

Abstract
The transition to a cooked diet represents an important shift in human ecology and evolution. Cooking requires a set of sophisticated cognitive abilities, including causal reasoning, self-control and anticipatory planning. Do humans uniquely possess the cognitive capacities needed to cook food? We address whether one of humans' closest relatives, chimpanzees (Pan troglodytes), possess the domain-general cognitive skills needed to cook. Across nine studies, we show that chimpanzees: (i) prefer cooked foods; (ii) comprehend the transformation of raw food that occurs when cooking, and generalize this causal understanding to new contexts; (iii) will pay temporal costs to acquire cooked foods; (iv) are willing to actively give up possession of raw foods in order to transform them; and (v) can transport raw food as well as save their raw food in anticipation of future opportunities to cook. Together, our results indicate that several of the fundamental psychological abilities necessary to engage in cooking may have been shared with the last common ancestor of apes and humans, predating the control of fire.
Keywords: primate cognition, future-oriented cognition, causal reasoning, cooking, human evolution
1. Introduction
The transition to a diet of cooked foods—involving the use of heat in food preparation—was a fundamental change for our species. Cooked foods are a universal dietary component of all known human societies, and modern humans have a suite of adaptations in the mouth and digestive tract for consuming cooked foods [1,2]. A recent hypothesis claims that cooking emerged early in hominin evolution [3,4] and played an important role in the evolution of several human-specific traits. For example, cooking increases the energy available from foods [5]. As brains are a metabolically expensive tissue [6], this shift to a cooked diet may have contributed to the emergence of larger brains in the human lineage. However, other models suggest that fire was first controlled for other purposes such as for light or heat, and was only used for cooking after a significant delay [7–9]. Thus, understanding when and how this dietary shift occurred is a pressing problem in biology.
Importantly, cooking is a complex behaviour that requires the flexible deployment of multiple cognitive abilities. In order to cook food, humans must exhibit capacities ranging from causal reasoning (as cooking involves the transformation of raw food into more desirable cooked food) to future-oriented cognition (individuals must save raw food they acquire in order to cook it later). Consequently, illuminating the origins of cooking behaviours requires understanding the proximate cognitive mechanisms that support these behaviours. To disentangle what abilities necessary for cooking may have been shared with the last common ancestor of apes and humans, we experimentally tested whether our closest living relative, chimpanzees (Pan troglodytes), possess a suite of domain-general cognitive capacities required for cooking: motivation, patience, inhibitory control, causal understanding and planning. Such studies can illuminate how domain-general cognitive capacities can constrain or enable flexible shifts in behaviour across species, serving as a test case for considering how rapidly cooking may have been adopted after the control of fire. If the cognitive abilities necessary to engage in cooking are also present in chimpanzees, it would support models in which control of fire rapidly led to cooking [3,4,10]. By contrast, a lack of these skills would represent a significant psychological constraint on the evolution of cooking.
In the current set of studies, we therefore examine whether chimpanzees possess this suite of cognitive capacities required for cooking. As chimpanzees neither control fire nor cook food in their natural behaviour, these experiments therefore focus not on whether chimpanzees can actually cook food, but rather whether they can apply their cognitive skills to novel problems that emulate cooking. Importantly, while our experiments focus on how chimpanzees think about the transformation of raw into cooked food, the cognitive skills we explore are not necessarily specific to this context. Many other types of food processing or foraging problems might require similar skills; for example, inhibitory control problems are ubiquitous for foraging animals who must choose between resources of differing values. Our approach is therefore to test a suite of domain-general abilities that are critical prerequisites for cooking behaviour in humans. As humans must flexibly integrate multiple capacities when cooking foods, a lack of any of these skills would therefore represent a significant psychological constraint on the evolution of cooking.
We test a set of five capacities across nine experiments (see figure 1 for overview). First, we confirm that chimpanzees prefer cooked over raw food, and thus possess the basic motivation to engage in cooking. In particular, we replicate the basic finding that chimpanzees prefer cooked foods [10], here using a more minimal cooking procedure than previous studies to better approximate the time delays used in our subsequent tasks. Second, we examine whether chimpanzees are willing to wait to acquire cooked foods: cooking inherently requires an ability to forgo immediately available raw foods to wait for cooked foods, and humans are thought to be unique in their high levels of patience [11]. Third, we assess whether chimpanzees can refrain from eating raw food they possess in order to have it be cooked, an important component of successful cooking. Many primate species, including chimpanzees, have difficulty giving up food already in their possession and show limitations in their self-control when faced with food [12,13]. Fourth, we assess whether chimpanzees have a causal understanding of the transformation of food in a novel cooking process and can flexibly generalize this understanding to new contexts. There are important differences between how humans and other primates reason about causality [14], which could hinder the chimpanzees' ability to exhibit flexible cooking behaviours. Finally, we examine if chimpanzees can transport food across distances and save raw food in anticipation of future opportunities to cook. Cooking often requires that individuals collect food in order to cook it later, but the ability of non-humans to engage in prospection is debated [15,16]. Moreover, previous evidence has suggested that apes can save tools for the future [17,18], but it is unclear whether they can save consumable food items in the same way.
Figure 1.

Study overview. Schematic diagram of all nine experiments.
Our studies present chimpanzees with a novel device that transforms raw food into cooked food—as opposed to cooking with heat, a process that this chimpanzee population have probably seen humans use previously. Moreover, this allowed us to contrast a ‘cooking device’ with a control device that a human acted on in an identical fashion, but which did not alter the state of the chimpanzees' raw food. Such a control is critical to make inferences about the cognitive processes supporting the chimpanzees' behaviours. These studies therefore follow a long tradition in experimental psychology by using novel—and purposefully artificial—contexts to measure the flexible deployment of cognitive abilities. Importantly, we use an analytic approach with a series of experiments that target a specific essential prerequisite for cooking, to assess which aspects are present or absent in chimpanzees. Our experiments build in complexity to assess which of these abilities chimpanzees can use to transform raw into cooked food.
