

Highlights
-
•
22q11DS offers a compelling model to understand the neural substrates of attentional dysfunction.
-
•
First study directly comparing neural function in 22q11DS vs. ADHD patients
-
•
22q11DS and ADHD patients show a shared deficit in RI-related activation.
-
•
ADHD patients showed greater activity in the middle frontal gyrus than 22q11DS during RI.
-
•
Neural activity is inversely correlated with self-reported Cognitive Impulsivity in 22q11DS.
1. Introduction
Attention deficit hyperactivity disorder (ADHD) is a behaviorally defined disorder characterized by a persistent pattern of inattention and/or hyperactivity–impulsivity that is more frequent and severe than typically observed in individuals at a comparable level of development (American Psychiatric Association, 2013). Substantial heterogeneity – at both the genetic and phenotypic level – has significantly impeded our understanding of the underlying biological mechanisms of ADHD. As the past two decades of research on causal genetic and neurobiological risk factors for this illness have been met with limited success (for reviews, see Elia and Devoto, 2007; Franke et al., 2009), one promising strategy for investigating the pathogenesis of ADHD involves the investigation of a disorder with known genetic etiology that shares similar phenotypic characteristics — an approach that is supported by findings that large, rare chromosomal deletions and duplications (copy number variants; CNVs) are implicated in ADHD etiology (Williams et al., 2010). 22q11.2 Deletion Syndrome (22q11DS; velocardiofacial syndrome) is a particularly compelling model, as it is a relatively common CNV that is highly penetrant for ADHD; approximately 40% of 22q11DS patients meet clinical criteria for the disorder (Green et al., 2009). However, it is not yet known whether the neural substrates of attentional dysfunction in 22q11DS are shared with idiopathic ADHD.
Impaired response inhibition (RI) has been consistently implicated as a hallmark of idiopathic ADHD (Dickstein et al., 2006; Durston, 2003; Epstein et al., 2007; Schulz et al., 2004; Tamm et al., 2004; van Rooij et al., 2015). Most commonly measured with either Go/No-Go or Stop-signal task paradigms, successful inhibition of a prepotent response has been shown to rely upon engagement of a right-lateralized fronto-striatal network in healthy individuals (Aron and Poldrack, 2006; Aron et al., 2007; Chambers et al., 2009). Studies on the neural underpinnings of RI in adults with ADHD have yielded inconsistent results, with some studies reporting less RI-related activation in ADHD patients than controls (e.g., Mulligan et al., 2011; Sebastian et al., 2012; van Rooij et al., 2015) and others either reporting no differences in activity (Carmona et al., 2012; Congdon et al., 2010) or greater activity for ADHD patients as compared to controls (Dillo et al., 2010; Karch et al., 2010). However, the bulk of the evidence suggests a disturbed pattern of RI-related neural activation in adults with ADHD. Discrepancies in previous findings may be attributable to several factors, including differences in task parameters, medication status, symptom severity, and small sample size.
Consistent with findings of impaired performance on RI-related behavioral tasks in 22q11DS (Shapiro et al., 2013), using functional magnetic resonance imaging (fMRI) we previously found that young adults with 22q11DS show significantly less activation in RI-related regions during successful RI relative to healthy controls (Montojo et al., 2015). Further, we found that RI-related activity within the left middle frontal gyrus and basal ganglia was associated with the severity of self-reported Cognitive Impulsivity, suggesting that reduced engagement of RI-related regions in 22q11DS may be relevant to the behavioral manifestations of the disorder.
Despite symptomatic overlap between 22q11DS and idiopathic ADHD, it is not yet known whether disturbances in the neural circuitry underlying inhibitory control difficulties are shared between these clinical populations. Using fMRI, we examined neural activity in patients with 22q11DS, idiopathic ADHD, and demographically comparable healthy controls during performance of a Stop-signal task. We hypothesized that, relative to controls, individuals with 22q11DS and those with idiopathic ADHD would show reduced recruitment of fronto-striatal regions involved in inhibitory control. In addition, we explored the relationship of neural activity during task performance to dimensional measures of cognitive and Behavioral Impulsivity, to determine whether brain–behavior associations were similar across groups. Based on prior literature in 22q11DS and ADHD (Montojo et al., 2015; van Rooij et al., 2015) we hypothesized that, while 22q11DS and idiopathic ADHD patients would share similar dysfunction within RI-related neural systems, this shared dysfunction would show different relationships to behavior. Specifically, we hypothesized that 22q11DS patients would show an inverse relationship between neural activity during RI and Cognitive Impulsivity (Montojo et al., 2015). Given the phenotypic and genetic heterogeneity known to exist within the ADHD population, we hypothesized that we would not observe a significant relationship between neural activity and cognitive or Behavioral Impulsivity within this group.
2. Material and methods
2.1. Participants
Seventy-two participants (19 patients with 22q11DS, 23 with an ADHD diagnosis, and 30 healthy adults) were included in the study. 22q11DS participants include individuals with a molecularly confirmed diagnosis of 22q11.2 Deletion Syndrome recruited from an ongoing longitudinal study at the University of California Los Angeles (UCLA). Individuals with ADHD were recruited from the Los Angeles area as part of the Consortium for Neuropsychiatric Phenomics (CNP) at UCLA. The current study includes a partially overlapping subset of patients with 22q11DS and ADHD that were included in previous publications (Congdon et al., 2014; Montojo et al., 2015). Four 22q11DS participants whose data were collected for the Stop-signal task were excluded from analysis for the following reasons: low signal-to-noise ratio (n = 1), too much translational movement (n = 1), and poor registration between functional and anatomical scans (n = 2). Thus, our final sample consisted of 15 patients with 22q11DS, 23 with ADHD, and 30 healthy adults. Demographic information for each group is presented in Table 1. All diagnoses were determined using the Structured Clinical Interview for the Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition (DSM-IV) (SCID; First et al., 2004) by trained raters who participated in an ongoing quality assurance program (Ventura et al., 1998). ADHD diagnoses in the CNP sample were confirmed with the Adult ADHD Interview (a structured interview derived from the Kiddie Schedule for Affective Disorders and Schizophrenia, Present and Lifetime Version (KSADS-PL) (Kaufman et al., 1997)), in order to provide a more detailed characterization of lifetime history of ADHD in adults. Prior to conducting diagnostic interviews, clinicians had to achieve good to excellent reliability with a set of gold standard ratings (kappa coefficients of .90 or greater for categorical diagnoses and intra-class correlation coefficients (ICCs) ranging from 0.85 to 1.00 for quantitative symptom ratings). Diagnostic formulations for all 22q11DS cases were additionally reviewed in consensus diagnosis meetings led by the study director, a board-certified psychologist (CEB). Training, reliability and ongoing quality assurance procedures for psychiatric diagnostic assessments are detailed in a prior publication (Jalbrzikowski et al., 2013).
Table 1.
Demographic and clinical characteristics of study participants.
| 22q11.2 participants (n = 15) |
ADHD participantsa (n = 23) |
Control participants (n = 30) |
p-Value |
Tukey's HSD post-hoc contrast p-value |
|
|---|---|---|---|---|---|
| Age (years, ±SD) |
22.73 (5.25) |
24.28 (2.97) | 23.00 (4.35) |
p = 0.341 |
NA |
| Participant education (years, ±SD) |
12.33 (1.23) |
14.91 (1.90) | 14.60 (1.73) |
p < 0.001 |
22q11DS < Con(p < 0.01)ADHD = Con(p = 0.785)ADHD > 22q11DS(p < 0.01) |
| Gender (N, % female) |
7 (47%) |
12 (52%) | 12 (40%) |
p = 0.675 |
NA |
| Ethnicity (N, % Latino) |
1 (6%) |
6 (17%) | 10 (33%) |
p = 0.132 |
NA |
| Full scale IQ (mean, ±SD)b | 75.00 (14.36) | 120.60 (16.88) | 123.93 (12.39) |
p < 0.001 |
22q11DS < Con(p < 0.01)ADHD = Con(p = 0.687)ADHD > 22q11DS(p < 0.01) |
| Barratt Impulsivity Scale: Cognitive Impulsivity (mean, ±SD)c |
7.97 (2.19) | 8.74 (1.02) | 6.09 (1.56) | p < 0.001 | 22q11DS > Con(p < 0.01)ADHD > Con(p < 0.01)ADHD = 22q11DS(p = 0.309) |
| Barratt Impulsivity Scale: Behavioral Impulsivity (mean, ±SD)c |
5.38 (2.05) | 8.00 (1.55) | 6.01 (1.03) | p < 0.001 | 22q11DS = Con(p = 0.425)ADHD > Con(p < 0.01)ADHD > 22q11DS(p < 0.01) |
| Psychotropic medication (N, none/antidepressant/psychostimulant/anti-anxiety/antipsychotic)d |
7/5/3/2/1 | 15/2/7/0/0 | 28/2/0/0/0 |
χ2 for ADHD vs. 22q11DS psycho-stimulants: p = 0.475 |
NA |
ADHD participants met criteria for the following subtypes: Predominantly Hyperactive–Impulsive Type (1 participant), Combined Type (10 participants), Predominantly Inattentive Type (12 participants).
Based on 2-subtest Wechsler Abbreviated Scale of Intelligence (Vocabulary and Matrix Reasoning).
BIS measures were available for 23/30 control participants, p-values are based on ANCOVAs controlling for age and years of education. Tukey's HSD post-hoc contrast p-values represent pairwise contrasts run following the ANOVA.
Three 22q11DS and two ADHD participants were prescribed multiple psychotropic medications.
Demographically comparable healthy controls were recruited from these two studies. Exclusion criteria for all study participants included the following: neurological or medical condition that might affect performance, insufficient fluency in English, substance or alcohol abuse and/or dependence with the past 6 months, any contraindications to scanning, and left-handedness (further details are provided in the Supplementary material). The SCID was used to ensure that healthy controls did not meet criteria for any current major mental disorder (see the Supplementary material and Thakkar et al., 2014 for additional details of inclusion/exclusion criteria). Stable medications were permitted in 22q11DS and ADHD participants, as it was not ethically permissible to ask participants to stop taking physician-prescribed medication in order to participate in the study. Any psychoactive medication use by controls was an exclusion factor.
