

Abstract
Antimalarial drug resistance is one of the major challenges in global efforts of malaria control and elimination. In 1998, chloroquine was abandoned and replaced with sulfadoxine/pyrimethamine, which in turn was replaced with artemether/lumefantrine for the treatment of uncomplicated falciparum malaria in 2004. Sulfadoxine/pyrimethamine resistance is associated with mutations in dihydrofolate reductase (Pfdhfr) and dihydropteroate synthase (Pfdhps) genes. The prevalence of mutation in Pfdhfr and Pfdhps genes were evaluated and compared for a total of 159 isolates collected in two different time points, 2005 and 2007/08, from Pawe hospital, in North Western Ethiopia. The frequency of triple Pfdhfr mutation decreased significantly from 50.8% (32/63) to 15.9% (10/63) (P<0.001), while Pfdhps double mutation remained high and changed only marginally from 69.2% (45/65) to 55.4% (40/65) (P = 0.08). The combined Pfdhfr/Pfdhps quintuple mutation, which is strongly associated with sulfadoxine/pyrimethamine resistance, was significantly decreased from 40.7% (24/59) to 13.6% (8/59) (P<0.0001). On the whole, significant decline in mutant alleles and re-emergence of wild type alleles were observed. The change in the frequency is explained by the reduction of residual drug-resistant parasites caused by the strong drug pressure imposed when sulfadoxine/pyrimethamine was the first-line drug, followed by lower fitness of these resistant parasites in the absence of drug pressure. Despite the decrease in the frequency of mutant alleles, higher percentages of mutation remain prevalent in the study area in 2007/08 in both Pfdhfr and Pfdhps genes. Therefore, further multi-centered studies in different parts of the country will be required to assess the re-emergence of sulfadoxine/pyrimethamine sensitive parasites and to monitor and prevent the establishment of multi drug resistant parasites in this region.
Introduction
For more than 40 years, chloroquine was the first line anti-malarial drug in Ethiopia for the treatment of uncomplicated Plasmodium falciparum malaria. In 1998, chloroquine (CQ) was abandoned and replaced with sulfadoxine/pyrimethamine (SP). In 2004, SP was replaced with artemether/lumefantrine (AL) for the treatment of uncomplicated falciparum malaria [1,2]. SP is no longer recommended for the treatment of malaria but it remains a vital tool to reduce the burden of malaria in Africa. The World Health Organization (WHO) recommends SP for intermittent preventive treatment in pregnancy (IPTp), in infancy (IPTi) and in childhood (IPTc) [3–5]. In sub-Saharan Africa, 39 countries had adopted intermittent preventive treatment in pregnancy between 1993 and 2007 [6].
Point mutations in P. falciparum dihydrofolate reductase (Pfdhfr) and dihydropteroate synthase (Pfdhps) genes are known to confer resistance to pyrimethamine and sulfadoxine, respectively [7,8]. Pyrimethamine resistance is generally caused by three mutations in Pfdhfr gene, namely: N51I, C59R, and S108N [9]; sulfadoxine resistance in sub Saharan Africa is caused by substitutions S/A436F, A437G, K540E, A581G, and A613S/T in a variety of combinations in Pfdhps gene [10]. Parasites carrying all five mutations, Pfdhfr triple mutant (51I, 59R, and 108N) and Pfdhps double mutant (437G and 540E), commonly called quintuple mutations have been strongly associated with SP treatment failure in sub-Saharan Africa [11–13]. Additionally, Pfdhfr (I164L) and increased prevalence of the Pfdhps A581G mutation has been well documented and are linked with increased therapeutic failure of SP in southeastern Africa [14–17]. WHO recommends the implementation of SP for intermittent preventive treatment only if the prevalence of the K540E mutation (and thus the quintuple mutation) is <50% [18].
Following the withdrawal of CQ in 1993, Pfcrt mutation (K76T) gradually decreased and disappeared completely by 2001 in Malawi [19]. Similar findings have been reported in Tanzania [20], Kenya [21] and China [22]. This decline was correlated with the return of the clinical efficacy of CQ for the treatment of falciparum malaria in Malawi [23]. In Cambodia, however, alleles conferring CQ and SP resistance occur at a high frequency after the withdrawal of these drugs [24]. In Venezuela, the complete fixations of mutant Pfdhfr and Pfdhps alleles eight years after the withdrawal of SP were reported [25]. Zhou et al., 2009, reported decrease in the frequency of Pfdhfr and Pfdhps gene mutations after the withdrawal of SP in Peru. Decline in Pfdhfr and Pfdhps mutations after the withdrawal of SP has also been reported from Northern Ethiopia [26], Tanzania [27] and southern Mozambique [28].
In Ethiopia, high level of Pfdhfr triple mutations and Pfdhfr/Pfdhps quintuple mutations was reported from Jimma, Dilla and Bahirdar in 2004/05 [26,29,30]. A study conducted in Northern Ethiopia (Bahirdar) showed a significant decline in the triple Pfdhfr mutation from 78.6% in 2005 to 56.4% in 2008 and the quintuple mutations reduced from 60.6% to 37.2% between 2005 and 2008 [31]. In this study we evaluated change in the frequency of the mutant and wild type Pfdhfr and Pfdhps markers after the withdrawal of SP in 2004 in a perennial transmission setting. The objective of this study was to determine the change in the frequency of Pfdhfr and Pfdhps mutant and wild alleles in P. falciparum isolates collected in 2005 and 2007/08.
Materials and Methods
Study area
The study was conducted in the rural town of Pawe, in the North Western part of Ethiopia (Fig 1). The area is located at an altitude of 1050 meters above sea level with a mean annual temperature ranging from 16.2°C to 32.2°C, and the mean annual rainfall between 980 and 1200 mm occurring in two seasons from March to May and from June to December (Pawe Agricultural Research Centre). Pawe was one of the 14 sentinel sites, that were eco-epidemiologically selected, for drug resistance studies by the Federal Ministry of Health [2].