2. General methods
(a). Participants
We tested semi-free-ranging chimpanzees from Tchimpounga Chimpanzee Sanctuary in the Republic of Congo (see the electronic supplementary material). All behavioural tests were approved by Harvard University IACUC (Protocol 11–12) and adhered to local laws (permit no. 013/MRS/DGRST/DMAST). All chimpanzees were socially housed, and most had access to large rainforest enclosures. They had ad libitum water and were not food deprived for testing; the majority of their diet comes from daily feedings of fruits and vegetables. Previous research indicates that these chimpanzees are psychologically healthy, rarely exhibiting aberrant behaviours [19]. Each subject was tested individually in a familiar dormitory room and all tests were voluntary.
(b). General procedure and set-up
The ape and human sat across from each other at a table with a sliding top, separated by wire mesh or bars. Across studies, the experimenter placed the relevant options on the tabletop, and then pushed it forward; chimpanzees could indicate their choice by pointing or touching one option. In studies where the chimpanzees could place food in devices, the same basic procedure was used except that the experimenter held the devices against the wire mesh, within the ape's reach. The experimenter always looked down or along the midline of the table (e.g. not at the options) in order to avoid potential social cuing. In all experiments, the sides on which options were presented were fully counterbalanced. All studies were video-recorded, and a second coder blind to the hypotheses always coded sessions with high reliability (see the electronic supplementary material for all details).
3. Experiment 1: preference for cooked food
In experiment 1, we addressed whether chimpanzees possess the underlying motivation to cook. Previous research has shown that chimpanzees prefer a variety of food when cooked, such as when food is baked until soft or boiled for an extended period [10]. Here, we tested whether chimpanzees preferred cooked white sweet potato, a food type not previously examined, using a different procedure involving minimal food processing. In particular, potato slices were roasted in a dry pan over medium heat (with no additional substances) for approximately 1.5 min, emulating the time ranges used in our cognitive tests. Therefore, the goal of this experiment was to identify a cooked food resource that was suitable for our subsequent cognitive experiments. Both raw and cooked foods were presented at room temperature during testing (see the electronic supplementary material, movie S1).
(a). Methods
Twenty-nine chimpanzees made dichotomous choices between a raw and a cooked slice (3 mm thick, 3 cm in diameter) of potato in a single session (see the electronic supplementary material for photos). This species of potato is a normal component of this populations' diet and is always provided raw. Chimpanzees first completed six exposure trials (three per item in blocks, with order counterbalanced), with only one option available at a time. Second, chimpanzees completed a number discrimination pretest (described in experiment 2). Finally, chimpanzees completed a preference test with 12 choice trials. In the first half of trials, the subject received a small taste of each item prior to their choice (following the methods in [10]), and in the second half they could only smell the slices. There was an inter-trial-interval (ITI) of 20 s after the subject put the food in their mouth.
(b). Results and discussion
Chimpanzees chose the cooked slices on M = 88.8% ± s.e. 2.4 of trials, above chance (t28 = 16.3, p < 0.0001). All 29 individuals chose the cooked potato more than the raw potato (range 66–100% of trials), and 19 apes were individually above chance (binomial: p < 0.05). With previous work [10], this supports the claim that apes have an intrinsic preference for cooked foods. Moreover, in the current test, chimpanzees showed this preference even following a minimal cooking process. We therefore used this food type for subsequent studies.
4. Experiment 2: patience when waiting for cooked food
In experiment 2, we examined whether chimpanzees were willing to forgo raw foods to acquire cooked foods later. While apes can delay gratification in several contexts [20,21], humans are thought to be unique in their high levels of patience [11]. We compared chimpanzees' temporal preferences in two conditions. In the raw-delay condition, chimpanzees chose between receiving one raw piece immediately, or waiting 1 min to receive three raw pieces (electronic supplementary material, movie S2). In the cooked-delay condition, the delayed option provided three cooked pieces (electronic supplementary material, movie S3). This allowed us to assess how cooking impacted the delayed option's value, while holding the immediate alternative constant. Previous studies have shown that chimpanzees are willing to wait longer for rewards of increasing relative amount [22], but can exhibit similar levels of patience for different types of rewards when their amounts are constant (e.g. choosing to delay gratification for larger amounts of food, compared with delaying for the same quantities of tokens [23]). However, it is unknown if chimpanzees would exhibit greater patience when faced with decisions about minimally cooked foods compared with the same quantity of raw food.
(a). Methods
We tested 16 chimpanzees that had completed experiment 1 and passed a basic quantity discrimination pretest (see the electronic supplementary material). Each condition consisted of an introduction session and a test session, administered on consecutive days; condition order was counterbalanced across subjects. In the introduction session, apes completed 14 exposure trials in which only one of the two options was available at a time, in order to introduce them to the delays and food types. In the test sessions, apes first completed an additional four exposure trials, and then completed 10 test trials where they could choose between the two options. In each trial, the experimenter allowed the ape to smell the slices, and then placed them on the respective sides of the table so the ape could choose. If the subject chose the smaller reward, they received it immediately. However, if the subject chose the larger reward, the experimenter waited 1 min before the chimpanzee could access it. There was a 20 s ITI, and each subject had a minimum of 2 days break between conditions.
