

Abstract
Our understanding of the role of myeloid-derived suppressor cells (MDSCs) in cancer is becoming increasingly complex. In addition to their eponymous role in suppressing immune responses, they directly support tumor growth, differentiation, and metastasis in a number of ways that are only now beginning to be appreciated. It is because of this increasingly complex role that these cells may become an important factor in the treatment of human cancer. In this Review, we discuss the most pertinent and controversial issues of MDSC biology and their role in promoting cancer progression and highlight how these cells may be used in the clinic, both as prognostic factors and as therapeutic targets.
Introduction
As immunotherapy has become a viable strategy for treating different types of cancer, it has become clear that it faces serious barriers that limit its clinical efficacy. The search for the factors blunting spontaneous or therapeutic immune responses in cancer has resulted in the discovery of myeloid-derived suppressor cells (MDSCs), a group of pathologically activated immature myeloid cells with potent immunosuppressive capacity. Although the term MDSC was formally introduced in 2007 (1), cells with similar characteristics were described over 35 years ago (2). In recent years, a vast amount of information has been generated detailing the biology and clinical significance of these cells in various pathologic conditions. In this Review, we will discuss recent progress as well as the main unresolved issues associated with these cells.
Definition of MDSC
MDSCs are broadly defined as myeloid cells and are distinct from terminally differentiated mature myeloid cells (i.e., macrophages, DCs, or neutrophils). MDSCs include a small group of myeloid progenitors as well as immature mononuclear cells, which are morphologically and phenotypically similar to monocytes (M-MDSCs), and immature polymorphonuclear (PMN) cells, which are morphologically and phenotypically similar to neutrophils (PMN-MDSCs) (Figure 1). In mice, MDSCs are broadly identified as CD11b+Gr1+ cells. The two major subsets of MDSCs can be differentiated by variable expression of the Gr-1 marker (Gr-1hi cells are mostly PMN-MDSCs, and Gr-1lo cells are mostly M-MDSCs) (3). The Gr-1 marker is not a singular molecule, but instead is a combination of the Ly6C and Ly6G markers, and these subsets can be more accurately identified based on Ly6C and Ly6G markers (M-MDSC as CD11b+Ly6ChiLy6G– and PMN-MDSC as CD11b+Ly6CloLy6G+) (4, 5).
Figure 1. MDSC ontogeny.

Differentiation of neutrophils and mononuclear cells in naive mice is shown by black arrows. Hematopoietic stem cells (HSC) differentiate into common myeloid progenitors (CMP), then into granulocyte-macrophage progenitors (GMP), which give rise to mature neutrophils via sequential steps of differentiation involving myeloblasts, promyelocytes, myelocytes, metamyelocytes, and band forms. Differentiation of macrophages and DCs involves macrophage/DC progenitors (MDP), DC progenitors (CDP), and pre-cDCs as well as several types of monocytes. The most prominent are Ly6C+ inflammatory monocytes and Ly6C– patrolling monocytes. Differentiation of myeloid cells in tumor-bearing mice is shown by red arrows. Tumor-derived signals affect all steps of granulocytic and monocytic cell differentiation, causing expansion of pathologically activated PMN-MDSCs and Ly6C+ M-MDSCs. During tumorigenesis, these cells become more prevalent in bone marrow and spleen than in their nonsuppressive counterparts. M-MDSCs acquire the ability to differentiate to PMN-MDSCs and, at the tumor site, differentiate to TAMs and DCs. The dashed line represents pathways that are not yet firmly established.
In humans, MDSCs are identified in the mononuclear fraction. PMN-MDSCs are CD14–CD11b+CD33+CD15+ or CD66b+ cells and M-MDSCs are CD14+HLA-DR–/lo cells. Populations of Lin–HLA-DR–CD33+ MDSCs represent a mixed group of cells enriched for myeloid progenitors. Accurate characterization of MDSCs in cancer patients requires the analysis of all three groups of cells. Several other markers have been suggested to characterize MDSCs further; however, none has emerged as a clear MDSC-specific marker. This subject has been recently reviewed (6, 7).
A topic of some controversy is the distinction of PMN-MDSCs from neutrophils. In some reports, neutrophils with immunosuppressive and protumor functions are called N2, as opposed to antitumor N1, neutrophils (8, 9). It is rather difficult to envision that very short-lived, terminally differentiated PMN cells could be effectively polarized in tumor tissues. It is more likely that N1 cells represent activated bona fide PMN cells, whereas N2 cells are in fact PMN-MDSCs. However, this question cannot be resolved without identification of markers that allow for the delineation of PMN cells and PMN-MDSCs. In mice, several parameters that could distinguish PMN-MDSCs from PMN cells have been suggested (10); however, none is sufficient, and more effort is needed to better distinguish these cells.
In humans, separation of neutrophils from PMN-MDSCs is done via density gradient, with PMN-MDSCs staying in the PBMC fraction while neutrophils are pelleted out over Ficoll (11). In healthy individuals, PMN-MDSCs are practically undetectable, and hence, few to no cells with this phenotype remain in the PBMC fraction. Although very useful in providing scientific information, the use of cell density as a clinical biomarker is limited by the fact that the density of PMN cells depends on the conditions of blood collection and storage and is prone to fluctuations that may affect the interpretation of the results (12). Development of definitive markers that will allow for one-step identification of MDSCs is necessary. A similar challenge exists for M-MDSCs; however, it is less critical in humans due to the fact that monocytes can be distinguished from M-MDSCs by their phenotype (CD14+HLA-DRhi vs. CD14+HLA-DR–/lo, respectively).
Immunosuppressive functions of MDSCs
As evident from their name, the initial defining feature of MDSCs is their ability to suppress immune cell function. Main factors implicated in MDSC-mediated immune suppression include expression of arginase (ARG1), inducible NOS (iNOS) (3–5), TGF-β (13–15), IL-10 (16), and COX2 (17–19), sequestration of cysteine (20), decreased expression of l-selectin by T cells (21), and induction of Tregs (22), among others. In recent years, it has become clear that M-MDSCs and PMN-MDSCs utilize different mechanisms of immune suppression. M-MDSCs suppress both antigen-specific and nonspecific T cell responses and, on a per cell basis, are consistently shown to have stronger suppressive activity than PMN-MDSCs. M-MDSCs primarily utilize mechanisms associated with production of NO and cytokines (reviewed in ref. 23). In contrast, PMN-MDSCs are capable of suppressing immune responses in an antigen-specific manner only. ROS production is essential for this function. Reaction of NO with superoxide generates peroxynitrite (PNT), which directly inhibits T cells by nitrating T cell receptors (TCRs) and reducing their responsiveness to cognate antigen-MHC complexes (24). Additionally, nitration reduces binding of antigenic peptides to MHC molecules on tumor cells (25) and blocks T cell migration by nitrating T cell–specific chemokines (26).
The relatively weak suppressive activity of PMN-MDSCs has raised the question of whether these cells play a role in immune suppression. A recent study made the claim that PMN-MDSCs were not involved (27), based on the fact that myeloid leukemia cell-1 (MCL-1) deletion, which results in deletion of granulocytes, including PMN-MDSCs, did not affect tumor incidence in mice. However, deletion of PMN-MDSCs in these mice was associated with a substantial increase in M-MDSCs, which could effectively compensate for any loss of PMN-MDSCs. Additionally, the impact of M-MDSC deletion on tumor incidence was not explored, nor were immune correlates investigated. In contrast to this report, a number of studies have demonstrated improvement of immune responses and antitumor immunity following depletion of PMN-MDSCs (but not M-MDSCs) with a Ly6G-specific antibody (28–30). It is also important to note that, although M-MDSCs are more suppressive on a per cell basis, PMN-MDSCs are generally much more prevalent in cancer (4). Thus, the role of PMN-MDSCs in tumor-associated immune suppression requires further elucidation.