Fig 1. Map of the study area.

Study population
Plasmodium falciparum samples were collected from 80 patients in 2005 by the Ethiopian Public Health Institute (formerly known as Ethiopian Health and Nutrition Research Institute (EHNRI)). In 2007/08, we screened 602 patients attending Pawe hospital and 79 of them were positive for Plasmodium falciparum malaria by light microscopy. We included patients of all age, both sex and only those who lived in the study area for at least two years. The same inclusion criteria were used in 2005. Patients with severe malaria [32], severe malnutrition and serious underlying diseases according to the physicians comment were excluded from the study.
A finger-prick blood sample was taken for thick and thin blood smear. An experienced microscopist examined the Giemsa-stained blood smears. If the patient was infected with P. falciparum mono-infection and fulfilled the inclusion criteria, two to three drops of blood was collected on to a whatman filter paper (Krackeler Scientific Inc., New York). The filter paper was labeled, dried and stored at -20°C. After the blood sample was collected, additional thin and thick blood smears were made labeled and transported to EHNRI for re-confirmation by a different microscopist and parasite count. From the thick smears, parasite density was determined by counting the number of asexual parasites against 200 white blood cells. The thin smear was used for species determination. The national and Addis Ababa University (Department of Biology) ethical boards independently approved this study. A written informed consent was translated to the local languages of the study area and approved by the ethical review boards. During sample collection a written informed consent was obtained from each participant and parents or guardian and appropriate drug was given free of charge for patients according to the physician subscription.
DNA extraction and PCR amplification
Parasite genomic DNA was extracted from dried blood spot as previously described [33]. Pfdhfr and Pfdhps genes were amplified from 2–3μl of genomic DNA for the primary PCR and 2μL of the PCR product for the nested PCR. The sequence of the primers, PCR and cycling conditions are shown in S1 Table. Positive control genomic DNA from 3D7, Dd2, T994, HB3, T996, SL/D6 and IEC513/86 (S2 Table) and negative control were included in all PCR reactions. PCR products were visualized in agarose gel containing ethidium bromide. The bands were analyzed and the PCR products were used for the dot blot hybridization experiment. PCRs were repeated and optimized for the negative samples.
Preparation of dot blots
PCR products were chemically denatured, heated and then neutralized with equal volume ammonium acetate (pH = 7). The denatured PCR product was blotted by direct application onto Gene screen nylon membranes using dot blotter (Bio-Rad, UK). The membrane was incubated for 30 minutes and neutralized by washing in saline-sodium citrate buffer (2X SSC) for 60 seconds. The membrane was immersed in 0.4M Sodium Hydroxide for 30–60 seconds to ensure complete denaturation of immobilized DNA, and then rinsed in neutralizing buffer for 30 seconds. The cross-linking between the applied DNA and the membranes was done using UV cross-linker as previously described [34]. In each blot appropriate positive controls for Pfdhfr and Pfdhps (S2 Table) and negative control were included.
Labeling of oligonucleotide probes
Ten oligonucleotide probes were labeled with radioactive 32P as described in previous studies (Table 1) [34,35]. Briefly, 1μl (10pMol) of each oligonucleotide probe (MWG Biotech, Germany) mixed with 1μl Ready-To-Go T4 Polynucleotide Kinase (Amersham Pharmacia Biotech, UK) and 47μl of DNase free water. 1μl of [γ-32P] ATP (Institutes of Isotopes, Hungary) was added and incubated for 30 minutes at 37°C. The reaction was stopped by 5μl of 250mM EDTA. The unincorporated [32p]-γ-ATP was removed by re-suspending the mixture in column according to the manufacturer guidelines (Amersham Pharmacia Biotech, UK). The purified sample was stored in -20°C until further use. Strict personal and environmental safety protocols of the national radiation regulation authority were followed.
Table 1. List of oligonucleotide probes used for the detection of polymorphism in Pfdhfr and Pfdhps genes.
| Gene | Amino acid | Probe sequences | Hybridization Temperature |
|---|---|---|---|
| Pfdhfr | Ser 108 | 5'-AACAAGCTGCGAAAGCATTCCAA-3' | 54°C |
| Asn 108 | 5'-AACAAACTGGGAAAACATTCCAA-3' | 54°C | |
| Ile 51 | 5'-CCATGGAAATGTATTTCGCTAG-3' | 45°C | |
| Asn 51 | 5'-CCATGGAAATGTAATTCGCTAG-3' | 54°C | |
| Arg 59 | 5'-GAAATATTTTCGTGCAGTTAC-3' | 48°C | |
| Cys 59 | 5'-GAAATATTTTTGTGCAGTTAC-3' | 58°C | |
| Pfdhps | Gly 437 | 5'-GAATCTTCTGGTCCTTTT-3' | 43°C |
| Ala 437 | 5'-GAATCCTCTGCTCCTTTT-3' | 51°C | |
| Lys 540 | 5'-CAATGGATAAACTAACAA-3' | 35°C | |
| Glu 540 | 5'-CAATGGATGAACTAACAA-3' | 35°C |
Dot blot hybridization
The blot membrane was immersed in a hybridization buffer (Final concentration: 5X SSPE, 5X Denhardt’s reagent, 0.5% SDS, 59.9% DNase-free water and 0.01 mg/ml sonicated salmon sperm) (Gibco BRL, UK) and incubated for 30 minutes at the specific hybridization temperature of each probe (Table 1). The 32P labeled oligonucleotide probe (1μl for every 1ml of the hybridization buffer) was added and hybridized overnight in rotary rocker hybridization oven. The membrane was washed once in 2X SSC and twice in 1X SSC/0.1% SDS with 20 and 10 minutes incubation, respectively. The washing solution was poured off and disposed according to local radiation regulations. The blot was wrapped in cling film and taped into an autoradiography cassette (Amersham Biosciences, France). It was exposed to X-ray film (Amersham Bioscience, France) at –70°C for 12–24 hours. The film was developed and results were scored. Following autoradiography, probes were stripped off by two washes in 0.1M Sodium Hydroxide and rinsed in 5XSSC [34,35]. The blots were then re-hybridized with other probes as required.
