one can easily identify. Since the introduction of assays to measure RT, FISH have been pre-dominantly used in various laboratories[32-39] because of its simplicity in scoring and rapidity. Generally, the dose was estimated by measuring the RT in painting few chromosomes and extrapolating to the whole genome translocation frequency; else if any exchanges between non-painted chromosomes go undetected. However, it was extrapolated to whole genome with assumptions that radiation induced break points and translocation formation are randomly distributed throughout the genome, frequency of translocation is directly proportional to the DNA content and size of chromosomes without any hotspots on selective chromosomes. However, literature evidenced that radiation induced break points are distributed randomly in A-bomb survivors[40]. In vitro exposure as well as non-randomly[41-43]. Many laboratories have established dose-response curves by a selective painting of few chromosomes (Figure 6). Rapid developments in the probe labelling methodology, optics and imaging modalities, the assay has evolved in different directions like m-FISH, SKY-FISH, and m-band[44] where exchanges involved in any chromosomes or regions within chromosomes can be identified easily similar to that, GTG-banding technique have been in use for the identification of aberrations in individual chromosomes[45] as well as in entire genomes. It was an attractive option for many years back; however, RT measurements with latest FISH technology, and G-banding, in dosimetry is limited because of either time factor and/or cost factor. However, it can provide a true estimation of translocation frequency by analyzing the individual chromosomes for chronic dose estimation.
Figure 6.

Metaphase chromosomes with (or) without translocation and dose response curve coefficients after whole chromosome painting.
γ-H2AX ASSAY
Markers based on the chromosome abnormalities and/or gene mutations are suitable to quantify the residual damage and not the actual amount of damages induced due to exposure. This is for the reason that, to score the aberrations, the exposed cells have to be cultured, arrested in to suitable stage and then to score sufficient number of cells does extrapolate into the dose. Exposures of living organisms to radiation can induce assortment of DNA damages including DSB. Many molecules of histone H2AX at the broken site are rapidly phosphorylated on serine 139 in the C-terminus among the living organisms. In turn multiple factors involved in DNA repair and chromatin remodelling are assembled at the broken site and forms the γ-H2AX foci[46]. The γ-H2AX are simply visualized with antibodies to γ-H2AX with each DSB yielding one focus. Currently, γ-H2AX foci frequency is measured by immunocytochemistry, Western blot analysis and single-well flow cytometry (Figure 7). Measurement of γ-H2AX foci from peripheral blood lymphocytes (PBL) is used as a prospective biomarker to assess the radiation dose[47]. A dose-dependent increase and time-dependent reduction of γ-H2AX foci has been reported in cancer cells after exposure to ionizing radiation, patients who underwent radiation therapy, and in personnel after computer tomography (CT) imaging[48]. The assay was reported to distinguish partial and total body irradiation[49]. Flow cytometry measurement of γ-H2AX fluorescence intensity suggests that more samples can be analyzed in a short duration with lower sensitivity when compared to foci counting using microscopy[50].
Figure 7.
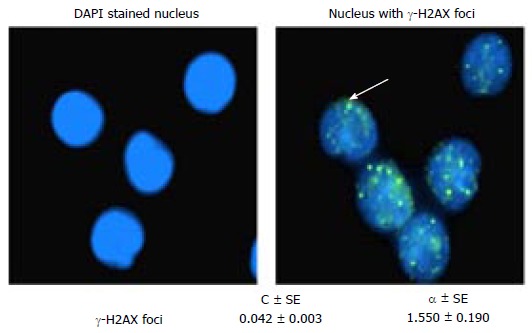
4’,6-diamidino-2-phenylindole, dihydrochloride stained nucleus with (or) without γ-H2AX foci and their coefficients obtained from peripheral blood lymphocytes.
In spite of rapid progress in technical development and advantages, variables like age[51], smoking[52], oxidative stress, inflammation[53], heat[54], genetic factors, etc., have been reported to influence the γ-H2AX foci levels. Inter-individual variability of γ-H2AX fluorescence intensity was observed in human PBL of healthy individuals[55]. Consistent with cytogenetic abnormalities, baseline γ-H2AX foci has been reported have been shown wide inter-individual variation[56,57]. Amusingly, though speed is an added advantage, a difference in γ-H2AX foci yields obtained from the same samples by different laboratories and methodical underestimation of doses was a major concern when using flow cytometry; variations in foci loss during shipment of blood samples, or variations in immunofluorescence staining quality were listed and should be minimized to reduce the uncertainties[58]. It was also emphasized that one should not look upon any calibration curves for this assay as set in standard like that of DC and MN assays; as an alternative, the γ-H2AX foci assay should be frequently recalibrated to take into account any drift in foci yields, and protocols should be optimized to reduce variability and ensure consistency. The mean γ-H2AX foci frequency obtained from our laboratory by scoring 26400 cells from healthy subjects (n = 130) is 0.042 ± 0.001 (unpublished data) which is comparable to that of reported values; thus the mean yield of γ-H2AX ± SD reported was 0.09 ± 0.05 with a range of 0.01 to 0.17 foci per lymphocyte[59].
