

Abstract
Oral vaccination can offer a painless and convenient method of vaccination. Furthermore, in addition to systemic immunity it has potential to stimulate mucosal immunity through antigen-processing by the gut-associated lymphoid tissues. In this study we propose the concept that pollen grains can be engineered for use as a simple modular system for oral vaccination. We demonstrate feasibility of this concept by using spores of Lycopodium clavatum (clubmoss) (LSs). We show that LSs can be chemically cleaned to remove native proteins to create intact clean hollow LS shells. Empty pollen shells were successfully filled with molecules of different sizes demonstrating their potential to be broadly applicable as a vaccination system. Using ovalbumin (OVA) as a model antigen, LSs formulated with OVA were orally fed to mice. LSs stimulated significantly higher anti-OVA serum IgG and fecal IgA antibodies compared to those induced by use of cholera toxin as a positive-control adjuvant. The antibody response was not affected by pre-neutralization of the stomach acid, and persisted for up to seven months. Confocal microscopy revealed that LSs can translocate in to mouse intestinal wall. Overall, this study lays the foundation of using LSs as a novel approach for oral vaccination.
Keywords: lycopodium spores, mucosal vaccination, oral vaccination, pollen shells, sporopollenin
1. Introduction
Oral vaccination is of significant interest because it is needle-free, painless, child-friendly, convenient, and amenable to self-administration. Furthermore, antigen delivery to the gastrointestinal tract has potential to induce mucosal immunity through antigen-processing via the gut-associated lymphoid tissues [1, 2]. Because majority of pathogens infect via mucosal surfaces, immunological defenses at mucosal surfaces can neutralize pathogens before they can cause infection [3].
However, oral vaccination remains daunting because the vaccine molecules experience a highly degradative environment in the stomach, and encounter a tough transport barrier offered by the tightly juxtaposed epithelial cells that line the intestinal mucosa [4]. To overcome these delivery challenges, live attenuated microorganisms such as bacteria and viruses, and particulate vaccine delivery systems such as liposomes, virosomes, polymeric micro and nanoparticles, and immune stimulating complexes have been examined [5, 6]. Various techniques that seek to target microfold (M) cells to enable higher antigen uptake through endocytosis have also been tested [7]. Toxins such as cholera toxin (CT) and heat labile enterotoxin (LT) that enable increased uptake of vaccine molecules have demonstrated the highest adjuvant effects [8, 9]. However, the toxicity of CT and LT naturally renders them unsuitable for human use [10]. The dangers of attenuated yet live strains of viruses and bacteria to become virulent, or their neutralization through pre-existing host immunity are some of the limitations of using microorganisms for oral vaccination [11]. Additionally, low encapsulation efficiency of antigens in polymeric particles, and poor vaccine stability resulting from exposure to organic solvents during particle synthesis has hindered clinical use of polymeric particles [12]. Plant-based edible vaccines expressed in rice [13], tobacco, tomato, carrot and potato have also been investigated [14]. Based on a twenty year development experience of an edible vaccine against hepatitis B, it has been found that just oral administration of edible plant-based hepatitis vaccine was insufficient for a protective response, yet when coupled in an oral-parenteral dosage regimen, it proved to be effective [14].
In this study we propose a novel concept to use pollen grains (PGs) as a natural system for oral vaccination. In nature, PGs facilitate pollination by functioning as a ‘safety-pod’ to carry the plant male gamete to the female reproductive organ, the ovary, located in the flower. During pollination PGs are often exposed to fluctuating temperatures and weather conditions, and to survive such conditions, they possess mechanical toughness and chemical resistance. Our postulate to use PGs for oral vaccination is based on such multiple natural properties of PGs that suggest their usefulness for oral vaccination: (i) they have a tough outer shell that can stay intact in the stomach environment [15, 16], (ii) the shell is naturally porous, and (iii) despite their relatively large size (tens of μm in diameter) it has been found that Lycopodium clavatum (clubmoss) spores (LSs) and Secale cereale (rye) pollen grains can cross the intestinal barrier as intact particles [17, 18]. Thus overall, we hypothesized that if (i) natural pores in the pollen wall could be used to clean and remove the allergy-causing native biomolecules from PGs, (ii) their clean ‘belly’ could be refilled with vaccine antigens through the natural pores in pollen walls, and (iii) the antigen-filled PGs could translocate across the intestinal epithelium into the body, then PGs might behave as natural ‘Trojan horses’ for oral vaccination ferrying the vaccines safely into the body. While LSs can survive the harsh acidic treatment, it has been suggested that enzymes in the body can degrade them [18, 19], thus providing a potentially safe natural carrier for oral vaccine and drug delivery. Indeed, using this conceptual framework LSs have recently been proposed for oral drug delivery. It has been shown that proteins as large as 540 kDa, a magnetic resonance imaging contrast agent, food oils including cod liver oil can be filled into LSs [19-23]. While these in vitro studies demonstrate the flexibility of filling LSs with different molecules, in vivo demonstrations on the effectiveness of pollens for oral drug and vaccine delivery are lacking.
Herein we demonstrate for the first time that LSs filled with ovalbumin (OVA) as a model antigen when fed orally to mice can induce a systemic and a mucosal immune response, which is superior to that stimulated by CT, a potent yet toxic mucosal adjuvant. We also investigated whether neutralization of stomach's acidic environment, prior to administration of the LS-based oral vaccine can affect the immune response. The durability of antigen-specific systemic and mucosal antibodies was also investigated, and it was found that OVA-specific antibodies could be detected in significant amounts up to seven months after vaccination. Overall this study lays the foundation for an oral vaccination platform that is simple to implement and has potential for applicability to a broad range of vaccines.
2. Materials and Methods
2.1. Pollens, chemicals, proteins and antibodies
LSs, dextran conjugated to fluorescein isothiocyanate (4000 Da and 2000 kDa), sulforhodamine (558 Da), and phosphate-citrate buffer tablets were purchased from Sigma-Aldrich (MO, USA). Pollens of chenopodium album (lambs quarters), helianthus annuus (sunflower), artemisia vulgaris (mugwort), and alnus glutinosa (alder black) were obtained from Pharmallerga (Lišov, Czech Republic). Acetone, potassium hydroxide, orthophosphoric acid, ethanol, hydrochloric acid, sodium hydroxide and tween 20 were purchased from Fisher Scientific (PA, USA). O-phenylenediamine (OPD) was obtained from Invitrogen (NY, USA). Milli-Q water with a resistance of 18.2 MΩ.cm was used in all experiments. OVA was purchased from MP Biomedicals (OH, USA). CT and CTB were purchased from Sigma-Aldrich (MO, USA). Goat anti-mouse IgG, IgG1, IgG2a, IgA, and IgE with the horseradish peroxidase (HRP) conjugate were bought from Southern Biotech (AL, USA). Texas-red labeled OVA and bovine serum albumin were purchased from Invitrogen (OR, USA).