Data analysis
The data was scored for each sample as mutant, wild and mixed (mutant and wild) based on the detection of the specific probes. The scored raw data is attached in S1 Dataset. We scored double, triple and quintuple mutations based on the detection of specific alleles in the isolates. For example, if we detect three Pfdhfr mutations in the same isolate, we score this isolate as a triple Pfdhfr mutant and the same for the Pfdhps and the quintuple mutation. The changes in the frequencies of mutations between groups were compared using the F-test. The data was analyzed using GraphPad Prism v6.0 (GraphPad Software Inc).
Results
Malaria prevalence in the study area
A retrospective data was obtained from clinical records at Pawe hospital for the year 2005, 2006 and a three-month record from November 2007 to January 2008. In 2005/06 prevalence of malaria by microscopy was 23.1% (2440/10569); 19.8% (1671/8444) in 2006 and 14.78% (89/602) in the three month period from November, 2007 to January, 2008. By microscopy, Plasmodium falciparum is the pre-dominant species in the study area with 88.6%, 69.24% and 88.76% of all malaria cases in 2005, 2006 and 2007/08 (3 month) respectively. P. vivax prevalence was 9.4% in 2005/06, 3.17% in 2006/07 and 7.87% in 2007/08 (3 month) and the rest were infected with mixed infections (P. falciparum and P.vivax) (Table 2). The retrospective data showed that between 2005 and 2007 malaria accounted for 60% of admission and about 30% mortality in the hospital.
Table 2. Prevalence of malaria based on clinical records in Pawe general hospital from 2005/06 to 2007/08.
| Year | Total (N) | Positive to malaria N (%) | P. falciparum N (%) | P.vivax N (%) | Mixed (Pf & Pv) N (%) |
|---|---|---|---|---|---|
| 2005/06 | 10569 | 2440 (23.1) | 2162(88.6) | 229(9.4) | 49(2) |
| 2006/07 | 8444 | 1671 (19.8) | 1157(69.24) | 53(3.17) | 461(27.59) |
| 2007/08* | 602 | 89 (14.78) | 79(88.76) | 7(7.87) | 3(3.37) |
*Only 3 months data is shown.
Characteristics of study participants
The data presented in this study is based on 159 samples tested for the change in the frequency of Pfdhfr and Pfdhps mutant and wild type alleles for the year 2005 (n = 80) and 2007/08 (n = 79). The study participants between the two study periods are comparable in terms of age, sex and parasite density (Table 3). The parasite counts were in the range of 2200–180,000/μl (Geometric mean = 13554.28/μl) and 1,840–200,000/μl (Geometric mean = 14757.68/μl) in 2005 and 2007/08, respectively. Among these 80 and 79 microscopy positive samples, we successfully amplified 63 samples for Pfdhfr in 2005 and 2007/08 and 65 for Pfdhps gene by the nested PCR.
Table 3. Characteristics of the study participants.
| Characteristics | 2005 N (%) | 2007/8 N (%) | |
|---|---|---|---|
| Sex | Male | 38(47.5) | 40 (50.63) |
| Female | 42(52.5) | 39(49.37) | |
| Age (year) | Range | 1–46 | 1/2–60 |
| ≤5 | 23(28.75) | 17(21.5) | |
| 6–14 | 17(21.25) | 28(35.4) | |
| ≥15 | 40(50) | 34(43.1) |
Prevalence of Pfdhfr and Pfdhps mutations
The prevalence of mutations in five codons, three for the Pfdhfr gene (N51I, C59R, and S108N) and two codons of the Pfdhps gene (A437G, and K540E) were analyzed to assess the change in the frequency from 2005 to 2007/8 after the withdrawal of SP from the study area. In 2005, 92% (58/63) isolates had mutation in the Pfdhfr codon 108, 82% (52/63) at codon 59 and 62% (39/63) at codon 51. About 75.4% (49/65) of the isolates had mutation in one of the Pfdhps codon in 2005. In 2007, 74.6% (47/63) of the isolates exhibited the Pfdhfr core mutation N108. In 2005, only four isolates displayed wild type alleles in Pfdhfr codon 108 (Table 4).
Table 4. Number and frequency of Pfdhfr and Pfdhps mutant and wild type alleles in 2005 and 2007/08 in Pawe, North Western, Ethiopia.