RECENT DEVELOPMENTS TOWARDS TRIAGE
Inter-laboratory comparisons
The preliminary dose estimation and segregation of exposed and non-exposed individuals are the main step in the management of triage. Moreover the first responders also need to be monitored periodically, to ensure the dose levels they exposed during evacuation. In such scenario, to meet the demand alternative strategies is being developed as classical cytogenetic methods (DC and MN) by manual scoring is time consuming; sharing of the workload among the expert groups, automation of analytical methods, and early markers to ionizing radiation are the recent advancements in biodosimetry. Available literature has demonstrated ample evidences that many laboratories are well equipped and use more than one methodology to estimate the dose in an exposed individual. The scoring of DC from the PBL of individuals exposed to radiation remains the “gold standard” in biological dosimetry[3]. However, it has its limitation in potential scenarios of radiation exposure resulting in mass casualties owing the time needed for analysis. Of late to handle mass radiation casualties, countries have developed competencies in biological dosimetry[60]. In parallel to handle radiation triage, an inter-laboratory comparison exercise has been carried out among the established facility with a good sensitivity and minimize uncertainties in dose estimation[61]. The International Atomic Energy Agency and International Organization for Standardization played a vital role and provided guidelines to achieve the above goal[60]. As per the directions WHO a revised regulation in the field of radio nuclear incidents has been well established among the “BioDoseNet”, connected laboratories[62,63]. Such a networking and quality assurance in biodosimetry, is well established in the United States[64], Canada[65], Japan[66], Europe[67], Portuguese[68] and India[20,69].
Inter-laboratory comparisons and automation of MN scoring
Significant efforts were made to harmonize protocol to utilize simple and rapid scoring of MN as an alternative to manage large scale radiation accidents[3]. To minimize the individual discrepancies in the scoring of MN, an intra and inter-laboratory exercise was carried out among 34 laboratories; however, it was emphasized that it is paramount important to rectify scorers variation in analyzing the micro nucleated cells[70] and this can be reduced minimally by an automation in scoring of MN as it reduces labor and individuals scoring variations in addition to enhancing throughput.
There are two different kinds of automated methods that are presently being used to analyze the MN are (1) flow cytometry[71]; and (2) MN counting by image analysis[72]. While, the common advantage of both the methods is fast acquisition and analysis of the data in less time, it was cautioned that sensitivity is a limitation in case of flow cytometry based scoring of MN due to unspecified debris[73]. In spite of the potential for rapid scoring of MN with flow-cytometry, difficulty in discriminating MN from artifacts leading to a false-positive interpretation[74] and compromise in the sensitivity; furthermore, sample preservation and re-analysis are added limitations[75]. Therefore the automated image cytometry is preferred, as improved computer algorithms and allow rapid image analysis on cell-by-cell basis with a higher sensitivity. Moreover, with automated imaging system one can score the same slides repeatedly provided steps are taken to reduce background signals of the slides, which can be accounted as the MN in binucleate (BN) cells[76]. Though the speed was increased using automated scoring, it is able to detect fifty percent of the BN cells and seventy five percent of the MN in those cells. It was attributed that relative high inaccuracy in the classification of the BN cells[77,78]. Of late, systems like Meta Systems Metafer MN Score[79], IMSTAR Pathfinder™ Screentox Auto-MN[80] and Compucyte iCyte® Laser Scanning cytometer[81], which are commercially available to increase the scoring speed of MN with a better accuracy in identifying the MN and BN cells. The RABiT system developed by the Columbia University can be used to estimate absorbed dose based in MN and γ-H2AX scoring in a large number of populations with less time and small quantity of sample[82,83]. But, however, all labs cannot have this fully automated facility and it is not feasible to use at all places due to its cost.
Considering the importance of the time, rapid analysis in case of large population exposures, methods is being developed for automated scoring of MN[78]. Nonetheless, it is significant to observe that there exists a variation in the yield of MN scored in BN cells stained with giemsa depend on the adopted scoring method; it was suggested that the difference in the MN yield due to scoring methods can be reduced when they were scored the cells stained with fluorescence dyes like propidium iodide (PI) and 4’, 6-diamidino-2-phenylindole[76]. In considering the potential of the methodology, we carried out a systemic analysis of MN frequencies induced for different doses of γ-radiation in giemsa and PI stained BN cells, obtained from PBL by manual and automated scoring methods in-lieu of biological dosimetry for triage medical management. Immediate triage and high throughput dosimetry are more important in the medical management of radiation accidents. At the same time, it is equally important that the accuracy of the assay and reliable dose estimation at a later time for important cases identified by triage. The obtained results suggest that automated MN scoring in PI stained slides analysed with Meta Systems would be a better choice for the segregation and dose estimation than scoring in the BN cells stained with giemsa[30].
Inter-laboratory comparison and automation of γ-H2AX scoring
In explicit during triage owing to its time factor as one need not culture the sample for a few days to enumerate the damage, the γ-H2AX foci assay is an emerging technology. The γ-H2AX changes after irradiation are quantified mainly using either microscopy or flow cytometry[84,85]. Of which, the microscope counting of γ-H2AX foci (manual and automated) is the most preferred method than the flow cytometry as, it permits in detecting very low doses of radiation, differentiate the partial body exposures from that of whole body uniform exposure, higher specificity and its capability to estimate the doses even after 24 h of irradiation[49]. However, scoring the foci frequency manually with a microscope is somewhat time consuming than that of automated scoring. Whereas, automatic scoring of γ-H2AX foci could decrease the time of analysis, albeit its associated complication, like a higher standard error linked with fitted coefficient, loss on the sensitivity, and inability to categorize the nature of exposure due to over dispersed foci in automated scoring[86]. Moreover, the γ-H2AX foci method is sensitive and accurate after exposure to low doses, at higher doses overlapping of foci leads to underestimation of doses. Relative fluorescence intensity measurement using flow cytometry looks as a better option in case of radiological emergency at higher doses[55]. Nevertheless, speed is an added advantage of this assay, the difference in foci yields obtained from the same samples by different laboratories and systematic underestimation of doses were reported[87]. Thus, improvements have been made to reduce processing time[88], analysis speed[89], and time required to access dose in case of radiological emergencies using the γ-H2AX assay[90].