(b). Results and discussion
Chimpanzees chose the delayed option on 60.0% ± 3.4 trials in the raw-delay condition (one-sample test: t15 = 2.93, p < 0.01), and on 84.4% ± 4.8 trials in the cooked-delay condition (t15 = 7.12, p < 0.0001). A comparison revealed that chimpanzees chose the delayed reward more often in the cooked-delay condition (t15 = 4.58, p < 0.0001; figure 2a). That is, chimpanzees were more willing to pay temporal costs to acquire the cooked food rewards. This study therefore provides one piece of evidence that apes exhibit elevated levels of patience in the context of acquiring more desirable cooked foods, an issue further explored in experiments 8 and 9.
Figure 2.
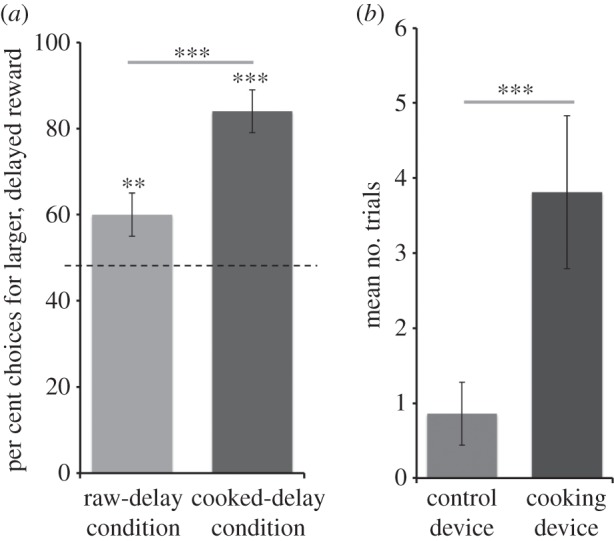
Patience and self-control for cooked foods. (a) Per cent choices for a larger, delayed reward (raw or cooked food across conditions) compared with a smaller, immediately available raw alternative (experiment 2). (b) Trials where chimpanzees placed their raw food into a cooking device, compared with a control device (experiment 4). Error bars indicate s.e.; **p < 0.01; ***p < 0.001.
5. Experiment 3: preference for cooking device
In experiment 3, we introduced the chimpanzees to a novel way to transform raw food into cooked food, designed to emulate a cooking process. When a raw slice of food was placed in the cooking device, it was transformed into a cooked piece after the experimenter shook the container (see the electronic supplementary material, movie S4). The experimenter could act on the control device in the same fashion, but the raw slice was unchanged—thus, controlling for any attraction due to the experimenter's actions. Importantly, these devices were both completely novel, ensuring that the chimpanzees had no previous experience with this transformation problem. Our major goal was to confirm that chimpanzees could infer the (cooked) contents of the cooking device without viewing it (as in [24]), as this basic comprehension was necessary for subsequent studies.
(a). Methods
We tested 23 chimpanzees. Apes first completed a preference test between cooked and raw slices as in experiment 1 (see the electronic supplementary material). They were then introduced to the devices in the exposure phase. The experimenter manipulated each device eight times so the chimpanzee could witness how the devices functioned (in blocks of four trials per device, order counterbalanced). The cooking device consisted of a bowl-shaped container and a lid with a second bowl affixed to its bottom. In each trial, the experimenter showed the chimpanzee a raw piece of food, visibly placed it in the device, put on the lid and shook it 10 times (approx. 5 s duration), and then opened it to give the cooked slice to the subject. The trick was that the main container had a false bottom that allowed us to surreptitiously hide a cooked piece of food (see the electronic supplementary material for photos); the bowl attached to the device's lid contained an object such that it made a noise (like the control device) when shaken. The control device consisted of a transparent plastic container with an opaque lid. The experimenter manipulated it in an identical fashion, but it did not change the raw slice. In 16 subsequent choice trials, the experimenter baited both devices with raw food and placed them on opposite sides of the table (side counterbalanced). The chimpanzee therefore had to select one of the devices before the (cooked) content of the cooking device was revealed, thereby making an inference about its contents. Once the chimpanzee chose, the experimenter then opened the device and gave them the food.
(b). Results and discussion
Chimpanzees selected the cooking device on 87.5% ± 2.7 of trials, above chance (t22 = 13.78, p < 0.0001). This preference was apparent from the first trial, with 17 out of 23 chimpanzees choosing the cooking device (binomial: p < 0.05). To meet our criterion for understanding the basic function of the device, chimpanzees had to individually choose the cooking device above chance (12 of 16 trials, one-tailed binomial). Nineteen out of 23 subjects reached this criterion in one session. The remaining individuals repeated a second session of demonstration and test trials; two additional chimpanzees then passed. Overall, this experiment showed that chimpanzees quickly learned to differentiate between the novel cooking device and a control device that did not alter the food. In addition, we identified 21 chimpanzees meeting criteria to participate in experiment 4.
6. Experiment 4: will chimpanzees choose to cook their own food?
Experiment 3 indicated that chimpanzees differentiated between a device that produced cooked food and one that did not alter the food. However, the critical test is whether chimpanzees can actively place their own raw food into the cooking device to transform it. In experiment 4, we gave chimpanzees raw food to see if they would choose to place it in the cooking device (compared with the control device; see the electronic supplementary material, movies S5 and S6). This problem is challenging both in terms of comprehending the devices' functions, as well as due to the inhibitory control demands. Primates have difficulty giving up food in their hand and show limitations in their self-control when faced with food [12,13]. While chimpanzees can learn to give up their food to acquire a more valuable alternative [22], previous studies have first trained individuals to give up non-food items, and then given them extensive experience with the exchange. Critically, our population of chimpanzees had not been trained to trade any items, so our test measured their spontaneous inferences following minimal experience.
