

Abstract
We consider a multistage cancer model in which cells are arranged in a d-dimensional integer lattice. Starting with all wild-type cells, we prove results about the distribution of the first time when two neutral mutations have accumulated in some cell in dimensions d ≥ 2, extending work done by Komarova [12] for d = 1.
Keywords: biased voter model, stochastic tunneling, cancer progression
1 Introduction
The accumulation of mutations is important not only for cancer initiation, progression, and metastasis, but also for the emergence of acquired resistance against chemotherapeutics, radiation therapy, or targeted drugs. For this reason there is a large and growing literature on the waiting time τk until some cell has acquired k prespecified mutations. In all the models we consider, type i individuals mutate to type (i + 1) at rate ui+1. The dynamics considered have most often been studied in multi-type Moran models with a homogeneously mixing population of constant size. Here we will concentrate on how results change when one considers a spatial Moran models, and as is the case for much earlier work we will concentrate on the behavior of τ2.
We suppose that cells of type 0 and type 1 have relative fitness 1 and λ. Since we will only consider the waiting time for the first type 2, the relative fitness of type 2’s is not important. In this work we will consider situation in which λ is so close to 1 that the mutations are essentially neutral. For cancer applications, this is a restrictive assumption, and it will be removed in the companion paper (part II) by Durrett, Foo and Leder [6]. However, the current result applies to the important case of tumor suppressor genes. In that case, when both copies of the gene are inactivated trouble develops, but while there is one working copy the cell can function normally.
We begin by recalling results for the Moran model in a homogeneously mixing population of size N. Here and in what follows the mutation rates ui and selection coefficient λ depend on N, even though this is not indicated in the notation, and we write aN ≪ bN if aN/bN → 0 as N → ∞. The next result made its first appearance on page 16230 of Nowak et al [17]. Since then it has appeared in print a number of times: [14], [18], [10], [11], and in Nowak’s excellent book [16] on Evolutionary Dynamics.
Theorem 1
In the neutral case of the Moran model, λ = 1, if we assume that
| (1) |
and let u1, u2 → 0 then we have
The same conclusion holds if .
Durrett and Schmidt [7] applied these ideas to study regulatory sequence evolution and to expose flaws in Michael Behe’s arguments for intelligent design. Durrett, Schmidt, and Schweinsberg [8], see also Schweinsberg [19], generalized this result to cover τk.
The conditions in the result may look mysterious but they can be derived by simple reasoning. Here and throughout the paper and f(u) ~ g(u) means f(u)/g(u) → 1 as u → 0. Suppose first that λ = 1.
-
(A1)
If we start the Moran model with k ≪ N type 1’s and the rest type 0, then the 1’s behave like a critical branching process. The time needed for the 1’s to die out is O(k) and the number of type-1 births before they die out is O(k2). Thus we expect the first type 2 to occur in a type 1 family that reaches size , and hence has births. The condition in (1) guarantees k1 ≪ N.
-
(A2)Since the voter model is a martingale, the probability a type-1 mutation creates a family that reaches size is . More to the point a simple computation (consider what happens at the first jump) shows that the probability a type-1 family gives rise to a type-2 before it dies out is . Since mutations to type 1 occur at times of a rate Nu1 Poisson process and with probability give rise to a type 2, it follows that if ρ2 is the birth time of the type-1 mutant that first gives rise to a type 2 then
To complete the proof we need to show that τ2 − ρ2 ≪ ρ2, and for this we need the condition N ≪ 1/u1 in (1).
-
(A3)
By the discussion of (A1), the first mutation will occur in a family that reaches a size . If , then computations with Girsanov’s formula show that (in the limit u2 → 0) the behavior of the Moran model, while it is , is indistinguishable from the case with no drift.
The assumption of a homogeneously mixing cell populations simplifies calculations considerably, but is not realistic for solid tumors. For this reason, Komarova [12] considered a spatial model, which is very similar to one introduced much earlier by Williams and Bjerknes [20]. Due to work of Bramson and Griffeath [2, 3], the second model is known to probabilists as the biased voter model.
