

Abstract
Floral thermogenesis plays a crucial role in pollination biology, especially in plant–pollinator interactions. We have recently explored how thermogenesis is related to pollinator activity and odour release in Magnolia sprengeri. By analyzing flower temperatures, emission of volatiles, and insect visitation, we found that floral blends released during pistillate and staminate stages were similar and coincided with sap beetle visitation. Thus, odour mimicry of staminate-stage flowers may occur during the pistillate stage and may be an adaptive strategy of Magnolia species to attract pollinators during both stages, ensuring successful pollination. In addition to the biological significance of floral thermogenesis in Magnolia species, we explored the underlying regulatory mechanisms via profiling miRNA expression in M. denudata flowers during thermogenic and non-thermogenic stages. We identified 17 miRNAs that may play regulatory roles in floral thermogenesis. Functional annotation of their target genes indicated that these miRNAs regulate floral thermogenesis by influencing cellular respiration and light reactions. These findings increase our understanding of plant–pollinator interactions and the regulatory mechanisms in thermogenic plants.
Keywords: Magnoliaceae, odour mimicry, pollination biology, regulatory mechanisms, thermogenesis
Thermogenic plants produce an unusually large amount of heat during the floral stages,1 which has been reported in flowers of several families of seed plants.2-8 Thermogenesis is considered a direct energy reward for the insect visitors that usually reside in a floral chamber.9 Thermogenesis is also associated with volatilisation of floral scents, which attract pollinators during the anthesis.10-12 Recently, we studied the relationship between thermogenesis and pollinator activity during flowering of M. sprengeri by investigating flower temperatures, emission of volatiles, and insect visitation. It was found that release of floral blends and insect visitation were closely associated with heat production. In addition, we found that floral odours during pistillate and staminate stages were fundamentally similar, indicative of odour mimicry of staminate-stage flowers during the pistillate stage. Based on these results, floral thermogenesis plays a role in pollination biology of M. sprengeri by promoting emission of floral scents, which act as a signal of food rewards for pollinators.13
Flowers of thermogenic plants have the ability to maintain a relatively higher temperature than ambient environments during anthesis.14-16 In this study, although the floral temperature of M. sprengeri changed constantly throughout anthesis, it was relatively higher than ambient air temperature, and heat production showed 2 noticeable peaks; one at the pistillate stage and the other at the staminate stage. The thermogenic pattern of M. sprengeri was identical to that of its close relative, M. denudata—both flower during early spring and produce heat in the daytime.13 The thermogenic peaks of these species are well-synchronised with the activity of their pollinators. It is interesting to note that pollinators of M. ovata (which produces heat at night) are active at night.16 These studies reveal an association between floral thermogenesis and pollinator visitation.
Thermogenesis is closely related to the volatilisation of floral odour in M. sprengeri. Since floral odour and heat production are dynamic processes, the relative proportions of the odour compounds varied across anthesis. We found that the floral blends released during pistillate and staminate stages were very similar in M. sprengeri, indicative of odour mimicry of staminate-stage flowers in pistillate-stage flowers. Previously, it was suspected that staminate-stage flowers mimic pistillate-stage flowers in M. hypoleuca, as the former offers no pollen reward but has strong fragrance.17 Our results confirm odour mimicry between the female and male stage flowers in a Magnolia species. In another study, a similarity in the smells of pistillate- and staminate-stage flowers of M. ovata was reported.18 These results reveal that flowers of Magnolia species at both pistillate and staminate stages attract pollinators through odour mimicry. Considering that odour emission coincided with the 2 thermogenic episodes, it is possible that thermogenesis promotes odour release, which attracts pollinators at both the pistillate and staminate stages, ensuring reproductive success at low temperatures during the early spring.
Although the ecological significance of thermogenesis is widely recognized,19-21 little information on the underlying regulatory mechanisms is available.20 miRNAs are crucial regulators of various biological processes in eukaryotic cells,22,23 but their roles in floral thermogenesis remain unclear. We recently profiled miRNA expression in M. denudata flowers during thermogenesis using high-throughput sequencing.21 A total of 82 conserved and 32 novel miRNAs were identified in M. denudata flowers, among which 17 were differentially expressed between thermogenic and non-thermogenic stages, and thus were thought to play roles in regulating floral thermogenesis. Gene Ontology (GO) enrichment analysis revealed that target genes of these thermogenesis-related miRNAs were enriched in the functional groups of ‘polyprenyl transferase activity’ and ‘photosynthetic electron transport’. Thus, we propose that regulation of floral thermogenesis may be associated with cellular respiration and photosynthesis in M. denudata.