2.2. LS treatment and characterization
LSs were chemically treated to produce intact clean spores by modifying a previously published process [24]. Briefly, 50 g of dry LSs were stirred in 300 mL of acetone under reflux overnight. Following filtration and overnight drying, they were stirred under reflux in 450 mL of 2M potassium hydroxide for 12h at 120 °C (renewed after 6h). They were then filtered and washed with hot water (5 × 300mL) and hot ethanol (5 × 300mL). After drying overnight, LSs were stirred under reflux for 7 days in 450mL of orthophosphoric acid at 180 °C. LS were filtered and washed sequentially with water (5 × 300mL), acetone (300mL), 2M HCl (300mL), 2M NaOH (300mL), water (5 × 300mL), acetone (300mL) and ethanol (300mL). Finally, they were dried at 60 °C until constant weight was achieved. The final protein concentration of the LSs was measured using nitrogen elemental analysis (PerkinElmer 2400 Series II CHNS/O Analyzer), which measures percent nitrogen in the sample. A multiplication factor of 6.25 was use to convert percent nitrogen to percent protein [25]. Scanning electron micrographs (SEM) of LSs before and after treatment were taken to confirm the removal of the biomolecules, and to determine if chemical treatment causes any damage to LSs.
2.3. Filling LSs with foreign molecules
To assess the ability to fill different molecules in to LS core, dry chemically-treated LSs were added to respective aqueous solutions of sulforhodamine, dextran conjugated to fluorescein isothiocyanate, ovalbumin conjugated to texas red, bovine serum albumin conjugated to texas red. Vacuum of about 25 inch of Hg was applied overnight by placing the aqueous suspensions of LSs in a vacuum chamber. LSs were then imaged using confocal microscopy (Ti-E inverted microscope with C2+ point scanning confocal system, Nikon Melville, NY).
2.4. Vaccine formulations
Vaccine formulations were prepared as follows in 0.3 ml PBS (dose per mouse): OVA alone: OVA(100 μg) - dose was selected based on a previous study [26]; LS1: OVA(100 μg)+LS(1 mg); LS5: OVA(100 μg)+LS(5 mg); CT1: OVA(100 μg)+CT(5 μg); CT2: OVA(5 mg)+CT(5 μg). Higher dose of OVA was used because it has been shown that CT is more effective when OVA dose is high [26]; CTB: OVA(100 μg)+CTB(50 μg). B-subunit of CT is a safer mucosal adjuvant than CT, but it is less immunogenic.
2.5. Immunizations and sample collection
BALB/c female mice 6-8 weeks-old were purchased from Charles River Laboratories (MA, USA) and were maintained at Texas Tech University Animal Care Services (TX, USA). All treatments were performed according to Texas Tech Animal Care and Use Committee (IACUC) approved procedures. All mice were fed an OVA-free diet. Mice (n=5 per group) were orally immunized by administering each mouse 0.3 ml of the vaccine formulation (OVA alone, CT1, CT2, CTB, LS1 or LS5) using a 27G blunt-tipped feeding needle. In a separate experiment, to examine the effect of neutralization of gastric acid prior to oral vaccination, mice were deprived of food and water 2 h prior to the immunizations. Next they were given an oral gavage of 0.3 ml of sodium bicarbonate (8 parts PBS + 2 parts 0.34 M sodium bicarbonate) [27, 28] to neutralize the acid in their stomach, and 30 min later were orally immunized with OVA alone (OVA-Na group) or LS5 formulation (LS5-Na group). Separate groups of mice also received OVA alone or LS5 without pre-feeding with sodium bicarbonate. In all experiments, mice were immunized on days 0 and 28. Blood was collected from animals via the retro-orbital plexus on days 0, 28 and 56. Sera were stored at -20 °C until analysis. For the determination of mucosal immune response about 10-15 fecal droppings were collected from each animal, processed in PBS, and the saline extracts were stored at -20 °C until analysis. For measuring the long term immune response, in some groups serum and fecal matter were also collected at 7 months after first oral dose (day 0).
2.6. Antibody measurement
OVA-specific antibodies in (i) pooled sera diluted from 1:50 to 1:6400, (ii) individual mouse sera at a dilution of 1:200, 1:400, or 1:25, and (iii) fecal samples at an individual dilution of 1:5 were analyzed by standard ELISA. The wells of a 96-well plate (Maxisorp-Nunc, Sigma-Aldrich, MO, USA) were coated with 50 μl of 5 μg/ml OVA solution in PBS and kept overnight at 4°C. The wells were then blocked using 100 μl of milk (5% in PBS containing 0.05% tween 20) for 1.5 h at room temperature. Pooled serum samples from each group were serially diluted (from 1:50 to 1:6400), added (50 μl) in the wells of the plate, and incubated at room temperature for 1.5 h. Secondary antibody (HRP-conjugated goat anti-mouse IgG) at a dilution of 1:4000 was then added to the wells and incubated for 1.5 h at room temperature. The plate was washed three times with PBS containing 0.05% tween 20 between each step, using the ELx405 microplate washer (BioTek, VT, USA). 50 μl OPD solution was finally added in the wells and incubated for 15 min at room temperature. The reaction was stopped by adding 50 μl sulfuric acid to each well. The plate was read at 490 nm using a microplate reader (SpectraMax Plus384, Molecular Devices LLC, Sunnyvale, CA). Optical Density (OD) at each dilution was determined to obtain a titration curve.
Using the titration curve a suitable dilution was selected and samples from individual animals for all groups were then diluted to this value, and the OD for individual animals was determined. Pooled serum samples at the same dilution were used to determine the IgG isotypes in serum. Similarly, serum samples from individual animals were diluted at 1:25 and analyzed by ELISA to determine anti-OVA IgE. Fecal samples at an individual dilution of 1:5 were analyzed by ELISA to determine anti-OVA IgA using the same procedure as described above.