In the usual formulation of the biased voter model, each site on the d-dimensional integer lattice
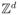 can be in state 0 or 1 indicating the presence of a cell with relative fitness 1 or λ > 1. Cells give birth at a rate equal to their fitness, and the offspring replaces a nearest neighbor chosen at random. When λ = 1 this is the voter model which was introduced independently by Clifford and Sudbury [4] and Holley and Liggett [9]. For a summary of what is known see Liggett [15].
can be in state 0 or 1 indicating the presence of a cell with relative fitness 1 or λ > 1. Cells give birth at a rate equal to their fitness, and the offspring replaces a nearest neighbor chosen at random. When λ = 1 this is the voter model which was introduced independently by Clifford and Sudbury [4] and Holley and Liggett [9]. For a summary of what is known see Liggett [15].
In the biased voter model births drive the process. In Komarova’s version cells die at rate 1 and are replaced by a copy of a nearest neighbor chosen with probability proportional to its fitness. A site with ni neighbors in state i makes
In d = 1 if the set of sites in state 1 is an interval [ℓ, r] with ℓ < r then any site that can change has n1 = n0 = 1 so Komarova’s model is a time change of the biased voter model. In d ≥ 2 this is not exactly true. However, we are interested in values of λ = 1 + s where s = 0.02 or even less, so we expect the two models to have very similar behavior. In any case, the difference between the two models is much less than their difference from reality, so we will choose to study the biased voter, whose duality with branching coalescing random walk (to be described below) gives us a powerful tool for doing computations.
Since we want a finite cell population we will restrict our process to be a subset of (−L/2, L/2]d. Komarova [12] uses “Dirichlet boundary conditions”, i.e., she assumes her space is an interval with no cells outside, but this is awkward because the set of type 1 cells may reach one end of the interval and then no further changes happen at that end. To avoid this, we will use periodic boundary conditions, i.e., we consider (
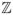 mod L)d. The resulting toroidal geometry is a little strange for studying cancer. However, using (
mod L)d has the advantage that the space looks the same seen from any point. Our results will show that for the parameter values the first type 2 will arise when the radius of the set of sites occupied by 1’s is ≪ L so the boundary conditions do not matter.
mod L)d. The resulting toroidal geometry is a little strange for studying cancer. However, using (
mod L)d has the advantage that the space looks the same seen from any point. Our results will show that for the parameter values the first type 2 will arise when the radius of the set of sites occupied by 1’s is ≪ L so the boundary conditions do not matter.
Let
be the set of cells equal to 1 in the voter model with no mutations from 0 to 1 on
starting from a single type 1 at 0. Let |
| be the number of cells in
, and let
| (2) |
This quantity, which is defined for the voter model without mutation, calculates the probability, which depends on the dimension d, that a mutation to type 1 gives rise to a type 2 before its family dies out. To see why this is true note that the integral gives the total number of man-hours in the type 1 family, and conditional on this the number of mutations that will occur is Poisson with mean .
Since mutations to type 1 in a population of N cells occur at rate Nu1 this suggests that
| (3) |
As we will explain in a moment, there is a constant γd so that νd ~ γdhd(u2) as u2 → 0 where
| (4) |
To state the result we need one more definition:
| (5) |
Theorem 2
In the neutral case of the biased voter model, λ = 1, if we assume
| (6) |
then there are constants λd given in (12) and (13) so that as u1, u2 → 0
The same conclusion holds if |λ − 1| ≪ hd(u2).
In d = 1 this result was proved by Komarova [12], see her equation (62) and assumption (60), then change notation u1 → u, u2 → u1. See also her survey paper [13]. Note that when d ≥ 3 the order of magnitude of the waiting time and the assumptions are the same as in Theorem 1. In d = 2 there are logarithmic corrections to the behavior in Theorem 1, so only in the case of d = 1 (which is relevant to cancer in the mammary ducts) does space make a substantial change in the waiting time.
The reasons for the conditions in Theorem 2 are the same as in Theorem 1.
-
(B1)
We will see that the mutation to type-2 will occur in a type-1 family that reaches size k = O(1/hd(u2)). The left hand assumption in (6) implies that k ≪ N, so the type 2 mutant arises before the 1’s reach fixation.
-
(B2)Let ρ2 be the time of the first type 1 mutation that begins the family that eventually leads to a type 2. Since mutations to type 1 occur at rate Nu1 and lead to a type 2 with probability νd, it is easy to see that
so to prove the result we need to show that with high probability τ2 − ρ2 ≪ ρ2. As the reader will see, this is guaranteed by the right-hand assumption in (6).