| Year | Codon | N | Wild type N (%) | Mutant type N (%) | Mixed N (%) |
|---|---|---|---|---|---|
| 2005 | Pfdhfr S108N | 63 | 4 (6.35) | 58 (92.1) | 1 (1.6) |
| Pfdhfr C59R | 63 | 8(12.7) | 52 (82.5) | 3 (4.8) | |
| Pfdhfr N51I | 63 | 13(20.6) | 39 (61.9) | 11 (17.5) | |
| Pfdhps A437G | 65 | 13 (20.0) | 49 (75.4) | 3 (75.4) | |
| Pfdhps K540E | 65 | 10 (15.4) | 52 (80.0) | 3 (4.6) | |
| 2007/08 | Pfdhfr S108N | 63 | 16 (25.4) | 47 (74.6) | 0 (0) |
| Pfdhfr C59R | 63 | 26 (41.3) | 35 (55.6) | 2 (3.2) | |
| Pfdhfr N51I | 63 | 30 (47.6) | 16 (25.4) | 17 (27.0) | |
| Pfdhps A437G | 65 | 21 (32.3) | 44 (67.7) | 0 (0) | |
| Pfdhps K540E | 65 | 24 (36.9) | 41 (63.1) | 0 (0) |
Temporal decline in Pfdhfr and Pfdhps mutations
The frequency of the Pfdhfr S108N mutation decreased from 92% (58/63) in 2005 to 74.6% (47/63) (P = 0.002) in 2007/08. The relative frequencies of the Pfdhfr C59R and N51I mutations decreased from 62% (39/63) to 25.4% (16/63) (P<0.0001) and from 82.5% (52/63) to 55.5% (35/63) (P <0.0001) in 2005 and 2007/08, respectively. Pfdhps mutation at position 437 decreased from 75.4% (49/65) to 69.8% (44/65) (P = 0.35) and at position 540 decreased from 80% (52/65) to 63% (41/65) in 2005 and 2007 (P = 0.012), respectively (Fig 2). Change in the allelic frequencies of the Pfdhfr triple (S108N/C59R/N51I), Pfdhps double (A437G/K540E) and quintuple (the three Pfdhfr and two Pfdhps) mutations mirrored the trends observed in the prevalence analyses. The frequency of parasites carrying quintuple mutations decreased from 40.7% (24/59) to 13.6% (8/59) (P<0.0001) between 2005 and 2007/08. A significant decrease in the frequency of Pfdhfr triple mutation (S108N/C59R/N51I), from 50.8% (32/63) to 15.87% (10/63) (P<0.001), was detected. For Pfdhps double mutation (A437G/K540E), there was only a marginal difference between the two surveys with a frequency 69.2% (45/65) and 55.4% (40/65) (P = 0.08) in 2005 and 2007/08, respectively (Fig 3).
Fig 2. Temporal changes in the frequency of Pfdhfr and Pfdhps single nucleotide mutations between 2005 and 2007/08.
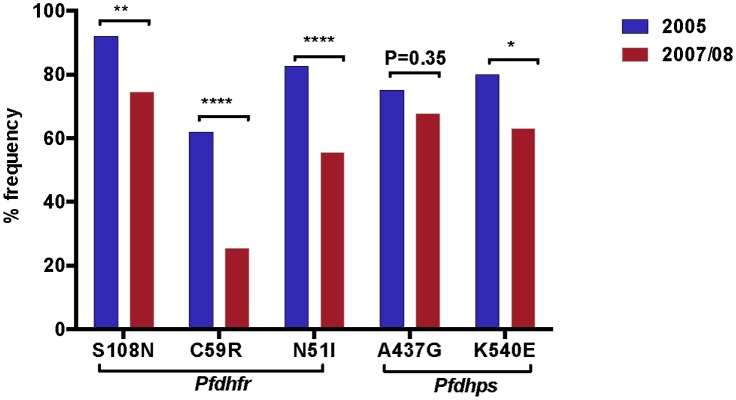
The asterisk indicates the statistical significance of the difference according to Fisher’s exact test.
Fig 3. Temporal changes in the frequency of Pfdhfr and Pfdhps combined mutations.

The percentage frequencies of Pfdhfr triple (S108N/C59R/N51I), Pfdhps double (A437G/K540E) and quintuple mutations (S108N/C59R/N51I/A437G/K540E) were compared between 2005 and 2007/08. The asterisk indicates the statistical significance of the difference according to Fisher’s exact test.
Temporal changes in Pfdhfr and Pfdhps wild types
A significant increase in the frequency of Pfdhfr and Pfdhps wild type was observed in 2007/08 as compared to 2005. Wild type allele in Pfdhfr codon 108-Ser increased from 6.35% (4/63) to 25.4% (16/63) (P = 0.0005), codon 51-Asn from 12.7%(8/63) to 41.3% (26/63) (P<0.0001), and codon 59-Cys from 20.63% (13/63) to 47.62% (30/63) (P<0.0001). For Pfdhps 437-Ala marginally increased from 20% (13/65) to 32.3% (21/65) (P = 0.08) and 540-Lys wild type allele significantly increased from 15% (10/65) to 36.9% (24/65) (P = 0.0006) (Fig 4). Furthermore, triple Pfdhfr wild types were not detected in 2005 but we found the triple wild type Pfdhfr in 11.11% (7/63) (P = 0.0007) of the isolates in 2007/08. Pfdhps double wild types increased from 13.8% (9/65) to 30.8% (20/65) (P = 0.0063). The Pfdhfr/Pfdhps quintuple wild type increased significantly from zero to 10.2% (P = 0.0015), after the withdrawal of SP in 2004 (Fig 4).
Fig 4. Change in the frequency of Pfdhfr and Pfdhps wild type alleles.

The percentage frequency of wild type alleles in Pfdhfr single and triple (Ser-108/Asn-51/Cys-59), Pfdhps single and double (Ala-437/Lys-540) and quintuple wild types (Ser-108/Asn-51/Cys-59/Ala-437/Lys-540) were compared between 2005 and 2007/08. The asterisk indicates the statistical significance of the difference according to Fisher’s exact test.