2.7. Evaluation of the ability of LSs to cross intestinal epithelium
Mice (n=3) were fed 5 mg of treated LSs, euthanized 24 h later, their intestines were isolated, cleaned, cut into small pieces, further cut longitudinally to expose the intestinal epithelium, laid flat between a cover slip and glass slide with the cover slip touching the epithelial surface, and visualized using confocal microscopy. LSs are naturally fluorescent over a broad emissionspectrum but were better visible under TRITC (570 nm) filter. Verification of the confocal microscopy technique to identify LSs and discriminate their presence on epithelial or serosal surfaces is presented in Supplementary Information-Materials and Methods.
2.8. Statistical analysis
All statistical analysis was performed using GraphPad Prism for Windows version 6.0 (GraphPad Software, Inc., CA, USA). Comparison of antibody levels between groups of mice was performed with a two-way analysis of variance (ANOVA) and a Tukey test of multiple comparisons at a value of p < 0.05 for statistical significance.
3. Results and Discussion
3.1. Creating intact LSs with a clean core
PGs are natural microparticles that plants use for safe transportation of their genetic material, often across large geographical distances with fluctuating climatic conditions. To perform this function, the pollen wall typically comprises of two main layers, the outer exine and the inner intine. Exine is composed of a tough biopolymer called sporopollenin, which is resistant to acetolysis, high temperature and decay, while intine is mostly made of cellulose and pectin [29]. Exine, is highly permeable [30] and the pores of the exine wall, which can be up to 40 nm wide, allow for the penetration of different substances into the central hollow core of pollens [21, 31, 32].
We postulated that the naturally-occurring pores in the pollen wall could be used to allow chemicals to penetrate the pollen core to facilitate removal of the intine layer, and biomolecules such as cellular organelle, proteins and lipids, and then could subsequently be used to load vaccine molecules in to the clean pollen core for oral delivery (Fig 1). PG size and shape varies from one plant species to another as can be seen from the scanning electron micrographs of PGs from four different plant species (Fig 2A-2D). However, within a plant species PGs possess size and morphological homogeneity as exemplified by LSs (Fig 2E). In this study we selected LSs as the model system and sequentially treated them with acetone, potassium hydroxide, and phosphoric acid; to dissolve lipids and hydrophobic molecules, proteins and cellulosic intine layer, respectively. By comparing unprocessed and processed LSs (Fig 2E vs 2F; 2G vs 2H; 2I vs 2J) it can be seen that the chemical treatment does not damage the LSs. Furthermore, comparing Fig 3A to 3B it can be qualitatively seen that the chemical treatment removes native biomolecules from the LS core. We also quantified the final protein concentration in the treated pollen to be 0.53% (w/w of pollen) as compared to 7.3% in untreated raw LSs, resulting in about 90% removal of proteins. It is important to note that protein concentration was determined by assuming that the entire amount of measured nitrogen originates from proteins, however, nitrogen may very well be part of the sporopollenin chemical structure. Thus, it is likely that our calculation underestimates the extent of protein removal, and in fact even lower amount of protein is expected to remain.
Fig 1. Schematic illustrating concept of pollens as a system for oral vaccination.

(A) A raw pollen grain with a naturally porous pollen wall. The pollen wall is often coated with a sticky material called pollen kit, which can contain allergens. PW: pollen wall, PK: pollen kit, CO&B: cellular organelle and biomolecules. (B) A chemically processed pollen grain whose native biomolecules, cellular organelle, pollen kit, and intine have been removed to produce an intact pollen with a clean core. (C) Pollen grain whose core is filled with a foreign molecule via the natural pores in the pollen wall, enabling use of pollens as drug and vaccine carriers.
Fig 2. Unprocessed and chemically processed pollens.
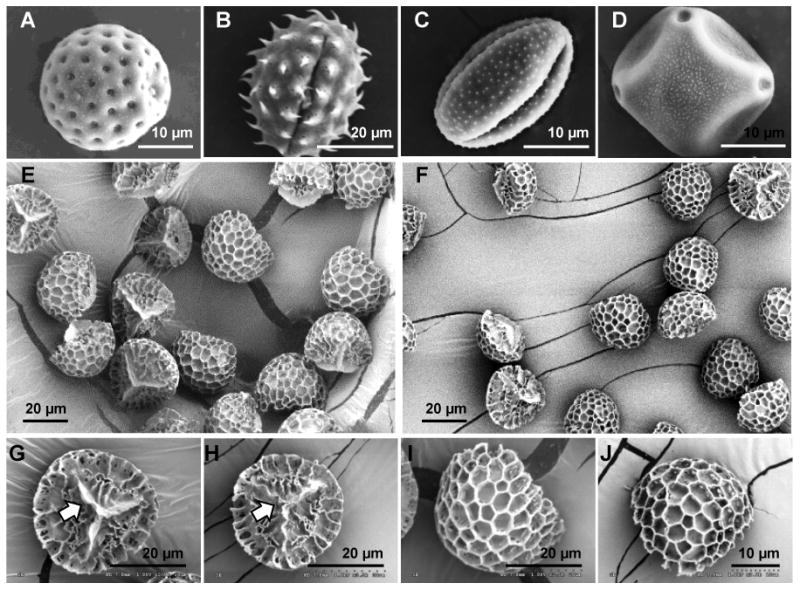
Scanning electron micrographs of unprocessed raw pollens showing diversity in shape, size and morphology: (A) Chenopodium album (lambs quarters) pollen grain, (B) Helianthus annuus (sunflower) pollen grain, (C) Artemisia vulgaris (mugwort) pollen grain, (D) Alnus glutinosa (alder black) pollen grain, and (E) Lycopodium clavatum (clubmoss) spore. Scanning electron micrographs of lycopodium spores before and after chemical processing: (E:before, F:after) low magnification images showing en bloc comparison, (G:before, H:after) higher magnification comparison of proximal face of the spore and its trilete scar (indicated by arrow), and (I:before, J:after) higher magnification comparison of distal face of the spore and its decorative ornamentation. No visible damage to the trilete scar or the surface ornamentation is seen after chemical processing.
Fig 3. Interior of lycopodium spore at different stages of processing.
