

Abbreviations
- AP
action potential
- EEG
electroencephalograph
- EPD
electrical potential difference
- MAP
microtubule-associated protein
- MT
microtubule
- Orch OR
orchestrated objective reduction
Whether or not plants exhibit consciousness in the sense that they possess a ‘sensation of self’ is uncertain. Unlike humans and certain animals, plants cannot be interrogated directly on this matter. In any case, it is possible that states of consciousness, like brains and nervous systems of animals, have evolved and developed in different ways, depending on their phylogeny. However, some of the criteria by which consciousness is inferred to be present in animals are met by plants. For example, plants display features of cognition (sensing and response) and of learning and memory which, in animals, contribute to the conscious state. Plants also possess a rudimentary nervous system similar to that found in basal animals and have, perhaps, a simple brain, as well as showing slow and fast transmissable electrical activity, all of which are strong correlates of consciousness in animals. However, the question of consciousness in general can be approached in a different way. This is by taking the Orch OR (Orchestrated Objective Reduction) hypothesis of Hameroff and Penrose1 as a starting point. The Orch OR hypothesis, which is based in quantum physics, proposes that, when a sufficient mass of tubulin molecules has assembled into cytoskeletal microtubules (MTs) within neuronal cells of the brain, these structures become sites of quantum computation and of quantum state reduction (OR) events resulting in moments of protoconsciousness. Because plant cells also have large populations of MTs, and because plant MTs share properties with those of animal neuronal MTs, which putatively orchestrate OR events, plant MTs might also be sites of quantum reduction events and, hence, lead to momentary protoconsciousness. The extent to which the Orch OR hypothesis is applicable to plants is examined, and it is argued that, within the plant body, the most likely tissue where OR events could be located and promote protoconsciousness is to be found in the system of ray cells of tree trunks in which bundled MTs and actin filaments are prevalent. A single complete ray complex is estimated to contain about as many tubulin molecules as a single human cerebral neuron.
Inferential evidence used by Hameroff and Penrose to support their Orch OR hypothesis leads in another direction. These authors presented estimates of the frequency of protoconscious or conscious moments. This frequency turns out to be similar to the frequency of successive Earthly quantal time units, according to a theory proposed by G. Dorda.2 These time units are also intimately linked with simultaneous changes of quantal mass, both mass and time being structured according to the motions of Earth and Moon around the Sun. In the case of cellular tissues, quantised mass takes the form of aggregates of water, and these are hypothesized to move in and out of cells in response to the passage of quantal time. In humans, the passage of time and its association with quantised water flux might account for a sense of self gained during meditative practices and govern also sleep-wakefulness rhythms, a principle which could apply to all living organisms. In plants, a similar quantised mass-time relationship might also confer an experience akin to one of selfhood. This relationship may be supplemented with the development of spontaneous action potentials which, in turn, lead to the development of electromagnetic fields. The presence of such fields surrounding plants and other living bodies are perhaps related to instances of ‘primary perception’ and also to certain cognitive features, such as kin recognition, which seem to lie at the fringe of consciousness.
Introduction
Despite the efforts of neurobiologists, philosophers and psychologists, the nature of consciousness remains elusive. The usual model taken for consideration is human consciousness, the major theories of which – from Descartes in the 17th century to quantum mechanics in the 20th century, and the manner in which each of these theories fits within the prevailing scientific view of the world – have been discussed by Seager.3 Not only is human consciousness the most advanced of its kind but it is, of all presumed types of consciousnesses, also the one most available to empirical enquiry.4,5 As Rosenberg (ref. 6, p. 93) suggests, “every theory of consciousness goes beyond the direct evidence we have, because we have direct evidence only in our own case.” Sometimes, there are also opportunities for experimental modulation and the observation of altered states of consciousness,7-9 which throw the usual type of consciousness into sharper relief. The focus on human consciousness and its subjective nature, however, need not deny that other, non-human organisms might also experience consciousness. Nevertheless, pursuit of other models of consciousness presents the difficulty that the corresponding organisms may have sense organs of different acuity, or even of different kind, to those of humans;10 it is, after all, through the senses that the concept of consciousness has come into being, and which provide the impetus for consciousness studies. Indeed, a systematic enumeration of criteria whereby the presence of consciousness can be inferred in human and non-human animals has been presented by Seth and his colleagues.11-13 These criteria can therefore be useful in determining whether consciousness is widespread, at least in the animal kingdom. However, notwithstanding the immensely long phylogenetic progression within which consciousness lies,14 not all of these criteria are met in every animal group. Moreover, it is possible that consciousness, in some form or other, has arisen more than once during the course of evolution.12 Nevertheless, it should be possible to reach some consensus on what qualities or properties comprise a ‘universal’ mode of consciousness. For Maxine Sheets-Johnstone,15 from whose work the first part of the title of the present article is derived, the key question relating to consciousness is not how consciousness arose in matter, but how consciousness comes “to be in the natural history of living creatures and to inhere in the animate?” This is the object of the present article, especially in regard to how this question of the inherency of consciousness relates to plants.