(a). Methods
We tested 21 chimpanzees (who met criteria in experiment 3) in one session with four blocks of trials. In each block, two experimenters first demonstrated the actions of the cooking device and the control device (four demonstrations per device per block) to remind the chimpanzees of devices' functions. One experimenter (E1) sat at the table, holding the one device within reach of the ape. A second experimenter (E2) placed a raw slice of potato on the centre of the table, and then visibly moved it into the cooking device (see the electronic supplementary material for photos). E1 then manipulated the device as in experiment 3 and gave the chimpanzee the resulting food.
Next, the chimpanzees completed four test trials per block (16 total). Here, E1 presented the two (visibly empty) containers within the ape's reach (approx. 15 cm apart on the table, side counterbalanced), while E2 placed a centred slice of raw potato on the table within the chimpanzees' reach (5 cm from the edge). The chimpanzees therefore could decide to eat the raw slice, place it in the cooking device, or place it in the control device. E1 manipulated the device appropriately if the chimpanzee placed food inside; E1 withdrew the devices if the subject ate the food, or after 30 s if they took it but did not eat it (a rare occurrence comprising approx. 1% of trials). Occasionally, the chimpanzees ate part of the slice and placed a smaller piece into the cooking device. When this occurred, E1 surreptitiously put a cooked slice matching its size into the device (out of the ape's view), and then manipulated the device as usual.
(b). Results and discussion
We first examined whether chimpanzees ever put raw food into the devices. Thirteen out of 21 chimpanzees put raw food in the cooking device at least once. Apes that placed food into a device chose the cooking device on 85.3% ± 6.5 of trials, above chance (t12 = 5.40, p < 0.001; figure 2b). We also examined their choices using a more stringent criterion, including only trials where chimpanzees placed a whole piece into a container; these results were similar (see the electronic supplementary material). Trial-by-trial analyses further showed that chimpanzees placed food in the control device at consistently low rates over time, but systematically increased how often they placed food in the cooking device compared with eating it (see the electronic supplementary material). Overall, this indicates that the chimpanzees made a novel inference about their own ability to cook food using these devices, overcoming the strong inhibitory constraints posed by this situation.
7. Experiment 5: replication
In experiment 5, we aimed to replicate our finding that chimpanzees would cook their own food. Our second goal was to identify those individual chimpanzees who cooked reliably, in order to further probe their cognition in subsequent studies.
(a). Methods, results and discussion
We tested 24 chimpanzees (13 participated in experiment 4 one year before, and 11 were naive) using the same procedures as the previous experiments (see the electronic supplementary material for all details). Experiment 5a first replicated the results of experiment 1, showing that chimpanzees preferred cooked over raw potatoes. Twenty-two preferred cooked slices (at the individual level). In experiment 5b, we then tested whether those chimpanzees also had a preference for the cooking device over the control device, as in experiment 3, and found that they all reliably chose the cooking device. Finally, in experiment 5c, we tested 21 chimpanzees' willingness to place their own food into the cooking device using the task from experiment 4. We replicated the finding that chimpanzees selectively place food in the cooking device over the control device and identified 13 chimpanzees who met our criteria by placing food in the cooking device more than half of trials.
8. Experiment 6: do cooking skills generalize to other foods?
Experiments 3 and 4 showed that chimpanzees understood the basic function of the cooking device: they preferred it to the control device, and selectively placed their food in it. However, it is unclear how flexible their knowledge was. In experiment 6, we examined how chimpanzees generalized their experience to new contexts by presenting them with different types of food that could be placed in the devices. In experiment 6a, we first confirmed that chimpanzees preferred a novel food type (carrots) when cooked. Experiment 6b then assessed whether chimpanzees could flexibly modulate whether they placed food in the cooking device. Chimpanzees made decisions about three different food types: raw potato (which they had previous experience with), raw carrot (which they had never experienced or seen being placed in the devices) and pre-cooked potato (which did not need to be transformed; see the electronic supplementary material, movie S7).
(a). Experiment 6a: do chimpanzees prefer cooked carrots?
We tested 29 chimpanzees' preferences for cooked versus raw carrots across 12 trials, following the methods from experiment 1. Chimpanzees chose cooked carrots on 60.1% ± 4.2 of trials, above chance (t28 = 2.42, p < 0.05). The subset of 12 chimpanzees who then participated in experiment 6b (as they met criteria in experiment 5) showed similar results to the overall sample (see the electronic supplementary material). A comparison of these chimpanzees' preferences across experiments 5a and 6a revealed that they had a stronger relative preference for cooked potato than carrots (t11 = 4.06, p < 0.01). That is, while chimpanzees preferred cooked slices for both food types, cooking had a greater impact on their preferences for potatoes.
(c). Experiment 6b: do cooking skills generalize to other foods?
We tested 12 chimpanzees' ability to selectively place raw potato, raw carrot and cooked potato in the cooking device, using the basic set-up from experiment 4. Chimpanzees only had prior experience with placing the raw potato in the cooking device, and the pre-cooked potato did not need to be transformed. If the chimpanzees put any of the items into the cooking device, it would be returned cooked; if they put it in the control device, it would be returned in the identical state. Chimpanzees completed 18 trials in blocks of three trials per type (order counterbalanced), for a total of six trials per type. There was no demonstration period, as these chimpanzees had experience with the devices in experiment 5c. Chimpanzees put the raw potato slice in the cooking device on 75.0% ± 9.3 of trials, more often than they ate it (t11 = 3.36, p < 0.01; figure 3a). By contrast, they preferred to eat the pre-cooked potatoes, placing them into the cooking device on only 26.9% ± 8.8 of trials (t11 = −2.29, p < 0.05). Finally, chimpanzees placed raw carrots in the cooking device in 50.0% ± 9.2 of trials, similar to the rate they ate them (t11 = 0.15, p > 0.7). Importantly, they placed the carrot into the cooking device more often than into the control device (t11 = 4.14, p < 0.005). Chimpanzees generally placed items in the control device at low rates (less than 9.0% of trials) and placed all food types into the cooking device significantly more often than the control device (p < 0.05 for all three types). This shows that chimpanzees could generalize their abilities to a new food type they had never seen being transformed with these devices. Moreover, they flexibly modulated their responses across food types: they placed raw items in the cooking device according to their relative preferences, but refrained from doing so with pre-cooked items.