The large number of different immunosuppressive mechanisms described for MDSCs does not mean that these mechanisms are simultaneously operational. The prevalence of a particular immunosuppressive mechanism depends on the type of MDSCs that expanded in response to disease as well as on the stage of the disease and the site where the suppression is occurring. It is likely that at any given time there is a dominant suppressive mechanism used by MDSCs and that this mechanism could change throughout the progression of the disease. This suggests that evaluation of the role of MDSCs in cancer probably cannot be based only on one or two factors produced by these cells, but should include analysis of their functional activity. It also suggests that, for therapeutic purposes, targeting of one mechanism may not be effective unless this mechanism has been determined to be dominant in the type of cancer being treated.
MDSCs in the regulation of tumor development and progression
The ability of MDSCs to support tumor growth and metastases can be broadly divided into four functions: (a) protection of tumor cells from immune-mediated killing, (b) remodeling of the tumor microenvironment, (c) establishment of a premetastatic niche, and (d) interaction with tumor cells to induce “stemness” and facilitate the epithelial-to-mesenchymal transition (EMT) (Figure 2).
Figure 2. Potential role of immature myeloid cells and MDSCs in the regulation of tumor development and progression.

(A) Immature myeloid cells (IMCs) with the typical phenotype of MDSCs are produced in response to inflammatory stimuli. However, these cells often lack immunosuppressive activity. They contribute to tumorigenesis by recruiting proinflammatory CD4+ T cells that promote epithelial cell proliferation. (B) Tumor development is associated with the expansion of cells with acquired immunosuppressive activity (MDSCs). These cells also promote tumor cell invasion and angiogenesis and neutralize tumor cell senescence. (C) In metastatic tumors, MDSCs, in addition to promoting tumor cell invasion and angiogenesis, can support EMT and differentiation of osteoclasts supporting bone resorption.
The mechanism of immune privilege recently was shown to be important in the increased permissiveness for metastasis seen in pregnancy. Specifically, decreased NK cell functionality in pregnant mice was dependent on MDSC accumulation. This accumulation was shown to be an essential part of pregnancy-associated metastasis, as no difference in the metastatic potential of NK cell–resistant tumor cell lines was observed during pregnancy (31). Additionally, an increase in the immunosuppressive potential of MDSCs was also shown to correlate with lymph node metastasis in breast cancer patients (32).
MDSCs are capable of supporting tumor growth through remodeling of the tumor microenvironment (33, 34). They have been shown to produce VEGF, bFGF, VEGF analogue Bv8, and MMP9, all essential mediators of neoangiogenesis and tissue invasion at the tumor site (35–37). Expression of these mediators has been linked to MDSC-mediated metastasis and is independent of their immunosuppressive capacity (38).
The concept of the premetastatic niche refers to the situation wherein bone marrow–derived cells prepare distant organs for the arrival of tumor cells (39, 40). Mouse models have shown that MDSCs appear in the lungs as early as two weeks prior to the appearance of metastases. The presence of these cells correlated with decreased immune function in the lungs. Importantly, myeloid-specific deletion of Mmp9 essentially eliminated metastasis, suggesting that the role of MDSCs in tissue remodeling, and not just their ability to suppress immune responses, was indispensable for their metastasis-promoting function (41). Tumor-derived hypoxia-induced lysyl oxidase, VEGF, S100A8/A9, IL-6, and IL-10 (via activation of the S1PR1/STAT3 axis in the lungs) have all been implicated in the recruitment and activation of MDSCs in the lungs and the establishment of the premetastatic niche (42–44).
The contribution of myeloid cells to EMT has only recently begun to emerge. Coculture of MDSCs with tumor cells induced a stem-like phenotype in tumor cells and increased their ability to metastasize in vivo (45). In the ret model of spontaneous mouse melanoma, CD11b+Gr1+ cells were shown to infiltrate primary tumors in a mechanism that was dependent on CXCL5. Depletion of these cells resulted in smaller, less nodal tumors and a drastic reduction in metastasis. In vitro coculture experiments showed that these cells induced a stem-like phenotype in tumor cells and that inhibition of the TGF-β, EGF, and/or HGF pathways could reverse this effect (46). Studies using the 4T1 model of transplantable breast cancer implicated MDSC-derived IL-6 in increased tumor cell stemness (47). Because IL-6 and HGF have been shown to upregulate each other’s receptors (48) and IL-6R/EGFR crosstalk has previously been implicated in EMT (49), it is possible that MDSC induction of EMT is mediated primarily though IL-6; however, more studies are needed to confirm this assumption.
Bona fide MDSCs are not produced in healthy individuals. Even in the early stages of cancer, cells with an MDSC-like phenotype may not have immunosuppressive activity (50). Recent evidence has linked accumulation of immature myeloid cells with an MDSC-like phenotype during chronic inflammation with the early stages of tumor development. Exposure of mice to cigarette smoke caused accumulation of these cells in lung and spleen (34); however, these cells became immunosuppressive MDSCs only after the development of lung cancer. Nevertheless, their depletion increased survival (34). In a model of skin carcinogenesis, accumulation of immature myeloid cells without suppressive function in the skin of mice strongly promoted tumor formation. That effect was caused by CCL4-mediated recruitment of IL-17–producing CD4+ T cells (51). CD11b+Gr1+ cells were also recently shown to oppose cellular senescence and promote a more aggressive disease in a mouse model of spontaneous prostate cancer by antagonizing IL-1α–mediated senescence (52). Thus, cells with an MDSC-like phenotype play a significant role in tumor development and progression via mechanisms that are not necessarily related to their ability to suppress tumor-specific immune responses (Figure 2).
Mechanisms of accumulation and activation of MDSCs in cancer
It is likely that, in addition to the common mechanisms regulating both expansion of immature myeloid cells and acquisition of suppressive function, there are distinct mechanisms that mediate these two effects. We previously suggested that accumulation of bona fide MDSCs requires two sets of signals (53). The first of these signals supports the expansion of immature myeloid cells and is mediated largely by granulocyte-macrophage CSF (GM-CSF), macrophage CSF (M-CSF), granulocyte CSF (G-CSF), and other growth factors produced by tumor cells and tumor stroma (54). Although expression of these factors is sufficient to expand cells with an MDSC-like phenotype, they do not imbue them with the suppressive phenotype that defines this cell type (55), which requires a second signal. Many different proinflammatory cytokines (IL-6, IL-1β, TNF-α, IL-13/4, IFN-γ, etc.) have been suggested as candidates; however, the effect of these cytokines has been observed mostly during extended culture of progenitor cells in vitro. Elimination through genetic knockout or in vivo blockade of one or several of these mediators has shown limited effect on the ability of MDSCs to suppress T cell responses (23). Additionally, models of chronic inflammation caused by smoking or skin irritation showed that these conditions were not sufficient to generate bona fide MDSCs (34, 51). Thus, the mechanism controlling the conversion of expanded immature myeloid cells to MDSCs is complex and requires further investigation.
Members of the STAT transcription factor family (STAT3, STAT5, and STAT6) have long been considered to be critical factors in the regulation of MDSC expansion and activity (56–59). In recent years, more information has emerged regarding the mechanism of MDSC regulation by STAT3. STAT3 can regulate arginase production by binding directly to the ARG1 promoter (60). The effect of STAT3 on MDSC accumulation and suppressive function was found to be mediated by C/EBPβ (55) and IFN regulatory factor-8 (IRF8) (61, 62). Pharmacological inhibition of STAT3 in MDSCs favored their conversion toward mature DCs, in part through inhibition of PKCβII (63).
HIF-1α has emerged as another important regulator of MDSC function and differentiation. At the tumor site, HIF-1α facilitates the differentiation of MDSCs to tumor-associated macrophages (TAMs) and promotes the suppressive activity of MDSCs via upregulation of iNOS and arginase and downregulation of components of the NADPH oxidase complex (64). A recent report demonstrated that HIF-1α bound to the promoter of programmed cell death ligand-1 (PD-L1) and upregulated its expression on myeloid cells (65). Further studies will be necessary to determine whether this mechanism represents a function of MDSCs or is indicative of their switch to TAMs. The canonical Wnt pathway has been shown to antagonize MDSC differentiation and support the differentiation of mature DCs (66, 67). A recent pathway analysis comparing MDSCs, both in the periphery and at the tumor site, with normal immature myeloid cells has provided a comprehensive look at the number of factors enriched in MDSCs in different physiological settings (68). It represents the first step toward a comprehensive understanding of how MDSC function is controlled.