Two types of thermogenesis have been identified among thermogenic plants. In some species, floral temperature is maintained within a constant range, independently of ambient temperature, throughout anthesis (Fig. 1A). These are thermoregulatory species, such as Philodendron selloum,24 Nelumbo nucifera,25 and Symplocarpus foetidus.26 In other thermogenic species, heat production usually corresponds to the period when female flower parts are most receptive to pollination and when floral scents are strongest (Fig. 1B), such as in Dracunculus vulgaris, Helicodiceros muscivorus,24,27 and Magnolia species.28 Such thermogenic plants are sometimes referred to as pseudo-thermoregulatory species. Thermogenesis directly rewards pollinators (beetles) with energy, especially at night when no floral scents are released.19,29 Compared with thermoregulatory flowers, which produce heat during anthesis, pseudo-thermoregulatory species use less energy, as their heat production is tied to the circadian cycle. They experience intense warming only during peaks of scent emission, promoting odour release to attract visiting insects. The pseudo-thermoregulatory strategy for attracting pollinators may be more efficient than the thermoregulatory strategy in having more benefits and fewer costs. The divergence of the 2 strategies may involve co-evolution with pollinator insects. Further comparative studies of the 2 types of thermogenesis may facilitate our understanding of evolution and of regulatory mechanisms driving floral thermogenesis.
Figure 1.
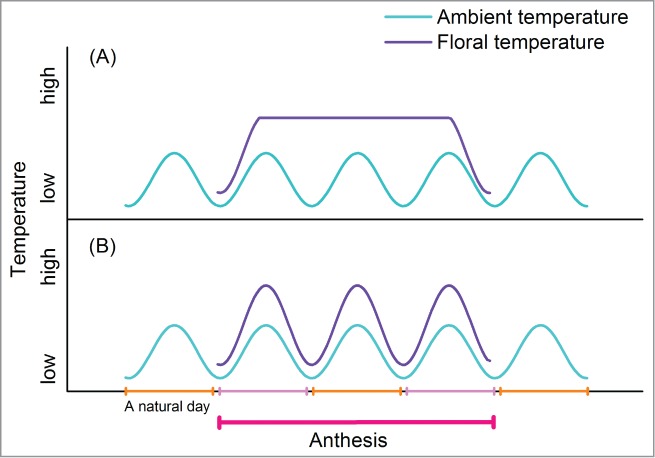
Different patterns of heat production in thermogenic flowers. (A) In some thermogenic flowers, heat production is physiologically regulated to maintain a relatively constant floral temperature, independently of ambient temperature, such as in skunk cabbage (Symplocarpus foetidus).26 (B) Thermogenesis is related to the circadian cycle and greatly increases when scent is released in most thermogenic flowers, including Magnolia spp.28
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed
Funding
This work was supported by the Fundamental Research Funds for Central Universities (No. YX 2014–14) and National Science Fund of China (No. 31100450 & J1103516).