Figure 3.
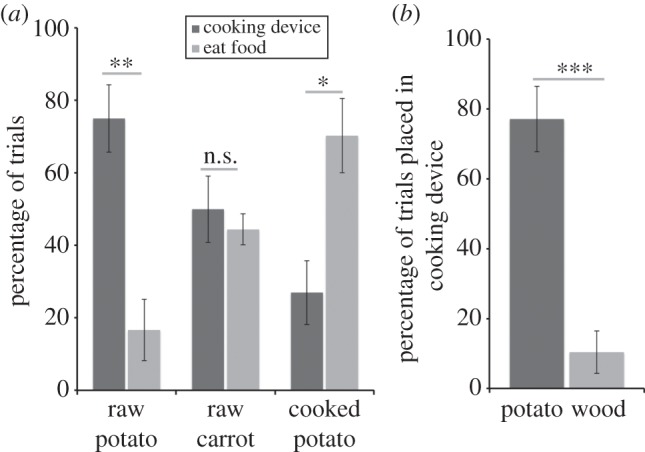
Comprehension of cooking transformation. (a) Percentage of trials where chimpanzees chose to eat versus cook raw potato, pre-cooked potato and novel raw carrot (experiment 6). (b) Percentage of trials where chimpanzees chose to place raw potato in the cooking device compared with an inedible item (experiment 7). Error bars indicate s.e.; *p < 0.05; **p < 0.01; ***p < 0.001.
9. Experiment 7: do chimpanzees selectively cook edible items?
The previous experiment showed that chimpanzees can flexibly generalize their behaviour to new foods. In experiment 7, we further probed what chimpanzees understood about the transformation by assessing their selectivity. We gave chimpanzees a raw potato slice and a visually similar but inedible wood chip simultaneously (electronic supplementary material, movie S8). If chimpanzees thought any item produced cooked slices when placed in the cooking device (e.g. ‘trading’), they should be equally likely to deposit either—and might even prefer to eat the potato and place the wood chip in the device to increase total rewards. By contrast, if chimpanzees view this as a transformation of raw into cooked food, they should specifically place the edible item in the device.
(a). Methods
We tested the chimpanzees from experiment 6b (immediately following that test) on 12 trials with the same basic set-up, except that E1 offered only the cooking device (centre on the table), and E2 placed both the potato and wood on opposite sides (side counterbalanced). If the chimpanzee put an item into the cooking device, E1 manipulated it as in the previous experiments; the raw potato resulting in a cooked slice, and the wood was unchanged.
(b). Results and discussion
Chimpanzees placed raw potato into the cooking device on 77.1% ± 9.4 of trials, but only did so with wood on 10.4% ± 6.1 of trials, a significant difference (t11 = 4.64, p < 0.001; figure 3b). Indeed, nine of 12 chimpanzees placed raw potato into the cooking device on their first trial. The same result was obtained when including only intact pieces of raw potato (see the electronic supplementary material). Importantly, chimpanzees never attempted to put both items into the device—even though results from experiment 9 show that they would place more than one food item in at the same time when given multiple raw slices. That is, they selectively tried to transform only edible items. Together, the results from experiments 6b and 7 indicate that chimpanzees generalized their knowledge to place novel raw food items in the cooking device, but did not place novel inedible items or pre-cooked food into the device. This therefore provides strong evidence that chimpanzees thought that the cooking device transformed raw into cooked food.
10. Experiment 8: will chimpanzees transport food to cook it?
In experiment 8, we examined whether chimpanzees could transport their food to cook it at a different location. Whereas apes eat on the go, human foragers typically collect food and then cook at it a central location [25]—posing a steeper inhibitory control problem given that the food is carried some distance. We contrasted the chimpanzees' behaviours in the near condition, where they received the raw food immediately adjacent to the cooking device (as in the prior experiments), to the far condition, where they had to carry the food across the room to place it in the cooking device (electronic supplementary material, movie S9).
(a). Methods
We tested 13 chimpanzees who met the testing criteria in experiment 5. In the near condition, E2 gave the chimpanzees the food at the testing table where E1 presented the devices (as in the previous studies). In the far condition, E2 provided the food on the other side of the room so chimpanzees had to carry the food approximately 4 m to the testing table (see the electronic supplementary material for diagram). Chimpanzees completed two identical sessions, alternating trial blocks of each condition, for a total of 32 trials per condition. Each session first started with demonstration trials at the testing table (as in experiment 4; four demos per device) followed by four test trials per condition in blocks. In the second half of the session, the demonstration and test trial procedure was repeated (condition block order counterbalanced within sessions).
(b). Results and discussion
Chimpanzees placed the food in the cooking device on 79.6% ± 5.5 of trials in the near condition, and on 60.6% ± 9.0 in the far condition (figure 4a). A repeated-measures ANOVA revealed a main effect of condition (F1,12 = 7.13, p < 0.05, 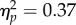 ): chimpanzees were more likely to place food in the devices in the near condition. There was also a main effect of device (F1,12 = 53.22, p < 0.001,
): chimpanzees were more likely to place food in the devices in the near condition. There was also a main effect of device (F1,12 = 53.22, p < 0.001, 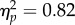 ), and an interaction between condition and device (F1,12 = 7.51, p < 0.05,
), and an interaction between condition and device (F1,12 = 7.51, p < 0.05, 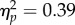 ): chimpanzees placed food in the cooking device more often than in the control device and used the control device at similar low rates across conditions (less than 7% of trials). Similar results were obtained for trials where chimpanzees put a whole piece into the devices (see the electronic supplementary material). These results show that having to transport the food to the cooking location was challenging, as chimpanzees were more successful at placing food in the cooking device in the near condition. Yet, they did succeed in more than half of the far trials, and all but one chimpanzee successfully transported food to the cooking device at least once.