Recently, ER stress was implicated in MDSC-mediated suppression. The ER stress response is an evolutionarily conserved process designed to protect cells from stress signals, including starvation, viral infections, changes in oxygen tension, etc. Transmission of ER stress from the tumor cells to macrophages and DCs has been shown to skew them toward promotion rather than suppression of tumor growth (69, 70). A recent study demonstrated that, compared with their counterparts from healthy animals, MDSCs from tumor-bearing mice and cancer patients showed increases in DNA damage-inducible transcript 3 (CHOP), spliced X-box binding protein 1 (XBP1), and Bip (71). These molecules are downstream effectors of the major pathways of the ER stress response (72, 73). Chop deletion in mice resulted in a decrease in tumor growth. Tumor MDSCs from Chop KO mice exhibited greater survival than wild-type MDSCs and were shifted toward stimulation, rather than suppression, of T cell responses. Decreased IL-6 production by MDSCs was observed and exogenous IL-6 could “rescue” the function of Chop KO MDSCs (74). However, the importance of CHOP in the immunosuppressive phenotype of MDSCs requires further characterization, since there is a lack of consensus among different groups on its role in tumor progression (75). Of particular interest is the differentiation of the role of CHOP in immunosuppression at the tumor site versus in the periphery. A recent report showed that ER stress inducer thapsigargin increased the suppressive capacity of MDSCs at the tumor site, but not in the periphery (76); however, this study evaluated the total population of CD11b+ cells, allowing for the interpretation that the effect of ER stress may not be MDSC specific.
Recent additions to the list of inflammatory mediators controlling MDSC function include high mobility group box 1 (HMGB1) and PPARγ. HMGB1 promotes the development of MDSCs and contributes to their ability to suppress antigen-driven activation of T cells (77). Overexpression of PPARγ resulted in expansion of PMN-MDSCs with immunosuppressive activity. This was associated with activation of STAT3, NF-κB, Erk1/2, and p38 (78).
The fate of MDSCs in the tumor-bearing host
It is known that cells with an M-MDSC phenotype can differentiate into macrophages and DCs (79–81). However, the role of this differentiation in tumor progression has only recently been expounded upon. At the tumor site, adoptively transferred MDSCs primarily differentiated into immunosuppressive TAMs through a mechanism that was shown to be dependent on HIF-1α. In the periphery, MDSCs transitioned equally to macrophages and DCs (64). However, a recent study suggested that MDSCs are capable of differentiating into CD11cloCD11bhi regulatory DCs both at the tumor site and in the periphery (82). Discrepancies between this and the previous study may be due to the use of an experimental lung metastatic model in the latter study, which reduced the level of hypoxia at the tumor site and likely reduced HIF-1α expression. Another study showed that MDSCs from mice bearing bone metastases were capable of differentiating into osteoclasts in an NO-dependent manner (83). These osteoclasts are capable of bone resorption and are involved in the bone remodeling necessary for the colonization of bone metastases. In the lung, MDSCs have been shown to differentiate into fibrocytes and myofibroblasts, which support the establishment of lung metastases. This transition was dependent on binding of the transcription factor Kruppel-like factor 4 (KLF4) to the fibroblast-specific protein 1 (Fsp1) promoter (84, 85). In fact, some studies have shown that fibrocytes represent a unique subset of MDSCs that differentiate from M-MDSCs in the presence of IL-4. These cells can suppress immune responses through expression of indoleamine oxidase (IDO) and expansion of Tregs (86, 87).
Accumulation of MDSCs in tumor-bearing hosts could indicate that these cells are protected from apoptosis. Several protective mechanisms have been suggested. TNF signaling in MDSCs has been shown to be important in maintaining their survival. Upregulation of the caspase-8 inhibitor c-FLIP was a prominent feature of TNF treatment in MDSCs, and inhibition of caspase-8 rescued decreased accumulation of MDSCs in Tnf KO mice (38). However, a recent study suggested that c-FLIP might not necessarily affect survival of MDSCs in tumor-bearing mice, but instead could affect MDSC development (27). No effect on the development of PMN-MDSCs in the absence of c-FLIP was shown; however, one could envision a scenario in which c-FLIP–deficient PMN-MDSCs may be phenotypically quite different from their wild-type counterparts, especially given the ability of M-MDSCs to serve as a source of PMN-MDSCs in tumor-bearing mice (88). Decreases in FasL-mediated killing have been related to increases in Bcl-xL in MDSCs relative to immature myeloid cells from tumor-free animals (89, 90). However, a recent study demonstrated that, in the periphery, MDSCs actually had a much shorter life span than their control counterparts, PMN cells and monocytes. This effect was mediated by changes in the expression of TRAIL receptors. Interestingly, MDSC cell death in the periphery directly contributed to enhanced expansion of MDSCs in bone marrow (71). These results may have substantial implications for the therapeutic targeting of MDSCs. An important distinction between this study and previous work was that it examined the survival of MDSCs directly in vivo. In vitro assays of MDSCs after isolation from tissues have inherent difficulties relating to the sensitivity of these cells to mechanical isolation, which may select for more apoptosis-resistant cells. More detailed studies are needed to establish the life span of MDSCs in the tumor-bearing host.
MDSCs as biomarker of tumor progression and responsiveness to therapy
Ample evidence supports a close association between MDSC accumulation and clinical outcome in cancer patients (91). Most solid tumors have expansion of PMN-MDSCs, with some exceptions, including melanoma, multiple myeloma, and prostate cancer. In these cancers, the M-MDSC population is more prevalent. Such preferential expansion is likely caused by variability in the factors that predominate among the different cancer types. This issue and the role MDSCs play in cancer prognosis have recently been reviewed for a number of tumor types (92). Although identification of MDSCs in the peripheral blood of cancer patients is now well established, the situation with the detection of these cells in tissues using immunohistochemistry is different. This is due to the fact that multicolor analysis of paraffin-embedded tissues, suitable for biomarker research, is very difficult. Recently, we found that double-staining of tissues with CD33 and S100A9 antibodies detected cells that phenotypically reflect the population of MDSCs and accumulate in melanoma and colon cancer patients as well as in the patients with some inflammatory conditions (51). More studies are needed to validate the use of this approach for detection of MDSCs.
In recent years, MDSCs have been implicated in resistance to anticancer therapies. Resistance to the tyrosine kinase inhibitor sunitinib was associated with increased GM-CSF levels and persistent high levels of MDSCs in the peripheral blood of patients with renal cell carcinoma. This effect correlated with elevated MMP9, MMP8, and IL-8 levels (93). Clinical response to a number of chemotherapeutics is negatively correlated with MDSC levels. In non–small cell lung cancer patients, circulating CD14+S100A9+ cells predicted a poorer response to cisplatin and other chemotherapeutics (94, 95). An earlier study showed similar findings with a population of PMN-MDSCs (CD11b+CD14–CD15+CD33+) (96). The cells in these studies were capable of suppressing CD8+ T cells. MDSCs have also been associated with an inhibitory effect of chemotherapy on the immune system. Cyclophosphamide has been shown to increase MDSC numbers in both preclinical and clinical settings (97), and this was accompanied by reduced CD8+ T cell and NK cell activity (98) and reduced antibody responses (99).
Recent studies suggest that MDSCs are an important factor regulating the response to immune therapies. The efficacy of ipilimumab, a monoclonal antibody against CTLA-4, was inversely correlated with the presence of M-MDSCs in melanoma patients prior to treatment (100, 101). Among patients receiving adoptive transfer of cytokine-induced killer cells, those with low MDSC numbers were significantly more likely to survive in the long term (102). Similar results were found in patients treated with high-dose IL-2 (103). In a cancer-preventative setting, the presence of MDSCs in patients with advanced colonic adenomas was inversely correlated with responsiveness to a vaccination against the tumor antigen mucin 1 (MUC1) (104). These studies highlight MDSCs as a valuable biomarker and suggest that targeting MDSCs may be an attractive strategy for improving the efficacy of concurrent therapies.