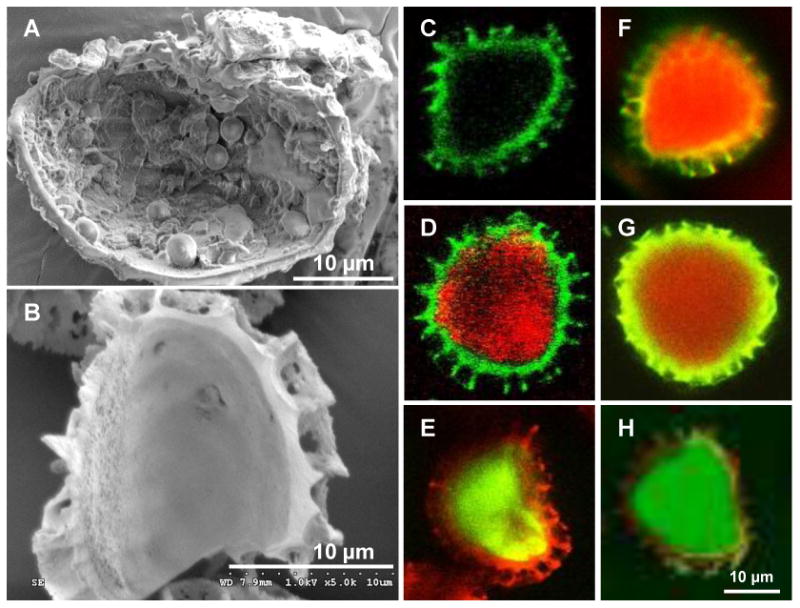
Scanning electron micrographs of lycopodium spores manually cracked (A) before processing showing cellular organelle and biomolecules in the core, and (B) after chemical processing showing a clean core. Confocal micrographs of chemically-processed lycopodium spores that are (C) empty, (D) filled with sulforhodamine (558 Da), (E) filled with dextran conjugated to fluorescein isothiocyanate (4000 Da), (F) filled with ovalbumin conjugated to texas red (45,000 Da), (G) filled with bovine serum albumin conjugated to texas red (67,000 Da), and (H) filled with dextran conjugated to fluorescein isothiocyanate (2,000,000 Da).
We expect chemically-treated clean LSs to be non-allergenic because native proteins and lipids found in PGs, which are the root cause of pollen allergies [33] are removed after chemical treatment. Despite this, it is important to consider if treated pollens might still cause allergies in patients known to have pollen allergies. In this regard, some studies [34, 35] have shown that even raw-unprocessed pollen when fed to patients of pollen hay fever did not present any systemic or gastrointestinal clinical symptoms. More so, oral delivery of pollen extracts, which contains allergens, to pollen allergy patients [36] did not result in oral hyposensitization, suggesting safety of pollens even in allergic subjects. Considerable anecdotal evidence also exists on the safety of oral uptake of PGs in their natural raw form without any chemical treatment. For example bee pollen and other pollen species are often used as a health supplement [37]. PGs and LSs are also part of traditional medicine across the world including India, China, American Indians, Turkish folk medicine, and Papua New Guniea to name a few [38]. While more thorough investigation is needed, this evidence suggests that chemically-treated pollens should be clinically acceptable and safe for oral ingestion without causing any allergies.
3.2. Filling the LS core with foreign molecules
After creating clean empty LSs we utilized the pores in their walls to fill different molecules in to their core by incubating LSs in aqueous solutions of fluorescently-labeled compounds and applying mild vacuum to facilitate removal of air from the LS core causing an ingress of dissolved molecules from the outer aqueous environment into the LS core. To verify encapsulation, LSs were imaged using a confocal microscope. LSs are naturally fluorescent over a wide range of emission wavelengths from blue to red. Using appropriate combination of excitation-emission filters both the pollen shell and the inner core was visualized. Treated but empty shell can be seen in Fig 3C, and they possess a non-fluorescent interior. Figs 3D, 3E, 3F, 3G, and 3H show treated LSs that are filled with sulforhodamine (558 Da), dextran (4 kDa), OVA (45 kDa), bovine serum albumin (67 kDa), and dextran (2000 kDa), respectively. This result is consistent with a recent study that successfully encapsulated oils, fats and proteins such as β-galactosidase (540 kDa) and horseradish peroxidase (100 kDa) in LSs [23]. Thus, a wide range of molecules can be filled in to LS core demonstrating the flexibility of the platform for vaccine delivery.
For the purpose of vaccination, OVA was used as an antigen. It is important to note that each oral dose comprised of 0.3 ml of OVA (100 μg) solution with 5 mg LSs suspended in it. Not all of the OVA was encapsulated in LSs because the OVA solution was in excess; it was present in the hollow core of LSs and their exterior. The excess solution helped to maintain equilibrium between OVA inside and outside the LSs, and kept them filled with OVA at all times.
3.3. Immune response after oral vaccination with LSs
3.3.1. Systemic immune response
To evaluate the potential of LSs for oral vaccination, we immunized Balb/c mice with OVA-filled LS formulations. CT and its B subunit (CTB) are well known adjuvants for oral immunization, and were used as positive controls, while OVA alone served as the negative control. After two doses (day 0 and 28), the antibody titration curve of day 56 pooled serum samples (Fig 4A) shows that OVA by itself is poorly immunogenic while addition of 5 mg LSs (LS5 group) to the formulation stimulated the highest antibody levels at all dilutions. Based on the dilution curve, a suitable dilution (1:200) was selected to analyze OVA-specific immunoglobulin G (IgG) response for individual mice to assess intergroup variability. From Fig 4B it can be seen that the LS5 group elicits significantly higher anti-OVA IgG response compared to use of CT and CTB (p<0.0001). Although, CT1, CT2, CTB, and LS1 groups produced an increase in anti-OVA IgG response compared to OVA alone, it was not statistically significant (p>0.05). A dose-dependent effect of LSs was seen wherein 5 mg LSs in the formulation stimulated a significantly higher (p=0.0005) anti-OVA IgG response in serum compared to 1 mg LSs (LS1 formulation).
Fig 4. Systemic and mucosal immune response against ovalbumin (OVA) with lycopodium spores as the oral vaccination system.

Groups of mice (n=5 per group) were orally vaccinated on days 0 and 28 with OVA and different adjuvants. Anti-OVA IgG and IgA were determined in serum and fecal droppings, respectively. (A) Titration curve of pooled day 56 mouse serum demonstrating the high anti-OVA IgG stimulated with 5 mg LSs (LS5 group) in comparison to all other formulations. (B) Anti-OVA IgG in individual mouse serum (diluted 1:200) at different time point (days 0, 28 and 56). LS5 group shows significantly higher antibody response compared to all groups. Each symbol represents a mouse, the error bar represents standard deviation, and the horizontal line represents the mean. (C) Anti-OVA IgG subtypes in day 56 pooled serum (dilution 1:200). Error bars indicate standard deviation. (D) Anti-OVA IgA in individual mouse fecal droppings (diluted 1:5) at different time points (days 0, 28 and 56). LS5 group shows significantly higher antibody response compared to OVA alone. Each symbol represents a mouse, the error bar represents standard deviation, and the horizontal line represents the mean. (E) Anti-OVA IgE in day 56 individual mouse serum (diluted 1:25). LS5 group does not show any statistically significant difference as compared to OVA and CT2 groups. ns: not statistically significant (p>0.05).