Discussion
There are widely differing malaria endemicity and transmission regions in Ethiopia ranging from a sporadic cases in highland fringe areas to a perennial transmission south western lowlands [2]. Considering the substantial burden of malaria in this country, in 2004 multi-site survey demonstrated mean SP treatment failure rates of 36% and 72% within 14 and 28 days of follow-up, respectively [1]. Following this report, SP was replaced with artemisinin-based combination therapy, artemether/lumefantrine (AL). Different studies show that CQ withdrawal from use for a number of years has reversed resistance based on prevalence of Pfcrt resistance marker [19]. This was only possible because CQ was totally banned and it was not available to health facilities. Similarly, SP was banned in Ethiopia since 2004 and unlike many other African countries it has not been used for intermittent preventive treatment. This gives us the opportunity to study the temporal changes in the frequency of molecular markers associated with sulfadoxine/pyrimethamine resistance in the absence of drug pressure in a perennial malaria transmission setting. We compared 159 samples collected in two time points (2005 and 2007/8) by PCR based dot blot hybridization technique. We found a significant declining trend of Sulfadoxine/pyrimethamine mutation markers and re-emergence of wild parasites after the withdrawal of the drug in 2004.
The quintuple mutation (Pfdhfr S108N/N51I/C59R/Pfdhps A437G/K540E), which was strongly correlated to SP treatment failure of P. falciparum malaria in south eastern Africa [13], was decreased significantly from 40.7% in 2005 to 13.6% in 2007/08. The level of mutation is much higher as compared to previous report of 1.4% prevalence in the absence of SP in the Ashanti Region of Ghana [36]. The quintuple predictor [13], Pfdhfr C59R and Pfdhps K540E double mutant, showed a statistically significant change from 51% in 2005 to 16.5% in 2007/08 (P<0.0001) supporting the hypothesis that the resistant strains are less fit in the absence of drug pressure.
Triple mutations in the Pfdhfr gene are associated with 60 to 70% rates of SP treatment failure [13]. The Pfdhfr triple mutations decreased from 51% to 16% (P<0.001) and the Pfdhps double mutation decreased from 69% and 55.4% (P = 0.08) indicating that Pfdhfr allele with triple mutations is being replaced faster than the Pfdhps allele with double mutations as the drug pressure was removed from the population, suggesting that pyrimethamine sensitivity is reemerging faster than sulfadoxine sensitivity. This is also supported by the increase in the triple wild Pfdhfr types from zero to 11.1% (P<0.0001) and Pfdhps double wild types from 13.8% to 30.8% (P = 0.006) in the two time points. These findings contrasts with the report from Peru, which showed a decrease in Pfdhps 437/540/581 mutation from 47% in 1997 to zero in 2005/6 while Pfdhfr triple mutation was decreased to 17% [37]. However, the transmission of malaria in this part of Ethiopia is higher than Peru and the time difference in the Peruvian study was five years. This indicates assessing the change in frequency of mutations in larger time points will give better resolution depending on the intensity of transmission. A similar study in southern Mozambique between 1999 and 2004 showed that the frequency of dhfr triple mutation increased from 1999 to 2004, however, the frequency of dhps double mutation increased to 2001 but declined to baseline levels by 2004. Quintuple mutations increased from 1999 to 2001 but decreased by 2004, which corresponded with replacement of SP with artemether—lumefantrine as malaria treatment policy in neighboring KwaZulu-Natal, South Africa [28]. Similarly, in Tanzania a community-based study compared the prevalence of mutation between IPTi intervention and comparison groups showed an increase in Pfdhfr triple mutation and stabilized frequencies of Pfdhps double mutant parasites following the withdrawal of SP as first-line treatment [27].
The slower decline in prevalence of mutations in Pfdhps (Figs 2 and 3) suggests that these mutations may be less deleterious to parasite fitness than are Pfdhfr mutations in this study area. Although the precise relationship between mutations in the Pfdhfr and Pfdhps genes in clinical sulfadoxine/pyrimethamine resistance is unclear, previous data showed that the presence of a sensitive Pfdhfr allele is highly predictive of sulfadoxine/pyrimethamine treatment success irrespective of the Pfdhps allele [38].
We found a significant decrease in all single mutations except Pfdhps A437G (from 75.4% to 69.8%, P = 0.35). These findings contradicts with a recent report from Bahir Dar, which is 230km away from this study area [26]. The prevalence of high level of single and combination mutations in Pfdhfr and Pfdhps alleles in 2005 is consistent with what was known about SP efficacy in this area during 2003. A mean treatment failure of 32.4% on the 14 days and 74.3% in the 28 days follow-up was reported before SP was withdrawn in 2003 [1]. Interestingly, SP-sensitive P. falciparum parasites have significantly reemerged after the withdrawal of the drug (Fig 3). Triple Pfdhfr wild alleles (Ser-108/Asn-51/Cys-59) increased from zero to 11.1%, which is in contrast with Ghana [36], a decrease from 13.3% in 2001 to 11.9% in 2003 in the absence of SP was reported. Double Pfdhps wild forms (Ala-437/Lys-540) increased from 13.8% to 30.8% and the most important quintuple Pfdhfr/Pfdhps wild forms increased from zero in 2005 to 10.2% in 2007/08, this finding is in accordance with previous report [23] and shows the reemergence SP sensitive parasites. These finding is also in contradiction with the reported changes in Bahir Dar [26].
The decrease in mutation and subsequent increase wild type parasites might be explained by the reduction of residual drug-resistant parasites, caused by the strong drug pressure imposed before 2004 when SP was the first-line drug, followed by lower fitness of these resistant parasites in the absence of drug pressure. Re-emergence of SP susceptible parasites support the hypothesis that drug-resistant P. falciparum parasite may be at competitive disadvantage when drug pressure is removed in agreement with epidemiological reports from northern Ethiopia [26], Tanzania [27], Southern Mozambique [28] and Peru for SP [37] and Malawi [19], Sudan [39] and Southeast Asia [40,41] for CQ. However, in Ghana, Cambodia and Venezuela SP resistance alleles have remained at a high frequency after the replacement of SP [24,25,36].