All participants underwent a verbal and written informed consent process. The UCLA Institutional Review Board (IRB) approved all study procedures and informed consent documents.
2.2. Procedure
In both studies, after screening and neuropsychological assessment, participants took part in a behavioral testing session immediately prior to a 1-hour scan. In the behavioral testing session, participants received training on the Stop-signal task in the form of one initial demonstration and trial run before completing one experimental run while inside the scanner. Stimulus presentation and timing of all stimuli and response events were achieved using Matlab (Mathworks) and the Psychtoolbox (http://www.psychtoolbox.org, Brainard, 1997) on an Apple Powerbook. For the experiment block administered in the scanner, each participant viewed the task through MRI-compatible goggles and responded with his or her right hand on an MR-compatible button box in the scanner.
2.3. Measures
2.3.1. Stop-signal task
Participants completed a tracking version of the Stop-signal task (Logan, 1994) enabled isolation of neural activation associated with the inhibition of an already-initiated motor response, and calculation of an individualized measure of inhibitory control (stop-signal reaction time, SSRT). Participants were instructed to respond quickly when a “go” stimulus was presented on the computer screen (which consisted of left- or right-ward pointing arrows), except on the subset of trials where the “go” stimulus was followed by a “stop” signal (a 500 Hz tone presented through headphones), in which case participants were instructed to withhold their response. The onset of the stop-signal, or stop-signal delay (SSD), was adjusted according to the participant's performance – such that the SSD increased following a successful inhibition (making the next trial more difficult) and decreased following a failed inhibition (making the next trial easier) – which ensured that subjects successfully inhibited on approximately 50% of stop trials. A complete description of the task and fMRI acquisition parameters is presented in the Supplementary material.
2.3.2. Barratt Impulsiveness Scale
The Barratt Impulsiveness Scale (BIS) Version 11 (BIS-11; Patton et al., 1995) is a 30-item questionnaire assessing aspects of impulsivity which has been applied to various clinical groups, including those with ADHD (Crunelle et al., 2013; Malloy-Diniz et al., 2007). In addition to calculating a Total BIS Score, we used the revised, two-factor scoring method of the BIS, which has been proposed as an alternative multidimensional structural representation of impulsivity (Reise et al., 2013). The revised method of scoring the BIS results into two correlated factors: (1) Cognitive Impulsivity, reflecting difficulties in attentional control, concentration, careful and deliberate thinking, and planning (e.g., “not a steady thinker”, “no self-control/concentration” and “not planful”), and (2) Behavioral Impulsivity (with some cognitive elements), which reflects acting impulsively, changing jobs, moving residences relatively often, and a scattered quick-paced cognitive tempo (e.g., “extraneous racing thoughts”, “acts impulsively”, and “changes, moves around”). We note that while the BIS-11 measures an individual's subjective view of his/her own behavior, there is a strong correlation between scores on this measure and clinical diagnoses of ADHD (Malloy-Diniz et al., 2007). For both scales, higher scores reflect higher levels of trait impulsivity.
2.3.3. Neurocognitive measures
Supervised clinical psychology doctoral students or PhD staff administered a comprehensive neurocognitive battery. IQ data were acquired for all participants using the Wechsler Abbreviated Scale of Intelligence (WASI; Wechsler, 1999) or the Wechsler Adult Intelligence Scale (WAIS-IV; Wechsler, 2008).
2.4. Behavioral data analysis
Stop-signal task data were analyzed as has been previously described (Congdon et al., 2010, 2012) and as detailed in the Supplementary material. Briefly, SSRT was estimated using the quantile method (Band et al., 2003), with longer SSRT values reflecting poorer inhibitory control. Additional performance measures included mean and standard deviation of reaction time (RT) on Go trials, percent inhibition on Stop trials (i.e., successful inhibition), and percent correct on Go trials.
To examine demographic differences between the three groups (22q11DS vs. ADHD vs. controls) we conducted univariate ANOVAs for continuous variables or chi-square tests for categorical variables. Pearson correlations were conducted to assess the relationship between demographics and Stop-signal task performance. To examine group differences with regard to Stop-signal task performance and BIS-11 scores, we conducted ANOVAs for each measure, with group as a fixed factor. As IQ and years of education showed significant differences between groups, ANCOVAs were also conducted with years of education included as a covariate, in order to verify ANOVA results (IQ was omitted given the high correlation between years of education and IQ: r(68) = 0.577, p < 0.01). Age was also included as a covariate given that this variable showed associations with task performance (see Results section). All analyses were conducted using SPSS software v. 21 (IBM).
2.5. fMRI data analysis
Analyses were performed using tools from the FMRIB software library (http://www.fmrib.ox.ac.uk/fsl), version 5.0 (S.M. Smith et al., 2004), and preprocessing steps are outlined in the Supplementary material. For each subject, Stop Successful–Go, Go–Null, and Stop Unsuccessful–Stop Successful contrasts were computed, and the output from the subject-specific analyses was then analyzed using a mixed-effects model with FLAME for between-group comparisons. To examine between-group differences, group-level statistics images were thresholded with a cluster-forming threshold of Z > 2.3 and a cluster probability of p < 0.05, corrected for whole-brain multiple comparisons using the Gaussian random field theory. As our primary goal was to examine between-group differences in RI-related activation, and as patterns associated with RI have been previously reported in each group alone (Congdon et al., 2014; Montojo et al., 2015), we present below only the results from Stop Successful–Go between-group comparisons. The whole-brain, between-group contrasts included the following: controls > 22q11DS, controls > ADHD, 22q11DS > ADHD, and ADHD > 22q11DS. Results for additional task contrasts are presented in the Supplementary material.
2.5.1. Conjunction analysis
In order to identify differences in neural activity during successful RI that were common to both clinical groups relative to controls, we used activity maps for the contrasts of controls > 22q11DS and controls > ADHD and performed a conjunction analysis to define a set of significantly overlapping regions between these two contrasts. We then extracted percent signal change values from this overlapping set of regions in order to visualize the patterns of group differences in neural activation for these regions of overlap (for details, see methods for Visualizing group differences below).
2.5.2. Relationship between inhibition-related neural activation and trait impulsivity
In order to investigate correlates of RI-associated neural activity in these clinical groups, we characterized relationships between neural activity and BIS-11 scores using whole-brain regressions, within 22q11DS and ADHD patients only, given the limited range of BIS-11 scores within control participants. First, to test for differences in the relationship between RI-related activation and BIS-11 scores between the 22q11DS and ADHD groups (i.e., an interaction effect), whole-brain regressions were defined for the Stop Successful–Go contrast with either BIS Cognitive or Behavioral Impulsivity subscale scores and group membership as covariates of interest, including age and sex as covariates of no interest. Second, to further explore the pattern of association between RI-related activation and BIS-11 scores within the 22q11DS and ADHD patient groups separately – without regard to group differences in these relationships – we defined whole-brain regressions for the Stop Successful–Go contrast with either BIS Cognitive or Behavioral Impulsivity subscale scores as covariates of interest, including age and sex as covariates of no interest. Each whole-brain regression was thresholded with a cluster-forming threshold of Z > 2.3 and a cluster probability of p < 0.05, corrected for whole-brain multiple comparisons using the Gaussian random field theory. This approach allowed us to: 1) test whether there were differences in the association between neural activity during task performance and BIS scores between groups (22q11DS > ADHD; ADHD > 22q11DS); and 2) subsequently investigate the associations between neural activity and BIS scores within each group alone, in order to understand the directionality of any significant interaction effects.
2.5.3. Visualizing common neural substrates of RI and clinical correlations
In order to visualize: 1) the common regions of reduced RI-related neural activity across both patient groups relative to controls; and 2) the relationship between RI-related neural activity and BIS-11 scores within each group, we extracted percent signal change values corresponding to a 1-s stimulus convolved with a double-gamma hemodynamic response function from each contrast of interest (following Mumford and Poldrack, 2007) following inspection of group contrasts. For the conjunction analysis, after inspecting clusters of activation that significantly differed between both patient groups as compared with controls, we intersected the group conjunction map with an anatomical atlas (the FSL Harvard–Oxford probabilistic atlas thresholded at 25%) to extract percent signal change from anatomically-constrained regions that contained the clusters of activation identified in the conjunction map. The anatomically constrained regions that contained these clusters of activation included the anterior cingulate cortex (ACC), cerebellum, inferior parietal lobe, middle frontal gyrus, middle temporal gyrus (posterior), paracingulate gyrus, superior frontal gyrus, and left and right thalamus. By intersecting our group conjunction map with an anatomical atlas, we were able to extract percent signal change from multiple clusters within anatomically-defined regions.
In order to visualize the relationship between stopping-related activation and self-reported impulsivity within each group, percent signal change was extracted from significant clusters identified in each whole brain regression of Stop Successful–Go activity on impulsivity scores. The resulting percent signal change values were then plotted for visualization purposes.
2.5.4. Secondary analyses
Follow-up analyses were conducted to investigate: 1) the effects of psychostimulant use and IQ on fMRI results, and 2) the effects of group differences in performance on Go trials on RI-related neural activity (see the Supplementary material).
3. Results
3.1. Behavioral results
3.1.1. Demographic and clinical characteristics
The total sample consisted of 68 participants (15 22q11DS, 23 ADHD, and 30 healthy adults, age 18–38 years old). As shown in Table 1, 22q11DS, ADHD, and control groups were matched on all demographic factors except for years of education and IQ. ADHD patients and controls had significantly higher IQ and more years of education than 22q11DS patients. There was no significant difference in years of education or IQ between ADHD patients and controls. Stable medications were permitted in 22q11DS and ADHD patients. Among the 22q11DS patients, 33% (5 out of 15) had a clinical diagnosis of ADHD, and 20% (3 out of 15) were taking psychostimulants. In ADHD patients, 40% (8 out of 23) reported currently taking psychostimulants.