): chimpanzees placed food in the cooking device more often than in the control device and used the control device at similar low rates across conditions (less than 7% of trials). Similar results were obtained for trials where chimpanzees put a whole piece into the devices (see the electronic supplementary material). These results show that having to transport the food to the cooking location was challenging, as chimpanzees were more successful at placing food in the cooking device in the near condition. Yet, they did succeed in more than half of the far trials, and all but one chimpanzee successfully transported food to the cooking device at least once.
Figure 4.
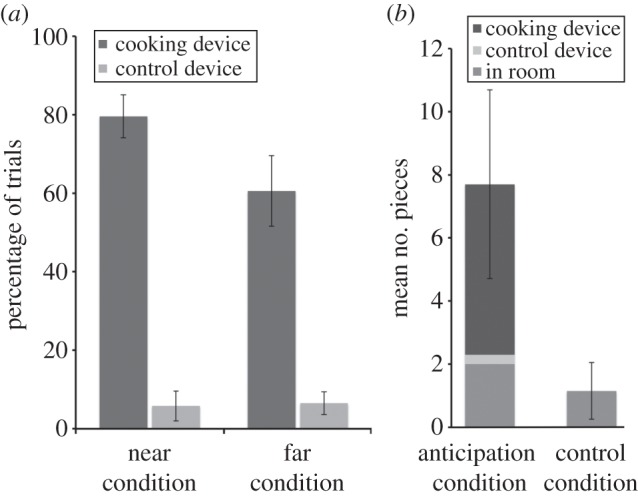
Transporting and saving raw food. (a) Percentage of trials where chimpanzees placed raw food in the cooking device when it was adjacent to them in the near condition, compared with when they had to transport it in the far condition (experiment 8). (b) Number of trials where chimpanzees saved their food in anticipation of future opportunity to place it in the cooking device (anticipation condition) compared with a control condition without such opportunities (experiment 9). Error bars indicate s.e.
11. Experiment 9: will chimpanzees save their food for future cooking?
In experiment 9, we examined whether chimpanzees could save raw food for future cooking opportunities. Previous research has shown that apes can save tools in order to use them in the future and even forgo immediate food rewards in order to obtain the tool [17,18], but human cooking behaviours require actually saving the food itself in order to cook it later. Saving food for future cooking poses a steeper inhibitory problem compared with saving artefacts, as the food has immediate value and could be eaten instantly. In the anticipation condition, chimpanzees initially received three pieces of food, but the experimenter only appeared with the devices after 3 min (electronic supplementary material, movie S10). Thus, chimpanzees had to anticipate the later opportunity to place food in the cooking device (without any concurrent cue). In the control condition, chimpanzees were allotted raw food on the same schedule, but never had any opportunity to place it in the devices. This control condition confirmed that chimpanzees consumed the entirety of the raw food allotment when there was no opportunity to transform it into cooked food.
(a). Methods
We tested the 13 chimpanzees from experiment 8. In the anticipation condition, the basic set-up was the same: E1 presented the devices on one side of the room, and E2 provided raw slices on the opposite side. The main difference was that the there was no testing table present to remove all cues for future opportunities to cook. Rather, E1 appeared with the devices and simply held them against the wire mesh within the chimpanzee's reach (side counterbalanced). At the beginning of a trial, E1 appeared with the two devices for 1 min, and then left. Once E1 was out of sight, E2 gave the chimpanzee three slices of raw potato on the other side of the room. After 3 min, E1 again appeared with the devices for 1 min. If the chimpanzee placed raw potato into a device during this period, E1 manipulated the device appropriately. Because chimpanzees could potentially place multiple slices into the device at the same time (as they received three slices per trial), E1 always matched the resultant cooked amount to the chimpanzee's behaviour. In the control condition, E2 gave three slices of raw potato to the chimpanzee at the same interval (every 4 min), but E1 never appeared with the devices at any point during the session. Chimpanzees completed two conditions of 10 trials (order counterbalanced; see the electronic supplementary material for a diagram).
(b). Results and discussion
We first examined the raw food that chimpanzees did not consume. Chimpanzees saved an average of 7.7 ± 3.0 pieces in the anticipation condition, but only 1.2 ± 0.9 in the control condition (t12 = 2.09, p = 0.058, two-tailed; figure 4b). Individually, seven chimpanzees ate all the raw food in both conditions, five saved more food in the anticipation condition (between seven and 28 slices) and one saved more in the control condition (and never tried to place any food in the cooking device in the anticipation condition). Next, we examined the individuals who placed food in a device at least once in the anticipation condition (individuals could not place food in devices in the control condition; uneaten slices reflect food left in the room). In the anticipation condition, chimpanzees placed an average of 14.0 ± 5.1 slices in the cooking device, but only 0.8 ± 0.5 slices in the control device (t4 = 2.46, p < 0.05, one-tailed). That is, chimpanzees who saved food in the anticipation condition selectively placed it in the cooking device. These results indicate that at least some chimpanzees are capable of saving their food in anticipation of future opportunities to cook it. Overall, chimpanzees saved more food in the anticipation condition, whereas they ate almost all of the food in the control condition—indicating that they were highly motivated to consume the raw food in the absence of cooking opportunities. Moreover, individuals who did save their food in the anticipation condition selectively placed it in the cooking device, compared with the control device.