Although the change in the frequency of mutation is significant, the resistant alleles are still abundant in the study area which is explained by the following factors: the time difference between the two surveys was only two and half years and after the 2004 discontinuation of SP for the treatment of P. falciparum malaria, availability of AL was limited and 85% of the populations were living in rural areas with restricted access to health care giving rise to a high rate of presumptive treatment with available drugs like SP and CQ [42]. The other important factor is cross-resistance between SP and Trimethoprim/ sulfamethoxazole [43,44]. The uses of Trimethoprim/ sulfamethoxazole as prophylaxis against human immunodeficiency virus (HIV) associated opportunistic infections most likely make an essential contribution to SP resistance. In this study, we do not have the data for the HIV prevalence or positivity of the study populations and we cannot sort out the impact of trimethoprim/sulfamethoxazole in maintaining the Pfdhfr and Pfdhps mutation. Cross-resistance between SP and Trimethoprim/sulfamethoxazole appears to be a contributing factor rather than the exclusive factor responsible for less rapid decrease of resistant parasites [43].
The declining trend of mutant alleles and re-emergence of SP sensitive parasites in this perennial transmission area is encouraging despite the limitations of this study. First, this study did not include the change in the frequency of Pfdhps A581G mutation in this study area. However, different studies showed the association of A581G mutation and SP resistance in this part of Africa. Second, the methodologies used in this study detect the mutant and wild type alleles at each locus independent of the other. Sequencing of the full-length Pfdhfr and Pfdhps genes will give a better resolution to determine combinations of mutations. Finally, this study reports the change in frequency of mutation only with two time points, which are 2.5 years apart. Large scale, multi-centered studies with multiple time points will be required to investigate the change in the frequency of mutation and re-emergence of SP sensitive parasites in different transmission areas of the country.
The recovery of SP-sensitive parasite populations in the study area after removal of drug pressure suggests an advantage of the native Pfdhfr and Pfdhps molecule over its mutant forms and points to the possible value of drug rotation strategies in antimalarial policies as we facing the advent of artemisinin resistance. The use of SP for the control and elimination of malaria together with the current tools will require much lower rates of mutation associated with SP resistance. Further multi-centered studies are required to monitor the change in the frequency of mutation, in order to monitor and prevent the establishment of multi drug resistant parasites in different parts of the country.
Supporting Information
(DOCX)
(DOCX)
(XLSX)
Acknowledgments
We are grateful to the participating communities and the staff of Pawe general Hospital for their involvement with this study. This study was supported by a grant from the IAEA (grant Eth/6012), Ethiopian Health and Nutrition Research Institute (EHNRI) and Department of Biology, Addis Ababa University.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
The project was funded by the International atomic energy agency (IAEA), Addis Ababa University and Ethiopian Health and Nutrition Research Institute (EHNRI). The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Jima D, Tesfaye G, Medhin A, Kebede A, Argaw D, Babaniyi O (2005) Efficacy of sulfadoxine-pyrimethamine for the treatment of uncomplicated falciparum malaria in Ethiopia. East Afr Med J 82: 391–395. [DOI] [PubMed] [Google Scholar]
- 2.FMOH (2004) Malaria Diagnosis and Treatment Guidelines for Health Workers in Ethiopia.
- 3. ter Kuile FO, van Eijk AM, Filler SJ (2007) Effect of sulfadoxine-pyrimethamine resistance on the efficacy of intermittent preventive therapy for malaria control during pregnancy: a systematic review. JAMA 297: 2603–2616. [DOI] [PubMed] [Google Scholar]
- 4. Aponte JJ, Schellenberg D, Egan A, Breckenridge A, Carneiro I, Critchley J, et al. (2009) Efficacy and safety of intermittent preventive treatment with sulfadoxine-pyrimethamine for malaria in African infants: a pooled analysis of six randomised, placebo-controlled trials. Lancet 374: 1533–1542. 10.1016/S0140-6736(09)61258-7 [DOI] [PubMed] [Google Scholar]
- 5. Dicko A, Diallo AI, Tembine I, Dicko Y, Dara N, Sidibe Y, et al. (2011) Intermittent preventive treatment of malaria provides substantial protection against malaria in children already protected by an insecticide-treated bednet in Mali: a randomised, double-blind, placebo-controlled trial. PLoS Med 8: e1000407 10.1371/journal.pmed.1000407 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. van Eijk AM, Hill J, Alegana VA, Kirui V, Gething PW, ter Kulie FO, et al. (2011) Coverage of malaria protection in pregnant women in sub-Saharan Africa: a synthesis and analysis of national survey data. Lancet Infect Dis 11: 190–207. 10.1016/S1473-3099(10)70295-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Wang P, Lee CS, Bayoumi R, Djimde A, Doumbo O, Swedberg G, et al. (1997) Resistance to antifolates in Plasmodium falciparum monitored by sequence analysis of dihydropteroate synthetase and dihydrofolate reductase alleles in a large number of field samples of diverse origins. Mol Biochem Parasitol 89: 161–177. [DOI] [PubMed] [Google Scholar]