Recently, a novel and apparently universal theory of consciousness, which brings a provisional, though theoretical answer to the question of Sheets-Johnstone, was proposed in its latest form by Stuart Hameroff and Sir Roger Penrose (ref. 1 – this publication is hereafter sometimes referred to simply as ‘Hameroff and Penrose’). The theory deals with the preconditions for a moment of consciousness. It places quantum-physical processes center-stage and deploys the argument that cytoskeletal microtubules (MTs), particularly those contained within neuronal cells, are sites where crucial ‘protoconscious’ events are initiated. The consequences of these events are then relayed to the brain where they are perceived as moments of ‘protoconscious experience’. Because MTs are widespread and abundant in all forms of life except bacteria (although bacteria do possess helices of the tubulin-related protein FtsZ16), it follows that the Hameroff-Penrose hypothesis could be considered in relation to organisms other than Man – that is, wherever intracellular tubulin and MTs are found. Hence, it should be possible to discover whether the elements that uphold the theory and which make possible a protoconscious event, are present in the biological makeup of non-human organisms and, hence, confer upon these organisms similar moments of proto-conscious experience. It should also be remarked that, although this is not the only theory to have placed consciousness and its neurobiological correlates within the domain of quantum physics – see ref. 17 for a survey, and note also the proposals of Walker18 and Bernroider and Summhammer19 for quantum effects in relation to neuronal functioning – the Hameroff-Penrose hypothesis is the one which is currently attracting a great deal of attention due not only to its testable proposals, but also to its theoretical underpinnings, which themselves are tending to lead toward a deepening consideration of gravity within the context of quantum mechanics in general.20,21 Although Hameroff and Penrose1 specify the conditions which, according to their quantum-physical theory, initiate what they term a ‘protoconscious event’, these authors have left it to others to describe what actually constitutes the broader aspects of consciousness; they do say, however, that consciousness ‘implies a sense of self’ and ‘defines our existence’. It is here that Damasio,5 for example, feels that the issue which is critical in any comprehensive account of consciousness lies in explaining “how we know that we own a mind;” and for this he does not regard quantum events as necessary, although he does hold that such events offer a possible means of explaining “how we have a mind.”
Hameroff and Penrose1 have also put forward evidence from observation of meditative states as being relevant to an understanding of the conscious state. The advantage of this less conventional, but nevertheless supportive viewpoint is that consciousness can hereby be considered not only from a top-down (holistic) perspective, starting from the correlates of higher states of consciousness, but can also be linked with the more everyday, but nevertheless fundamental observations of this state. This means that the question of consciousness need not rely entirely on the bottom-up (reductionist) perspective, centered on sensory stimuli and response mechanisms, such as espoused by Crick and Koch,22 but can also espouse more spiritual aspects of the question. Taking both viewpoints together, as was proposed by Sperry,23 suggests that not only could a fairly complete tiered structure of consciousness be prepared but mechanisms could also be uncovered by which this tiered structure comes about. Then would follow the possibility of examining the degree to which different non-human organisms, including plants, have developed the potentiality, and perhaps the capacity, for consciousness, a capacity associated, fundamentally, with sensitivity to the process of living within a particular environment, guided either by instinct or environment. However, as soon as the question of non-human and plant types of consciousness is mooted, investigators become cautious. Crick and Koch,22 for example, claimed that the time was not right for such an enquiry, and Sheets-Johnstone,15 while at pains not to trivialise “the ways in which plants are animate,” preferred to “narrow down the complexity of an already complex subject” and, hence, her discussion remained within the scope of animal models.