Strategies to therapeutically target MDSCs
The fact that MDSCs play an important role in the regulation of tumor growth has stimulated the search for a way to therapeutically target these cells. Several strategies are currently being tested in clinic (Figure 3). These strategies can broadly be placed into three categories: (a) elimination of MDSCs, (b) deactivation of MDSCs, and (c) skewing of myelopoeisis away from the accumulation of MDSCs. An extensive review of MDSC targeting has recently been published (105). Here, we will give a brief overview of this exciting field and highlight some new studies.
Figure 3. Therapeutic strategies targeting MDSCs.
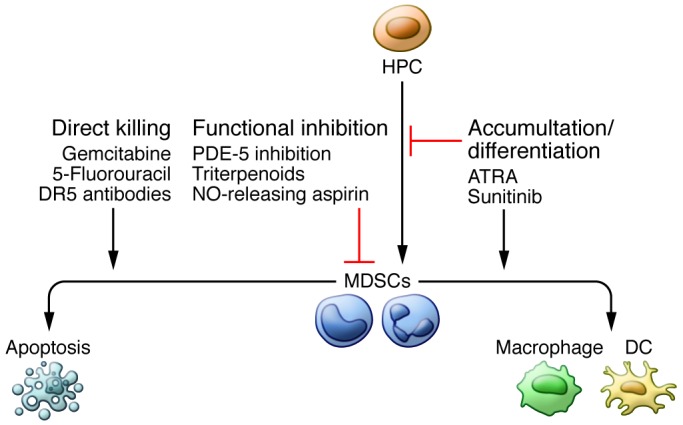
MDSCs can be targeted in one of three broadly defined ways. First, they can be directly killed. Low-dose gemcitabine, 5-fluorouracil, and TRAIL receptor ligation have shown efficacy in doing so, both clinically and preclinically. The second category of therapeutic is functional inhibition of MDSC-suppressive machinery. PDE-5 inhibitors, such as taldalafil, as well as NO-releasing aspirin and synthetic triterpenoids act effectively in this manner, in part by reducing the expression of ROS, reactive nitrogen species, and arginase, all central to the ability of MDSCs to inhibit immune responses. Finally, myelopoeisis can be skewed such that MDSC accumulation is inhibited and/or MDSCs are forcefully differentiated into more terminally differentiated, immunostimulatory myeloid cells such as DCs and macrophages. Sunitinib and ATRA have shown efficacy in these two functions, respectively.
MDSCs can be eliminated with relatively low doses of chemotherapy. In murine models, gemcitabine has been shown to deplete MDSCs without deleterious effects on T cells, resulting in decreased tumor growth and prolonged survival (106, 107) as well as enhanced responsiveness to immune therapy (108). Cisplatin and 5-fluorouracil have been shown to preferentially deplete MDSCs compared with other immune cells (109, 110), which resulted in increased CD8+ T cell responses (111). Several trials that examine the ability of low-dose chemotherapy to augment vaccine-induced immune responses are now underway. Recently, it was shown that selective upregulation of TRAIL receptor DR5 on mouse MDSCs and downregulation of decoy receptors DCR1 and DCR2 in human MDSCs make TRAIL receptor targeting a viable and specific method of MDSC depletion. This has been shown to inhibit tumor growth in mice in a CD8+ T cell–dependent manner (71). Newly engineered peptibodies consisting of S100A9-derived peptides conjugated to antibody Fc fragments have shown promise in eliminating MDSCs in mouse models (112); however, the specificity of this targeting and its applicability to human cancer needs to be established.
MDSCs can be functionally inactivated by targeting their suppressive machinery. ROS and NO are essential components of MDSC-suppressive machinery. Nf-E2–related factor 2 (NRF2) is a transcription factor that plays an important role in protecting cells against free radical damage. NRF2 modulates the expression of several antioxidant enzymes that scavenge ROS and NO, and upregulation of NRF2 by a synthetic triterpenoid has been shown to reduce the production of ROS by MDSCs and attenuate their suppressive activity ex vivo (113). Both arginase and iNOS expression have been shown to be downregulated in response to phosphodiesterase-5 (PDE-5) inhibition (114). A recent clinical report indicated that head and neck cancer patients treated with the PDE-5 inhibitor tadalafil had fewer circulating MDSCs, lower iNOS and arginase expression in these cells, and a greater number of spontaneously generated tumor-specific T cells (115). This study was published alongside another showing that tadalfil treatment decreased Tregs and increased tumor antigen–specific CD8+ T cells in addition to reducing MDSC numbers (116). Finally, inhibition of COX-2 decreased the production of immunosuppressive prostaglandin E2, and nitroaspirin has been shown to downregulate NO production (117, 118).
In addition to targeting suppressive functions, myelopoeisis can be diverted away from generating MDSCs for therapeutic benefit. All-trans-retinoic acid (ATRA) has been shown to differentiate MDSCs into DCs and improve their immunostimulatory capacity (119, 120). Treatment of renal cell carcinoma patients with ATRA substantially decreased the presence of MDSCs in peripheral blood (121). A recent study demonstrated that in lung cancer patients, p53 vaccine–generated immune responses were improved if patients received a short course of ATRA (122). STAT3 targeting can affect MDSC expansion and differentiation and lead to differentiation of MDSCs into immunogenic DCs (81, 123). Additionally, sunitinib, which inhibits STAT3 as well as VEGF, c-kit, and M-CSF signaling, has been shown to reduce MDSC levels in renal cell carcinoma patients and may provide an avenue for improving antitumor immunity in these patients (124).
Summary
The field of MDSC research is experiencing a renaissance. As more information regarding the clinical significance and the intriguing biology of these cells has become available, the critical role they play in the regulation of immune responses and tumor progression has become more apparent. These cells represent a unique model for studying pathological activation of myeloid cells and may serve as both a powerful biomarker for improved patient selection and as a therapeutic target that could potentially enhance not only anticancer immune therapeutics, but also most anticancer therapies.
Acknowledgments
Footnotes
Conflict of interest: Dmitry I. Gabrilovich receives some research support from Jannsen Pharmaceutical Companies, Daiichi-Sankyo, Peregrine Pharmaceuticals, and Reata Pharmaceuticals.
Reference information:J Clin Invest. 2015;125(9):3356–3364. doi:10.1172/JCI80005.
Address for correspondence: Dmitry I. Gabrilovich, The Wistar Institute, Room 118, 3601 Spruce St., Philadelphia, Pennsylvania 19014, USA. Phone: 215.495.6955; E-mail: dgabrilovich@wistar.org.
This work was supported by NIH grants CA 100062 and CA 141438.