3.1.2. Between-group comparisons of Stop-signal task performance
As shown in Table 2, there were no significant differences in SSRT between the three groups and − given the tracking nature of the task design – percent inhibition on Stop trials was close to 50% in each group. However, there were significant differences in percent correct on Go trials and median reaction time (RT) on Go trials, with ADHD patients and controls showing higher accuracy and faster response times on Go trials as compared to 22q11DS patients. There were no significant differences between patients with ADHD and controls in accuracy or median RT on Go trials. These results remained unchanged when controlling for age and years of education.
Table 2.
Behavioral performance on stop signal task: ADHD, 22q11DS participants and healthy controls.
| Stop signal fMRI task: behavioral results | 22q11.2 participants (n = 15) |
ADHD participants (N = 23) |
Control participants (N = 30) | F-value |
p-Value |
Tukey's HSD post-hoc contrast p-value |
|---|---|---|---|---|---|---|
| Median Go RT (ms, ±SD) | 574.67 (163.88) | 458.28 (83.47) | 440.80 (81.35) | 5.260 | p = 0.008 | 22q11DS > Con(p < 0.01)ADHD = Conp = 0.821ADHD < 22q11DS(p < 0.01) |
| Go trials: % correct (%, ±SD) |
75.98% (8.93) | 98.10% (6.51) | 92.46% (11.44) | 13.99 | p < 0.001 | 22q11DS < Con(p < 0.01)ADHD = Conp = 0.089ADHD > 22q11DS(p < 0.01) |
| Stop trials: % inhibition (%, ±SD) |
53.54% (10.49) | 49.46% (8.41) | 49.59% (10.24) | 1.266 | p = 0.289 | NA |
| SSRT RT (ms, ±SD) |
194.04 (60.71) | 189.32 (58.49) | 167.17 (71.08) | 0.709 | p = 0.496 | NA |
We covaried for age and participant education in group comparisons of task performance variables, as age was associated with task performance and there was a significant difference between groups in years of education. The F-value and associated p-values in the table represent results from the ANCOVA. Tukey's HSD post-hoc contrast p-values represent pairwise contrasts run following on the ANOVA.
3.1.3. Demographic associations with task performance
Across the three groups, there was a significant relationship between percent correct on Go trials with age (r = 0.365, p < 0.01) and years of education (r = 0.575, p < 0.01). Participants who were older or had more years of education performed with higher accuracy on Go trials relative to younger or less educated participants. There was no association between any of the other demographic or performance variables (see the Supplementary material).
3.1.4. Between-group comparisons of trait impulsivity
As shown in Table 1 and Supplementary Fig. 1, there were significant group differences in Cognitive Impulsivity. Specifically, 22q11DS and ADHD patients showed significantly elevated Cognitive Impulsivity scores relative to controls, while there were no significant differences between ADHD and 22q11DS patients. There were also significant group differences in Behavioral Impulsivity. Specifically, ADHD patients showed significantly elevated Behavioral Impulsivity scores relative to 22q11DS patients and controls, while there was no significant difference between 22q11DS patients and controls. These results remained unchanged when controlling for years of education (see the Supplementary material).
3.2. fMRI results
3.2.1. Successful stopping: contrasts of clinical groups compared to controls
As shown in Fig. 1 and Table 3, a direct comparison between controls and 22q11DS patients during successful RI (Stop Successful–Go contrast) revealed significantly increased activation in controls relative to 22q11DS patients in the bilateral medial frontal cortex/ACC/paracingulate gyrus, bilateral middle/inferior/superior frontal gyrus, bilateral occipital and parietal regions, right middle temporal gyrus, posterior cingulate cortex (PCC), bilateral basal ganglia (thalamus) and cerebellum. There were no regions showing greater activation for 22q11DS patients as compared to controls. A direct comparison between controls and ADHD patients revealed a similar pattern of increased activation for controls relative to ADHD patients in the medial frontal cortex/ACC paracingulate, right middle frontal gyrus, bilateral inferior parietal cortex, bilateral middle temporal gyrus, bilateral occipital cortex, bilateral basal ganglia (thalamus), and cerebellum, while there were no regions showing greater activation for ADHD patients as compared to controls.
Fig. 1.

Neural activity during successful RI (Stop-Successful vs. Go contrast): 22q11DS and ADHD participants versus controls. (a) Green clusters represent activity from the contrast of controls > 22q11DS, and dark blue clusters represent activity from the contrast of controls > ADHD. (b) Red colors represent regions showing a conjunction of neural activity from the contrasts of controls > 22q11DS and controls > ADHD.
Table 3.
Regions showing between-group differences in activation for patients with ADHD, 22q11DS, and healthy controls during successful stopping (Stop successful vs. Go). Montreal Neurological Institute (MNI) coordinates represent the location of maximum activation for significant clusters.
| Trial Type | Contrast | Region | Voxel # | Max Z-Score | Max X (mm) | Max Y (mm) | Max Z (mm) |
|---|---|---|---|---|---|---|---|
| Successful Stopping | |||||||
| Controls > 22q11DS | |||||||
| Right middle/superior and medial frontal gyrus | 2731 | 5.32 | 16 | 12 | 64 | ||
| Bilateral caudate, thalamus | 2112 | 4.21 | 8 | -4 | 14 | ||
| Left lateral occipital cortex | 2008 | 4.37 | -8 | -80 | -40 | ||
| Left middle, inferior frontal gyrus | 1499 | 4.72 | -40 | 24 | 44 | ||
| Right middle temporal gyrus | 1105 | 4.52 | 70 | -22 | -12 | ||
| Right lateral occipital cortex | 1092 | 4.4 | 40 | -68 | -44 | ||
| Right posterior cingulate gyrus, precuneus cortex | 943 | 5.11 | 10 | -68 | 44 | ||
| Left lateral parietal cortex | 461 | 4.94 | -44 | -52 | 48 | ||
| Cerebellum | 433 | 4.04 | -8 | -54 | -42 | ||
| Controls > ADHD | |||||||
| Medial frontal cortex, right middle frontal gyrus, cingulate cortex | 2558 | 4.51 | -2 | 20 | 62 | ||
| Left Cerebellum | 931 | 4.61 | -6 | -86 | -32 | ||
| Right middle temporal gyrus | 900 | 3.87 | 60 | -46 | 4 | ||
| Left middle temporal gyrus | 633 | 4.23 | -46 | -54 | 12 | ||
| Thalamus | 463 | 3.82 | -14 | -6 | 10 | ||
| Left lateral parietal cortex, angular gyrus | 343 | 3.9 | -42 | -50 | 42 | ||
| Right lateral parietal cortex, occipital fusiform gyrus | 341 | 4.25 | 28 | -88 | -14 | ||
| ADHD > 22q11DS | |||||||
| Left middle frontal gyrus | 491 | 3.57 | -24 | 38 | 32 | ||
| Right middle frontal gyrus | 381 | 4.18 | 30 | 48 | 40 | ||
3.2.2. Conjunction analysis: common substrates of RI-related activity in 22q11DS and ADHD
As controls showed greater RI-related activation than both 22q11DS and ADHD patients across multiple brain regions, we conducted a conjunction analysis in order to determine the regions common to both clinical groups (Price and Friston, 1997). As illustrated in Fig. 1, conjunction regions that showed overlap for 22q11DS patients and ADHD patients compared to controls included the medial frontal cortex/anterior cingulate/paracingulate gyrus, right middle frontal gyrus, left inferior parietal cortex, cerebellum (bilaterally), thalamus (bilaterally), and right middle temporal gyrus. Corresponding percent signal change plots are illustrated in Fig. 2, showing that controls show greater activity than both ADHD and 22q11DS patients within RI-related regions, including the ACC, bilateral cerebellum, left inferior parietal cortex, right middle frontal gyrus, right middle temporal gyrus, paracingulate gyrus, superior frontal gyrus, and bilateral thalamus.
Fig. 2.

Conjunction analysis showing significant differences in RI-related neural activity common to 22q11DS and ADHD patients compared to controls. Nine anatomically-defined ROIs were intersected with the conjunction group map to extract percent signal change. The X-axis represents the group (controls = gray, ADHD patients = dotted, 22q11DS = black) and the Y-axis represents percent signal change.
3.2.3. Successful stopping: idiopathic ADHD compared to 22q11DS
As shown in Fig. 3 and Table 3, a direct comparison between ADHD and 22q11DS patients revealed significantly increased activation that was specific to the bilateral middle frontal gyrus for patients with ADHD relative to those with 22q11DS. There were no regions showing greater activation for 22q11DS patients as compared with ADHD patients. Since we observed a significant difference in performance for Go trials, which may affect the neural activity findings for the Stop Successful–Go contrast, we conducted a follow-up analysis on the Stop Successful–Null contrast and found similar results to those observed for the Stop Successful–Go contrast (see the Supplementary material for details).
Fig. 3.