Realizing the prospective, many researchers have established the assay with modifications for a variety of applications in addition to biodosimetry and radiation triage. Similar to the well-established radiation specific DC assay, while many laboratories established their own dose-response curve[56], an inter-laboratory exercise has been carried out among the five European laboratories. Even though, there is no significant difference between the manual and automated scoring, the sensitivity of the assay is compromised and was unable to distinguish the partial exposures[86], NATO biodosimetry inter-comparison on γ-H2AX assay as tool for triage, revealed an increased time delay was inversely proportional to the foci frequency, in blood samples measured at 2 and 24 h post irradiation; variations in foci loss during shipment of blood samples or by differences in the immuno-fluorescence staining quality were listed as variables and should be minimized to reduce the uncertainties. Lately, technological advancement permitted of tele scoring of γ-H2AX foci among RENEB (Realizing European Network of Biodosimetry) laboratories; while the participant laboratories were able to distinguish critically high (> 2 Gy) and low dose and triage segregation of samples at 4 h, triage segregation of the 24 h samples shows high unpredictability. Apart from the variation in the shipment, variability in the staining quality under microscope (spectral and brightness differences in the light sources and fluorophores, wavelength ranges between different filters) and antibody could influence the foci analysis[58]. While, the manual scoring with the microscopy has the higher sensitivity of dose estimation, automated scoring with image analyser is a faster method for triage. However, flow cytometry can be employed for larger population. Despite the fact, those methods provides early dose estimation and radiation triage, it should be employed within 48 h post exposure, because the kinetic study demonstrated a reduction and reaches base line level of γ-H2AX foci.
CRITERIA FOR AN IDEAL BIOMARKER
There are many biomarkers reported for radiation exposure. An ideal biomarker should be specific, sensitive, and reproducible. Moreover, able to discriminate the nature of exposure (whole body from that of partial body) and could provide a reliable dose estimation irrespective of the time of exposures. Analysis of the marker and quantification of the dose should be rapid in particular at the time of triage. An, additional desirable characteristic is the possibility of using non-invasive and easy procedures for collection of biological samples[91]. Finally, validity of the assay measuring the biomarker and known variables influences the assay methods should be clearly established.
CONCLUSION
While all the techniques discussed in this review demonstrate the hallmark characteristic features the sensitivity and reproducibility, other features differ among the techniques. The DC is specific, sensitive (0.1 Gy), able to differentiate the nature of exposure partial body exposure from that of whole body exposure. Moreover being the unstable type of aberration, quantification of chronic exposure is difficult and it require more expertise and time despite the automated scoring as it need manual intervention even to score limited number of cells (about 50) in triage application. In alternate the MN is simple, rapid to score and easy to automate with a less sensitivity (0.25 Gy). Similarly, being an unstable type of aberration gets eliminated over a period of time and not suitable for chronic exposure as well as unable to discriminate the nature of exposure. Alternatively, the translocation measurement with fluorescence in situ hybridization is an expensive or labour intensive in case of G bands by trypsin using giemsa (GTG); nonetheless it provides an estimate of chronic and retrospective dose estimation with an equal sensitivity to DC, an essential criterion for occupational workers though it is not specific to radiation. However, the time needed to culture to look for all those aberrations is not needed, in γ-H2AX assay; thus the interphase cells could provide a reliable dose estimate with a sensitivity of 1 mGy using microscopy or triage with flow cytometry within 24-48 h beyond which is of limited use. To conclude none of the assay could fulfil all the criteria of ideal biomarkers. However with the wider choice an appropriate assay can be employed based on the need.
Footnotes
Supported by Department of Science and Technology, Government of India, No. SR-SO/HS-127/2012; Defence Research and Development Organisation, Government of India, No. DLS/81/48222/LSRB-261; and Atomic Energy Regulatory Board, Government of India (AERB/CSRP/Proj. No.58/04/2014).
Conflict-of-interest statement: The authors declare no conflict of interest.