-
(B3)
As in the discussion of Theorem 1, once we know that the mutation to type-2 will occur in a type-1 family that reaches size k = O(1/hd(u2)), it follows that if |λ − 1| ≪ hd(u2) then (in the limit u2 → 0) the behavior of the size of the biased voter | | is, while it is O(1/hd(u2)), indistinguishable from the case with no drift.
2 The Key to the Proof
The size of the voter model, when , is a time change of symmetric simple random walk, with jumps happening at two times the size of the boundary | |, which is the number of nearest neighbor pairs with and . The one dimensional case is easy because when the boundary . The key to the study of the process in d ≥ 2 is the observation that there are constants βd so that
| (7) |
where means that when | | is large, is close to 1 with high probability.
The intuition behind this result is that the voter model is dual to a collection of coalescing random walks, so in d ≥ 3 neighbors of points in will be unoccupied with probability ≈ βd, the probability two simple random walks started at 0 and e1 = (1, 0, … 0) never hit. In dimension d = 2, the recurrence of random walks implies that when is large, most neighbors of points in will be occupied, but due to the fat tail of the recurrence time sites will be vacant with probability ~β2/log k, where β2 = π.
Before we try to explain why (7) is true, we will list an important consequence. Let Tk be the first time . Let
Lemma 1
Let be the unbiased voter model (i.e., λ = 1) starting from a single occupied site.
| (8) |
where ⇒ indicates convergence in distribution of the stochastic processes and the limit has
where Bt is a one dimensional Brownian motion. In d = 1 the process is stopped when it hits 0. In d ≥ 2, 0 is an absorbing boundary so we don’t need to stop the process.
In d = 1 this result is trivial. If one accepts (7) then (8) can be proved easily by computing infinitesimal means and variances and using standard weak convergence results. In d ≥ 2, (7) and (8) are almost consequences of work of Cox, Durrett, and Perkins [5]. They speed up time at rate an, scale space by , and assign each point occupied in the voter model mass 1/n to define a measure-valued diffusion Xn which they prove converges to super-Brownian motion. See their Theorem 1.2. (Their scaling is a little different in d = 2 but this makes no difference to the limit.)
Let be the fraction of sites adjacent to x in state 0 at time s (with the prime indicating that we multiply this by log n in d = 2, see page 196). A key step in the proof in [5] is to show, see (I1) on page 202, that for nice test functions ϕ
| (9) |
where denote the integral of the function f against the measure . The result in (9) shows that when we integrate in time and average in space (multiplying by a test function to localize the average) then (7) is true.
From the convergence of the measure valued diffusion Xn to super-Brownian motion, (8) follows by considering the total mass. Earlier we said (8) is almost a consequence of [5], since they start their process from an initial measure (i.e., O(n) initial 1’s) while consider a single occupied site and condition on reaching nε. However, this defect can be remedied by citing the work of Bramson, Cox, and LeGall [1], who have a result, Theorem 4 on page 1012 that implies (8) in d ≥ 2.
The result in (8) is enough for Section 3, but for the calculations in Section 4 we will need a version of (7). In that section we will compute under the assumption that if
| (10) |
If one wants to give a rigorous proof of the estimates there, then small values of k, can be treated with the inequalities
and one can control large values of k using (9) and estimates such as (J1) and (J2) on page 208 of [5]. We will assume (10) in order to avoid getting bogged down in technicalities.
3 Proof, part I
Let be the probability defined in (2) ignoring mutations to type 2 that occur before Tnε. The size of the voter model, | |, is a martingale, so if we let P1 to denote the law of the voter model starting from one occupied site P1(Tnε < ∞) = 1/nε. Applying (8) now,
| (11) |
where T0 = min{t : Yt = 0}, Eε is the expected value for (Yt|Y0 = ε). We have
so if we set n = 1/hd(u2) then (4) implies nanu2 → 1 and using (11) gives
Thus the type-2 mutation will occur in a family that reaches sizes O(1/hd(u2)), and we must assume 1/hd(u2) ≪ N.
If we ignore the time to reach size 1/hd(u2), the time needed to generate the type-2 mutation is, by (8), of order
where we have written a(n) for an for readability. Thus for (B2) we need a(1/hd(u2)) ≪ 1/Nu1hd(u2), which means N ≪ gd(u2)/u1.