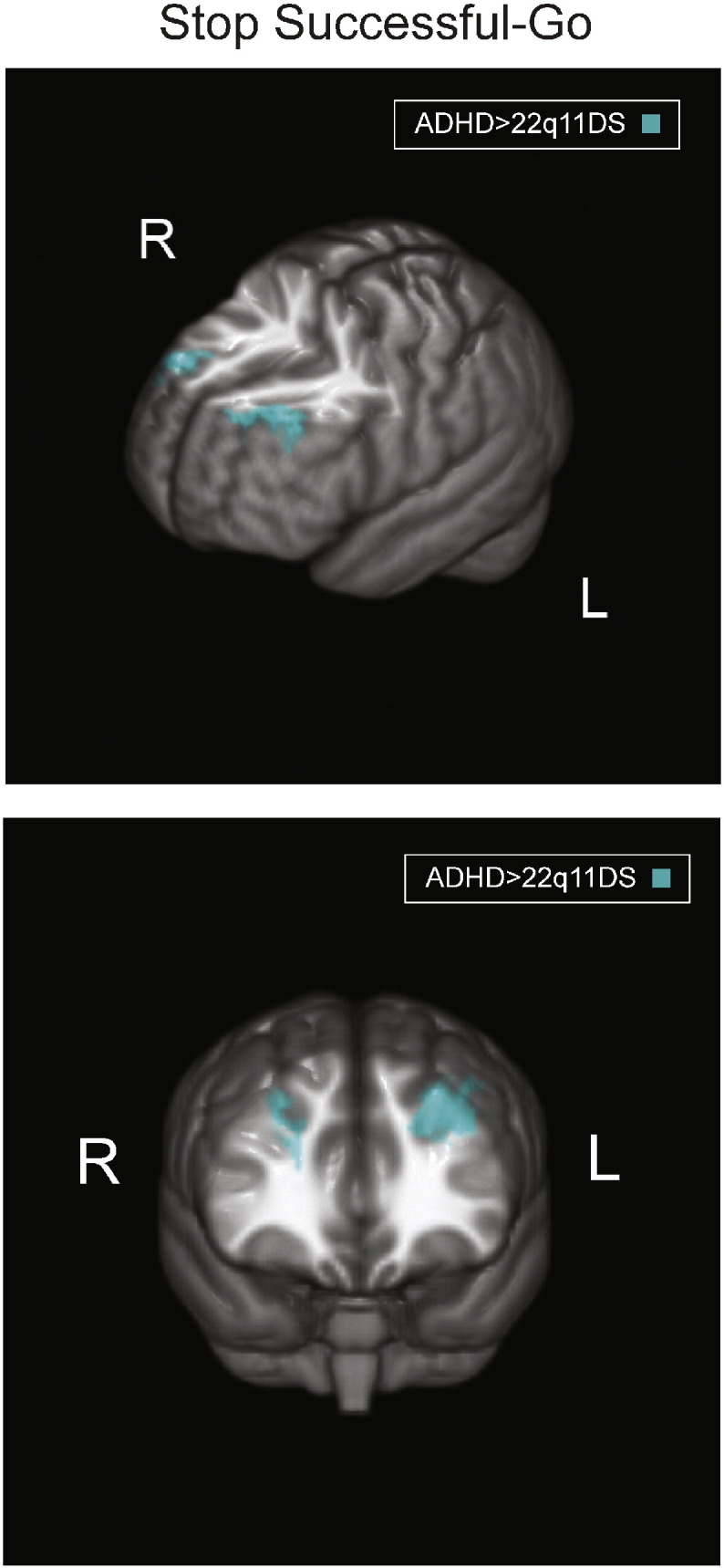
Neural activity differences for successful stopping: 22q11DS versus ADHD. Turquoise maps represent activity from the contrast of Stop Successful–Go, ADHD > 22q11DS, to investigate differences in activity related to response inhibition. Brain orientations are labeled such that S = superior, I = inferior, P = posterior, and A = anterior; R = right and L = left.
3.2.4. Relationship between RI-related activation and trait impulsivity
To further explore differences in RI-related activation between clinical groups, we examined the relationship between RI-related activation and trait impulsivity as a function of group status (22q11DS vs. ADHD). As illustrated in Fig. 4a and detailed in Table 4, a significant interaction was observed between group and Cognitive Impulsivity, such that 22q11DS patients showed an inverse correlation between activity in the medial frontal cortex and precuneus with Cognitive Impulsivity, while ADHD patients showed a positive correlation in these same regions. Follow-up regressions within 22q11DS patients alone confirmed a negative association between Cognitive Impulsivity and RI-related activation in the medial frontal gyrus, precuneus/posterior cingulate cortex, and inferior parietal cortex (Table 4). No regions were positively associated with Cognitive Impulsivity in the 22q11DS patients, and no significant correlation with Cognitive Impulsivity was evident in the ADHD group alone after correction for multiple comparisons.
Fig. 4.

Relationship between RI-related neural activity and trait impulsivity in 22q11DS and ADHD patients. Activity maps display regions that showed a significant relationship between Stop Successful–Go activity and trait impulsivity values, using the following color scheme: turquoise = ADHD > 22q11DS, yellow = 22q11DS alone, and magenta = 22q11DS > ADHD. No regions showed a significant correlation within ADHD alone. Brain orientations are labeled such that S = superior, I = inferior, P = posterior, and A = anterior; R = right and L = left. The X-axis values represent the (a) Cognitive Impulsivity subscore or (b) Behavioral Impulsivity subscore of the Barratt Impulsivity Scale, and the Y-axis values represent the percent signal change during Stop Successful–Go (with age and years of education included in the analysis as covariates of non-interest). Higher scores on cognitive or Behavioral Impulsivity scales indicate higher levels of trait impulsivity. ADHD patient data are shown as open circles, 22q11DS patient data are shown as black squares, the ADHD trend line is shown as a dashed line, and the 22q11DS trend line is shown as a solid line.
Table 4.
Brain regions showing significant relationships between successful stopping activity and impulsivity symptoms for 22q11DS and ADHD patients. Coordinates for location of maximum activation of significant clusters in MNI space.
| Trial Type | Contrast | Region | Voxel # | Max Z-Score | Max X (mm) | Max Y (mm) | Max Z (mm) |
|---|---|---|---|---|---|---|---|
| Successful Stopping | |||||||
| Barratt Impulsivity Scale: Cognitive Impulsivity | |||||||
| ADHD > 22q11DS | |||||||
| Precuneus | 645 | 3.45 | 16 | -42 | 12 | ||
| Medial frontal gyrus | 442 | 3.62 | 4 | 40 | -6 | ||
| 22q11DS negative correlation | |||||||
| Medial frontal gyrus | 1244 | 3.8 | 2 | 36 | -10 | ||
| Precuneus, posterior cingulate cortex | 533 | 4.22 | -38 | -70 | 44 | ||
| Left inferior parietal cortex | 334 | 3.68 | -2 | -46 | 26 | ||
| Barratt Impulsivity Scale: Behavioral Impulsivity | |||||||
| 22q11DS > ADHD | |||||||
| Left motor cortex | 438 | 3.78 | -60 | -14 | 12 | ||
| 22q11DS positive correlation | |||||||
| Left motor cortex | 815 | 4.23 | -50 | -24 | -4 | ||
As illustrated in Fig. 4b and detailed in Table 4, a significant interaction in the opposite direction was observed between patient group and Behavioral Impulsivity, 22q11DS patients showed a positive correlation between activity in the left motor cortex and Behavioral Impulsivity, while ADHD patients showed an inverse correlation. Follow-up regressions within 22q11DS patients alone confirmed a positive association between successful stopping-related activation in the left motor cortex and Behavioral Impulsivity, such that increased neural activity in this region during successful stopping was associated with more severe self-reported Behavioral Impulsivity. No brain regions were inversely correlated with Behavioral Impulsivity in the 22q11DS patients, and no significant correlation with Behavioral Impulsivity was evident in the ADHD group alone after correction for multiple comparisons. In a follow-up analysis, we excluded one outlier that was evident within the 22q11DS sample for the Behavioral Impulsivity score. While the interaction was no longer significant, the 22q11DS within-group correlation between RI-related activity and Behavioral Impulsivity remained significant.
4. Discussion
To our knowledge, this is the first study to directly compare the neural substrates of response inhibition (RI) across distinct clinical populations associated with attentional dysfunction (22q11DS and idiopathic ADHD) and healthy controls. We hypothesized that, while 22q11DS and idiopathic ADHD patients would share similar dysfunction within RI-related neural systems, this shared dysfunction would show different relationships to behavior. Specifically, we anticipated that 22q11DS patients would show an inverse relationship with Cognitive Impulsivity (Montojo et al., 2015), whereas we would not observe a significant relationship between neural activity and cognitive or Behavioral Impulsivity within the ADHD cohort.
Supporting our hypotheses, our fMRI conjunction findings indicate that, during performance of the Stop-signal task, 22q11DS and ADHD patients show reduced RI-related neural activity compared to healthy controls within multiple overlapping brain regions, specifically the ACC, middle and superior frontal gyri, middle temporal gyrus, inferior parietal cortex, paracingulate gyrus, bilateral thalamus and cerebellum. In addition to this shared pattern of hypoactivation across both clinical groups, 22q11DS patients showed reduced RI-related neural activity relative to ADHD patients within the bilateral middle frontal gyrus. Also in agreement with our predictions, in 22q11DS patients (but not ADHD patients), reduced neural activity within the medial frontal gyrus, precuneus/posterior cingulate, and inferior parietal cortex was associated with greater self-reported Cognitive Impulsivity. This measure of Cognitive Impulsivity was elevated in 22q11DS patients, in agreement with prior findings regarding the nature of ADHD symptomatology in this population (Antshel et al., 2008; Niklasson et al., 2001). These findings suggest that 22q11DS and idiopathic ADHD patient groups share a pattern of reduced neural engagement during RI, relative to healthy controls, but that 22q11DS and idiopathic ADHD patient groups differ with regard to the relationship of RI-related neural activity to trait impulsivity. Specifically, these findings suggest that neural dysfunction during RI in 22q11DS is associated with variability in Cognitive Impulsivity, and not Behavioral Impulsivity. This relationship was not observed in patients with ADHD.
4.1. Shared deficits in response inhibition-related neural activity for 22q11DS and idiopathic ADHD
In the current study, both 22q11DS and idiopathic ADHD patients showed significantly less engagement of several RI-related regions that have been implicated in prior research (Aron et al., 2007; Aron and Poldrack, 2006; Whelan et al., 2012). The use of a tracking Stop-signal task ensured that participants successfully inhibited responses 50% of the time, which results in an equal number of successful stop trials for comparison and controls for difficulty level across participants. As such, findings of reduced activation in these cortical regions and the thalamus during successful inhibition in both patient groups are consistent with models of RI suggesting that the right inferior frontal cortex, via connections with a prefrontal-basal ganglia network, implements inhibition of a prepotent motor response (Aron et al., 2014). Furthermore, findings of reduced RI-related neural activation in these patient groups, both of which are characterized by deficits in inhibitory control, are consistent with prior research (Mulligan et al., 2011; Sebastian et al., 2012; van Rooij et al., 2015).
