

GATA2, a member of the GATA transcription factor family, plays a critical role in hematopoiesis,1 vascular2 and neural development. Mutations in the exons and intron 5 of this gene have been identified as the cause of several hematologic disorders.3 GATA2-related disorders include familial myelodysplastic syndromes (MDS)/acute myeloid leukemia (AML);4,5 chronic myeloid leukemia (CML);6 monocytopenia and mycobacterial infection (MonoMAC) syndrome;7 and dendritic cell, monocyte, B and NK lymphoid (DCML) deficiency.8 Patients with MonoMAC syndrome or DCML deficiency exhibit increased susceptibility to infection and often progress to MDS and AML.2,8 Because GATA2 is associated with the development of vascular and lymphatic systems, patients with GATA2 deficiency may present with lymphedema, monosomy 7, and MDS, known as Emberger syndrome.2,9
Monosomy 7 is often identified in patients with MDS, AML, and MDS-related disorders such as mixed myelodysplastic syndrome/myeloproliferative disorder (MDS/MPD) and acquired aplastic anemia (AA). Monosomy 7 has been established as a predictor of poor clinical outcome. Recent studies have identified SETBP1 mutations in hematological malignancies, including atypical chronic myelogenous leukemia (aCML), MDS, and juvenile myelomonocytic leukemia (JMML).10,11 SETBP1 mutations are occasionally accompanied by monosomy 7 and are independent predictors of poor outcome. The incidence of coexistent monosomy 7 and SETBP1 mutation in other hematological disorders is currently unknown.
We aimed to clarify the prevalence of GATA2 and other mutations linked to myeloid malignancies in pediatric hematologic disorders related to MDS and AML. We investigated the incidence of GATA2 mutations in Japanese children with AML (n = 75), JMML (n = 96), AA (n = 75), and familial MDS (n = 4). All exons and intron 5 (wherein pathogenic mutations were documented)3 of GATA2 were analyzed using Sanger sequencing (Online Supplementary Table S1). We detected two mutations in the 75 children with AML. One was a heterozygous c.953C>T, p.A318V missense mutation that had been previously reported in adult AML.12 Furthermore, a t(8;21) chromosomal translocation was found in this patient. The other was a heterozygous c.599insG, p.S201X frameshift mutation. Using CD3+ cells as germline control, we confirmed that these two mutations were of somatic origin (Online Supplementary Figure S1A). In familial MDS, GATA2 mutations were detected in all three families. We did not detect any mutations in our cohort of children with AA or JMML.
We constructed three pedigrees of familial MDS (Table 1). Family 1 had histories of lymphatic malignancy and leukemia. The probands (Patients 1 and 2) were diagnosed with MDS and MonoMAC syndrome with mixed karyotypes. The younger brother (Patient 2) was treated using hematopoietic stem cell transplantation (HSCT). The proband of Family 2 (Patient 4) who suffered from Emberger syndrome died despite treatment using HSCT. Family 3 was also associated with Emberger syndrome. The mother (Patient 6) was diagnosed with lymphedema. The proband (Patient 5) was diagnosed with MDS and received HSCT. In all families analyzed, chromosomal abnormalities affecting chromosome 7 were detected (reciprocal translocation between chromosomes 1 and 7 in Family 1, and monosomy 7 in Families 2 and 3).
Table 1.
Patient characteristics and somatic driver mutations in familial MDS.