- 8. Triglia T, Menting JGT, Wilson C, Cowman AF (1997) Mutations in dihydropteroate synthase are responsible for sulfone and sulfonamide resistance in Plasmodium falciparum. P Natl Acad Sci USA 94: 13944–13949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Cowman AF, Morry MJ, Biggs BA, Cross GAM, Foote SJ (1988) Amino-Acid Changes Linked to Pyrimethamine Resistance in the Dihydrofolate-Reductase Thymidylate Synthase Gene of Plasmodium-falciparum. P Natl Acad Sci USA 85: 9109–9113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Brooks DR, Wang P, Read M, Watkins WM, Sims PF, Hyde JE (1994) Sequence variation of the hydroxymethyldihydropterin pyrophosphokinase: dihydropteroate synthase gene in lines of the human malaria parasite, Plasmodium falciparum, with differing resistance to sulfadoxine. Eur J Biochem 224: 397–405. [DOI] [PubMed] [Google Scholar]
- 11. Nzila AM, Nduati E, Mberu EK, Sibley CH, Monks SA, Winstanley PA, et al. (2000) Molecular evidence of greater selective pressure for drug resistance exerted by the long-acting antifolate Pyrimethamine/Sulfadoxine compared with the shorter-acting chlorproguanil/dapsone on Kenyan Plasmodium falciparum. J Infect Dis 181: 2023–2028. [DOI] [PubMed] [Google Scholar]
- 12. Nzila AM, Mberu EK, Sulo J, Dayo H, Winstanley PA, Sibley CH, et al. (2000) Towards an understanding of the mechanism of pyrimethamine-sulfadoxine resistance in Plasmodium falciparum: genotyping of dihydrofolate reductase and dihydropteroate synthase of Kenyan parasites. Antimicrob Agents Chemother 44: 991–996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Kublin JG, Dzinjalamala FK, Kamwendo DD, Malkin EM, Cortese JF, Martino LM, et al. (2002) Molecular markers for failure of sulfadoxine-pyrimethamine and chlorproguanil-dapsone treatment of Plasmodium falciparum malaria. J Infect Dis 185: 380–388. [DOI] [PubMed] [Google Scholar]
- 14. Nzila A, Ochong E, Nduati E, Gilbert K, Winstanley P, Ward S, et al. (2005) Why has the dihydrofolate reductase 164 mutation not consistently been found in Africa yet? Trans R Soc Trop Med Hyg 99: 341–346. [DOI] [PubMed] [Google Scholar]
- 15. Lynch C, Pearce R, Pota H, Cox J, Abeku TA, Rwakimari J, et al. (2008) Emergence of a dhfr mutation conferring high-level drug resistance in Plasmodium falciparum populations from southwest Uganda. J Infect Dis 197: 1598–1604. 10.1086/587845 [DOI] [PubMed] [Google Scholar]
- 16. Alifrangis M, Lusingu JP, Mmbando B, Dalgaard MB, Vestergaard LS, Ishengoma D, et al. (2009) Five-year surveillance of molecular markers of Plasmodium falciparum antimalarial drug resistance in Korogwe District, Tanzania: accumulation of the 581G mutation in the P. falciparum dihydropteroate synthase gene. Am J Trop Med Hyg 80: 523–527. [PubMed] [Google Scholar]
- 17. Karema C, Imwong M, Fanello CI, Stepniewska K, Uwimana A, Nakeesathit S, et al. (2010) Molecular correlates of high-level antifolate resistance in Rwandan children with Plasmodium falciparum malaria. Antimicrob Agents Chemother 54: 477–483. 10.1128/AAC.00498-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.WHO (2010) WHO Policy recommendation on Intermittent Preventive Treatment during infancy with sulphadoxine-pyrimethamine (SP-IPTi) for Plasmodium falciparum malaria control in Africa. Geneva
- 19. Kublin JG, Cortese JF, Njuniu EM, Mukadam RAG, Wirima JJ, Kazembe PN, et al. (2003) Reemergence of chloroquine-sensitive Plasmodium falciparum malaria after cessation of chloroquine use in Malawi. J Infect Dis 187: 1870–1875. [DOI] [PubMed] [Google Scholar]
- 20. Mohammed A, Ndaro A, Kalinga A, Manjurano A, Mosha JF, Mosha DF, et al. (2013) Trends in chloroquine resistance marker, Pfcrt-K76T mutation ten years after chloroquine withdrawal in Tanzania. Malar J 12: 415 10.1186/1475-2875-12-415 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Mwai L, Ochong E, Abdirahman A, Kiara SM, Ward S, Kokwaro G, et al. (2009) Chloroquine resistance before and after its withdrawal in Kenya. Malaria J 8:106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Wang X, Mu J, Li G, Chen P, Guo X, Fu L, et al. (2005) Decreased prevalence of the Plasmodium falciparum chloroquine resistance transporter 76T marker associated with cessation of chloroquine use against P. falciparum malaria in Hainan, People's Republic of China. Am J Trop Med Hyg 72: 410–414. [PubMed] [Google Scholar]
- 23. Laufer MK, Thesing PC, Eddington ND, Masonga R, Dzinjalamala FK, Takala SL, et al. (2006) Return of chloroquine antimalarial efficacy in Malawi. N Engl J Med 355: 1959–1966. [DOI] [PubMed] [Google Scholar]