We further determined Th1 and Th2 bias induced by the vaccine formulations by measuring serum anti-OVA IgG1 and IgG2a antibody subtypes. LS5 formulation was seen to induce high anti-OVA IgG1 antibody levels compared to IgG2a subtype (Fig 4C), indicating a Th2-biased immune response. The same trend was exhibited by CT1, CT2 and CTB formulations, which is in agreement with previous studies reporting higher Th2 (IgG1) response than Th1 (IgG2a) response [39] through use of these adjuvants.
We also evaluated whether the LS5 formulation stimulates IgE antibody. Day 56 serum samples from individual animals at 1:25 dilution were analyzed through ELISA. We compared the LS5 formulation with, OVA alone and OVA with CT (CT2 formulation) as the negative control and positive control groups, respectively. Analysis of serum samples with ELISA showed low anti-OVA IgE for all the groups (Fig 4E), and no statistically significant difference was observed between LS5, CT2 and OVA groups.
3.3.2. Mucosal immune response
Mucosal surfaces act as portals of entry for pathogens. The human body produces immunoglobulin A (IgA) at these surfaces, which can bind invading pathogens and can prevent them from entering systemic circulation. Hence, it is beneficial for a vaccine to produce a mucosal immune response in addition to a systemic immune response [1, 2]. Thus, we investigated stimulation of anti-OVA IgA in intestinal secretions. We compared the LS5 formulation (since it induced maximum serum IgG stimulation), OVA alone or with CT as adjuvant (CT2 formulation) as the negative and positive control groups, respectively. Analysis of saline extracts of fecal droppings through ELISA showed that (Fig 4D) on day 28 while the LS5 group exhibited a small increase in anti-OVA IgA, it was not statistically significant (p > 0.05). However, following a booster dose on day 28, the LS5 group demonstrated a significant increase in anti-OVA IgA compared to CT2 formulation (p = 0.0411) and OVA alone (p = 0.0013).
3.4. Effect of neutralization of gastric acid on anti-OVA antibody response
Degradation of an antigen in the hostile gastric environment is a common problem associated with oral vaccines. One way to deal with this problem is to neutralize the gastric acid to create a favorable environment in which the antigen can survive [27, 28]. The commercial rotavirus vaccine employs this strategy to enhance vaccine efficacy [40]. We thus sought to determine whether neutralizing the stomach acid prior to oral vaccination with LSs might further enhance the immune response. Since LS5 formulation was found to be the most successful formulation in the previous experiment, we focused on it to evaluate the gastric neutralization effect. Based on an ELISA performed on day 56 pooled sera (Fig 5A) it was seen that pre-neutralization of gastric acid did not affect serum anti-OVA IgG levels for both OVA alone and LS5 formulations. This result was further confirmed through analysis of OVA-specific IgGs in individual mice serum (Fig 5B), wherein again no significant difference (p>0.05) was observed between LS5 and LS5-Na formulations. Fig 5B also shows that LS5 formulation (with and without gastric neutralization) produces a significantly (p<0.0001) higher anti-OVA IgG response compared to OVA alone (with and without gastric neutralization) reaffirming the effectiveness of LSs for oral vaccination. As seen in Fig 5C, an elevated IgG1 response compared to IgG2a was seen for both LS5 and LS5-Na groups, indicating that pre-neutralization of gastric acid does not change the character of LS-induced immune response.
Fig 5. Effect of gastric acid neutralization on stimulation of ovalbumin-specific serum IgGs.

Groups of mice (n=5 per group) were fed sodium bicarbonate buffer before orally administering ovalbumin (OVA)-based vaccine formulations. Vaccinations were performed on days 0 and 28, and serum samples were analyzed to determine anti-OVA IgG. (A) Titration curve of day 56 pooled mouse serum demonstrating the high anti-OVA IgG stimulation from groups receiving OVA with 5 mg LSs either with (LS5-Na group) or without (LS5 group) sodium bicarbonate pre-feed. No effect of gastric acid neutralization is seen. (B) Anti-OVA IgG in individual mouse serum at days 0 and 56 at a dilution of 1:400. Both LS5 and LS5-Na groups show significantly higher anti-OVA IgG response compared to OVA alone administered either with (OVA-Na group) or without (OVA group) sodium bicarbonate pre-feed. No significant difference is observed between LS5 and LS5-Na groups (p>0.05). Each symbol represents a mouse, the error bar represents standard deviation, and the horizontal line represents the mean. ns: not statistically significant. (C) Anti-OVA IgG subtypes in day 56 pooled serum samples (diluted 1:400). Error bars indicate standard deviation.
3.5. Duration of anti-OVA serum and mucosal antibodies
Persisting antigen-specific antibodies are needed to maintain long-term humoral immunity [41]. Commercial vaccines induce high titers of antigen-specific antibodies, which either persist over a long time, or periodical booster doses must be administered to maintain them at high levels for sustained protection. For example, hepatitis B vaccine given during childhood is expected to provide life-long immunity [42], while the tetanus vaccine requires a booster dose every 10 years [43]. Thus we sought to determine if LSs could induce long term circulating and mucosal antibodies. We analyzed the serum and fecal droppings of animals at month 7 following two oral vaccine doses at day 0 and day 28. From Fig 6A it can be seen that anti-OVA serum IgG and fecal IgA antibodies could be measured in the animals even seven months after the first vaccine dose at statistically significant levels (IgG: p=0.0263; IgA: p=0.0018). Furthermore, no change in IgG1 versus IgG2a bias was observed (Fig 6B) suggesting no long term change in the character of the immune response. Overall, this shows that LSs have the potential to induce long-term systemic and mucosal immune responses.
Fig 6. Long term systemic and mucosal immune response.