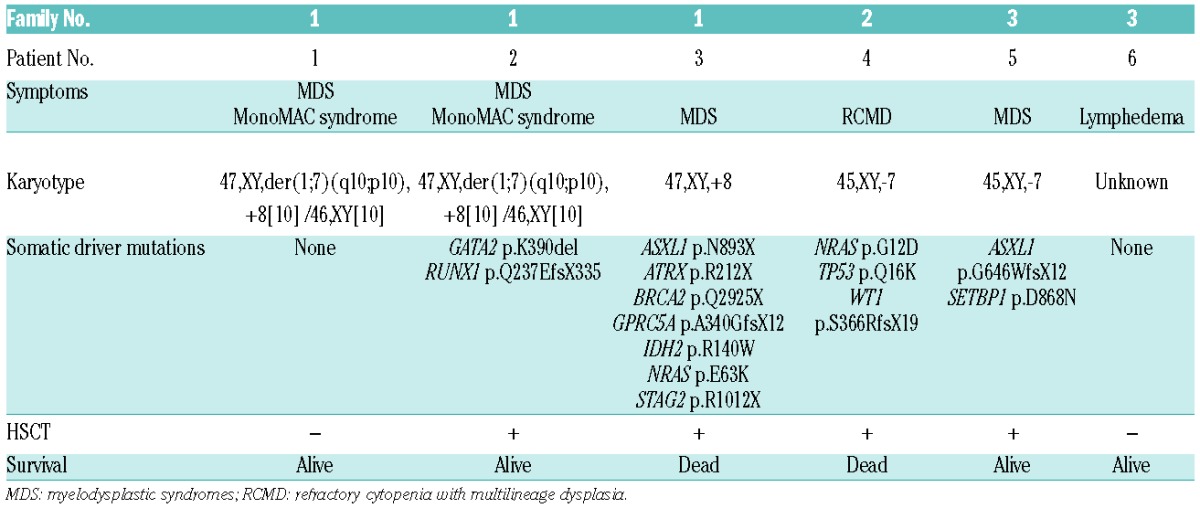
Pedigrees constructed for the three families indicated an autosomal dominant mode of inheritance for the prevalent diseases (Figure 1A). The probands and father from Family 1 had the same germline mutations in GATA2 (Patients 1, 2, and 3; c.892dupT, p.C298LfsX86). The younger brother carried an additional GATA2 mutation (c.1168_1170delAAG, p.K390del). We failed to collect blood samples from all the members of Family 2, thus, using their family history, we inferred that the proband had familial MDS with a confirmed germline nonsense mutation (Patient 4, c.802G>T, p.G268X, Online Supplementary Figure S1B). We examined all five members of Family 3 for mutations and found co-segregation of the clinical phenotype and GATA2 mutations (Patients 5 and 6, c.1018-2A>G, splice site). We confirmed the effect of c.1018-2A>G mutation on splicing (Online Supplementary Figure S2). All mutations were heterozygous. Among the four mutations, three mutations resulted in truncated proteins, wherein two zinc finger domains were lacking, suggesting a complete loss of protein function (Figure 1B). The remaining mutation was an in-frame deletion mutation.
Figure 1.
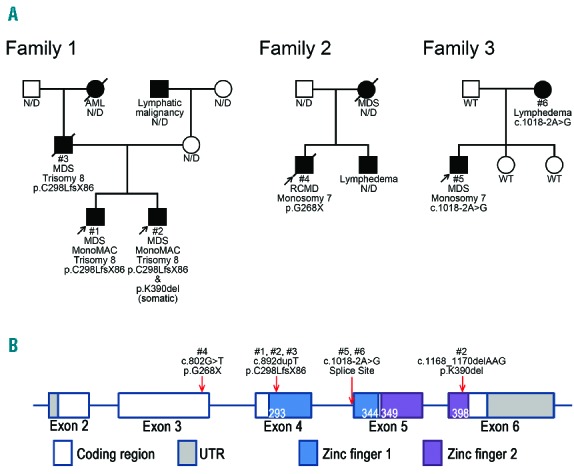
The characteristics of families with familial MDS. (A) Pedigrees of the three included families. Clinical diagnoses and results of genetic analysis are indicated. WT indicates wild type for GATA2 and N/D indicates not done for genetic analysis. Arrows indicate probands. Family 1 was associated with MDS, hematological malignancies, and MonoMAC syndrome. Families 2 and 3 were diagnosed with Emberger syndrome. (B) The distribution of GATA2 mutations. Frameshift mutations identified in Patients 1, 2, and 3 were located in the zinc finger 1 domain, and the 3-bp deletion mutation identified in Patient 2 was located within the zinc finger 2 domain. The nonsense mutation observed in Patient 4 was identified in exon 3. Splice site mutations observed in Patients 5 and 6 were identified in IVS4.
In addition to the inherited mutation, we detected a 3-bp deletion mutation in the younger brother from Family 1 (Patient 2; c.1168_1170delAAG, p.K390del, Figure 2A). To test whether these mutations in exons 4 and 6 occurred in different alleles, we performed polymerase chain reaction (PCR) spanning the exons and cloned the product into a TA cloning vector (Online Supplementary Figure S3). The haplotype of these two mutations was determined using Sanger sequencing of each cloned plasmid. The two mutations were detected in different alleles, indicating that these mutations were compound heterozygous. A haplotype with no mutations was also observed. We concluded that the c.1168_1170delAAG mutation was of somatic origin.
Figure 2.