It is some years since the topic of consciousness of plants was discussed critically and in depth,24 even though a number of authors have not been shy of writing prolifically about subjects such as of ‘plant intelligence’,25,26 ‘plant decision-making’,27 and ‘plant cognition’,28,29 all of which are relevant for defining certain areas related to consciousness, but none of which, individually, are evidence for consciousness. Because of the difficulties of language and idiom, the 3 mentioned consciousness-related subjects are bedevilled by pitfalls of metaphor and anthropomorphism which can lead to unsatisfying subjective conclusions.30 Nevertheless, some metaphors of consciousness and attention can lead to testable hypotheses.31 Alexandra Nagel,24 although anxious to explore a possible conscious aspect of plant life, did not define consciousness, either in general terms or in terms of what it might entail for the plant. The examples she chose were rather of a plant being ‘conscious’ of its environment, i.e., the plant's response to many types of unusual stimuli – even ‘psychic’ stimuli – and, hence, she tended to treat ‘consciousness’ as the ability to perceive inputs to which a response could be observed. The position outlined in the present paper is somewhat different: it is proposed that, in essence, consciousness is a sense of self, perceived during a state of awareness. In its purest form, this sensation of self may be received in circumstances free from the implications of learning, memory and thought, as well as emotional, intellectual and locomotory reactions to stimuli. As this indicates, consciousness would then seem to be something more than what reductionists would claim it to be: that it is simply a matter of how sensory signals (e.g., visual signals) are perceived and interpreted by the brain;22 it may be more akin to a special kind of ‘feeling’ (“I know that I know”) which accompanies the emerging sense of self.4,5
The topic of consciousness of plants is therefore broached in the light of the Hameroff-Penrose theory, drawing, as did these authors,1 upon 2500 years of Buddhist thinking on this subject, especially the Buddhist theory of momentariness.32 Also recognized are the perceptive insights of Henri Bergson, who is now regarded as one of the first so-called ‘process philosophers’ (prior to AN Whitehead, who posited consciousness to be ‘moments of experience’), and whose ideas on the natural history of consciousness, authored 100 years ago, chime with many of the newer ideas emerging from modern studies of evolution, behavior, intelligence and consciousness.33,34
What is plant consciousness?
Alexandra Nagel,24 when considering the question of plant consciousness, surveyed both scientific and non-scientific literature and analyzed the variously held beliefs of whether plants were or were not conscious. The evidence amassed did not lead to a decisive answer because much depended upon subjective opinions rather than upon hypotheses and experiments.
Definitions of consciousness usually start with the supposition that it is a property of a brain fed, moment by moment, with inputs from the sense organs via a nervous system. In humans, these various momentary sensations are bound together by the brain to produce a feeling of continuous awareness and, at the same time, generate a representation of the organism's experience of its internal and external world.35 The immediate product of such an integrative system can be regarded as a ‘core’ consciousness, which is subjective and private to the organism. The manner in which the various sensory inputs are handled by the brain leads to the conceptualization of a range of functional attributes to which terms such as cognition, memory, feelings, thought, and so on, are applied. In the case of plants, cognition and memory have been critically discussed in the scientific literature28 and are reasonably well accepted as attributes of plants, as are the notions that plants possess both a simple brain and a simple nervous system.36 Cognition is a source of experience; and cognitive experiences may well be assimilated by plants not only to institute immediate responses but also to be stored as a memory. Both circumstances form a basis for primary plant consciousness. However, it is entirely another matter whether a plant is able to bind together the manifold varieties of its experience and thereby form – and here one needs the language of metaphor - a representation of itself and of its world upon the screen of a plant mind (phyto-mind). That is the deep question of plant consciousness.