Open-Access: This article is an open-access article which was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: May 27, 2015
First decision: June 18, 2015
Article in press: August 3, 2015
P- Reviewer: Storto G, Vinh-Hung V S- Editor: Tian YL L- Editor: A E- Editor: Wu HL
References
- 1.Little JB. Radiation carcinogenesis. Carcinogenesis. 2000;21:397–404. doi: 10.1093/carcin/21.3.397. [DOI] [PubMed] [Google Scholar]
- 2.Meenakshisundaram V, Rajagopal V, Mathiyarasu R. Lecture notes on radiation protection (Nucleonic well logging training programme) India: Institute of petroleum safety and environmental management, ONGC, DAE, GOI; 2005. pp. 14–24. [Google Scholar]
- 3.World Health Organization. Cytogenetic dosimetry: applications in preparedness for and response to radiation emergencies. Vienna, Austria: International Atomic Energy Agency, Department of Nuclear Safety and Security, Incident and Emergency Centre; 2011. [Google Scholar]
- 4.Mettler FA, Bhargavan M, Thomadsen BR, Gilley DB, Lipoti JA, Mahesh M, McCrohan J, Yoshizumi TT. Nuclear medicine exposure in the United States, 2005-2007: preliminary results. Semin Nucl Med. 2008;38:384–391. doi: 10.1053/j.semnuclmed.2008.05.004. [DOI] [PubMed] [Google Scholar]
- 5.Bedford JS, Dewey WC. Radiation Research Society. 1952-2002. Historical and current highlights in radiation biology: has anything important been learned by irradiating cells? Radiat Res. 2002;158:251–291. doi: 10.1667/0033-7587(2002)158[0251:hachir]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 6.Hall EJ, Giaccia AJ. Radiobiology for the Radiologist. Lippincott Williams and Wilkins. Vienna, Austria: International Atomic Energy Agency, Department of Nuclear Safety and Security, Incident and Emergency Centre; 2006. [Google Scholar]
- 7.Amundson SA, Fornace AJ. Gene expression profiles for monitoring radiation exposure. Radiat Prot Dosimetry. 2001;97:11–16. doi: 10.1093/oxfordjournals.rpd.a006632. [DOI] [PubMed] [Google Scholar]
- 8.Guipaud O, Benderitter M. Protein biomarkers for radiation exposure: towards a proteomic approach as a new investigation tool. Ann Ist Super Sanita. 2009;45:278–286. [PubMed] [Google Scholar]
- 9.Pernot E, Hall J, Baatout S, Benotmane MA, Blanchardon E, Bouffler S, El Saghire H, Gomolka M, Guertler A, Harms-Ringdahl M, et al. Ionizing radiation biomarkers for potential use in epidemiological studies. Mutat Res. 2012;751:258–286. doi: 10.1016/j.mrrev.2012.05.003. [DOI] [PubMed] [Google Scholar]
- 10.Romm H, Oestreicher U, Kulka U. Cytogenetic damage analysed by the dicentric assay. Ann Ist Super Sanita. 2009;45:251–259. [PubMed] [Google Scholar]
- 11.Sancar A, Lindsey-Boltz LA, Unsal-Kaçmaz K, Linn S. Molecular mechanisms of mammalian DNA repair and the DNA damage checkpoints. Annu Rev Biochem. 2004;73:39–85. doi: 10.1146/annurev.biochem.73.011303.073723. [DOI] [PubMed] [Google Scholar]
- 12.Sasaki MS. Advances in the biophysical and molecular bases of radiation cytogenetics. Int J Radiat Biol. 2009;85:26–47. doi: 10.1080/09553000802641185. [DOI] [PubMed] [Google Scholar]
- 13.Manning G, Rothkamm K. Deoxyribonucleic acid damage-associated biomarkers of ionising radiation: current status and future relevance for radiology and radiotherapy. Br J Radiol. 2013;86:20130173. doi: 10.1259/bjr.20130173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sax K. Chromosome Aberrations Induced by X-Rays. Genetics. 1938;23:494–516. doi: 10.1093/genetics/23.5.494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Revell SH. A new hypothesis for chromatid exchanges. Radiobiolog (Proc. Symp. Liège, 1954), London: Butterworths; 1955. pp. 243–253. [Google Scholar]
- 16.Lea D, Catcheside D. The mechanism of the induction by radiation on chromosome aberrations in tradesoantia. Joumal of Genetics. 1942;44:216–245. [Google Scholar]
- 17.Durante M, Bedford JS, Chen DJ, Conrad S, Cornforth MN, Natarajan AT, van Gent DC, Obe G. From DNA damage to chromosome aberrations: joining the break. Mutat Res. 2013;756:5–13. doi: 10.1016/j.mrgentox.2013.05.014. [DOI] [PubMed] [Google Scholar]
- 18.Lloyd DC. Chromosomal analysis to assess radiation dose. Stem Cells. 1997;15 Suppl 2:195–201. doi: 10.1002/stem.5530150727. [DOI] [PubMed] [Google Scholar]
- 19.Paul SF, Venkatachalam P, Jeevanram RK. A comparative study of synchronised and conventional culture methods on the micronucleus dose-response curve. Mutat Res. 1997;391:91–98. doi: 10.1016/s0165-1218(97)00038-4. [DOI] [PubMed] [Google Scholar]
- 20.Bhavani M, Tamizh Selvan G, Kaur H, Adhikari JS, Vijayalakshmi J, Venkatachalam P, Chaudhury NK. Dicentric chromosome aberration analysis using giemsa and centromere specific fluorescence in-situ hybridization for biological dosimetry: An inter- and intra-laboratory comparison in Indian laboratories. Appl Radiat Isot. 2014;92:85–90. doi: 10.1016/j.apradiso.2014.06.004. [DOI] [PubMed] [Google Scholar]