References
- 1. Seymour RS, Gibernau M. Respiration of thermogenic inflorescences of Philodendron melinonii: natural pattern and responses to experimental temperatures. J Exp Bot 2008; 59:1353-62; PMID:18375932; http://dx.doi.org/ 10.1093/jxb/ern042 [DOI] [PubMed] [Google Scholar]
- 2. Endress PK. Diversity and evolutionary biology of tropical flowers. Cambridge, England; New York, NY, USA: Cambridge University Press; 1994. [Google Scholar]
- 3. Küechmeister H, Webber AC, Silberbauer-Gottsberger I, Gottsberger G. Pollination and its relationship to thermogenesis in species of Arecaceae and Annonaceae of central Amazonia. Acta Amazonica 1998; 28:217-45 http://www.ncbi.nlm.nih.gov/pubmed/NOT_FOUND [Google Scholar]
- 4. Ervik F, Barfod A. Thermogenesis in palm inflorescences and its ecological significance. Acta Biol Venez 1999; 22:195-212 http://www.ncbi.nlm.nih.gov/pubmed/NOT_FOUND [Google Scholar]
- 5. Thien LB, Azuma H, Kawano S. New perspectives on the pollination biology of basal angiosperms. Int J Plant Sci 2000; 161:225-35http://www.ncbi.nlm.nih.gov/pubmed/NOT_FOUND; http://dx.doi.org/ 10.1086/317575 [DOI] [Google Scholar]
- 6. Seymour RS, Maass E, Bolin JF. Floral thermogenesis of three species of Hydnora (Hydnoraceae) in Africa. Ann Bot 2009; 104:823-32; PMID:19584128; http://dx.doi.org/ 10.1093/aob/mcp168 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Takacs S, Bottomley H, Andreller I, Zaradnik T, Schwarz J, Bennett R, Strong W, Gries G. Infrared radiation from hot cones on cool conifers attracts seed-feeding insects. Proc Biol Sci 2009; 276:649-55; PMID:18945664; http://dx.doi.org/ 10.1098/rspb.2008.0742 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Roemer RB, Terry LI, Marler TE. Cone thermogenesis and its limits in the tropical Cycas micronesica (Cycadaceae): association with cone growth, dehiscence, and post-dehiscence phases. Am J Bot 2013; 100:1981-90; PMID:2408114; http://dx.doi.org/ 10.3732/ajb.1300047 [DOI] [PubMed] [Google Scholar]
- 9. Seymour RS, Matthews PG. The role of thermogenesis in the pollination biology of the Amazon waterlily Victoria amazonica. Ann Bot 2006; 98:1129-35; PMID:17018568; http://dx.doi.org/ 10.1093/aob/mcl201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. McCallum KP, McDougall FO, Seymour RS. A review of the energetics of pollination biology. J Comp Physiol B 2013; 183:867-76; PMID:23653068; http://dx.doi.org/ 10.1007/s00360-013-0760-5 [DOI] [PubMed] [Google Scholar]
- 11. Suinyuy TN, Donaldson JS, Johnson SD. Patterns of odour emission, thermogenesis and pollinator activity in cones of an African cycad: what mechanisms apply? Ann Bot 2013; 112:891-902; PMID:23887092; http://dx.doi.org/ 10.1093/aob/mct159 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Dieringer G, Leticia Cabrera R, Mottaleb M. Ecological relationship between floral thermogenesis and pollination in Nelumbo lutea (Nelumbonaceae). Am J Bot 2014; 101:357-64; PMID:24458119; http://dx.doi.org/ 10.3732/ajb.1300370 [DOI] [PubMed] [Google Scholar]
- 13. Wang RH, Liu XY, Mou SL, Xu S, Zhang ZX. Temperature regulation of floral buds and floral thermogenicity in Magnolia denudata (Magnoliaceae). Trees 2013; 27:1755-62 http://www.ncbi.nlm.nih.gov/pubmed/NOT_FOUND; http://dx.doi.org/ 10.1007/s00468-013-0921-x [DOI] [Google Scholar]
- 14. Li JK, Huang SQ. Flower thermoregulation facilitates fertilization in Asian sacred lotus. Ann Bot 2009; 103:1159-63; PMID:19282320; http://dx.doi.org/ 10.1093/aob/mcp051 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Seymour RS, Lindshau G, Ito K. Thermal clamping of temperature-regulating flowers reveals the precision and limits of the biochemical regulatory mechanism. Planta 2010; 231:1291-300; PMID:20221632; http://dx.doi.org/ 10.1007/s00425-010-1128-7 [DOI] [PubMed] [Google Scholar]