Seth et al.11 proposed 17 criteria which conformed to the neuronal, brain-based conscious state of humans by which the degree of consciousness in non-human mammals and other animals could then be assessed, using their patterns of behavior as a reference. Humans therefore serve as the bench-mark for discussions of consciousness in general. However, not only are many of the 17 criteria inapplicable to many animal species because of their much simplified behavioral patterns, but many are also inapplicable to plants. Nevertheless, 2 of the criteria, ‘learning’ and ‘decision making’ (nos. 14 and 17 of Seth et al.'s criteria), can be adopted in modified form and analogized to plant processes which indicate corresponding experience of, and response to, the outside world. There may also be traces of conditions that fulfil 4 other criteria (nos. 1, 3, 7 and 12), ‘electroencephalographic (EEG) signature’, ‘widespread brain activity’, ‘internal consistency’ (i.e., discrimination between 2 inconsistent stimuli and response to only one), and ‘subjectivity’ (i.e., existence of a private flow of events perceived only by the experiencing subject). These will be mentioned later, in sections 7 and 13. One further criterion, of ‘self-attribution’ (no. 10 of Seth et al.,11), seems essential for the elevation of consciousness from a primary, or ‘core’, consciousness to a secondary, mind-based type of consciousness. This attribute can also be discussed (see the section ‘The idea of plant ‘oneness’): namely, whether plants possess a ‘sense of self’ – a sense of participation and interaction with their external environment but at the same time being aware of their separateness from that environment. This ‘sense’ of self would appear, at least from work with humans, to be the emergent property of ‘mind’ derived from the integration, or ‘sensory binding’ within the brain,35,37 of most of the processes implied by the aforementioned definitional criteria. In the particular context of plant consciousness, the question arises as to the number and type of processes which can be considered critical in support not only a primary type of plant consciousness, but also a sense of self-awareness. Unfortunately, unlike studies of human and some animals, it is not possible to interrogate plants directly; answers relating to cognition and memory, for example, can be received only indirectly through behavioral responses and, then, be comprehended only in the light of human knowledge systems, personal subjective experience, and degree of understanding. Nevertheless, relevant and intelligible responses from plants could satisfy criterion no. 11, ‘accurate reportability’, of Seth et al.11 In terms of Hameroff and Penrose's Orch OR hypothesis,1 the properties to which their experiential moments of protoconsciousness would be relevant are the above-mentioned criteria 1 and 3 of Seth et al.,11 ‘EEG signature’ and ‘widespread brain activity’. Thus, at least 7 of the 17 criteria of consciousness proposed by Seth et al. 11 are fulfilled.
Although specific criteria for consciousness were identified and extrapolated to plants (see above), it is ventured that the remaining 10 criteria of Seth et al.11 can also, in one way or another, be envisaged as relevant to some process or property of plants. The major problem, it seems, for both plant and animal consciousness, is how exactly the processes which are embodied by the criteria of consciousness are played out in the 2 respective ‘theatres of the mind’ to engender a sensation of existence within a world of sensory impressions. Humans create and recognize this world; and plants are presumably also constantly receptive of their environment; but are plants (and many animals) aware of this receptivity which brings into being some type of representation of the world in which they live?
The Penrose-Hameroff Orch OR model of consciousness
In an update of their controversial 20-year-old theory of consciousness published in Physics of Life Reviews, Hameroff and Penrose1 claim that consciousness derives from deep-level, fine-scale activities within brain neurons. The authors propose 3 variants of a possible way in which consciousness came to be developed in biological organisms, one of which is that consciousness is the product of discrete physical events that have always existed in the Universe, and which are governed by its laws, even if these are not fully understood at present. The resultant physical events are termed ‘protoconscious events’. With the advent of living forms during the course of evolution of the Universe following its inception, protoconscious events have become embodied or, as the authors put it, “orchestrated,” within living forms by virtue of their particular cellular composition and construction, and now find their most developed expression in neuronal activity of animals. From their site of inception in the brain (and here Hameroff and Penrose mean the human brain) the protoconscious events somehow lead to momentary ‘states of mind’ which humans, from experience, call ‘consciousness’.
For Hameroff and Penrose, the mediators of each protoconscious event are the proteinaceous dimers of tubulin which, together with their associated proteins (MAPs), are assembled into microtubules (MTs). All these macromolecules are ubiquitous in eukaryotic cells and, with the participation of actin and intermediate filaments, develop a scaffold, or cytoskeleton, which supports internal cellular structure and thereby cell function. Tubulin and MTs are particularly abundant in neurons. Because neurons do not divide (although new cerebral and spinal cord neurons can be generated from astrocytes), the MTs and cytoskeleton are conserved as relatively stable intraneuronal components, apart from a possible slow turnover of their tubulin and other molecular constituents, a process characteristic of MTs in all situations.38 Furthermore, MTs and their intracellular arrays show properties that link them to quantum events, such as quantum computing and quantum coherence1,39 and, hence, MTs are prime candidates for the quantum reduction events that generate protoconsciousness and which lead to the firing of brain neurons and axons that underpin episodes of consciousness. The recent discovery40 of quantum vibrations in MTs within brain neurons is considered to corroborate the theoretical expectation that MTs participate in consciousness by means of quantum reduction. Also relevant is the finding that MTs can generate electrical fields and, hence, provide a passage for electrical currents.41-43 The electric charge developed at the minus ends of MTs can also assist in cellular processes, such as mitosis.44 Furthermore, Hameroff and Penrose1 suggest that deep-level quantum MT vibrations are the source of certain frequencies found with electro-encephalograph (EEG) records.