- 21.Bender MA, Awa AA, Brooks AL, Evans HJ, Groer PG, Littlefield LG, Pereira C, Preston RJ, Wachholz BW. Current status of cytogenetic procedures to detect and quantify previous exposures to radiation. Mutat Res. 1988;196:103–159. doi: 10.1016/0165-1110(88)90017-6. [DOI] [PubMed] [Google Scholar]
- 22.Fenech M, Morley A. Solutions to the kinetic problem in the micronucleus assay. Cytobios. 1985;43:233–246. [PubMed] [Google Scholar]
- 23.Odagiri Y, Uchida H, Shibazaki S. Interindividual variation in cytogenetic response to X-ray and colchicine measured with the cytokinesis-block micronucleus assay. Mutat Res. 1997;381:1–13. doi: 10.1016/s0027-5107(97)00138-3. [DOI] [PubMed] [Google Scholar]
- 24.Matter B, Schmid W. Trenimon-induced chromosomal damage in bone-marrow cells of six mammalian species, evaluated by the micronucleus test. Mutat Res. 1971;12:417–425. doi: 10.1016/0027-5107(71)90092-3. [DOI] [PubMed] [Google Scholar]
- 25.Fenech M. Cytokinesis-block micronucleus assay evolves into a “cytome” assay of chromosomal instability, mitotic dysfunction and cell death. Mutat Res. 2006;600:58–66. doi: 10.1016/j.mrfmmm.2006.05.028. [DOI] [PubMed] [Google Scholar]
- 26.Lee TK, O’Brien KF, Naves JL, Christie KI, Arastu HH, Eaves GS, Wiley AL, Karlsson UL, Salehpour MR. Micronuclei in lymphocytes of prostate cancer patients undergoing radiation therapy. Mutat Res. 2000;469:63–70. doi: 10.1016/s1383-5718(00)00072-3. [DOI] [PubMed] [Google Scholar]
- 27.Monsieurs MA, Bacher K, Brans B, Vral A, De Ridder L, Dierckx RA, Thierens HM. Patient dosimetry for 131I-lipiodol therapy. Eur J Nucl Med Mol Imaging. 2003;30:554–561. doi: 10.1007/s00259-002-1108-7. [DOI] [PubMed] [Google Scholar]
- 28.Sridevi K, Rao KP. Modification of genetic damage by dihomo-gamma-linolenic acid. Bull Environ Contam Toxicol. 1994;52:457–464. doi: 10.1007/BF00197837. [DOI] [PubMed] [Google Scholar]
- 29.Paillole N, Voisin P. Is micronuclei yield variability a problem for overexposure dose assessment to ionizing radiation? Mutat Res. 1998;413:47–56. doi: 10.1016/s1383-5718(98)00015-1. [DOI] [PubMed] [Google Scholar]
- 30.Selvan GT, Bhavani M, Vijayalakshmi J, Paul Solomon FD, Chaudhury NK, Venkatachalam P. Delayed mitogenic stimulation decreases DNA damage assessed by micronucleus assay in human peripheral blood lymphocytes after (60)co irradiation. Dose Response. 2014;12:498–508. doi: 10.2203/dose-response.13-060.Selvan. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Fenech M. Important variables that influence base-line micronucleus frequency in cytokinesis-blocked lymphocytes-a biomarker for DNA damage in human populations. Mutat Res. 1998;404:155–165. doi: 10.1016/s0027-5107(98)00109-2. [DOI] [PubMed] [Google Scholar]
- 32.Pinkel D, Gray JW, Trask B, van den Engh G, Fuscoe J, van Dekken H. Cytogenetic analysis by in situ hybridization with fluorescently labeled nucleic acid probes. Cold Spring Harb Symp Quant Biol. 1986;51 Pt 1:151–157. doi: 10.1101/sqb.1986.051.01.018. [DOI] [PubMed] [Google Scholar]
- 33.Natarajan AT, Vyas RC, Darroudi F, Vermeulen S. Frequencies of X-ray-induced chromosome translocations in human peripheral lymphocytes as detected by in situ hybridization using chromosome-specific DNA libraries. Int J Radiat Biol. 1992;61:199–203. doi: 10.1080/09553009214550821. [DOI] [PubMed] [Google Scholar]
- 34.Straume T, Lucas JN, Tucker JD, Bigbee WL, Langlois RG. Biodosimetry for a radiation worker using multiple assays. Health Phys. 1992;62:122–130. doi: 10.1097/00004032-199202000-00001. [DOI] [PubMed] [Google Scholar]
- 35.Schmid E, Zitzelsberger H, Braselmann H, Gray JW, Bauchinger M. Radiation-induced chromosome aberrations analysed by fluorescence in situ hybridization with a triple combination of composite whole chromosome-specific DNA probes. Int J Radiat Biol. 1992;62:673–678. doi: 10.1080/09553009214552621. [DOI] [PubMed] [Google Scholar]
- 36.Bauchinger M, Schmid E, Zitzelsberger H, Braselmann H, Nahrstedt U. Radiation-induced chromosome aberrations analysed by two-colour fluorescence in situ hybridization with composite whole chromosome-specific DNA probes and a pancentromeric DNA probe. Int J Radiat Biol. 1993;64:179–184. doi: 10.1080/09553009314551271. [DOI] [PubMed] [Google Scholar]
- 37.Nakano M, Nakashima E, Pawel DJ, Kodama Y, Awa A. Frequency of reciprocal translocations and dicentrics induced in human blood lymphocytes by X-irradiation as determined by fluorescence in situ hybridization. Int J Radiat Biol. 1993;64:565–569. doi: 10.1080/09553009314551781. [DOI] [PubMed] [Google Scholar]
- 38.Finnon P, Lloyd DC, Edwards AA. Fluorescence in situ hybridization detection of chromosomal aberrations in human lymphocytes: applicability to biological dosimetry. Int J Radiat Biol. 1995;68:429–435. doi: 10.1080/09553009514551391. [DOI] [PubMed] [Google Scholar]