- 24. Khim N, Bouchier C, Ekala MT, Incardona S, Lim P, Legrand E, et al. (2005) Countrywide survey shows very high prevalence of Plasmodium falciparum multilocus resistance genotypes in Cambodia. Antimicrob Agents Chemother 49: 3147–3152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. McCollum AM, Mueller K, Villegas L, Udhayakumar V, Escalante AA (2007) Common origin and fixation of Plasmodium falciparum dhfr and dhps mutations associated with sulfadoxine-pyrimethamine resistance in a low-transmission area in South America. Antimicrob Agents Chemother 51: 2085–2091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Hailemeskel E, Kassa M, Taddesse G, Mohammed H, Woyessa A, Sleshi M, et al. (2013) Prevalence of sulfadoxine-pyrimethamine resistance-associated mutations in dhfr and dhps genes of Plasmodium falciparum three years after SP withdrawal in Bahir Dar, Northwest Ethiopia. Acta Trop 128: 636–641. [DOI] [PubMed] [Google Scholar]
- 27. Pearce RJ, Ord R, Kaur H, Lupala C, Schellenberg J, Shirma K, et al. (2013) A community-randomized evaluation of the effect of intermittent preventive treatment in infants on antimalarial drug resistance in southern Tanzania. J Infect Dis 207: 848–859. 10.1093/infdis/jis742 [DOI] [PubMed] [Google Scholar]
- 28. Raman J, Sharp B, Kleinschmidt I, Roper C, Streat E, Kelly V, et al. (2008) Differential effect of regional drug pressure on dihydrofolate reductase and dihydropteroate synthetase mutations in southern Mozambique. Am J Trop Med Hyg 78: 256–261. [PMC free article] [PubMed] [Google Scholar]
- 29. Schunk M, Kumma WP, Miranda IB, Osman ME, Roewer S, Alano A, et al. (2006) High prevalence of drug-resistance mutations in Plasmodium falciparum and Plasmodium vivax in southern Ethiopia. Malaria J 5:54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Gebru-Woldearegai T, Hailu A, Grobusch MP, Kun JFJ (2005) Molecular surveillance of mutations in dihydrofolate reductase and dihyropteroate synthase genes of Plasmodium falciparum in Ethiopia. Am J Trop Med Hyg 73: 1131–1134. [PubMed] [Google Scholar]
- 31. Vacas E, Fernandez-Martinez AB, Bajo AM, Sanchez-Chapado M, Schally AV, Prieto JC, et al. (2012) Vasoactive intestinal peptide (VIP) inhibits human renal cell carcinoma proliferation. Biochim Biophys Acta 1823: 1676–1685. [DOI] [PubMed] [Google Scholar]
- 32. WHO (2000) Severe falciparum malaria. World Health Organization, Communicable Diseases Cluster. Trans R Soc Trop Med Hyg 94 Suppl 1: S1–90. [PubMed] [Google Scholar]
- 33. Plowe CV, Djimde A, Bouare M, Doumbo O, Wellems TE (1995) Pyrimethamine and Proguanil Resistance-Conferring Mutations in Plasmodium-Falciparum Dihydrofolate-Reductase—Polymerase Chain-Reaction Methods for Surveillance in Africa. Am J Trop Med Hyg 52: 565–568. [DOI] [PubMed] [Google Scholar]
- 34. Abdel-Muhsin AM, Ranford-Cartwright LC, Medani AR, Ahmed S, Suleiman S, Khan B, et al. (2002) Detection of mutations in the Plasmodium falciparum dihydrofolate reductase (dhfr) gene by dot-blot hybridization. Am J Trop Med Hyg 67: 24–27. [DOI] [PubMed] [Google Scholar]
- 35. Roper C, Pearce R, Bredenkamp B (2003) Antifolate antimalarial resistance in southeast Africa: a population-based analysis (vol 361, pg 1174, 2003). Lancet 362: 922–922. [DOI] [PubMed] [Google Scholar]
- 36. Marks F, Evans J, Meyer CG, Browne EN, Flessner C, von Kalckreuth V, et al. (2005) High prevalence of markers for sulfadoxine and pyrimethamine resistance in Plasmodium falciparum in the absence of drug pressure in the Ashanti region of Ghana. Antimicrob Agents Chemother 49: 1101–1105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Zhou ZY, Griffing SM, de Oliveira AM, McCollum AM, Quezada WM, Arrospide N, et al. (2008) Decline in sulfadoxine-pyrimethamine-resistant Alleles after change in drug policy in the Amazon region of Peru. Antimicrob Agents Chemother 52: 1900–1900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Wongsrichanalai C, Pickard AL, Wernsdorfer WH, Meshnick SR (2002) Epidemiology of drug-resistant malaria. Lancet Infect Dis 2: 209–218. [DOI] [PubMed] [Google Scholar]
- 39. Abdel-Muhsin AM, Mackinnon MJ, Ali E, Nassir el KA, Suleiman S, Ahmed S, et al. (2004) Evolution of drug-resistance genes in Plasmodium falciparum in an area of seasonal malaria transmission in Eastern Sudan. J Infect Dis 189: 1239–1244. [DOI] [PubMed] [Google Scholar]
- 40. Liu DQ, Liu RJ, Ren DX, Gao DQ, Zhang CY, Qui CP, et al. (1995) Changes in the resistance of Plasmodium falciparum to chloroquine in Hainan, China. Bull World Health Organ 73: 483–486. [PMC free article] [PubMed] [Google Scholar]
- 41. Thaithong S, Suebsaeng L, Rooney W, Beale GH (1988) Evidence of Increased Chloroquine Sensitivity in Thai Isolates of Plasmodium falciparum. Trans R Soc Trop Med Hyg 82: 37–38. [DOI] [PubMed] [Google Scholar]
- 42. Deressa W, Ali A, Enqusellassie F (2003) Self-treatment of malaria in rural communities, Butajira, southern Ethiopia. Bull World Health Organ 81: 261–268. [PMC free article] [PubMed] [Google Scholar]
- 43. Iyer JK, Milhous WK, Cortese JF, Kublin JG, Plowe CV (2001) Plasmodium falciparum cross-resistance between trimethoprim and pyrimethamine. Lancet 358: 1066–1067. [DOI] [PubMed] [Google Scholar]
- 44. Khalil I, Ronn AM, Alifrangis M, Gabar HA, Satti GM, Bygbjerg IC (2003) Dihydrofolate reductase and dihydropteroate synthase genotypes associated with in vitro resistance of Plasmodium falciparum to pyrimethamine, trimethoprim, sulfadoxine, and sulfamethoxazole. Am J Trop Med Hyg 68: 586–589. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
(DOCX)
(XLSX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.