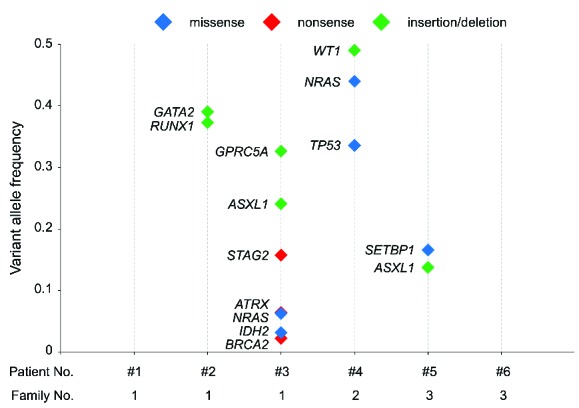
Secondary mutations in familial MDS. Somatic driver mutations detected by target gene sequencing are shown with their variant allele frequencies (VAFs). Blue, red, and green markers indicate missense, nonsense, and insertion/deletion (INDEL) mutations, respectively. The variant allele frequency (VAF) of each mutation is plotted on the y-axis. No somatic mutations were detected in Patients 1 or 6. Regarding the WT1 mutation in Patient 4, half of the raw VAF (0.98) is shown as a loss-of-heterozygosity event, and is strongly indicated by its VAF.
To further investigate somatic mutational events, we performed target gene sequencing of hematological malignancy-related genes in 6 familial MDS patients and 2 AML patients (Online Supplementary Table S2). By covering the entire coding region of 82 genes with 400x coverage, we detected 62 alterations (Online Supplementary Table S3). Based on literature and database searches, we identified 27 of these as driver mutations. Deep sequencing enabled us to measure the variant allele frequency (VAF) of each somatic mutation (Figure 2). In Patients 1 and 6, no somatic events were detected. In the other 4 patients, at least 2 somatic driver mutations were detected in each patient, in addition to a GATA2 germline mutation. In Patient 2, somatic GATA2 (p.K390del) and RUNX1 (p.Q237EfsX335) mutations were detected at similar VAFs (0.39 and 0.37, respectively), suggesting the presence of a single dominant clone. In Patients 3 and 4, VAFs varied even within patients, possibly reflecting a subclonal composition of clonally expanded cells. In Patient 3, at least 7 identifiable driver mutations were detected. GPRC5A (p.R339fs), ASXL1 (p.S892fs), and STAG2 (p.R1012X) mutations were detected at relatively high VAFs, whereas ATRX (p.R212X), NRAS (p.E63K), IDH2 (p.R140W), and BRCA2 (p.Q2925X) mutations were detected at relatively low VAFs. In Patient 4, WT1 (p. A365fs), NRAS (p.G12D), and TP53 (p.Q16K) mutations appeared to occur in tandem. In Patient 5, a SETBP1 (p.D868N) hotspot mutation and an ASXL1 (p.G642fs) mutation were identified. ASXL1 and NRAS mutations were identified as recurrent somatic events in familial MDS.
In our study, we identified germline GATA2 alterations in all 3 families with familial MDS and somatic GATA2 alterations in pediatric sporadic AML (2/75, 3%). In a comprehensive study of adult AML, GATA2 mutations were found in 2% of patients and were considered possible driver mutations.13 The frequency of GATA2 mutations in our pediatric cohort was comparable with that of adult AML. Our findings support the role of GATA2 mutations as driver mutations in AML. A recent report described bi-allelic GATA2 mutations as potential driver mutations in JMML.14
Despite separate GATA2 mutations in Families 2 and 3, haploinsufficiency in this gene led to the clinical manifestation of Emberger syndrome. Although the same germline mutation was detected, the clinical manifestation resulting from germline mutations differed among family members. These findings corroborate previous reports in demonstrating a clear association between GATA2 mutations and familial MDS.5 Our data strongly suggests that additional somatic events are modifiers of these GATA2-related disorders. We detected somatic driver mutations in 4 out of 6 patients with familial MDS. A subclonal composition, suggested by VAFs, was present in 2 patients. In Patient 5, monosomy 7 was detected by clinical karyotyping and a SETBP1 mutation was detected at low VAF. This suggests a serial acquisition of monosomy 7 and subsequent SETBP1 mutation in this patient. A relationship between clinical presentation and mutational status was also observed. In Family 1, Patient 2 carried two driver mutations and required HSCT. In contrast, Patient 1 carried no detectable mutations and HSCT was not required. In Family 3, Patient 6 carried no detectable somatic mutations and presented with lymphedema alone, whereas Patient 5 carried two driver mutations and required HSCT. Overall, serial clonal evolution explains at least part of the variation seen on the clinical presentation of GATA2-related disorders.
We detected at least 20 driver mutations in familial MDS patients. Of these, 7 involved GATA2 (6 germline and 1 somatic) mutations. ASXL1 and NRAS were recurrently found to be mutated. Other driver mutations identified included RUNX1, STAG2, IDH2, TP53, and SETBP1 mutations, which are frequently mutated in adult MDS,15 but have not thus far been reported in patients with GATA2 deficiency. In adult MDS, somatic mutations in several genes predict clinical outcome. Therefore, it may be possible to predict the clinical course of patients with GATA2-related disorders by somatic mutations. For the additional detection of driver events and confirmation of clinical significance, a larger number of patients and a more comprehensive approach, such as whole-exome sequencing, is required.