- 39.Lucas JN, Hill F, Burk C, Fester T, Straume T. Dose-response curve for chromosome translocations measured in human lymphocytes exposed to 60Co gamma rays. Health Phys. 1995;68:761–765. doi: 10.1097/00004032-199506000-00001. [DOI] [PubMed] [Google Scholar]
- 40.Tanaka K, Popp S, Fischer C, Van Kaick G, Kamada N, Cremer T, Cremer C. Chromosome aberration analysis in atomic bomb survivors and Thorotrast patients using two- and three-colour chromosome painting of chromosomal subsets. Int J Radiat Biol. 1996;70:95–108. doi: 10.1080/095530096145373. [DOI] [PubMed] [Google Scholar]
- 41.Garcia-Canton C, Anadón A, Meredith C. γH2AX as a novel endpoint to detect DNA damage: applications for the assessment of the in vitro genotoxicity of cigarette smoke. Toxicol In Vitro. 2012;26:1075–1086. doi: 10.1016/j.tiv.2012.06.006. [DOI] [PubMed] [Google Scholar]
- 42.Rave-Fränk M, Virsik-Peuckert P, Schmidberger H, Rodemann HP. Reciprocal translocation frequency in irradiated sensitive and resistant human tumor cells in correlation with clonogenic in vitro cell survival: a possibility of tumor radiosensitivity prediction? Radiother Oncol. 1996;38:163–170. doi: 10.1016/0167-8140(95)01654-6. [DOI] [PubMed] [Google Scholar]
- 43.Boei JJ, Vermeulen S, Natarajan AT. Differential involvement of chromosomes 1 and 4 in the formation of chromosomal aberrations in human lymphocytes after X-irradiation. Int J Radiat Biol. 1997;72:139–145. doi: 10.1080/095530097143356. [DOI] [PubMed] [Google Scholar]
- 44.Trask BJ. Human cytogenetics: 46 chromosomes, 46 years and counting. Nat Rev Genet. 2002;3:769–778. doi: 10.1038/nrg905. [DOI] [PubMed] [Google Scholar]
- 45.Kumagai E, Tanaka R, Kumagai T, Onomichi M, Sawada S. Effects of long-term radiation exposure on chromosomal aberrations in radiological technologists. J Radiat Res. 1990;31:270–279. doi: 10.1269/jrr.31.270. [DOI] [PubMed] [Google Scholar]
- 46.Rogakou EP, Boon C, Redon C, Bonner WM. Megabase chromatin domains involved in DNA double-strand breaks in vivo. J Cell Biol. 1999;146:905–916. doi: 10.1083/jcb.146.5.905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Redon CE, Dickey JS, Bonner WM, Sedelnikova OA. γ-H2AX as a biomarker of DNA damage induced by ionizing radiation in human peripheral blood lymphocytes and artificial skin. Adv Space Res. 2009;43:1171–1178. doi: 10.1016/j.asr.2008.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Löbrich M, Rief N, Kühne M, Heckmann M, Fleckenstein J, Rübe C, Uder M. In vivo formation and repair of DNA double-strand breaks after computed tomography examinations. Proc Natl Acad Sci USA. 2005;102:8984–8989. doi: 10.1073/pnas.0501895102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Horn S, Barnard S, Rothkamm K. Gamma-H2AX-based dose estimation for whole and partial body radiation exposure. PLoS One. 2011;6:e25113. doi: 10.1371/journal.pone.0025113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Tanaka T, Halicka D, Traganos F, Darzynkiewicz Z. Cytometric analysis of DNA damage: phosphorylation of histone H2AX as a marker of DNA double-strand breaks (DSBs) Methods Mol Biol. 2009;523:161–168. doi: 10.1007/978-1-59745-190-1_11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Mah LJ, El-Osta A, Karagiannis TC. GammaH2AX as a molecular marker of aging and disease. Epigenetics. 2010;5:129–136. doi: 10.4161/epi.5.2.11080. [DOI] [PubMed] [Google Scholar]
- 52.Ishida M, Ishida T, Tashiro S, Uchida H, Sakai C, Hironobe N, Miura K, Hashimoto Y, Arihiro K, Chayama K, et al. Smoking cessation reverses DNA double-strand breaks in human mononuclear cells. PLoS One. 2014;9:e103993. doi: 10.1371/journal.pone.0103993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Li Z, Yang J, Huang H. Oxidative stress induces H2AX phosphorylation in human spermatozoa. FEBS Lett. 2006;580:6161–6168. doi: 10.1016/j.febslet.2006.10.016. [DOI] [PubMed] [Google Scholar]
- 54.Kaneko H, Igarashi K, Kataoka K, Miura M. Heat shock induces phosphorylation of histone H2AX in mammalian cells. Biochem Biophys Res Commun. 2005;328:1101–1106. doi: 10.1016/j.bbrc.2005.01.073. [DOI] [PubMed] [Google Scholar]
- 55.Ismail IH, Wadhra TI, Hammarsten O. An optimized method for detecting gamma-H2AX in blood cells reveals a significant interindividual variation in the gamma-H2AX response among humans. Nucleic Acids Res. 2007;35:e36. doi: 10.1093/nar/gkl1169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Sharma PM, Ponnaiya B, Taveras M, Shuryak I, Turner H, Brenner DJ. High throughput measurement of γH2AX DSB repair kinetics in a healthy human population. PLoS One. 2015;10:e0121083. doi: 10.1371/journal.pone.0121083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Sedelnikova OA, Horikawa I, Redon C, Nakamura A, Zimonjic DB, Popescu NC, Bonner WM. Delayed kinetics of DNA double-strand break processing in normal and pathological aging. Aging Cell. 2008;7:89–100. doi: 10.1111/j.1474-9726.2007.00354.x. [DOI] [PubMed] [Google Scholar]