References
- 1.Gabrilovich DI, et al. The terminology issue for myeloid-derived suppressor cells. Cancer Res. 2007;67(1):425–426. doi: 10.1158/0008-5472.CAN-06-3037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bennett JA, Rao VS, Mitchell MS. Systemic bacillus Calmette-Guerin (BCG) activates natural suppressor cells. Proc Natl Acad Sci U S A. 1978;75(10):5142–5144. doi: 10.1073/pnas.75.10.5142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Dolcetti L, et al. Hierarchy of immunosuppressive strength among myeloid-derived suppressor cell subsets is determined by GM-CSF. Eur J Immunol. 2010;40(1):22–35. doi: 10.1002/eji.200939903. [DOI] [PubMed] [Google Scholar]
- 4.Youn JI, Nagaraj S, Collazo M, Gabrilovich DI. Subsets of myeloid-derived suppressor cells in tumor-bearing mice. J Immunol. 2008;181(8):5791–5802. doi: 10.4049/jimmunol.181.8.5791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Movahedi K, et al. Identification of discrete tumor-induced myeloid-derived suppressor cell subpopulations with distinct T cell-suppressive activity. Blood. 2008;111(8):4233–4244. doi: 10.1182/blood-2007-07-099226. [DOI] [PubMed] [Google Scholar]
- 6.Talmadge JE, Gabrilovich DI. History of myeloid-derived suppressor cells. Nat Rev Cancer. 2013;13(10):739–752. doi: 10.1038/nrc3581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Damuzzo V, et al. Complexity and challenges in defining myeloid-derived suppressor cells. Cytometry B Clin Cytom. 2015;88(2):77–91. doi: 10.1002/cyto.b.21206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Fridlender ZG, et al. Polarization of tumor-associated neutrophil phenotype by TGF-β: “N1” versus “N2” TAN. Cancer Cell. 2009;16(3):183–194. doi: 10.1016/j.ccr.2009.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mishalian I, Bayuh R, Levy L, Zolotarov L, Michaeli J, Fridlender ZG. Tumor-associated neutrophils (TAN) develop pro-tumorigenic properties during tumor progression. Cancer Immunol Immunother. 2013;62(11):1745–1756. doi: 10.1007/s00262-013-1476-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Youn JI, Collazo M, Shalova IN, Biswas SK, Gabrilovich DI. Characterization of the nature of granulocytic myeloid-derived suppressor cells in tumor-bearing mice. J Leukoc Biol. 2012;91(1):167–181. doi: 10.1189/jlb.0311177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Schmielau J, Finn OJ. Activated granulocytes and granulocyte-derived hydrogen peroxide are the underlying mechanism of suppression of T-cell function in advanced cancer patients. Cancer Res. 2001;61(12):4756–4760. [PubMed] [Google Scholar]
- 12.Trellakis S, et al. Granulocytic myeloid-derived suppressor cells are cryosensitive and their frequency does not correlate with serum concentrations of colony-stimulating factors in head and neck cancer. Innate Immun. 2013;19(3):328–336. doi: 10.1177/1753425912463618. [DOI] [PubMed] [Google Scholar]
- 13.Huang B, et al. Gr-1+CD115+ immature myeloid suppressor cells mediate the development of tumor-induced T regulatory cells and T-cell anergy in tumor-bearing host. Cancer Res. 2006;66(2):1123–1131. doi: 10.1158/0008-5472.CAN-05-1299. [DOI] [PubMed] [Google Scholar]
- 14.Li H, Han Y, Guo Q, Zhang M, Cao X. Cancer-expanded myeloid-derived suppressor cells induce anergy of NK cells through membrane-bound TGF-β1. J Immunol. 2009;182(1):240–249. doi: 10.4049/jimmunol.182.1.240. [DOI] [PubMed] [Google Scholar]
- 15.Elkabets M, et al. IL-1β regulates a novel myeloid-derived suppressor cell subset that impairs NK cell development and function. Eur J Immunol. 2010;40(12):3347–3357. doi: 10.1002/eji.201041037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hu CE, Gan J, Zhang RD, Cheng YR, Huang GJ. Up-regulated myeloid-derived suppressor cell contributes to hepatocellular carcinoma development by impairing dendritic cell function. Scand J Gastroenterol. 2011;46(2):156–164. doi: 10.3109/00365521.2010.516450. [DOI] [PubMed] [Google Scholar]
- 17.Mao Y, et al. Melanoma-educated CD14+ cells acquire a myeloid-derived suppressor cell phenotype through COX-2-dependent mechanisms. Cancer Res. 2013;73(13):3877–3887. doi: 10.1158/0008-5472.CAN-12-4115. [DOI] [PubMed] [Google Scholar]
- 18.Nagaraj S, Nelson A, Youn JI, Cheng P, Quiceno D, Gabrilovich DI. Antigen-specific CD4(+) T cells regulate function of myeloid-derived suppressor cells in cancer via retrograde MHC class II signaling. Cancer Res. 2012;72(4):928–938. doi: 10.1158/0008-5472.CAN-11-2863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Fujita M, et al. COX-2 blockade suppresses gliomagenesis by inhibiting myeloid-derived suppressor cells. Cancer Res. 2011;71(7):2664–2674. doi: 10.1158/0008-5472.CAN-10-3055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Srivastava MK, Sinha P, Clements VK, Rodriguez P, Ostrand-Rosenberg S. Myeloid-derived suppressor cells inhibit T-cell activation by depleting cystine and cysteine. Cancer Res. 2010;70(1):68–77. doi: 10.1158/0008-5472.CAN-09-2587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hanson EM, Clements VK, Sinha P, Ilkovitch D, Ostrand-Rosenberg S. Myeloid-derived suppressor cells down-regulate L-selectin expression on CD4+ and CD8+ T cells. J Immunol. 2009;183(2):937–944. doi: 10.4049/jimmunol.0804253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lindau D, Gielen P, Kroesen M, Wesseling P, Adema GJ. The immunosuppressive tumour network: myeloid-derived suppressor cells, regulatory T cells and natural killer T cells. Immunology. 2013;138(2):105–115. doi: 10.1111/imm.12036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gabrilovich DI, Ostrand-Rosenberg S, Bronte V. Coordinated regulation of myeloid cells by tumours. Nat Rev Immunol. 2012;12(4):253–268. doi: 10.1038/nri3175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Nagaraj S, et al. Altered recognition of antigen is a mechanism of CD8+ T cell tolerance in cancer. Nat Med. 2007;13(7):828–835. doi: 10.1038/nm1609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lu T, et al. Tumor-infiltrating myeloid cells induce tumor cell resistance to cytotoxic T cells in mice. J Clin Invest. 2011;121(10):4015–4029. doi: 10.1172/JCI45862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Molon B, et al. Chemokine nitration prevents intratumoral infiltration of antigen-specific T cells. J Exp Med. 2011;208(10):1949–1962. doi: 10.1084/jem.20101956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Haverkamp JM, et al. Myeloid-derived suppressor activity is mediated by monocytic lineages maintained by continuous inhibition of extrinsic and intrinsic death pathways. Immunity. 2014;41(6):947–959. doi: 10.1016/j.immuni.2014.10.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Srivastava MK, et al. Myeloid suppressor cell depletion augments antitumor activity in lung cancer. PLoS One. 2012;7(7):e40677. doi: 10.1371/journal.pone.0040677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Tsukamoto H, Nishikata R, Senju S, Nishimura Y. Myeloid-derived suppressor cells attenuate TH1 development through IL-6 production to promote tumor progression. Cancer Immunol Res. 2013;1(1):64–76. doi: 10.1158/2326-6066.CIR-13-0030. [DOI] [PubMed] [Google Scholar]
- 30.Fischer MA, et al. CD11b+, Ly6G+ cells produce type I interferon and exhibit tissue protective properties following peripheral virus infection. PLoS Pathog. 2011;7(11):e1002374. doi: 10.1371/journal.ppat.1002374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mauti LA, et al. Myeloid-derived suppressor cells are implicated in regulating permissiveness for tumor metastasis during mouse gestation. J Clin Invest. 2011;121(7):2794–2807. doi: 10.1172/JCI41936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yu J, et al. Myeloid-derived suppressor cells suppress antitumor immune responses through IDO expression and correlate with lymph node metastasis in patients with breast cancer. J Immunol. 2013;190(7):3783–3797. doi: 10.4049/jimmunol.1201449. [DOI] [PubMed] [Google Scholar]