The next order of business is to compute νd. Stochastic calculus (or calculations with infinitesimal generators) tells us that
is the unique function on [0, ∞) with values in [0, 1], v(0) = 1 and
In d = 1 all solutions have the form:
where Ai and Bi are Airy functions
Since Bi is unbounded and Ai is decreasing on [0, ∞), we take β = 0 and set α = 32/3Γ(2/3) to satisfy the boundary condition, v(0) = 1. Letting ε → 0 we conclude that
| (12) |
In d ≥ 2, , and we have
| (13) |
4 Proof, part II: Missing details for λ = 1
In the previous section we have calculated the probability that a type 1 family reaches size ε/hd(u2) and then gives rise to a type 2. To let ε → 0 and prove Theorem 2 we need to consider the possibility of a mutation to type 2 in a family that (i) never reaches size nε, or (ii) will reach nε but hasn’t yet. To have a convenient name we will call these small families. Families of the first kind arise at rate Nu1(1 − 1/nε) and families of the second kind arise at rate Nu1/nε. We will now calculate the expected rate at which type-2’s are born from these small families. In the proof of Theorem 2, we will let ε → 0 slowly as n → ∞ so we can and will assume nε → ∞.
Consider the voter model starting from a single 1 at the origin at time 0. Let Vk be the total time spent at level k, i.e., |{ }| and let Nk be the total number of returns to level k before leaving the interval (0, nε). Recalling our assumption in (10), we let q(k) the rate jumps occur at level k.
Let Sk be the embedded discrete time chain, which is a simple random walk, and let .
| (14) |
where the bar indicates conditioning on T0 < Tnε. A similar argument shows that
| (15) |
where the hat indicates conditioning on Tnε < T0.
The three conditional probabilities we need can be computed using facts about simple random walk that follow from the fact that it is a martingale.
| (16) |
For the next two we note that the first step has to be in the correct direction for these events to happen.
| (17) |
| (18) |
Thus the expected total man-hours for a family that will die out before reaching size nε is
| (19) |
and in families that have yet to reach size nε,
| (20) |
The next result shows that the contribution of small families are indeed negligible. Note that in all three cases the order of magnitude of the contributions from small families is the same as the overall rate, but contains a constant that → 0 as ε → 0.
Lemma 2
The expected total man-hours in small families is
Proof
In one dimension, q(k) = 2. The sum in (19) is dominated by
Thus, families of the first kind produce type 2’s at rate ≤ Nu1u2(nε)2/12. The expression in (20) is dominated by
Thus, families of the second kind produce type-2’s at rate ≤ Nu1u2(nε)2/6. Adding the last two conclusions gives the result for d = 1.
In d ≥ 3, (10) implies q(k) = 2dβdk, so (19) becomes
The sum is bounded above by the integral
so with our choice of , families of the first kind produce type 2’s at rate bounded above by . Setting q(k) = 2dβdk, (20) becomes
The sum is bounded above by the integral
Thus, families of the second kind produce type-2’s at . Adding the last two conclusions gives the result for d ≥ 3.
In d = 2, (10) implies q(k) = 4β2k/log k, so (19) becomes
Each term in the sum is bounded above by log(nε), so the sum is less than nε log nε. Since , families of the first kind produce type 2’s at rate bounded above by
Taking q(k) = 4β2k/log k, (20) becomes
The sum is bounded above by
Thus families of the second kind produce type 2’s at rate bounded above by
Adding the last two conclusions gives the result for d = 2 and completes the proof.
5 Proof, part III: Almost neutral mutations
In the biased voter model, whose law we denote by Pλ, jumps occur at rate 1 + λ times the size of the boundary. To compensate for this we need to run the unbiased (λ = 1) voter at rate (1 + λ)/2. If we do this, call the resulting law P̃0, and let ωT is a realization of run up to time T then the Radon-Nikodym derivative
where n+ and n− are the number of up jumps in ωt when 0 ≤ t ≤ T.