- 58.Barnard S, Ainsbury EA, Al-hafidh J, Hadjidekova V, Hristova R, Lindholm C, Monteiro Gil O, Moquet J, Moreno M, Rößler U, et al. The first gamma-H2AX biodosimetry intercomparison exercise of the developing European biodosimetry network RENEB. Radiat Prot Dosimetry. 2015;164:265–270. doi: 10.1093/rpd/ncu259. [DOI] [PubMed] [Google Scholar]
- 59.Roch-Lefèvre S, Mandina T, Voisin P, Gaëtan G, Mesa JE, Valente M, Bonnesoeur P, García O, Voisin P, Roy L. Quantification of gamma-H2AX foci in human lymphocytes: a method for biological dosimetry after ionizing radiation exposure. Radiat Res. 2010;174:185–194. doi: 10.1667/RR1775.1. [DOI] [PubMed] [Google Scholar]
- 60.Roy L, Buard V, Delbos M, Durand V, Paillole N, Grégoire E, Voisin P. International intercomparison for criticality dosimetry: the case of biological dosimetry. Radiat Prot Dosimetry. 2004;110:471–476. doi: 10.1093/rpd/nch349. [DOI] [PubMed] [Google Scholar]
- 61.Garcia OF, Ramalho AT, Di Giorgio M, Mir SS, Espinoza ME, Manzano J, Nasazzi N, López I. Intercomparison in cytogenetic dosimetry among five laboratories from Latin America. Mutat Res. 1995;327:33–39. doi: 10.1016/0027-5107(94)00066-e. [DOI] [PubMed] [Google Scholar]
- 62.Di Giorgio M, Barquinero JF, Vallerga MB, Radl A, Taja MR, Seoane A, De Luca J, Oliveira MS, Valdivia P, Lima OG, et al. Biological dosimetry intercomparison exercise: an evaluation of triage and routine mode results by robust methods. Radiat Res. 2011;175:638–649. doi: 10.1667/RR2425.1. [DOI] [PubMed] [Google Scholar]
- 63.Christie DH, Chu MC, Carr Z. Global networking for biodosimetry laboratory capacity surge in radiation emergencies. Health Phys. 2010;98:168–171. doi: 10.1097/HP.0b013e3181abaad4. [DOI] [PubMed] [Google Scholar]
- 64.Wilkins RC, Romm H, Oestreicher U, Marro L, Yoshida MA, Suto Y, Prasanna PG. Biological Dosimetry by the Triage Dicentric Chromosome Assay - Further validation of International Networking. Radiat Meas. 2011;46:923–928. doi: 10.1016/j.radmeas.2011.03.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Miller SM, Ferrarotto CL, Vlahovich S, Wilkins RC, Boreham DR, Dolling JA. Canadian Cytogenetic Emergency network (CEN) for biological dosimetry following radiological/nuclear accidents. Int J Radiat Biol. 2007;83:471–477. doi: 10.1080/09553000701370860. [DOI] [PubMed] [Google Scholar]
- 66.Yoshida MA, Hayata I, Tateno H, Tanaka K, Sonta S. The chromosome network for biodosimetry in Japan. Radiat Mes. 2007;42:1125–1127. [Google Scholar]
- 67.Wojcik A, Lloyd D, Romm H, Roy L. Biological dosimetry for triage of casualties in a large-scale radiological emergency: capacity of the EU member states. Radiat Prot Dosimetry. 2010;138:397–401. doi: 10.1093/rpd/ncp279. [DOI] [PubMed] [Google Scholar]
- 68.Martins V, Antunes AC, Monteiro Gil O. Implementation of a dose-response curve for γ-radiation in the Portuguese population by use of the chromosomal aberration assay. Mutat Res. 2013;750:50–54. doi: 10.1016/j.mrgentox.2012.09.009. [DOI] [PubMed] [Google Scholar]
- 69.Bakkiam D, Bhavani M, Anantha Kumar AA, Sonwani S, Venkatachalam P, Sivasubramanian K, Venkatraman B. Dicentric assay: inter-laboratory comparison in Indian laboratories for routine and triage applications. Appl Radiat Isot. 2015;99:77–85. doi: 10.1016/j.apradiso.2015.02.015. [DOI] [PubMed] [Google Scholar]
- 70.Fenech M, Chang WP, Kirsch-Volders M, Holland N, Bonassi S, Zeiger E. HUMN project: detailed description of the scoring criteria for the cytokinesis-block micronucleus assay using isolated human lymphocyte cultures. Mutat Res. 2003;534:65–75. doi: 10.1016/s1383-5718(02)00249-8. [DOI] [PubMed] [Google Scholar]
- 71.Bryce SM, Avlasevich SL, Bemis JC, Lukamowicz M, Elhajouji A, Van Goethem F, De Boeck M, Beerens D, Aerts H, Van Gompel J, et al. Interlaboratory evaluation of a flow cytometric, high content in vitro micronucleus assay. Mutat Res. 2008;650:181–195. doi: 10.1016/j.mrgentox.2007.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Böcker W, Streffer C, Müller WU, Yu C. Automated scoring of micronuclei in binucleated human lymphocytes. Int J Radiat Biol. 1996;70:529–537. doi: 10.1080/095530096144734. [DOI] [PubMed] [Google Scholar]
- 73.Schreiber GA, Beisker W, Braselmann H, Bauchinger M, Bögl KW, Nüsse M. An automated flow cytometric micronucleus assay for human lymphocytes. Int J Radiat Biol. 1992;62:695–709. doi: 10.1080/09553009214552651. [DOI] [PubMed] [Google Scholar]
- 74.Laingam S, Froscio SM, Humpage AR. Flow-cytometric analysis of in vitro micronucleus formation: comparative studies with WIL2-NS human lymphoblastoid and L5178Y mouse lymphoma cell lines. Mutat Res. 2008;656:19–26. doi: 10.1016/j.mrgentox.2008.06.015. [DOI] [PubMed] [Google Scholar]