With regard to behavioral performance, it is somewhat surprising that there were no differences in SSRT between groups, given that deficits in response inhibition are widely reported in ADHD. Along these lines, we observed non-significantly increased accuracy on Go trials in our ADHD sample as compared with controls. Because this study included adult participants with ADHD who were able to complete up to 2 days of testing and from whom we have complete, usable data, it is possible that these individuals are not characterized by pronounced deficits in behavioral indices of response inhibition, or that they have acquired compensatory mechanisms to overcome response inhibition deficits. From that standpoint, the findings of anomalous patterns of neural activity during RI in the ADHD group – despite intact behavioral performance – are particularly notable.
We previously found that healthy controls showed significantly greater activation than 22q11DS patients within the frontal cortical and basal ganglia regions during successful RI (Montojo et al., 2015). There were no regions where 22q11DS patients exhibited greater activity than controls. One prior study on adolescent 22q11DS patients showed greater activation in the left parietal regions compared to healthy controls during performance of a Go/No-Go task (Gothelf et al., 2007). However, in addition to a difference in tasks used, the different findings may be accounted for by age differences, as differences in neural activity during this age range may reflect changes in neural trajectories during this developmentally sensitive period.
In terms of RI-related activation in idiopathic ADHD patients, although reports are mixed, the bulk of the evidence supports a pattern of reduced neural activation during RI as compared with controls (Cortese et al., 2012; Mulligan et al., 2011; Sebastian et al., 2012; van Rooij et al., 2015). Strong support comes from a recent large-scale study reporting a pattern of relative hypoactivity in 185 adolescents with ADHD, as well as in their unaffected siblings, as compared with healthy controls. Employing the Stopsignal task, the authors found reduced activation during successful RI seen in the left inferior and superior frontal gyri in ADHD patients as compared with unaffected siblings and controls, and reduced RI-related activation seen in the left supramarginal gyrus, right postcentral gyrus, and right temporal–parietal junction in both ADHD patients and their unaffected siblings as compared with controls (van Rooij et al., 2015). These findings suggest that RI-related deficits in activity within these regions may be useful endophenotypes that extend beyond the affected individuals of the family. While there are also reports of greater fronto-striatal activation (Dillo et al., 2010; Karch et al., 2010), or no differences in activation (Carmona et al., 2012; Congdon et al., 2014), during RI in ADHD patients as compared with controls, a review of this heterogeneous literature suggests that differences in the cognitive paradigm employed (e.g., Stop-signal task vs. Go/No-Go task), medication status, symptom severity, and small sample size may account for these discrepant findings. Our findings of common deficits in neural activity during RI between idiopathic ADHD and patients with a specific genetic mutation that is highly penetrant for the illness suggest that this pattern of hypoactivation during RI may be a valuable neural endophenotype underlying attentional dysfunction.
The direct comparison in RI-related neural activity between 22q11DS and ADHD patients was of particular interest to determine whether the clinical groups differed in RI-related neural circuitry. This analysis revealed that ADHD patients showed greater activity in the bilateral middle frontal gyrus regions compared to 22q11DS patients. Although the right inferior frontal cortex has been shown to be central to RI, it does so through a network of prefrontal–basal ganglia regions (Aron et al., 2014) and the middle frontal gyrus is often engaged during suppression of a prepotent response (e.g., Dambacher et al., 2014). Neuroimaging studies have additionally implicated the middle frontal gyrus in the processing of higher order information, including functions of working memory (Leung et al., 2002; McCarthy et al., 1994) and episodic retrieval (Rajah et al., 2011). Our findings indicate substantial overlap across clinical groups in terms of hypoactivation in RI-related neural circuitry relative to controls, but with additional dysfunction in the middle frontal gyrus in 22q11DS patients.
4.2. Trait impulsivity and neural activity during response inhibition
Impulsivity is a key symptom of ADHD (Robbins et al., 2012) and has been defined as a trait that leads to “actions which are poorly conceived, prematurely expressed, unduly risky or inappropriate to the situation and that often result in undesirable consequences” (Daruna and Barnes, 1993). The complexity of defining impulsivity has led researchers to suggest that impulsivity may not be a unitary construct (Dalley et al., 2011). In line with this, a recent study proposed that a two-factor model including Cognitive and Behavioral Impulsivity may best describe the multidimensional structural representation of self-reported trait impulsivity as measured by the BIS-11 (Reise et al., 2013).
In terms of trait impulsivity, both 22q11DS and ADHD patients showed greater Cognitive Impulsivity compared to healthy controls, as measured by the BIS-11. The elevated Cognitive Impulsivity scores that we observed within the 22q11DS group are of interest given that 1) the Cognitive Impulsivity subscale specifically captures difficulties in attentional control (Reise et al., 2013), and 2) ADHD diagnoses in 22q11DS are primarily of the inattentive subtype (Antshel et al., 2008; Niklasson et al., 2001). In line with this, we observed that while ADHD patients showed elevated Behavioral Impulsivity relative to both 22q11DS patients and controls, 22q11DS patients did not show increased Behavioral Impulsivity relative to controls. Thus, these findings are in general agreement with previous literature and suggest that 22q11DS and ADHD patients share elevated Cognitive Impulsivity, while adult ADHD patients exhibit elevations in both Cognitive and Behavioral Impulsivity.
Our findings further indicate that deficits in RI-related neural activation are associated with increased Cognitive Impulsivity in 22q11DS patients, but not Behavioral Impulsivity. Specifically, as Cognitive Impulsivity increases, RI-related neural activity decreases within the medial frontal cortex, precuneus/posterior cingulate, and inferior parietal cortex. We have previously reported an inverse correlation between RI-related activation and Cognitive Impulsivity in 22q11DS patients (Montojo et al., 2015) within a set of a priori RI-related regions of interest (primarily frontal and basal ganglia regions). Here, we extend those findings in a whole-brain regression analysis, to demonstrate an association between RI-related neural activity and Cognitive Impulsivity in a broader set of regions. Together, these complementary analysis approaches provide convergent evidence that self-reported Cognitive Impulsivity is inversely associated with RI-related neural activation in multiple brain regions. In contrast, the finding of a positive association between Behavioral Impulsivity and RI-related neural activity in the left motor cortex in 22q11DS patients is novel. Although this association may reflect a compensatory mechanism in patients required to achieve sufficient RI, the presence of an outlier driving the interaction effect leads us to exercise caution in interpreting these findings and suggests that replication is necessary.
While pathophysiologic models of ADHD have primarily focused on dysfunction within fronto-striatal neural circuitry (Arnsten, 2009; Cortese et al., 2012; Krain and Castellanos, 2006), recent studies have suggested that cingulate–precuneus interactions may constitute a new locus of dysfunction in ADHD (Castellanos and Proal, 2012; Castellanos et al., 2008). Specifically, a prior study reported reduced functional connectivity between the anterior cingulate cortex and precuneus/posterior cingulate cortex regions during resting state within ADHD patients (Castellanos et al., 2008). We extend those findings here to patients with attentional dysfunction associated with homogeneous genetic etiology (22q11.2 deletions), by reporting an inverse correlation between RI-related neural activity and Cognitive Impulsivity symptoms within the precuneus/posterior cingulate. In addition, 22q11DS patients show decreased cortical volume and surface area within the precuneus (Jalbrzikowski et al., 2013). These findings are in agreement with the idea of this region as another locus of dysfunction, with variability in neural activity being related to the severity of trait impulsivity.
With regard to the medial prefrontal cortex, both primate and human studies implicate this region in cognitive control functions (Ridderinkhof et al., 2004), including: the detection of unfavorable outcomes (Ito et al., 2003; Shidara and Richmond, 2002; Stuphorn et al., 2000), response errors (Gemba et al., 1986; Ito et al., 2003), response conflict (Botvinick et al., 2001), and decision uncertainty (Botvinick et al., 2001). The inferior parietal cortex is activated during Go/No-Go tasks (Braveret al., 2001; Garavan et al., 1999; Kiehl et al., 2000) and has been related to response conflict (Braver et al., 2001; Carter et al., 2000) and, more generally, multiple aspects of sensory processing and sensorimotor integration (Caspers et al., 2006; Clower et al., 2001). We suggest that reduced engagement of neural circuitry within these regions has potential significance for the clinical presentation of 22q11DS.
Despite clear elevations in trait impulsivity in idiopathic ADHD, the relationship between impulsivity and RI-related neural activation is still an active area of investigation. In a large-scale study of ADHD patients, van Rooij et al. (2015) reported an inverse correlation between ADHD symptom count (based on hyperactive/impulsive and inattentive symptoms according to DSM-IV criteria) and neural activation in the inferior frontal gyrus during successful RI. Here, our within-group analyses did not reveal an association between RI-related neural activity and Cognitive or Behavioral Impulsivity for ADHD patients that survived correction. This may be related to the relatively small sample of ADHD patients included in our study. Given the genetic and phenotypic heterogeneity within ADHD patients, it may be necessary to include a larger sample to detect neural function–behavior relationships. These findings also suggest that there may be more power to detect associations between dysfunctional RI-related neural activity and trait impulsivity within 22q11DS patients, given the more homogeneous genetic etiology.
4.3. Implications for genetic studies
Notably, the 22q11.2 locus includes the catechol-O-methyltransferase (COMT) gene, which codes for an enzyme involved in prefrontal dopamine metabolism (Egan et al., 2001; Fallgatter and Lesch, 2007; Gothelf et al., 2007; Lachman et al., 1996) which may be relevant to behavioral manifestations of inattention and impulsivity in 22q11DS (Cole et al., 2013; Cools et al., 2007; Gothelf et al., 2007; Shashi et al., 2006; Soeiro-De-Souza et al., 2013).
Investigation of the relationships between impulsive behavior, response inhibition-related neural circuitry, and allelic variation and expression of COMT and other genes involved in dopaminergic function is warranted in future, larger-scale studies of this syndrome.