If then the difference 0 ≤ n+ − n− = O(K). Since under P̃0, | | is a time change of simple random walk, we see that the total number of jumps n+ + n− = O(K2). Taking K = 1/hd(u2) and assuming |λ − 1| ≪ hd(u2), when u2 is small the Radon Nikodym derivative is
The last result implies that (8) extends to almost neutral mutations, and the computations in Section 2 are valid. To extend the part of the proof in Section 3, we need to check that (16)–(18) are true asymptotically for almost neutral mutations. To do this we recall that if a < x < b
| (21) |
When 0 ≤ a < x ≤ b = O(1/hd(u2)) and |λ − 1| ≪ hd(u2) we have
To show that the sums come out the same we need the following uniform version which follows from (21). If |λ − 1|hd(u2) → 0 then for any C fixed
References
- 1.Bramson M, Cox T, Le Gall J. Super-Brownian limits of voter model clusters. Ann Probab. 2001;29:1001–1032. [Google Scholar]
- 2.Bramson M, Griffeath D. On the Williams-Bjerknes tumour growth model. II. Math Proc Camb Phil Soc. 1980;88:339–357. [Google Scholar]
- 3.Bramson M, Griffeath D. On the Williams-Bjerknes tumour growth model. I. Ann Probab. 1981;9:173–185. [Google Scholar]
- 4.Clifford P, Sudbury A. A model for spatial conflict. Biometrika. 1973;60:581–588. [Google Scholar]
- 5.Cox JT, Durrett R, Perkins EA. Rescaled voter models converge to super-Brownian motion. Ann Probab. 2000;28:185–234. [Google Scholar]
- 6.Durrett R, Foo J, Leder K. Spatial Moran models, II. Tumor growth and progression 2012 [Google Scholar]
- 7.Durrett R, Schmidt D. Waiting for two mutations: with applications to regulatory sequence evolution and the limits of Darwinian evolution. Genetics. 2008;180:1501–1509. doi: 10.1534/genetics.107.082610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Durrett R, Schmidt D, Schweinsberg J. A waiting time problem arising from the study of multi-stage carcinogenesis. Ann Appl Probab. 2009;19:676–718. [Google Scholar]
- 9.Holley RA, Liggett TM. Ergodic theorems for weakly interacting infinite systems and the voter model. Ann Probab. 1975;3:643–663. [Google Scholar]
- 10.Iwasa Y, Michor F, Nowak MA. Stochastic tunnels in evolutionary dynamics. Genetics. 2004;166:1571–1579. doi: 10.1534/genetics.166.3.1571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Iwasa Y, Michor F, Komarova NL, Nowak MA. Population genetics of tumor suppressor genes. J Theor Biol. 2005;233:15–23. doi: 10.1016/j.jtbi.2004.09.001. [DOI] [PubMed] [Google Scholar]
- 12.Komarova NL. Spatial stochastic models of cancer initiation and progression. Bull Math Biol. 2006;68:1573–1599. doi: 10.1007/s11538-005-9046-8. [DOI] [PubMed] [Google Scholar]
- 13.Komarova NL. Loss and gain-of-function mutations in cancer: Mass action-action, sptail and hierarchical models. J Stat Phys. 2007;128:413–448. [Google Scholar]
- 14.Komarova NL, Sengupta A, Nowak MA. Mutation-selection networks of cancer initiation: tumor suppressor genes and chromosomal instability. J Theor Biol. 2003;223:433–450. doi: 10.1016/s0022-5193(03)00120-6. [DOI] [PubMed] [Google Scholar]
- 15.Liggett TM. Stochastic interacting systems: contact, voter and exclusion processes. Springer; New York: 1999. [Google Scholar]
- 16.Nowak MA. Evolutionary Dynamics: Exploring the Equations of Life. Belknap Press; Cambridge, MA: 2006. [Google Scholar]
- 17.Nowak MA, Komarova NL, Sengupta A, Jallepalli PV, Shih IM, Vogelstein B, Lengauer C. The role of chromosomal instability in tumor initiation. Proc Natl Acad Sci. 99:16226–16231. doi: 10.1073/pnas.202617399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Nowak MA, Michor F, Komarova NL, Iwasa Y. Evolutionary ddynamics of tumor suppressor gene inactivation. Proc Natl Acad Sci. 2004;101:10635–10638. doi: 10.1073/pnas.0400747101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Schweinsberg J. The waiting time for m mutations. Electron J Probab. 2008;13:1442–1478. [Google Scholar]
- 20.Williams T, Bjerknes R. Stochastic model for abnormal clone spread through epithelial basal layer. Nature. 1972;235:19–21. doi: 10.1038/236019a0. [DOI] [PubMed] [Google Scholar]