The proportion of GATA2 mutations in children with sporadic AML, AA, and JMML was previously believed to be low. We detected the presence of not only germline mutations, but also of a somatic mutation of GATA2 in familial MDS. Newly detected secondary mutations may affect the clinical course of familial MDS, and therefore be of prognostic value for clinical decision making, such as transplantation indication and assessment. Further genomic investigations including whole-genome sequencing will lead to an increased knowledge of secondary mutations in GATA2-related disorders.
Acknowledgments
The authors would like to thank all of the clinicians and families who made this study possible by providing samples. The authors also would like to thank Ms. Yoshie Miura, Ms. Yuko Imanishi, and Ms. Hiroe Namizaki for their valuable assistance. The authors acknowledge the Division for Medical Research Engineering, Nagoya University Graduate School of Medicine for technical support of cell sorting and next-generation sequencing.
Footnotes
Funding: this work was supported by “Research on Measures for Intractable Diseases” Project from Ministry of Health Labour and Welfare, Grant-in-Aids from the Ministry of Health, Labor and Welfare of Japan (H23-TA012).
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Rodrigues NP, Janzen V, Forkert R, et al. Haploinsufficiency of GATA-2 perturbs adult hematopoietic stem-cell homeostasis. Blood. 2005;106(2):477–484. [DOI] [PubMed] [Google Scholar]
- 2.Kazenwadel J, Secker GA, Liu YJ, et al. Loss-of-function germline GATA2 mutations in patients with MDS/AML or MonoMAC syndrome and primary lymphedema reveal a key role for GATA2 in the lymphatic vasculature. Blood. 2012;119(5):1283–1291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hsu AP, Johnson KD, Falcone EL, et al. GATA2 haploinsufficiency caused by mutations in a conserved intronic element leads to MonoMAC syndrome. Blood. 2013;121(19):3830–3837, S1–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hahn CN, Chong CE, Carmichael CL, et al. Heritable GATA2 mutations associated with familial myelodysplastic syndrome and acute myeloid leukemia. Nat Genet. 2011;43(10):1012–1017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bodor C, Renneville A, Smith M, et al. Germ-line GATA2 p.THR354MET mutation in familial myelodysplastic syndrome with acquired monosomy 7 and ASXL1 mutation demonstrating rapid onset and poor survival. Haematologica. 2012;97(6):890–894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Zhang SJ, Ma LY, Huang QH, et al. Gain-of-function mutation of GATA-2 in acute myeloid transformation of chronic myeloid leukemia. Proc Natl Acad Sci U S A. 2008;105(6):2076–2081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hsu AP, Sampaio EP, Khan J, et al. Mutations in GATA2 are associated with the autosomal dominant and sporadic monocytopenia and mycobacterial infection (MonoMAC) syndrome. Blood. 2011;118(10):2653–2655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dickinson RE, Griffin H, Bigley V, et al. Exome sequencing identifies GATA-2 mutation as the cause of dendritic cell, monocyte, B and NK lymphoid deficiency. Blood. 2011;118(10):2656–2658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ostergaard P, Simpson MA, Connell FC, et al. Mutations in GATA2 cause primary lymphedema associated with a predisposition to acute myeloid leukemia (Emberger syndrome). Nat Genet. 2011;43(10):929–931. [DOI] [PubMed] [Google Scholar]
- 10.Makishima H, Yoshida K, Nguyen N, et al. Somatic SETBP1 mutations in myeloid malignancies. Nat Genet. 2013;45(8):942–946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sakaguchi H, Okuno Y, Muramatsu H, et al. Exome sequencing identifies secondary mutations of SETBP1 and JAK3 in juvenile myelomonocytic leukemia. Nat Genet. 2013;45(8):937–941. [DOI] [PubMed] [Google Scholar]
- 12.Fasan A, Eder C, Haferlach C, et al. GATA2 mutations are frequent in intermediate-risk karyotype AML with biallelic CEBPA mutations and are associated with favorable prognosis. Leukemia. 2013;27(2):482–485. [DOI] [PubMed] [Google Scholar]
- 13.Brewin J, Horne G, Chevassut T. Genomic landscapes and clonality of de novo AML. N Engl J Med. 2013;369(15):1472–1473. [DOI] [PubMed] [Google Scholar]
- 14.Stieglitz E, Liu YL, Emanuel PD, et al. Mutations in GATA2 are rare in juvenile myelomonocytic leukemia. Blood. 2014;123(9):1426–1427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Haferlach T, Nagata Y, Grossmann V, et al. Landscape of genetic lesions in 944 patients with myelodysplastic syndromes. Leukemia. 2014;28(2):241–247. [DOI] [PMC free article] [PubMed] [Google Scholar]