Groups of mice (n=5 per group) were orally vaccinated on days 0 and 28 with OVA, with or without 5 mg lycopodium spores. Serum and fecal droppings were collected seven months later and analyzed to determine anti-OVA IgG and IgA. (A) Anti-OVA IgG in individual mouse serum (diluted 1:400), and anti-OVA IgA in individual mouse fecal droppings (diluted 1:5). Use of 5 mg LSs (LS5 group) results in elevated IgG and IgA levels even seven months after vaccination (IgG: p=0.0263; IgA: p=0.0018). Each symbol represents a mouse, the error bar represents standard deviation, and the horizontal bar represents the mean. (B) Anti-OVA IgG subtypes in month seven pooled serum samples (diluted 1:400). Error bars indicate standard deviation.
3.6. Penetration of LSs across the intestinal epithelium
To better understand how LSs stimulate systemic and mucosal immune responses, we investigated their ability to penetrate the mucosal epithelium. Mice (n=3) were fed LSs, and 24 h later parts of their intestine (after washing) were imaged using a confocal microscope. Fig 7 shows that LSs can translocate in to the intestinal wall, suggesting that LSs could perhaps carry the antigen enabling its interaction with the local population of immune cells to stimulate an antigen-specific immune response. LSs due to their particulate form are also expected to stimulate immune cells. Similar translocation of LSs was observed in the other two mice (see supplementary Fig S3). LSs are approximately 25 μm in size. Previous studies investigating oral transport of particles have determined that particles approximately 2 μm or less are appropriately-sized for uptake via M-cells of the Peyer's patches [44]. However, the same study also reported uptake of 20 μm diameter particles, although with reduced frequency. Furthermore, others have reported uptake of larger particles across the intestinal wall [45], including that of LSs in human subjects [18]. Uptake of larger particles (>10 μm) is thought to occur via the villous epithelium as opposed to M-cells [46], and this phenomenon has been termed ‘persorption’ [47]. It is likely that LSs penetrate the villous epithelium via persorption, a phenomenon not yet clearly understood. Although, the current study demonstrates ability of LSs to penetrate the intestinal wall, additional questions remain to be answered, for example, what is the mechanism by which LSs stimulate the immune response, how efficient is the co-transport of antigens with LSs, whether the vaccination approach translates successfully to other sub-unit vaccines, and whether other pollens exhibit a similar adjuvant effect. Our laboratory is performing experiments to answer these questions.
Fig 7. Penetration of LSs across intestinal epithelium in to the intestinal wall.
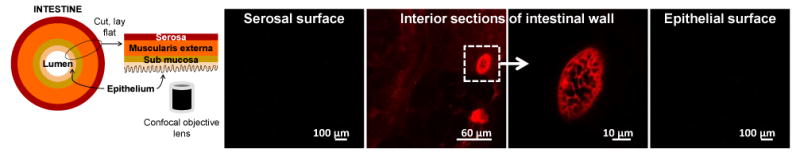
Mice (n=3) were fed 5 mg LSs, and 24 h later their intestinal tissues were imaged using a confocal microscope. Representative confocal micrographs at serosal surface, interior part of intestinal wall, and epithelial surface are shown. Presence of LSs in inner parts of the wall demonstrates that LSs can cross the epithelial surface and enter the intestinal wall.
3.7. Discussion on overall potential of LSs for oral vaccination
In this study we show that LSs elicit anti-OVA antibodies in serum (IgG) and intestinal secretions (IgA) at levels significantly higher than use of CT as an adjuvant, and these antibodies persisted up to seven months. LSs also induced anti-OVA IgE levels, which were not significantly different from those obtained by OVA alone or OVA (5 mg) + CT (5 μg). These findings are significant because they suggests that LSs could be a viable alternative to CT for oral vaccine development. CT is considered a highly effective oral adjuvant, albeit it's inherent toxicity prevents it from being used in humans [6]. LSs on the other hand are more effective and potentially safer. Clearly, use of 1 mg LSs can match the immune response produced by CT2 (5 mg of OVA + 5 μg of CT) with just 1/50th of the OVA dose (100 μg). By increasing the LS dose to 5 mg, the immune response was further significantly improved. LSs are relatively inexpensive, ∼ $0.2/g (Sigma Aldrich), and could be even cheaper if obtained in bulk. To formulate vaccine antigens with LSs, organic solvents or other processing conditions that could damage the vaccine are also not required. Instead just mild application of vacuum is needed. This LS-filling step could be easily integrated into the existing vaccine manufacturing process without significant expenditure or re-engineering for each vaccine. Because LSs can be filled with a broad range of molecules they offer a modular approach of vaccine delivery via the oral route, much like a hypodermic needle, which offers complete flexibility over the type of vaccine to be injected. Because, hypodermic needles, syringes or additional devices would not be required anymore, the overall cost may be even lower than current vaccine prices. For developing nations this would also help in reducing the extraordinary burden they face for safe disposal of used-needle medical waste [48]. More broadly, LS-based oral vaccination technology could offer a child-friendly and painless formulation that could potentially be available over the counter for self-administration.
4. Conclusion
We propose the novel concept that PGs can be engineered for oral vaccination. LSs were chemically cleaned to produce intact empty spores, which could be filled with foreign molecules up to 2000 kDa in size. LSs filled with OVA as a model antigen were used for oral immunization. LS5 formulation with 5 mg spores enabled stimulation of a strong anti-OVA IgG (systemic) and IgA (mucosal) antibody response, which was higher than that produced by use of CT and its B subunit - CTB as oral mucosal adjuvants. The amounts of anti-OVA IgG and IgA produced by the LS5 formulation were not affected by pre-neutralization of the gastric acid at the time of immunization. Furthermore, OVA-specific IgG and IgA antibodies were seen to persist in the mice for a prolonged period of time up to seven months. Confocal microscopy of intact intestinal tissues demonstrated the presence of LSs in the intestinal wall suggesting that translocation of LSs across the intestinal epithelium may be involved in stimulation of the immune system. Overall this study demonstrates the potential of LSs for development of oral vaccines, which could have wide applicability since a broad spectrum of antigens could potentially be loaded in to LSs for oral vaccination.
Supplementary Material
Acknowledgments
Research reported in this publication was supported in parts by the Eunice Kennedy Shriver National Institute of Child Health & Human Development of the National Institutes of Health (NIH) under Award Number DP2HD075691 to HSG, and Defense Advanced Research Projects Agency (DARPA) - Young Faculty Award under grant number N66001-12-1-4251 to HSG.