- 33.Sevko A, Umansky V. Myeloid-derived suppressor cells interact with tumors in terms of myelopoiesis, tumorigenesis and immunosuppression: thick as thieves. J Cancer. 2013;4(1):3–11. doi: 10.7150/jca.5047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ortiz M, Lu L, Ramachandran I, Gabrilovich D. Myeloid-derived suppressor cells in the development of lung cancer. Cancer Immunol Res. 2014;2(1):50–58. doi: 10.1158/2326-6066.CIR-13-0129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Tartour E, et al. Angiogenesis and immunity: a bidirectional link potentially relevant for the monitoring of antiangiogenic therapy and the development of novel therapeutic combination with immunotherapy. Cancer Metastasis Rev. 2011;30(1):83–95. doi: 10.1007/s10555-011-9281-4. [DOI] [PubMed] [Google Scholar]
- 36.Casella I, et al. Autocrine-paracrine VEGF loops potentiate the maturation of megakaryocytic precursors through Flt1 receptor. Blood. 2003;101(4):1316–1323. doi: 10.1182/blood-2002-07-2184. [DOI] [PubMed] [Google Scholar]
- 37.Shojaei F, et al. G-CSF-initiated myeloid cell mobilization and angiogenesis mediate tumor refractoriness to anti-VEGF therapy in mouse models. Proc Natl Acad Sci U S A. 2009;106(16):6742–6747. doi: 10.1073/pnas.0902280106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zhao X, et al. TNF signaling drives myeloid-derived suppressor cell accumulation. J Clin Invest. 2012;122(11):4094–4104. doi: 10.1172/JCI64115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kaplan RN, Psaila B, Lyden D. Bone marrow cells in the ‘pre-metastatic niche’: within bone and beyond. Cancer Metastasis Rev. 2006;25(4):521–529. doi: 10.1007/s10555-006-9036-9. [DOI] [PubMed] [Google Scholar]
- 40.Smith HA, Kang Y. The metastasis-promoting roles of tumor-associated immune cells. J Mol Med. 2013;91(4):411–429. doi: 10.1007/s00109-013-1021-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Yan HH, et al. Gr-1+CD11b+ myeloid cells tip the balance of immune protection to tumor promotion in the premetastatic lung. Cancer Res. 2010;70(15):6139–6149. doi: 10.1158/0008-5472.CAN-10-0706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hiratsuka S, Watanabe A, Aburatani H, Maru Y. Tumour-mediated upregulation of chemoattractants and recruitment of myeloid cells predetermines lung metastasis. Nat Cell Biol. 2006;8(12):1369–1375. doi: 10.1038/ncb1507. [DOI] [PubMed] [Google Scholar]
- 43.Erler JT, et al. Hypoxia-induced lysyl oxidase is a critical mediator of bone marrow cell recruitment to form the premetastatic niche. Cancer Cell. 2009;15(1):35–44. doi: 10.1016/j.ccr.2008.11.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Deng J, et al. S1PR1-STAT3 signaling is crucial for myeloid cell colonization at future metastatic sites. Cancer Cell. 2012;21(5):642–654. doi: 10.1016/j.ccr.2012.03.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Cui TX, et al. Myeloid-derived suppressor cells enhance stemness of cancer cells by inducing microRNA101 and suppressing the corepressor CtBP2. Immunity. 2013;39(3):611–621. doi: 10.1016/j.immuni.2013.08.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Toh B, et al. Mesenchymal transition and dissemination of cancer cells is driven by myeloid-derived suppressor cells infiltrating the primary tumor. PLoS Biol. 2011;9(9):e1001162. doi: 10.1371/journal.pbio.1001162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Oh K, et al. A mutual activation loop between breast cancer cells and myeloid-derived suppressor cells facilitates spontaneous metastasis through IL-6 trans-signaling in a murine model. Breast Cancer Res. 2013;15(5):R79. doi: 10.1186/bcr3473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.To Y, et al. A two-way interaction between hepatocyte growth factor and interleukin-6 in tissue invasion of lung cancer cell line. Am J Respir Cell Mol Biol. 2002;27(2):220–226. doi: 10.1165/ajrcmb.27.2.4804. [DOI] [PubMed] [Google Scholar]
- 49.Colomiere M, et al. Cross talk of signals between EGFR and IL-6R through JAK2/STAT3 mediate epithelial-mesenchymal transition in ovarian carcinomas. Br J Cancer. 2009;100(1):134–144. doi: 10.1038/sj.bjc.6604794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Eruslanov EB, et al. Tumor-associated neutrophils stimulate T cell responses in early-stage human lung cancer. J Clin Invest. 2014;124(12):5466–5480. doi: 10.1172/JCI77053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ortiz ML, et al. Immature myeloid cells directly contribute to skin tumor development by recruiting IL-17–producing CD4+ T cells. J Exp Med. 2015;212(3):351–367. doi: 10.1084/jem.20140835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Di Mitri D, et al. Tumour-infiltrating Gr-1+ myeloid cells antagonize senescence in cancer. Nature. 2014;515(7525):134–137. doi: 10.1038/nature13638. [DOI] [PubMed] [Google Scholar]
- 53.Condamine T, Gabrilovich DI. Molecular mechanisms regulating myeloid-derived suppressor cell differentiation and function. Trends Immunol. 2011;32(1):19–25. doi: 10.1016/j.it.2010.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Bayne LJ, et al. Tumor-derived granulocyte-macrophage colony-stimulating factor regulates myeloid inflammation and T cell immunity in pancreatic cancer. Cancer Cell. 2012;21(6):822–835. doi: 10.1016/j.ccr.2012.04.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Marigo I, et al. Tumor-induced tolerance and immune suppression depend on the C/EBPβ transcription factor. Immunity. 2010;32(6):790–802. doi: 10.1016/j.immuni.2010.05.010. [DOI] [PubMed] [Google Scholar]
- 56.Gabrilovich DI, Nagaraj S. Myeloid-derived suppressor cells as regulators of the immune system. Nat Rev Immunol. 2009;9(3):162–174. doi: 10.1038/nri2506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Sinha P, Clements VK, Ostrand-Rosenberg S. Interleukin-13-regulated M2 macrophages in combination with myeloid suppressor cells block immune surveillance against metastasis. Cancer Res. 2005;65(24):11743–11751. doi: 10.1158/0008-5472.CAN-05-0045. [DOI] [PubMed] [Google Scholar]
- 58.Bronte V, et al. IL-4-induced arginase 1 suppresses alloreactive T cells in tumor-bearing mice. J Immunol. 2003;170(1):270–278. doi: 10.4049/jimmunol.170.1.270. [DOI] [PubMed] [Google Scholar]
- 59.Ko JS, et al. Direct and differential suppression of myeloid-derived suppressor cell subsets by sunitinib is compartmentally constrained. Cancer Res. 2010;70(9):3526–3536. doi: 10.1158/0008-5472.CAN-09-3278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Vasquez-Dunddel D, et al. STAT3 regulates arginase-I in myeloid-derived suppressor cells from cancer patients. J Clin Invest. 2013;123(4):1580–1589. doi: 10.1172/JCI60083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Stewart TJ, et al. Interferon regulatory factor-8 modulates the development of tumour-induced CD11b+Gr-1+ myeloid cells. J Cell Mol Med. 2009;13(9B):3939–3950. doi: 10.1111/j.1582-4934.2009.00685.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Waight JD, et al. Myeloid-derived suppressor cell development is regulated by a STAT/IRF-8 axis. J Clin Invest. 2013;123(10):4464–4478. doi: 10.1172/JCI68189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Farren MR, et al. Tumor-induced STAT3 signaling in myeloid cells impairs dendritic cell generation by decreasing PKCβII abundance. Sci Signal. 2014;7(313):ra16. doi: 10.1126/scisignal.2004656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Corzo CA, et al. HIF-1alpha regulates function and differentiation of myeloid-derived suppressor cells in the tumor microenvironment. J Exp Med. 2010;207(11):2439–2453. doi: 10.1084/jem.20100587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Noman MZ, et al. PD-L1 is a novel direct target of HIF-1alpha, and its blockade under hypoxia enhanced MDSC-mediated T cell activation. J Exp Med. 2014;211(5):781–790. doi: 10.1084/jem.20131916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Liu H, Zhou J, Cheng P, Ramachandran I, Nefedova Y, Gabrilovich DI. Regulation of dendritic cell differentiation in bone marrow during emergency myelopoiesis. J Immunol. 