12. General discussion
Together, these results indicate that chimpanzees and humans share several of the essential psychological capacities needed to cook food. Chimpanzees preferred cooked foods over raw foods and were even willing to pay greater temporal costs in order to acquire cooked foods. Chimpanzees further exhibited self-control by giving up food in their own possession in order to transform it by placing it in a novel cooking device. They had a practical understanding of this basic cooking transformation after minimal experience, generalizing their knowledge about the device to new food types that they had never seen cooked, and selectively attempting to place only edible items in the cooking device. Finally, chimpanzees could transport their food to the cooking device, and even saved their food in anticipation of future cooking opportunities not immediately available. To our knowledge, this is the first evidence that apes can plan for the future by saving food for future transformation.
If chimpanzees possess this set of fundamental psychological capacities for cooking, why do wild chimpanzees not actually cook their food? One obvious reason is that chimpanzees do not control fire, which limits their ability to cook their food and represents another critical capacity that has emerged in the human lineage [26]. But our results also suggest that other psychological constraints may be important. First, the impact of cooking on chimpanzees' typical diet is critical. Chimpanzees rarely consume tubers and roots in the wild, whereas hominin diets uniquely incorporate starchy tubers [27]. Whereas chimpanzees preferred cooked tubers, cooking has little impact on their preferences for more typical chimpanzee foods such as fruits [10]. The psychological barriers (such as inhibitory control) to cooking raw tubers are also probably reduced compared with fruits, as raw tubers are more difficult to digest and have low caloric value [5]. Second, human cooking is generally social in nature: humans pool resources and engage in cooking in the company of others. However, the social nature of cooking creates significant opportunities for theft [3], so increases in social tolerance may have been necessary for cooking to evolve. Chimpanzees may be unwilling to engage in these behaviours when multiple chimpanzees have access to food and cooking devices, monopolizing rather than sharing these resources. Thus, future research should explore how social context influences the expression of these cognitive capacities in chimpanzees.
Overall, these results show that many of the domain-general capacities necessary for cooking are shared with chimpanzees—even though chimpanzees do not naturally engage in cooking. Our results therefore suggest that the earliest adoption of fire may have led rapidly to the development of cooking, supporting claims that cooking originated early in human evolution [3,4]. Furthermore, these results provide some suggestion of how the earliest forms of cooking originated. While many theoretical accounts have focused on the control of fire [26], these results highlight the possible importance of exploitative fire use in the emergence of cooking [28]. Given that chimpanzees in these studies inferred novel opportunities to transform their food after minimal experience, this suggests that early hominins may have also been able to detect and use existing opportunities in their environment to cook foods. Indeed, wild chimpanzees will calmly monitor the movements of natural fires [29], and even actively seek out roasted seeds from burnt habitats [30]. Thus, some opportunistic use of natural fires—underpinned by the types of cognitive capacities examined in the current studies—may have played an important role in bootstrapping the emergence of more complex cooking behaviours that required the active control and maintenance of fire.
Supplementary Material
Supplementary Material
Acknowledgements
We thank Colin Bannard, Steven Pinker, Laurie Santos, Victoria Wobber and Richard Wrangham for comments, and Lauren Fields, Xiuying Jin, Kelly Pitman, Sophie Scolnik-Brower and Deborah Tor for coding assistance. At Tchimpounga, we thank Rebeca Atencia, Debby Cox, the chimpanzee caretakers, Jane Goodall Institute USA, and the Congolese Ministry of Research for hosting our research.
Ethics
All behavioural tests were approved by Harvard University IACUC (protocol 11-12) and adhered to local laws (permit no. 013/MRS/DGRST/DMAST).
Data accessibility
Data are available on Dryad Digital Repository: (doi:10.5061/dryad.rr4bj).
Authors' contributions
F.W. and A.G.R. designed the study, collected and analysed the data, and wrote the manuscript. Both authors gave final approval for publication.
Competing interests
We declare we have no competing interests.
Funding
F.W. was supported by Harvard University and A.G.R. was supported by a L.S.B. Leakey Foundation grant.