Footnotes
Conflict of Interest Statement: A patent has been filed by the authors regarding use of pollen grains for oral vaccination through Texas Tech University. This potential conflict of interest has been disclosed and is managed by Texas Tech University.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.De Magistris MT. Mucosal delivery of vaccine antigens and its advantages in pediatrics. Adv Drug Deliv Rev. 2006;58:52–67. doi: 10.1016/j.addr.2006.01.002. [DOI] [PubMed] [Google Scholar]
- 2.Neutra MR, Kozlowski PA. Mucosal vaccines: the promise and the challenge. Nat Rev Immunol. 2006;6:148–158. doi: 10.1038/nri1777. [DOI] [PubMed] [Google Scholar]
- 3.Acheson DW, Luccioli S. Microbial-gut interactions in health and disease. Mucosal immune responses. Best Pract Res Clin Gastroenterol. 2004;18:387–404. doi: 10.1016/j.bpg.2003.11.002. [DOI] [PubMed] [Google Scholar]
- 4.Yu LX, Lipka E, Crison JR, Amidon GL. Transport approaches to the biopharmaceutical design of oral drug delivery systems: prediction of intestinal absorption. Adv Drug Deliv Rev. 1996;19:359–376. doi: 10.1016/0169-409x(96)00009-9. [DOI] [PubMed] [Google Scholar]
- 5.Kersten G, Hirschberg H. Antigen delivery systems. Expert Rev Vaccines. 2004;3:453–462. doi: 10.1586/14760584.3.4.453. [DOI] [PubMed] [Google Scholar]
- 6.Holmgren J, Czerkinsky C. Mucosal immunity and vaccines. Nat Med. 2005;11:S45–53. doi: 10.1038/nm1213. [DOI] [PubMed] [Google Scholar]
- 7.Clark MA, Jepson MA, Hirst BH. Exploiting M cells for drug and vaccine delivery. Adv Drug Deliv Rev. 2001;50:81–106. doi: 10.1016/s0169-409x(01)00149-1. [DOI] [PubMed] [Google Scholar]
- 8.Holmgren J, Harandi AM, Czerkinsky C. Mucosal adjuvants and anti-infection and anti-immunopathology vaccines based on cholera toxin, cholera toxin B subunit and CpG DNA. Expert Rev Vaccines. 2003;2:205–217. doi: 10.1586/14760584.2.2.205. [DOI] [PubMed] [Google Scholar]
- 9.Rappuoli R, Pizza M, Douce G, Dougan G. Structure and mucosal adjuvanticity of cholera and Escherichia coli heat-labile enterotoxins. Immunol Today. 1999;20:493–500. doi: 10.1016/s0167-5699(99)01523-6. [DOI] [PubMed] [Google Scholar]
- 10.Levine MM, Kaper JB, Black RE, Clements ML. New knowledge on pathogenesis of bacterial enteric infections as applied to vaccine development. Microbiol Rev. 1983;47:510–550. doi: 10.1128/mr.47.4.510-550.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Woodrow KA, Bennett KM, Lo DD. Mucosal vaccine design and delivery. Annu Rev Biomed Eng. 2012;14:17–46. doi: 10.1146/annurev-bioeng-071811-150054. [DOI] [PubMed] [Google Scholar]
- 12.Schwendeman SP. Recent advances in the stabilization of proteins encapsulated in injectable PLGA delivery systems. Crit Rev Ther Drug Carrier Syst. 2002;19:73–98. doi: 10.1615/critrevtherdrugcarriersyst.v19.i1.20. [DOI] [PubMed] [Google Scholar]
- 13.Nochi T, Takagi H, Yuki Y, Yang L, Masumura T, Mejima M, Nakanishi U, Matsumura A, Uozumi A, Hiroi T, Morita S, Tanaka K, Takaiwa F, Kiyono H. Rice-based mucosal vaccine as a global strategy for cold-chain- and needle-free vaccination. Proc Natl Acad Sci USA. 2007;104:10986–10991. doi: 10.1073/pnas.0703766104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Pniewski T. The twenty-year story of a plant-based vaccine against hepatitis B: stagnation or promising prospects? Int J Mol Sci. 2013;14:1978–1998. doi: 10.3390/ijms14011978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Shivanna KR. Pollen Biology and Biotechnology. Science Publishers, Enfield; NH, USA: 2003. [Google Scholar]
- 16.Roulston TH, Cane JH. Pollen nutritional content and digestibility for animals. Pl Syst Evol. 2000;222:187–209. [Google Scholar]
- 17.Barrier S. Physical and chemical properties of sporopollenin exine particles. The University of Hull; Hull: 2008. [Google Scholar]
- 18.Jorde W, Linskens HF. Zur persorption von pollen und spoken durch die intake darmschleimhaut. Acta Allergol. 1974;29:165–175. [PubMed] [Google Scholar]
- 19.Lorch M, Thomasson MJ, Diego-Taboada A, Barrier S, Atkin SL, Mackenzie G, Archibald SJ. MRI contrast agent delivery using spore capsules: controlled release in blood plasma. Chem Commun. 2009:6442–6444. doi: 10.1039/b909551a. [DOI] [PubMed] [Google Scholar]
- 20.Barrier S, Rigby AS, Diego-Taboada A, Thomasson MJ, Mackenzie G, Atkin SL. Sporopollenin exines: A novel natural taste masking material. LWT-Food Sci Technol. 2010;43:73–76. [Google Scholar]
- 21.Paunov VN, Mackenzie G, Stoyanov SD. Sporopollenin micro-reactors for in-situ preparation, encapsulation and targeted delivery of active components. J Mater Chem. 2007;17:609–612. [Google Scholar]
- 22.Wakil A, Mackenzie G, Diego-Taboada A, Bell J, Atkin S. Enhanced Bioavailability of Eicosapentaenoic Acid from Fish Oil After Encapsulation Within Plant Spore Exines as Microcapsules. Lipids. 2010;45:645–649. doi: 10.1007/s11745-010-3427-y. [DOI] [PubMed] [Google Scholar]
- 23.Barrier S, Diego-Taboada A, Thomasson MJ, Madden L, Pointon JC, Wadhawan JD, Beckett ST, Atkin SL, Mackenzie G. Viability of plant spore exine capsules for microencapsulation. J Mater Chem. 2011;21:975–981. [Google Scholar]