2013;191(4):1916–1926. doi: 10.4049/jimmunol.1300714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Capietto AH, et al. Down-regulation of PLCgamma2-beta-catenin pathway promotes activation and expansion of myeloid-derived suppressor cells in cancer. J Exp Med. 2013;210(11):2257–2271. doi: 10.1084/jem.20130281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Aliper AM, Frieden-Korovkina VP, Buzdin A, Roumiantsev SA, Zhavoronkov A. Interactome analysis of myeloid-derived suppressor cells in murine models of colon and breast cancer. Oncotarget. 2014;5(22):11345–11353. doi: 10.18632/oncotarget.2489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Mahadevan NR, Anufreichik V, Rodvold JJ, Chiu KT, Sepulveda H, Zanetti M. Cell-extrinsic effects of tumor ER stress imprint myeloid dendritic cells and impair CD8+ T cell priming. PLoS One. 2012;7(12):e51845. doi: 10.1371/journal.pone.0051845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Mahadevan NR, Rodvold J, Sepulveda H, Rossi S, Drew AF, Zanetti M. Transmission of endoplasmic reticulum stress and pro-inflammation from tumor cells to myeloid cells. Proc Natl Acad Sci U S A. 2011;108(16):6561–6566. doi: 10.1073/pnas.1008942108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Condamine T, et al. ER stress regulates myeloid-derived suppressor cell fate through TRAIL-R–mediated apoptosis. J Clin Invest. 2014;124(6):2626–2639. doi: 10.1172/JCI74056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Walter P, Ron D. The unfolded protein response: from stress pathway to homeostatic regulation. Science. 2011;334(6059):1081–1086. doi: 10.1126/science.1209038. [DOI] [PubMed] [Google Scholar]
- 73.Claudio N, Dalet A, Gatti E, Pierre P. Mapping the crossroads of immune activation and cellular stress response pathways. EMBO J. 2013;32(9):1214–1224. doi: 10.1038/emboj.2013.80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Thevenot PT, et al. The stress-response sensor chop regulates the function and accumulation of myeloid-derived suppressor cells in tumors. Immunity. 2014;41(3):389–401. doi: 10.1016/j.immuni.2014.08.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Condamine T, Gabrilovich DI. Can the suppressive activity of myeloid-derived suppressor cells be “chop”ped? Immunity. 2014;41(3):341–342. doi: 10.1016/j.immuni.2014.08.016. [DOI] [PubMed] [Google Scholar]
- 76.Lee BR, et al. Elevated endoplasmic reticulum stress reinforced immunosuppression in the tumor microenvironment via myeloid-derived suppressor cells. Oncotarget. 2014;5(23):12331–12345. doi: 10.18632/oncotarget.2589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Parker KH, et al. HMGB1 enhances immune suppression by facilitating the differentiation and suppressive activity of myeloid-derived suppressor cells. Cancer Res. 2014;74(20):5723–5733. doi: 10.1158/0008-5472.CAN-13-2347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Wu L, et al. Inhibition of PPARgamma in myeloid-lineage cells induces systemic inflammation, immunosuppression, and tumorigenesis. Blood. 2012;119(1):115–126. doi: 10.1182/blood-2011-06-363093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Narita Y, Wakita D, Ohkur T, Chamoto K, Nishimura T. Potential differentiation of tumor bearing mouse CD11b+Gr-1+ immature myeloid cells into both suppressor macrophages and immunostimulatory dendritic cells. Biomed Res. 2009;30(1):7–15. doi: 10.2220/biomedres.30.7. [DOI] [PubMed] [Google Scholar]
- 80.Yang L, et al. Expansion of myeloid immune suppressor Gr+CD11b+ cells in tumor-bearing host directly promotes tumor angiogenesis. Cancer Cell. 2004;6(4):409–421. doi: 10.1016/j.ccr.2004.08.031. [DOI] [PubMed] [Google Scholar]
- 81.Nefedova Y, Nagaraj S, Rosenbauer A, Muro-Cacho C, Sebti SM, Gabrilovich DI. Regulation of dendritic cell differentiation and antitumor immune response in cancer by pharmacologic-selective inhibition of the janus-activated kinase 2/signal transducers and activators of transcription 3 pathway. Cancer Res. 2005;65(20):9525–9535. doi: 10.1158/0008-5472.CAN-05-0529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Zhong H, et al. Origin and pharmacological modulation of tumor-associated regulatory dendritic cells. Int J Cancer. 2014;134(11):2633–2645. doi: 10.1002/ijc.28590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Sawant A, et al. Myeloid-derived suppressor cells function as novel osteoclast progenitors enhancing bone loss in breast cancer. Cancer Res. 2013;73(2):672–682. doi: 10.1158/0008-5472.CAN-12-2202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Shi Y, et al. Deficiency of Kruppel-like factor KLF4 in myeloid-derived suppressor cells inhibits tumor pulmonary metastasis in mice accompanied by decreased fibrocytes. Oncogenesis. 2014;24(3):44. doi: 10.1038/oncsis.2014.44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Niedermeier M, et al. CD4+ T cells control the differentiation of Gr1+ monocytes into fibrocytes. Proc Natl Acad Sci U S A. 2009;106(42):17892–17897. doi: 10.1073/pnas.0906070106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Zoso A, et al. Human fibrocytic myeloid-derived suppressor cells express IDO and promote tolerance via Treg-cell expansion. Eur J Immunol. 2014;44(11):3307–3319. doi: 10.1002/eji.201444522. [DOI] [PubMed] [Google Scholar]
- 87.Zhang H, et al. Fibrocytes represent a novel MDSC subset circulating in patients with metastatic cancer. Blood. 2013;122(7):1105–1113. doi: 10.1182/blood-2012-08-449413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Youn JI, et al. Epigenetic silencing of retinoblastoma gene regulates pathologic differentiation of myeloid cells in cancer. Nat Immunol. 2013;14(3):211–220. doi: 10.1038/ni.2526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Chornoguz O, Grmai L, Sinha P, Artemenko KA, Zubarev RA, Ostrand-Rosenberg S. Proteomic pathway analysis reveals inflammation increases myeloid-derived suppressor cell resistance to apoptosis. Mol Cell Proteomics. 2011;10(3):M110.002980. doi: 10.1074/mcp.M110.002980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Hu X, et al. Deregulation of apoptotic factors Bcl-xL and Bax confers apoptotic resistance to myeloid-derived suppressor cells and contributes to their persistence in cancer. J Biol Chem. 2013;288(26):19103–19115. doi: 10.1074/jbc.M112.434530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Messmer MN, Netherby CS, Banik D, Abrams SI. Tumor-induced myeloid dysfunction and its implications for cancer immunotherapy. Cancer Immunol Immunother. 2015;64(1):1–13. doi: 10.1007/s00262-014-1639-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Solito S, Marigo I, Pinton L, Damuzzo V, Mandruzzato S, Bronte V. Myeloid-derived suppressor cell heterogeneity in human cancers. Ann N Y Acad Sci. 2014;1319(1):47–65. doi: 10.1111/nyas.12469. [DOI] [PubMed] [Google Scholar]