References
- 1.Brace CL. 1995. The stages of human evolution. Englewood Cliffs, NJ: Prentice-Hall. [Google Scholar]
- 2.Lucas PW, Ang KY, Sui Z, Agrawal KR, Prinz JF, Dominy NJ. 2006. A brief review of the recent evolution of the human mouth in physiological and nutritional contexts. Physiol. Behav. 89, 36–38. ( 10.1016/j.physbeh.2006.03.016) [DOI] [PubMed] [Google Scholar]
- 3.Wrangham R. 2009. Catching fire: how cooking made us human. New York, NY: Basic Books. [Google Scholar]
- 4.Wrangham RW, Jones JH, Laden G, Pilbeam D, Conklin-Brittain N. 1999. The raw and the stolen: cooking and the ecology of human origins. Curr. Anthropol. 40, 567–594. ( 10.1086/300083) [DOI] [PubMed] [Google Scholar]
- 5.Carmody RN, Weintraub GS, Wrangham RW. 2011. Energetic consequences of thermal and nonthermal food processing. Proc. Natl Acad. Sci. USA 108, 19 199–19 203. ( 10.1073/pnas.1112128108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Aiello LC, Wheeler P. 1995. The expensive tissue hypothesis: the brain and the digestive system in human and primate evolution. Curr. Anthropol. 36, 199–221. ( 10.1086/204350) [DOI] [Google Scholar]
- 7.Bellomo RV. 1994. Methods of determining early hominid behavioral activities associated with the controlled use of fire at FXJJ-20 main, Koobi-Fora, Kenya. J. Hum. Evol. 27, 173–195. ( 10.1006/jhev.1994.1041) [DOI] [Google Scholar]
- 8.Burton F. 2009. Fire: the spark that ignited human evolution. Albuquerque, New Mexico: University of New Mexico Press. [Google Scholar]
- 9.Stahl AB. 1989. Comment on James (1989). Curr. Anthropol. 30, 18–19. [Google Scholar]
- 10.Wobber V, Hare B, Wrangham R. 2008. Great apes prefer cooked food. J. Hum. Evol. 55, 340–348. ( 10.1016/j.jhevol.2008.03.003) [DOI] [PubMed] [Google Scholar]
- 11.Roberts WA. 2002. Are animals stuck in time? Psychol. Bull. 128, 473–489. ( 10.1037/0033-2909.128.3.473) [DOI] [PubMed] [Google Scholar]
- 12.Brosnan SF, Jones OD, Lambeth SP, Mareno MC, Richardson AS, Schapir SJ. 2007. Endowment effects in chimpanzees. Curr. Biol. 17, 1704–1707. ( 10.1016/j.cub.2007.08.059) [DOI] [PubMed] [Google Scholar]
- 13.Boysen ST, Berntson GG, Hannan MB, Cacioppo JT. 1996. Quantity-based interference and symbolic representations in chimpanzees (Pan troglodytes). J. Exp. Psychol. Anim. Behav. 22, 76–86. ( 10.1037/0097-7403.22.1.76) [DOI] [PubMed] [Google Scholar]
- 14.Tomasello M, Call J. 1997. Primate cognition. Oxford, UK: Oxford University Press. [Google Scholar]
- 15.Suddendorf T, Corballis MC. 2007. The evolution of foresight: what is mental time travel, and is it unique to humans? Behav. Brain Sci. 30, 299–351. ( 10.1017/S0140525X07001975) [DOI] [PubMed] [Google Scholar]
- 16.Osvath M, Martin-Ordas G. 2014. The future of future-orientd cognition in non-humans: theory and the empirical case of the great apes. Phil. Trans. R. Soc. B 369, 20130486 ( 10.1098/rstb.2013.0486) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mucalhy N, Call J. 2006. Apes save tools for future use. Science 312, 1038–1040. ( 10.1126/science.1125456) [DOI] [PubMed] [Google Scholar]
- 18.Osvath M, Osvath H. 2008. Chimpanzees (Pan troglodytes) and orangutans (Pongo abelii) forethought: self-control and pre-experience in the face of future tool use. Anim. Cog. 11, 661–674. ( 10.1007/s10071-008-0157-0) [DOI] [PubMed] [Google Scholar]
- 19.Wobber VT, Hare B. 2011. Psychological health of orphan bonobos and chimpanzees in African sanctuaries. PLoS ONE 6, e17147 ( 10.1371/journal.pone.0017147) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Rosati AG, Stevens JR, Hare B, Hauser MD. 2007. The evolutionary origins of human patience: temporal preferences in chimpanzees, bonobos, and human adults. Curr. Biol. 17, 1663–1668. ( 10.1016/j.cub.2007.08.033) [DOI] [PubMed] [Google Scholar]
- 21.Evans TA, Beran MJ. 2007. Chimpanzees use self-distraction to cope with impulsivity. Biol. Lett. 3, 599–602. ( 10.1098/rsbl.2007.0399) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Dufour V, Pelé M, Sterck EHM, Thierry B. 2007. Chimpanzee (Pan troglodytes) anticipation of food return: coping with waiting time in an exchange task. J. Comp. Psychol. 121, 145–155. ( 10.1037/0735-7036.121.2.145) [DOI] [PubMed] [Google Scholar]
- 23.Evans TA, Beran MJ, Paglieri F, Addessi E. 2012. Delaying gratification for food and tokens in capuchin monkeys (Cebus apella) and chimpanzees (Pan troglodytes): when quantity is salient, symbolic stimuli do not improve performance. Anim. Cogn. 15, 539–548. ( 10.1007/s10071-012-0482-1) [DOI] [PubMed] [Google Scholar]
- 24.Call J. 2004. Inferences about the location of food in the great apes. J. Comp. Psychol. 118, 232–241. ( 10.1037/0735-7036.118.2.232) [DOI] [PubMed] [Google Scholar]
- 25.Marlowe FW. 2005. Hunter-gathers and human evolution. Evol. Anthropol. 14, 54–67. ( 10.1002/evan.20046) [DOI] [Google Scholar]
- 26.Wrangham R, Carmody R. 2010. Human adaptation to the control of fire. Evol. Anthropol. 19, 187–199. ( 10.1002/evan.20275) [DOI] [Google Scholar]
- 27.Laden G, Wrangham R. 2005. The rise of the hominids as an adaptive shift in fallback foods: plant underground storage organs (USOs) and australopith origins. J. Hum. Evol. 49, 482–498. ( 10.1016/j.jhevol.2005.05.007) [DOI] [PubMed] [Google Scholar]
- 28.Clark JD, Harris JWK. 1985. Fire and its roles in early hominid lifeways. Afr. Arch. Rev. 3, 3–27. ( 10.1007/bf01117453) [DOI] [Google Scholar]
- 29.Pruetz JD, LaDuke TC. 2010. Reaction to fire by savanna chimpanzees (Pan troglodytes verus) at Fongoli, Senegal: conceptualization of “fire behavior” and the case for a chimpanzee model. Am. J. Phys. Anthropol. 141, 646–650. [DOI] [PubMed] [Google Scholar]
- 30.McGrew WC. 1992. Chimpanzee material culture: implications for human evolution. Cambridge, UK: Cambridge University Press. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data are available on Dryad Digital Repository: (doi:10.5061/dryad.rr4bj).