- 75.Fenech M, Kirsch-Volders M, Rossnerova A, Sram R, Romm H, Bolognesi C, Ramakumar A, Soussaline F, Schunck C, Elhajouji A, et al. HUMN project initiative and review of validation, quality control and prospects for further development of automated micronucleus assays using image cytometry systems. Int J Hyg Environ Health. 2013;216:541–552. doi: 10.1016/j.ijheh.2013.01.008. [DOI] [PubMed] [Google Scholar]
- 76.Varga D, Johannes T, Jainta S, Schuster S, Schwarz-Boeger U, Kiechle M, Patino Garcia B, Vogel W. An automated scoring procedure for the micronucleus test by image analysis. Mutagenesis. 2004;19:391–397. doi: 10.1093/mutage/geh047. [DOI] [PubMed] [Google Scholar]
- 77.Tates AD, van Welie MT, Ploem JS. The present state of the automated micronucleus test for lymphocytes. Int J Radiat Biol. 1990;58:813–825. doi: 10.1080/09553009014552191. [DOI] [PubMed] [Google Scholar]
- 78.Castelain P, Van Hummelen P, Deleener A, Kirsch-Volders M. Automated detection of cytochalasin-B blocked binucleated lymphocytes for scoring micronuclei. Mutagenesis. 1993;8:285–293. doi: 10.1093/mutage/8.4.285. [DOI] [PubMed] [Google Scholar]
- 79.Schunck C, Johannes T, Varga D, Lörch T, Plesch A. New developments in automated cytogenetic imaging: unattended scoring of dicentric chromosomes, micronuclei, single cell gel electrophoresis, and fluorescence signals. Cytogenet Genome Res. 2004;104:383–389. doi: 10.1159/000077520. [DOI] [PubMed] [Google Scholar]
- 80.Decordier I, Papine A, Plas G, Roesems S, Vande Loock K, Moreno-Palomo J, Cemeli E, Anderson D, Fucic A, Marcos R, et al. Automated image analysis of cytokinesis-blocked micronuclei: an adapted protocol and a validated scoring procedure for biomonitoring. Mutagenesis. 2009;24:85–93. doi: 10.1093/mutage/gen057. [DOI] [PubMed] [Google Scholar]
- 81.Darzynkiewicz Z, Smolewski P, Holden E, Luther E, Henriksen M, François M, Leifert W, Fenech M. Laser scanning cytometry for automation of the micronucleus assay. Mutagenesis. 2011;26:153–161. doi: 10.1093/mutage/geq069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Thierens H, Vral A. The micronucleus assay in radiation accidents. Ann Ist Super Sanita. 2009;45:260–264. [PubMed] [Google Scholar]
- 83.Nakamura A, Sedelnikova OA, Redon C, Pilch DR, Sinogeeva NI, Shroff R, Lichten M, Bonner WM. Techniques for gamma-H2AX detection. Methods Enzymol. 2006;409:236–250. doi: 10.1016/S0076-6879(05)09014-2. [DOI] [PubMed] [Google Scholar]
- 84.Redon CE, Nakamura AJ, Sordet O, Dickey JS, Gouliaeva K, Tabb B, Lawrence S, Kinders RJ, Bonner WM, Sedelnikova OA. γ-H2AX detection in peripheral blood lymphocytes, splenocytes, bone marrow, xenografts, and skin. Methods Mol Biol. 2011;682:249–270. doi: 10.1007/978-1-60327-409-8_18. [DOI] [PubMed] [Google Scholar]
- 85.Muslimovic A, Ismail IH, Gao Y, Hammarsten O. An optimized method for measurement of gamma-H2AX in blood mononuclear and cultured cells. Nat Protoc. 2008;3:1187–1193. doi: 10.1038/nprot.2008.93. [DOI] [PubMed] [Google Scholar]
- 86.Rothkamm K, Barnard S, Ainsbury EA, Al-Hafidh J, Barquinero JF, Lindholm C, Moquet J, Perälä M, Roch-Lefèvre S, Scherthan H, et al. Manual versus automated γ-H2AX foci analysis across five European laboratories: can this assay be used for rapid biodosimetry in a large scale radiation accident? Mutat Res. 2013;756:170–173. doi: 10.1016/j.mrgentox.2013.04.012. [DOI] [PubMed] [Google Scholar]
- 87.Rothkamm K, Horn S, Scherthan H, Rössler U, De Amicis A, Barnard S, Kulka U, Lista F, Meineke V, Braselmann H, et al. Laboratory intercomparison on the γ-H2AX foci assay. Radiat Res. 2013;180:149–155. doi: 10.1667/RR3238.1. [DOI] [PubMed] [Google Scholar]
- 88.Moquet J, Barnard S, Rothkamm K. Gamma-H2AX biodosimetry for use in large scale radiation incidents: comparison of a rapid ‘96 well lyse/fix’ protocol with a routine method. PeerJ. 2014;2:e282. doi: 10.7717/peerj.282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Barber P, Locke R, Pierce G, Rothkamm K, Vojnovic B. γ-H2AX foci counting: image processing and control software for high-content screening. Proceedings of SPIE. 2007;6441:64411M–1. [Google Scholar]
- 90.Turner HC, Sharma P, Perrier JR, Bertucci A, Smilenov L, Johnson G, Taveras M, Brenner DJ, Garty G. The RABiT: high-throughput technology for assessing global DSB repair. Radiat Environ Biophys. 2014;53:265–272. doi: 10.1007/s00411-014-0514-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Ohtaki K, Shimba H, Awa AA, Sofuni T. Comparison of type and frequency of chromosome aberrations by conventional and G-staining methods in Hiroshima atomic bomb survivors. J Radiat Res. 1982;23:441–449. doi: 10.1269/jrr.23.441. [DOI] [PubMed] [Google Scholar]