- 93.Finke J, Ko J, Rini B, Rayman P, Ireland J, Cohen P. MDSC as a mechanism of tumor escape from sunitinib mediated anti-angiogenic therapy. Int Immunopharmacol. 2011;11(7):856–861. doi: 10.1016/j.intimp.2011.01.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Feng PH, et al. CD14(+)S100A9(+) monocytic myeloid-derived suppressor cells and their clinical relevance in non-small cell lung cancer. Am J Respir Crit Care Med. 2012;186(10):1025–1036. doi: 10.1164/rccm.201204-0636OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Vetsika E-K, et al. A circulating subpopulation of monocytic myeloid-derived suppressor cells as an independent prognostic/predictive factor in untreated non-small lung cancer patients. J Immunol Res. 2014;2014:659294. doi: 10.1155/2014/659294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Liu CY, et al. Population alterations of L-arginase- and inducible nitric oxide synthase-expressed CD11b+/CD14(–)/CD15+/CD33+ myeloid-derived suppressor cells and CD8+ T lymphocytes in patients with advanced-stage non-small cell lung cancer. J Cancer Res Clin Oncol. 2010;136(1):35–45. doi: 10.1007/s00432-009-0634-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Diaz-Montero CM, Salem ML, Nishimura MI, Garrett-Mayer E, Cole DJ, Montero AJ. Increased circulating myeloid-derived suppressor cells correlate with clinical cancer stage, metastatic tumor burden, and doxorubicin-cyclophosphamide chemotherapy. Cancer Immunol Immunother. 2009;58(1):49–59. doi: 10.1007/s00262-008-0523-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Sevko A, et al. Cyclophosphamide promotes chronic inflammation-dependent immunosuppression and prevents antitumor response in melanoma. J Invest Dermatol. 2013;133(6):1610–1619. doi: 10.1038/jid.2012.444. [DOI] [PubMed] [Google Scholar]
- 99.Segre M, Tomei E, Segre D. Cyclophosphamide-induced suppressor cells in mice: suppression of the antibody response in vitro and characterization of the effector cells. Cell Immunol. 1985;91(2):443–454. doi: 10.1016/0008-8749(85)90242-4. [DOI] [PubMed] [Google Scholar]
- 100.Meyer C, et al. Frequencies of circulating MDSC correlate with clinical outcome of melanoma patients treated with ipilimumab. Cancer Immunol Immunother. 2014;63(3):247–257. doi: 10.1007/s00262-013-1508-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Tarhini AA, et al. Immune monitoring of the circulation and the tumor microenvironment in patients with regionally advanced melanoma receiving neoadjuvant ipilimumab. PLoS One. 2014;9(2):e87705. doi: 10.1371/journal.pone.0087705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Wang Z, et al. Association of myeloid-derived suppressor cells and efficacy of cytokine-induced killer cell immunotherapy in metastatic renal cell carcinoma patients. J Immunother. 2014;37(1):43–50. doi: 10.1097/CJI.0000000000000005. [DOI] [PubMed] [Google Scholar]
- 103.Finkelstein SE, et al. Changes in dendritic cell phenotype after a new high-dose weekly schedule of interleukin-2 therapy for kidney cancer and melanoma. J Immunother. 2010;33(8):817–827. doi: 10.1097/CJI.0b013e3181ecccad. [DOI] [PubMed] [Google Scholar]
- 104.Kimura T, et al. MUC1 vaccine for individuals with advanced adenoma of the colon: a cancer immunoprevention feasibility study. Cancer Prev Res. 2013;6(1):18–26. doi: 10.1158/1940-6207.CAPR-12-0275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Wesolowski R, Markowitz J, Carson W. Myeloid derived suppressor cells - a new therapeutic target in the treatment of cancer. J Immunother Cancer. 2013;1(1):10. doi: 10.1186/2051-1426-1-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Suzuki E, Kapoor V, Jassar AS, Kaiser LR, Albelda SM. Gemcitabine selectively eliminates splenic Gr-1+/CD11b+ myeloid suppressor cells in tumor-bearing animals and enhances antitumor immune activity. Clin Cancer Res. 2005;11(18):6713–6721. doi: 10.1158/1078-0432.CCR-05-0883. [DOI] [PubMed] [Google Scholar]
- 107.Le HK, Graham L, Cha E, Morales JK, Manjili MH, Bear HD. Gemcitabine directly inhibits myeloid derived suppressor cells in BALB/c mice bearing 4T1 mammary carcinoma and augments expansion of T cells from tumor-bearing mice. Int Immunopharmacol. 2009;9(7–8):900–909. doi: 10.1016/j.intimp.2009.03.015. [DOI] [PubMed] [Google Scholar]
- 108.Gujar SA, Clements D, Dielschneider R, Helson E, Marcato P, Lee PW. Gemcitabine enhances the efficacy of reovirus-based oncotherapy through anti-tumour immunological mechanisms. Br J Cancer. 2014;110(1):83–93. doi: 10.1038/bjc.2013.695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Naiditch H, Shurin MR, Shurin GV. Targeting myeloid regulatory cells in cancer by chemotherapeutic agents. Immunol Res. 2011;50(2–3):276–285. doi: 10.1007/s12026-011-8213-2. [DOI] [PubMed] [Google Scholar]
- 110.Vincent J, et al. 5-Fluorouracil selectively kills tumor-associated myeloid-derived suppressor cells resulting in enhanced T cell-dependent antitumor immunity. Cancer Res. 2010;70(8):3052–3061. doi: 10.1158/0008-5472.CAN-09-3690. [DOI] [PubMed] [Google Scholar]
- 111.Ugel S, et al. Immune tolerance to tumor antigens occurs in a specialized environment of the spleen. Cell Rep. 2012;2(3):628–639. doi: 10.1016/j.celrep.2012.08.006. [DOI] [PubMed] [Google Scholar]
- 112.Qin H, et al. Generation of a new therapeutic peptide that depletes myeloid-derived suppressor cells in tumor-bearing mice. Nat Med. 2014;20(6):676–681. doi: 10.1038/nm.3560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Nagaraj S, et al. Anti-inflammatory triterpenoid blocks immune suppressive function of MDSCs and improves immune response in cancer. Clin Cancer Res. 2010;16(6):1812–1823. doi: 10.1158/1078-0432.CCR-09-3272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Serafini P, et al. Phosphodiesterase-5 inhibition augments endogenous antitumor immunity by reducing myeloid-derived suppressor cell function. J Exp Med. 2006;203(12):2691–2702. doi: 10.1084/jem.20061104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Califano JA, et al. Tadalafil augments tumor specific immunity in patients with head and neck squamous cell carcinoma. Clin Cancer Res. 2015;21(1):30–38. doi: 10.1158/1078-0432.CCR-14-1716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Weed DT, et al. Tadalafil reduces myeloid-derived suppressor cells and regulatory T cells and promotes tumor immunity in patients with head and neck squamous cell carcinoma. Clin Cancer Res. 2015;21(1):39–48. doi: 10.1158/1078-0432.CCR-14-1711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Rodriguez PC, et al. Arginase I in myeloid suppressor cells is induced by COX-2 in lung carcinoma. J Exp Med. 2005;202(7):931–939. doi: 10.1084/jem.20050715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.De Santo C, et al. Nitroaspirin corrects immune dysfunction in tumor-bearing hosts and promotes tumor eradication by cancer vaccination. Proc Natl Acad Sci U S A. 2005;102(11):4185–4190. doi: 10.1073/pnas.0409783102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Kusmartsev S, et al. All-trans-retinoic acid eliminates immature myeloid cells from tumor-bearing mice and improves the effect of vaccination. Cancer Res. 2003;63(15):4441–4449. [PubMed] [Google Scholar]
- 120.Kusmartsev S, et al. Reversal of myeloid cell-mediated immunosuppression in patients with metastatic renal cell carcinoma. Clin Cancer Res. 2008;14(24):8270–8278. doi: 10.1158/1078-0432.CCR-08-0165. [DOI] [PubMed] [Google Scholar]
- 121.Mirza N, et al. All-trans-retinoic acid improves differentiation of myeloid cells and immune response in cancer patients. Cancer Res. 2006;66(18):9299–9307. doi: 10.1158/0008-5472.CAN-06-1690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Iclozan C, Antonia S, Chiappori A, Chen DT, Gabrilovich D. Therapeutic regulation of myeloid-derived suppressor cells and immune response to cancer vaccine in patients with extensive stage small cell lung cancer. Cancer Immunol Immunother. 2013;62(5):909–918. doi: 10.1007/s00262-013-1396-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Sansone P, Bromberg J. Targeting the interleukin-6/Jak/stat pathway in human malignancies. J Clin Oncol. 2012;30(9):1005–1014. doi: 10.1200/JCO.2010.31.8907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Ko JS, et al. Sunitinib mediates reversal of myeloid-derived suppressor cell accumulation in renal cell carcinoma patients. Clin Cancer Res. 2009;15(6):2148–2157. doi: 10.1158/1078-0432.CCR-08-1332. [DOI] [PubMed] [Google Scholar]