5. Conclusions
The findings reported here help to elucidate the neurobiological basis of attentional dysfunction in the following ways. First, we directly compared patients with ADHD, a group that traditionally exhibits substantial heterogeneity at both genotypic and phenotypic levels, with 22q11DS patients, a group representing a relatively homogeneous genetic etiology with common downstream phenotypic characteristics. 22q11DS offers a compelling model for the investigation of the neurobiological substrates of attentional dysfunction in ADHD. Direct comparison of our ADHD and 22q11DS patients allowed us to demonstrate a shared deficit in RI-related neural activation between groups, as compared with controls, but circumscribed differences in ADHD patients as compared with 22q11DS patients. Second, we investigated the relationship between RI-related neural activity and different aspects of trait impulsivity — Cognitive and Behavioral. We used a dimensional approach to assess these aspects of impulsivity symptoms, which is in agreement with the RDoC initiative (Cuthbert and Kozak, 2013; Insel et al., 2010) that aims to characterize psychopathology on dimensions of observable behaviors or neurobiological measures rather than traditional diagnostic categories. As such, our results provide initial evidence for how dimensional assessments of impulsivity may characterize distinct etiologies of downstream ADHD diagnosis. Finally, our ability to detect significant within-group correlations between impulsivity measures and RI-related neural activation in 22q11DS patients, but not ADHD patients, has implications for future efforts, as there may be more power to detect associations between dysfunctional RI-related neural activity and trait impulsivity within 22q11DS patients, given the more homogeneous genetic etiology.
Taken together, these findings suggest a pattern of reduced neural engagement during RI in both idiopathic ADHD and 22q11DS, seen in the ACC, cerebellum, inferior parietal cortex, middle frontal gyrus, middle temporal gyrus, paracingulate gyrus, superior frontal gyrus, and bilateral thalamus. Notably, 22q11DS patients differed relative to idiopathic ADHD patients in that they showed hypoactivation of the bilateral frontal regions. Our results further indicate that deficits in RI-related activation are associated with increased Cognitive Impulsivity in 22q11DS patients, but not increased Behavioral Impulsivity, consistent with the behavioral presentation of the disorder. These results offer initial evidence that similar underlying RI-related deficits in ADHD and 22q11DS may manifest as different forms of real-world trait impulsivity between patient groups.
Disclosures
None of the authors have relevant financial disclosures or conflicts of interest.
Funding sources
This manuscript was supported by grants from the National Institute of Mental Health: RO1 MH085953 (CEB), NIH/NICHD grant P50-HD-055784 (Pilot Project Grant to CEB), NIH/NIMH5T32MH073526-05 and NIH/NIMH T32MH096682 (Postdoctoral Training Program Fellowships given to CAM), NIH/NIMH5T32MH073526-05 (Postdoctoral Neurogenetics Training Program Fellowship given to MJ), and UL1-DE019580 and PL1MH083271 (NIH Roadmap grants to RMB). These funding sources had no further role in study design, in the collection, analysis, and interpretation of data, in the writing of the report, and in the decision to submit the paper for publication.
The following are the supplementary data related to this article.
Distributions of trait impulsivity scores for 22q11DS and ADHD patients. The y-axis represents the percent of subjects who scored at each value of cognitive or behavioral impulsivity, as measured by the BIS-11. Gray bars represent 22q11DS patients and black bars represent ADHD patients. The x-axis represents: (a) cognitive impulsivity scores and (b) behavioral impulsivity scores.
Differences in activation during response initiation and inhibition failure for 22q11DS and ADHD patients. Turquoise maps represent activity from the contrast of ADHD > 22q11DS. (a) Activation maps represent the contrast of Go–Null, to investigate activity related to response initiation. (b) Activation maps represent the contrast of Stop Unsuccessful–Stop Successful, to investigate activity related to inhibition failure.
Regions showing between group differences for ADHD patients, 22q11DS patients, and healthy controls during response initiation (Go vs. Null) and inhibitory failure (Stop Unsuccessful vs. Stop Successful) contrasts. Montreal Neurological Institute (MNI) coordinates represent the location of maximum activation for significant clusters.
Supplementary material.
Acknowledgments
We would like to thank the participants and their families for being a part of our research. We would also like to thank Ms. Chelsea Gilbert, Dr. Sarah Marvin, and Dr. Laurie Brenner, who assisted in conducting clinical assessments and administering neuropsychological measures to our participants, and Wen-Ching Tran, who assisted in data management and scoring. We would like to thank Sabrina Domicoli for quality control pre-processing of neuroimaging data. We would like to thank Angelica Bato and Eric Miller for their assistance with data collection from individuals recruited through the Center for Neuropsychiatric Phenomics.
Footnotes
This work was performed at the Semel Institute for Neuroscience and Human Behavior (Department of Psychiatry & Biobehavioral Sciences, UCLA) and was presented in part at the Human Brain Mapping conference, June 14th–18th, 2015, Honolulu, HI.
References
- American Psychiatric Association . fifth edition. APA; Washington, DC, USA: 2013. Diagnostic and Statistical Manual of Mental Disorders. [Google Scholar]
- Antshel K.M., Fremont W., Kates W.R. The neurocognitive phenotype in velo-sardio-facial syndrome: a developmental perspective. Dev. Disabil. Res. Rev. 2008;14(1):43–51. doi: 10.1002/ddrr.7. 18636636 [DOI] [PubMed] [Google Scholar]
- Arnsten A.F.T. Toward a new understanding of attention-deficit hyperactivity disorder pathophysiology: an important role for prefrontal cortex dysfunction. C.N.S. Drugs. 2009;23:33–41. doi: 10.2165/00023210-200923000-00005. 19621976 [DOI] [PubMed] [Google Scholar]
- Aron A.R., Durston S., Eagle D.M., Logan G.D., Stinear C.M., Stuphorn V. Converging evidence for a fronto-basal-ganglia network for inhibitory control of action and cognition. J. Neurosci. 2007;27(44):11860–11864. doi: 10.1523/JNEUROSCI.3644-07.2007. 17978025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aron A.R., Poldrack R.A. Cortical and subcortical contributions to stop signal response inhibition: role of the subthalamic nucleus. J. Neurosci. 2006;26(9):2424–2433. doi: 10.1523/JNEUROSCI.4682-05.2006. 16510720 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aron A.R., Robbins T.W., Poldrack R.A. Inhibition and the right inferior frontal cortex: one decade on. Trends Cogn. Sci. (Regul. Ed.) 2014;18(4):177–185. doi: 10.1016/j.tics.2013.12.003. 24440116 [DOI] [PubMed] [Google Scholar]
- Band G.P.H., van der Molen M.W., Logan G.D. Horse-race model simulations of the stop-signal procedure. Acta Psychol. 2003;112(2):105–142. doi: 10.1016/s0001-6918(02)00079-3. 12521663 [DOI] [PubMed] [Google Scholar]
- Botvinick M.M., Braver T.S., Barch D.M., Carter C.S., Cohen J.D. Conflict monitoring and cognitive control. Psychol. Rev. 2001;108(3):624–652. doi: 10.1037/0033-295x.108.3.624. 11488380 [DOI] [PubMed] [Google Scholar]
- Brainard D.H. The psychophysics toolbox. Spat. Vis. 1997;10(4):433–436. 9176952 [PubMed] [Google Scholar]
- Braver T.S., Barch D.M., Gray J.R., Molfese D.L., Snyder A. Anterior cingulate cortex and response conflict: effects of frequency, inhibition and errors. Cerebral Cortex (New York, N.Y.: 1991) 2001;11(9):825–836. doi: 10.1093/cercor/11.9.825. 11532888 [DOI] [PubMed] [Google Scholar]
- Carmona S., Hoekzema E., Ramos-Quiroga J.A., Richarte V., Canals C., Bosch R. Response inhibition and reward anticipation in medication-naïve adults with attention-deficit/hyperactivity disorder: a within-subject case–control neuroimaging study. Hum. Brain Mapp. 2012;33(10):2350–2361. doi: 10.1002/hbm.21368. 21826761 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carter C.S., Macdonald A.M., Botvinick M., Ross L.L., Stenger V.A., Noll D., Cohen J.D. Parsing executive processes: strategic vs. evaluative functions of the anterior cingulate cortex. Proc. Natl. Acad. Sci. U. S. A. 2000;97(4):1944–1948. doi: 10.1073/pnas.97.4.1944. 10677559 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caspers S., Geyer S., Schleicher A., Mohlberg H., Amunts K., Zilles K. The human inferior parietal cortex: cytoarchitectonic parcellation and interindividual variability. Neuroimage. 2006;33(2):430–448. doi: 10.1016/j.neuroimage.2006.06.054. 16949304 [DOI] [PubMed] [Google Scholar]
- Castellanos F.X., Margulies D.S., Kelly C., Uddin L.Q., Ghaffari M., Kirsch A. Cingulate–precuneus interactions: a new locus of dysfunction in adult attention-deficit/hyperactivity disorder. Biol. Psychiatry. 2008;63(3):332–337. doi: 10.1016/j.biopsych.2007.06.025. 17888409 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castellanos F.X., Proal E. Large-scale brain systems in ADHD: beyond the prefrontal–striatal model. Trends Cogn. Sci. (Regul. Ed.) 2012;16(1):17–26. doi: 10.1016/j.tics.2011.11.007. 22169776 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chambers C.D., Garavan H., Bellgrove M.A. Insights into the neural basis of response inhibition from cognitive and clinical neuroscience. Neurosci. Biobehav. Rev. 2009;33(5):631–646. doi: 10.1016/j.neubiorev.2008.08.016. 18835296 [DOI] [PubMed] [Google Scholar]