- 16. Seymour RS, Silberbauer-Gottsberger I, Gottsberger G. Respiration and temperature patterns in thermogenic flowers of Magnolia ovata under natural conditions in Brazil. Funct Plant Biol 2010; 37:870-8; http://dx.doi.org/ 10.1071/FP10039 [DOI] [Google Scholar]
- 17. Kikuzawa K, Mizui N. Flowering and fruiting phenology of Magnolia hypoleuca. Plant Spec Biol 1990; 5:255-61; http://dx.doi.org/ 10.1111/j.1442-1984.1990.tb00185.x [DOI] [Google Scholar]
- 18. Gottsberger G, Silberbauer-Gottsberger I, Seymour RS, Dotterl S. Pollination ecology of Magnolia ovata may explain the overall large flower size of the genus. Flora 2012; 207:107-18; http://dx.doi.org/ 10.1016/j.flora.2011.11.003 [DOI] [Google Scholar]
- 19. Seymour RS, White CR, Gibernau M. Environmental biology: heat reward for insect pollinators. Nature 2003; 426:243-4; PMID:14628037; http://dx.doi.org/ 10.1038/426243a [DOI] [PubMed] [Google Scholar]
- 20. Ito-Inaba Y, Hida Y, Matsumura H, Masuko H, Yazu F, Terauchi R, Watanabe M, Inaba T. The gene expression landscape of thermogenic skunk cabbage suggests critical roles for mitochondrial and vacuolar metabolic pathways in the regulation of thermogenesis. Plant Cell Environ 2012; 35:554-66; PMID:21955303; http://dx.doi.org/ 10.1111/j.1365-3040.2011.02435.x [DOI] [PubMed] [Google Scholar]
- 21. Liu XY, Cao DC, Ji XY, Zhang ZX, Wang YW, Wang RH. miRNAs play essential roles in the floral thermogenesis of Magnolia denudata (Magnoliaceae). Trees 2014; 1-8; http://dx.doi.org/ 10.1007/s00468-014-1051-9 [DOI] [Google Scholar]
- 22. Romanel E, Silva TF, Correa RL, Farinelli L, Hawkins JS, Schrago CE, Vaslin MF. Global alteration of microRNAs and transposon-derived small RNAs in cotton (Gossypium hirsutum) during Cotton leafroll dwarf polerovirus (CLRDV) infection. Plant Mol Biol 2012; 80:443-60; PMID:22987114; http://dx.doi.org/ 10.1007/s11103-012-9959-1 [DOI] [PubMed] [Google Scholar]
- 23. Wang ZJ, Huang JQ, Huang YJ, Li Z, Zheng BS. Discovery and profiling of novel and conserved microRNAs during flower development in Carya cathayensis via deep sequencing. Planta 2012; 236:613-21; PMID:22481137; http://dx.doi.org/ 10.1007/s00425-012-1634-x [DOI] [PubMed] [Google Scholar]
- 24. Seymour RS, Schultze-Motel P. Respiration, temperature regulation and energetics of thermogenic inflorescences of the dragon lily Dracunculus vulgaris (Araceae). Proc Biol Sci 1999; 266:1975-83; http://dx.doi.org/ 10.1098/rspb.1999.0875 [DOI] [Google Scholar]
- 25. Seymour RS, Schultze-Motel P, Lamprecht I. Heat production by sacred lotus flowers depends on ambient temperature, not light cycle. J Exp Bot 1998; 49, 1213-17; http://dx.doi.org/ 10.1093/jxb/49.324.1213 [DOI] [Google Scholar]
- 26. Seymour RS. Dynamics and precision of thermoregulatory responses of eastern skunk cabbage Symplocarpus foetidus. Plant Cell Environ 2004; 27, 1014-22; http://dx.doi.org/ 10.1111/j.1365-3040.2004.01206.x [DOI] [Google Scholar]
- 27. Seymour RS, Gibernau M, Ito K. Thermogenesis and respiration of inflorescences of the dead horse arum Helicodiceros muscivorus, a pseudo-thermoregulatory aroid associated with fly pollination. Funct Ecol 2003; 17, 886-94; http://dx.doi.org/ 10.1111/j.1365-2435.2003.00802.x [DOI] [Google Scholar]
- 28. Wang RH, Xu S, Liu XY, Zhang YY, Wang JZ, Zhang ZX. Thermogenesis, flowering and the association with variation in floral odour attractants in Magnolia sprengeri (Magnoliaceae). PLoS One 2012; 9(6): e99356; http://dx.doi.org/ 10.1371/journal.pone.0099356 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Seymour RS, Schultze-Motel P. Heat-producing flowers. Endeavour 1997; 21, 125-9; http://dx.doi.org/ 10.1016/S0160-9327(97)80222-0 [DOI] [Google Scholar]