- 24.Atkin SL, Barrier S, Cui Z, Fletcher PDI, Mackenzie G, Panel V, Sol V, Zhang X. UV and visible light screening by individual sporopollenin exines derived from Lycopodium clavatum (club moss) and Ambrosia trifida (giant ragweed) J Photochem Photobiol B. 2011;102:209–217. doi: 10.1016/j.jphotobiol.2010.12.005. [DOI] [PubMed] [Google Scholar]
- 25.B ST, Atkin Stephen Lawrence, Mackenzie Grahame. Dosage form. USA: 2005. p. 9. [Google Scholar]
- 26.Lauterslager TG, Stok W, Hilgers LA. Improvement of the systemic prime/oral boost strategy for systemic and local responses. Vaccine. 2003;21:1391–1399. doi: 10.1016/s0264-410x(02)00687-4. [DOI] [PubMed] [Google Scholar]
- 27.Vajdy M, Lycke NY. Cholera toxin adjuvant promotes long-term immunological memory in the gut mucosa to unrelated immunogens after oral immunization. Immunology. 1992;75:488–492. [PMC free article] [PubMed] [Google Scholar]
- 28.Marinaro M, Staats HF, Hiroi T, Jackson RJ, Coste M, Boyaka PN, Okahashi N, Yamamoto M, Kiyono H, Bluethmann H, Fujihashi K, McGhee JR. Mucosal adjuvant effect of cholera toxin in mice results from induction of T helper 2 (Th2) cells and IL-4. J Immunol. 1995;155:4621–4629. [PubMed] [Google Scholar]
- 29.Hesse M, Halbritter H, Weber M, Buchner R, Frosch-Radivo A, Ulrich S. Pollen terminology : an illustrated handbook. Springer, Wien; New York: 2009. [Google Scholar]
- 30.Hose E, Clarkson DT, Steudle E, Schreiber L, Hartung W. The exodermis: a variable apoplastic barrier. J Exp Bot. 2001;52:2245–2264. doi: 10.1093/jexbot/52.365.2245. [DOI] [PubMed] [Google Scholar]
- 31.Rowley JR, Skvarla JJ, El-Ghazaly G. Transfer of material through the microspore exine from the loculus into the cytoplasm. Can J Bot. 2003;81:1070–1082. [Google Scholar]
- 32.Wittborn J, Rao KV, El-Ghazaly G, Rowley JR. Nanoscale Similarities in the Substructure of the Exines of Fagus pollen grains and lycopodium spores. Ann Bot. 1998;82:141–145. [Google Scholar]
- 33.Larche M, Akdis CA, Valenta R. Immunological mechanisms of allergen-specific immunotherapy. Nat Rev Immunol. 2006;6:761–771. doi: 10.1038/nri1934. [DOI] [PubMed] [Google Scholar]
- 34.Taudorf E, Laursen LC, Djurup R, Kappelgaard E, Pedersen CT, Søsborg M, Wilkinson P, Weeke B. Oral Administration of Grass Pollen to Hay Fever Patients. Allergy. 1985;40:321–335. doi: 10.1111/j.1398-9995.1985.tb00243.x. [DOI] [PubMed] [Google Scholar]
- 35.Taudorf E, Weeke B. Orally Administered Grass Pollen. Allergy. 1983;38:561–564. doi: 10.1111/j.1398-9995.1983.tb04140.x. [DOI] [PubMed] [Google Scholar]
- 36.Urbanek R, Bürgelin KH, Kahle S, Kuhn W, Wahn U. Oral immunotherapy with grass pollen in enterosoluble capsules. Eur J Pediatr. 1990;149:545–550. doi: 10.1007/BF01957689. [DOI] [PubMed] [Google Scholar]
- 37.Linskens HF, Jorde W. Pollen as Food and Medicine: A Review. Econ Bot. 1997;51:78–86. [Google Scholar]
- 38.Orhan I, Kupeli E, Sener B, Yesilada E. Appraisal of anti-inflammatory potential of the clubmoss, Lycopodium clavatum L. J Ethnopharmacol. 2007;109:146–150. doi: 10.1016/j.jep.2006.07.018. [DOI] [PubMed] [Google Scholar]
- 39.Tochikubo K, Isaka M, Yasuda Y, Kozuka S, Matano K, Miura Y, Taniguchi T. Recombinant cholera toxin B subunit acts as an adjuvant for the mucosal and systemic responses of mice to mucosally co-administered bovine serum albumin. Vaccine. 1998;16:150–155. doi: 10.1016/s0264-410x(97)00194-1. [DOI] [PubMed] [Google Scholar]
- 40.Vesikari T. Rotavirus vaccination: a concise review. Clin Microbiol Infect. 2012;18(Suppl 5):57–63. doi: 10.1111/j.1469-0691.2012.03981.x. [DOI] [PubMed] [Google Scholar]
- 41.Amanna IJ, Carlson NE, Slifka MK. Duration of Humoral Immunity to Common Viral and Vaccine Antigens. N Engl J Med. 2007;357:1903–1915. doi: 10.1056/NEJMoa066092. [DOI] [PubMed] [Google Scholar]
- 42.Banatvala J, Van Damme P, Oehen S. Lifelong protection against hepatitis B: the role of vaccine immunogenicity in immune memory. Vaccine. 2000;19:877–885. doi: 10.1016/s0264-410x(00)00224-3. [DOI] [PubMed] [Google Scholar]
- 43.Cook TM, Protheroe RT, Handel JM. Tetanus: a review of the literature. Br J Anaesth. 2001;87:477–487. doi: 10.1093/bja/87.3.477. [DOI] [PubMed] [Google Scholar]
- 44.Carr KE, Hazzard RA, Reid S, Hodges GM. The effect of size on uptake of orally administered latex microparticles in the small intestine and transport to mesenteric lymph nodes. Pharm Res. 1996;13:1205–1209. doi: 10.1023/a:1016064320334. [DOI] [PubMed] [Google Scholar]
- 45.Volkheimer G. Passage of Particles through the Wall of of the Gastrointestinal Tract. Environ Health Perspect. 1974;9:215–225. doi: 10.1289/ehp.749215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Smyth SH, Feldhaus S, Schumacher U, Carr KE. Uptake of inert microparticles in normal and immune deficient mice. Int J Pharm. 2008;346:109–118. doi: 10.1016/j.ijpharm.2007.06.049. [DOI] [PubMed] [Google Scholar]
- 47.Volkheimer G, Schulz FH. The phenomenon of persorption. Digestion. 1968;1:213–218. doi: 10.1159/000196856. [DOI] [PubMed] [Google Scholar]
- 48.Hutin YJF, Hauri AM, Armstrong GL. Use of injections in healthcare settings worldwide, 2000: literature review and regional estimates. BMJ. 2003;327:1075. doi: 10.1136/bmj.327.7423.1075. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.