- Clower D.M., West R.A., Lynch J.C., Strick P.L. The inferior parietal lobule is the target of output from the superior colliculus, hippocampus, and cerebellum. J. Neurosci. 2001;21(16):6283–6291. doi: 10.1523/JNEUROSCI.21-16-06283.2001. 11487651 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cole D.M., Oei N.Y.L., Soeter R.P., Both S., van Gerven J.M.A., Rombouts S.A.R.B., Beckmann C.F. Dopamine-dependent architecture of cortico-subcortical network connectivity. Cerebral Cortex (New York, N.Y.: 1991) 2013;23(7):1509–1516. doi: 10.1093/cercor/bhs136. 22645252 [DOI] [PubMed] [Google Scholar]
- Congdon E., Altshuler L.L., Mumford J.A., Karlsgodt K.H., Sabb F.W., Ventura J. Neural activation during response inhibition in adult attention-deficit/hyperactivity disorder: preliminary findings on the effects of medication and symptom severity. Psychiatry Res. 2014;222(1–2):17–28. doi: 10.1016/j.pscychresns.2014.02.002. 24581734 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Congdon E., Mumford J.A., Cohen J.R., Galvan A., Aron A.R., Xue G. Engagement of large-scale networks is related to individual differences in inhibitory control. Neuroimage. 2010;53(2):653–663. doi: 10.1016/j.neuroimage.2010.06.062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Congdon E., Mumford J.A., Cohen J.R., Galvan A., Canli T., Poldrack R.A. Measurement and reliability of response inhibition. Front. Psychol. 2012;3:37. doi: 10.3389/fpsyg.2012.00037. 22363308 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cools R., Sheridan M., Jacobs E., D'Esposito M. Impulsive personality predicts dopamine-dependent changes in frontostriatal activity during component processes of working memory. J. Neurosci. 2007;27(20):5506–5514. doi: 10.1523/JNEUROSCI.0601-07.2007. 17507572 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cortese S., Kelly C., Chabernaud C., Proal E., Di Martino A., Milham M.P., Castellanos F.X. Toward systems neuroscience of ADHD: a meta-analysis of 55 fMRI studies. Am. J. Psychiatry. 2012;169(10):1038–1055. doi: 10.1176/appi.ajp.2012.11101521. 22983386 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crunelle C.L., Veltman D.J., van Emmerik-van Oortmerssen K., Booij J., van den Brink W. Impulsivity in adult ADHD patients with and without cocaine dependence. Drug Alcohol Depend. 2013;129(1–2):18–24. doi: 10.1016/j.drugalcdep.2012.09.006. 23026814 [DOI] [PubMed] [Google Scholar]
- Cuthbert B.N., Kozak M.J. Constructing constructs for psychopathology: the NIMH research domain criteria. J. Abnorm. Psychol. 2013;122(3) doi: 10.1037/a0034028. 24016027 [DOI] [PubMed] [Google Scholar]
- Dalley J.W., Everitt B.J., Robbins T.W. Impulsivity, compulsivity, and top-down cognitive control. Neuron. 2011;69(4):680–694. doi: 10.1016/j.neuron.2011.01.020. 21338879 [DOI] [PubMed] [Google Scholar]
- Dambacher F., Sack A.T., Lobbestael J., Arntz A., Brugman S., Schuhmann T. A network approach to response inhibition: dissociating functional connectivity of neural components involved in action restraint and action cancellation. Eur. J. Neurosci. 2014;39(5):821–831. doi: 10.1111/ejn.12425. 24289860 [DOI] [PubMed] [Google Scholar]
- Daruna J., Barnes P. A neurodevelopmental view of impulsivity. In: McCown W., Johnson J., Shure M., editors. The Impulsive Client: Theory, Research, and Treatment. American Psychological Association; Washington, DC: 1993. pp. 23–27. [Google Scholar]
- Dickstein S.G., Bannon K., Castellanos F.X., Milham M.P. The neural correlates of attention deficit hyperactivity disorder: an ALE meta-analysis. J. Child Psychol. Psychiatry Allied Discip. 2006;47(10):1051–1062. doi: 10.1111/j.1469-7610.2006.01671.x. 17073984 [DOI] [PubMed] [Google Scholar]
- Dillo W., Göke A., Prox-Vagedes V., Szycik G.R., Roy M., Donnerstag F. Neuronal correlates of ADHD in adults with evidence for compensation strategies — a functional MRI study with a Go/No-go paradigm. Ger Med Sci. 2010;8:Doc09. doi: 10.3205/000098. 20421953 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durston S. A review of the biological bases of ADHD: what have we learned from imaging studies? Ment. Retard. Dev. Disabil. Res. Rev. 2003;9(3):184–195. doi: 10.1002/mrdd.10079. 12953298 [DOI] [PubMed] [Google Scholar]
- Egan M.F., Goldberg T.E., Kolachana B.S., Callicott J.H., Mazzanti C.M., Straub R.E. Effect of COMT Val108/158 Met genotype on frontal lobe function and risk for schizophrenia. Proc. Natl. Acad. Sci. U. S. A. 2001;98(12):6917–6922. doi: 10.1073/pnas.111134598. 11381111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elia J., Devoto M. ADHD genetics: 2007 update. Curr. Psychiatry Rep. 2007;9(5):434–439. doi: 10.1007/s11920-007-0057-z. 17915085 [DOI] [PubMed] [Google Scholar]
- Epstein J.N., Casey B.J., Tonev S.T., Davidson M.C., Reiss A.L., Garrett A. ADHD- and medication-related brain activation effects in concordantly affected parent–child dyads with ADHD. J. Child Psychol. Psychiatry Allied Discip. 2007;48(9):899–913. doi: 10.1111/j.1469-7610.2007.01761.x. 17714375 [DOI] [PubMed] [Google Scholar]
- Fallgatter A.J., Lesch K.P. 22q11.2 deletion syndrome as a natural model for COMT haploinsufficiency-related dopaminergic dysfunction in ADHD. Int. J. Neuropsychopharmacol. 2007;10(3):295–299. doi: 10.1017/S1461145706006985. 16848928 [DOI] [PubMed] [Google Scholar]
- First M.B., Spitzer R.L., Gibbon M., Williams J.B. New York Biometrics Research, New York State Psychiatric Institute; 2004. Structured Clinical Interview for DSM-IV-TR Axis I Disorders, Research Version (Patient Edition SCID-IP) [Google Scholar]
- Franke B., Neale B.M., Faraone S.V. Genome-wide association studies in ADHD. Hum. Genet. 2009;126(1):13–50. doi: 10.1007/s00439-009-0663-4. 19384554 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garavan H., Ross T.J., Stein E.A. Right hemispheric dominance of inhibitory control: an event-related functional MRI study. Proc. Natl. Acad. Sci. U. S. A. 1999;96(14):8301–8306. doi: 10.1073/pnas.96.14.8301. 10393989 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gemba H., Sasaki K., Brooks V.B. “Error” potentials in limbic cortex (anterior cingulate area 24) of monkeys during motor learning. Neurosci. Lett. 1986;70(2):223–227. doi: 10.1016/0304-3940(86)90467-2. 3774229 [DOI] [PubMed] [Google Scholar]
- Gothelf D., Hoeft F., Hinard C., Hallmayer J.F., Stoecker J.V.D., Antonarakis S.E. Abnormal cortical activation during response inhibition in 22q11.2 deletion syndrome. Hum. Brain Mapp. 2007;28(6):533–542. doi: 10.1002/hbm.20405. 17427209 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green T., Gothelf D., Glaser B., Debbané M., Frisch A., Kotler M. Psychiatric disorders and intellectual functioning throughout development in velocardiofacial (22q11.2 deletion) syndrome. J. Am. Acad. Child Adolesc. Psychiatry. 2009;48(11):1060–1068. doi: 10.1097/CHI.0b013e3181b76683. 19797984 [DOI] [PubMed] [Google Scholar]
- Insel T., Cuthbert B., Garvey M., heinssen R., Pine D.S., Quinn K. Research domain criteria (RDoC): toward a new classification framework for research on mental disorders. Am. J. Psychiatry. 2010;167(7):748–751. doi: 10.1176/appi.ajp.2010.09091379. 20595427 [DOI] [PubMed] [Google Scholar]
- Ito S., Stuphorn V., Brown J.W., Schall J.D. Performance monitoring by the anterior cingulate cortex during saccade countermanding. Science. 2003;302(5642):120–122. doi: 10.1126/science.1087847. 14526085 [DOI] [PubMed] [Google Scholar]
- Jalbrzikowski M., Jonas R., Senturk D., Patel A., Chow C., Green M.F., Bearden C.E. Structural abnormalities in cortical volume, thickness, and surface area in 22q11.2 microdeletion syndrome: relationship with psychotic symptoms. Neuroimage Clin. 2013;3:405–415. doi: 10.1016/j.nicl.2013.09.013. 24273724 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karch S., Thalmeier T., Lutz J., Cerovecki A., Opgen-Rhein M., Hock B. Neural correlates (ERP/fMRI) of voluntary selection in adult ADHD patients. Eur. Arch. Psychiatry Clin. Neurosci. 2010;260(5):427–440. doi: 10.1007/s00406-009-0089-y. 19907927 [DOI] [PubMed] [Google Scholar]
- Kaufman J., Birmaher B., Brent D., Rao U., Flynn C., Moreci P. Schedule for affective disorders and schizophrenia for school-age children-present and lifetime version (K-SADS-PL): initial reliability and validity data. J. Am. Acad. Child Adolesc. Psychiatry. 1997;36(7):980–988. doi: 10.1097/00004583-199707000-00021. 9204677 [DOI] [PubMed] [Google Scholar]
- Kiehl K.A., Smith A.M., Hare R.D., Liddle P.F. An event-related potential investigation of response inhibition in schizophrenia and psychopathy. Biol. Psychiatry. 2000;48(3):210–221. doi: 10.1016/s0006-3223(00)00834-9. 10924664 [DOI] [PubMed] [Google Scholar]
- Krain A.L., Castellanos F.X. Brain development and ADHD. Clin. Psychol. Rev. 2006;26(4):433–444. doi: 10.1016/j.cpr.2006.01.005. 16480802 [DOI] [PubMed] [Google Scholar]
- Lachman H.M., Morrow B., Shprintzen R., Veit S., Parsia S.S., Faedda G. Association of codon 108/158 catechol-O-methyltransferase gene polymorphism with the psychiatric manifestations of velo-cardio-facial syndrome. Am. J. Med. Genet. 1996;67(5):468–472. doi: 10.1002/(SICI)1096-8628(19960920)67:5<468::AID-AJMG5>3.0.CO;2-G. 8886163 [DOI] [PubMed] [Google Scholar]
- Leung H.-C., Gore J.C., Goldman-Rakic P.S. Sustained mnemonic response in the human middle frontal gyrus during on-line storage of spatial memoranda. J. Cogn. Neurosci. 2002;14(4):659–671. doi: 10.1162/08989290260045882. 12126506 [DOI] [PubMed] [Google Scholar]
- Logan G.D. D., Dagenbach T.H., Carr, editors. xiv. Academic Press; Zur., Switzerland: 1994. (Inhibitory Processes in Attention, Memory, and Language). [Google Scholar]
- Malloy-Diniz L., Fuentes D., Leite W.B., Correa H., Bechara A. Impulsive behavior in adults with attention deficit/ hyperactivity disorder: characterization of attentional, motor and cognitive impulsiveness. J. Int. Neuropsychol. Soc. 2007;13(4):693–698. doi: 10.1017/S1355617707070889. 17521490 [DOI] [PubMed] [Google Scholar]
- McCarthy G., Blamire A.M., Puce A., Nobre A.C., Bloch G., Hyder F. Functional magnetic resonance imaging of human prefrontal cortex activation during a spatial working memory task. Proc. Natl. Acad. Sci. U. S. A. 1994;91(18):8690–8694. doi: 10.1073/pnas.91.18.8690. 8078943 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Montojo C.A., Jalbrzikowski M., Congdon E., Domicoli S., Chow C., Dawson C. Neural substrates of inhibitory control deficits in 22q11.2 deletion syndrome. Cerebral Cortex (New York, N.Y.: 1991) 2015;25(4):1069–1079. doi: 10.1093/cercor/bht304. 24177988 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mulligan R.C., Knopik V.S., Sweet L.H., Fischer M., Seidenberg M., Rao S.M. Neural correlates of inhibitory control in adult attention deficit/hyperactivity disorder: evidence from the Milwaukee longitudinal sample. Psychiatry Res. 2011;194(2):119–129. doi: 10.1016/j.pscychresns.2011.02.003. 21937201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mumford J.A., Poldrack R.A. Modeling group fMRI data. Soc. Cogn. Affect. Neurosci. 2007;2(3):251–257. doi: 10.1093/scan/nsm019. 18985145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niklasson L., Rasmussen P., Oskarsdóttir S., Gillberg C. Neuropsychiatric disorders in the 22q11 deletion syndrome. Genet. Med. 2001;3(1):79–84. doi: 10.1097/00125817-200101000-00017. 11339385 10. [DOI] [PubMed] [Google Scholar]
- Patton J.H., Stanford M.S., Barratt E.S. Factor structure of the Barratt impulsiveness scale. J. Clin. Psychol. 1995;51(6):768–774. doi: 10.1002/1097-4679(199511)51:6<768::aid-jclp2270510607>3.0.co;2-1. 8778124 [DOI] [PubMed] [Google Scholar]
- Price C.J., Friston K.J. Cognitive conjunction: a new approach to brain activation experiments. Neuroimage. 1997;5(4 Pt 1):261–270. doi: 10.1006/nimg.1997.0269. 9345555 [DOI] [PubMed] [Google Scholar]
- Rajah M.N., Languay R., Grady C.L. Age-related changes in right middle frontal gyrus volume correlate with altered episodic retrieval activity. Journal of Neuroscience. 2011;31(49):17941–17954. doi: 10.1523/JNEUROSCI.1690-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reise S.P., Moore T.M., Sabb F.W., Brown A.K., London E.D. The Barratt Impulsiveness Scale-11: reassessment of its structure in a community sample. Psychol. Assess. 2013;25(2):631–642. doi: 10.1037/a0032161. 23544402 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ridderinkhof K.R., Ullsperger M., Crone E.A., Nieuwenhuis S. The role of the medial frontal cortex in cognitive control. Science. 2004;306(5695):443–447. doi: 10.1126/science.1100301. 15486290 [DOI] [PubMed] [Google Scholar]
- Robbins T.W., Gillan C.M., Smith D.G., de Wit S., Ersche K.D. Neurocognitive endophenotypes of impulsivity and compulsivity: towards dimensional psychiatry. Trends Cogn. Sci. (Regul. Ed.) 2012;16(1):81–91. doi: 10.1016/j.tics.2011.11.009. 22155014 [DOI] [PubMed] [Google Scholar]
- Schulz K.P., Fan J., Tang C.Y., Newcorn J.H., Buchsbaum M.S., Cheung A.M., Halperin J.M. Response inhibition in adolescents diagnosed with attention deficit hyperactivity disorder during childhood: an event-related FMRI study. Am. J. Psychiatry. 2004;161(9):1650–1657. doi: 10.1176/appi.ajp.161.9.1650. 15337656 [DOI] [PubMed] [Google Scholar]
- Sebastian A., Gerdes B., Feige B., Klöppel S., Lange T., Philipsen A. Neural correlates of interference inhibition, action withholding and action cancelation in adult ADHD. Psychiatry Res. 2012;202(2):132–141. doi: 10.1016/j.pscychresns.2012.02.010. 22475505 [DOI] [PubMed] [Google Scholar]
- Shapiro H.M., Wong L.M., Simon T.J. A cross-sectional analysis of the development of response inhibition in children with chromosome 22q11.2 deletion syndrome. Front. Psychiatry. 2013;4:81. doi: 10.3389/fpsyt.2013.00081. 23966958 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shashi V., Keshavan M.S., Howard T.D., Berry M.N., Basehore M.J., Lewandowski E., Kwapil T.R. Cognitive correlates of a functional COMT polymorphism in children with 22q11.2 deletion syndrome. Clin. Genet. 2006;69(3):234–238. doi: 10.1111/j.1399-0004.2006.00569.x. 16542388 [DOI] [PubMed] [Google Scholar]
- Shidara M., Richmond B.J. Anterior cingulate: single neuronal signals related to degree of reward expectancy. Science. 2002;296(5573):1709–1711. doi: 10.1126/science.1069504. 12040201 [DOI] [PubMed] [Google Scholar]
- Smith S.M., Jenkinson M., Woolrich M.W., Beckmann C.F. Advances in functional and structural MR image analysis and implementation as FSL. Neuroimage. 2004 doi: 10.1016/j.neuroimage.2004.07.051. [DOI] [PubMed] [Google Scholar]
- Soeiro-De-Souza M.G., Stanford M.S., Bio D.S., Machado-Vieira R., Moreno R.A. Association of the COMT Met¹⁵⁸ allele with trait impulsivity in healthy young adults. Mol. Med. Rep. 2013;7(4):1067–1072. doi: 10.3892/mmr.2013.1336. 23440431 [DOI] [PubMed] [Google Scholar]
- Stuphorn V., Taylor T.L., Schall J.D. Performance monitoring by the supplementary eye field. Nature. 2000;408(6814):857–860. doi: 10.1038/35048576. 11130724 [DOI] [PubMed] [Google Scholar]
- Tamm L., Menon V., Ringel J., Reiss A.L. Event-related FMRI evidence of frontotemporal involvement in aberrant response inhibition and task switching in attention-deficit/hyperactivity disorder. J. Am. Acad. Child Adolesc. Psychiatry. 2004;43(11):1430–1440. doi: 10.1097/01.chi.0000140452.51205.8d. 15502603 [DOI] [PubMed] [Google Scholar]
- Thakkar K.N., Congdon E., Poldrack R.A., Sabb F.W., London E.D., Cannon T.D., Bilder R.M. Women are more sensitive than men to prior trial events on the stop-signal task. Br. J. Psychol. 2014;105(2):254–272. doi: 10.1111/bjop.12034. 24754812 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Rooij D., Hartman C.A., Mennes M., Oosterlaan J., Franke B., Rommelse N. Altered neural connectivity during response inhibition in adolescents with attention-deficit/hyperactivity disorder and their unaffected siblings. Neuroimage Clin. 2015;7:325–335. doi: 10.1016/j.nicl.2015.01.004. 25610797 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ventura J., Liberman R.P., Green M.F., Shaner A., Mintz J. Training and quality assurance with the Structured Clinical Interview for DSM-IV (SCID-I/P) Psychiatry Res. 1998;79(2):163–173. doi: 10.1016/s0165-1781(98)00038-9. 9705054 [DOI] [PubMed] [Google Scholar]
- Wechsler D. Psychological Corporation; San Antonio: 1999. WASI Manual. [Google Scholar]
- Wechsler D. Pearson; San Antonio: 2008. Wechsler Adult Intelligence Scale (WAIS-IV) Pearson Assessment. [Google Scholar]
- Whelan R., Conrod P.J., Poline J.-B., Lourdusamy A., Banaschewski T., Barker G.J. Adolescent impulsivity phenotypes characterized by distinct brain networks. Nat. Neurosci. 2012;15(6):920–925. doi: 10.1038/nn.3092. 22544311 [DOI] [PubMed] [Google Scholar]
- Williams N.M., Zaharieva I., Martin A., Langley K., Mantripragada K., Fossdal R. Rare chromosomal deletions and duplications in attention-deficit hyperactivity disorder: a genome-wide analysis. Lancet. 2010;376(9750):1401–1408. doi: 10.1016/S0140-6736(10)61109-9. 20888040 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Distributions of trait impulsivity scores for 22q11DS and ADHD patients. The y-axis represents the percent of subjects who scored at each value of cognitive or behavioral impulsivity, as measured by the BIS-11. Gray bars represent 22q11DS patients and black bars represent ADHD patients. The x-axis represents: (a) cognitive impulsivity scores and (b) behavioral impulsivity scores.
Differences in activation during response initiation and inhibition failure for 22q11DS and ADHD patients. Turquoise maps represent activity from the contrast of ADHD > 22q11DS. (a) Activation maps represent the contrast of Go–Null, to investigate activity related to response initiation. (b) Activation maps represent the contrast of Stop Unsuccessful–Stop Successful, to investigate activity related to inhibition failure.
Regions showing between group differences for ADHD patients, 22q11DS patients, and healthy controls during response initiation (Go vs. Null) and inhibitory failure (Stop Unsuccessful vs. Stop Successful) contrasts. Montreal Neurological Institute (MNI) coordinates represent the location of maximum activation for significant clusters.
Supplementary material.