The theory which embraces all these views and which postulates neuronal MTs as being a source of consciousness is called ‘Orchestrated Objective Reduction’ (Orch OR),1 so named in the belief that quantum vibrational computations and quantum coherence within MTs are ‘orchestrated’ (Orch) by synaptic inputs and neurophysiology. Moreover, MTs are held to be the sites where quantum state reduction (R) events take place which collapse to ‘objective reduction’ (OR) events in accordance with the Diósi-Penrose proposal in which quantised gravity plays a critical role.1,21 As mentioned, each OR event introduces to the brain a moment of protoconscious experience.
Following its proposal, Orch OR theory received much criticism (see refs. 1, 45 and 46 for replies to these criticisms, and see also refs. 47 and 48 for other sceptical remarks). At first, the animal/human brain was considered too “warm, wet, and noisy” for seemingly delicate quantum processes and quantum coherence, in particular, which previously had been observed only at sub-zero temperatures. However, evidence from plants has shown that quantum coherence participates in photosynthesis at ‘warm’ physiological temperatures.49,50 Furthermore, quantum coherence is presently considered to be the basis for both the navigational ability of birds and the process of olfaction.48,51 Interestingly, recent work on anesthesia, which selectively erases animal and human consciousness while sparing their non-conscious brain activities, indicates that it might result from a destabilization of MTs in brain neurons.52 This finding is also relevant to the question of a possible plant consciousness because some of these anesthetics affect plant growth and destabilize plant MTs also.
In their review,1 Hameroff and Penrose pose 3 questions relating to the origin and place of consciousness in the Universe. The first 2, roughly stated, are: (1) – If consciousness is not an intrinsic part of the universe, has it then evolved solely from complex computations within and among neurons, and those of the human brain, in particular? (This is a scientific and materialistic view of consciousness and would accord with known physical laws.) (2) – Has consciousness, in some sense or other, been present in the Universe all along, and by some extrinsic means is able to influence, or enter into, physical matter and human cognitive behavior? (This is a dualist, spiritual view and would not accord with present-day scientific paradigms.) The first of these 2 questions relates to consciousness as it manifests in the modern human brain, an organ which has come into existence during only the last 7 × 106 years. The second question, however, alludes to a deeper period of time, extending at least to the starting point of the present Universe, approximately 14 × 109 years ago; it also alludes to an unknown process whereby a presumed source of consciousness penetrated animate, biological material. Hameroff and Penrose believe that their Orch OR theory provides an intermediate, or third, way, posed here as question (3) – Does consciousness result from discrete physical processes which have always existed in the Universe, but which in recent times have become coupled with neuronal activity? (This accords with a scientific view of consciousness, although it incorporates not fully understood physical laws.) By means of this third way (the Orch OR theory) the authors believe that each of these seemingly different views of consciousness can be accommodated: if consciousness is a property of the material Universe then it follows that, as this Universe evolved, and with it the creation of conditions for the vivification of material condensates which we call “living organisms,” the physical processes requisite for consciousness have become enfolded within these living forms. It is possible that these forms have been constructed – and constrained – each in their unique way, as displays of evolutionary experimentation for the harnessing of consciousness in the most stable way.
Could Orch OR apply to plants?
Microtubules have been extensively studied in plant cells ever since their discovery in the root tips of Juniperus chinensis (Chinese juniper tree) and Phleum pratense (Timothy grass) by Ledbetter and Porter.53 The participation of MTs in the synthesis and orientation of linear chains of cellulose macromolecules from which plant cell walls are constructed accounts for the abundance of MTs in cells of all plant species. The cell-specific arrangements of MTs, as well as their spatial relationships with the nuclear envelope at which new MTs are seeded,54 appear to have relevance not only to cellular morphogenesis55 but also for the facilitation of specific pathways of gene activity, and hence, to the regulation of growth.56,57 Given the ubiquity of MTs throughout the Eukaryota, it is legitimate, within the framework of Orch OR, to ask whether MTs participate in the initiation of protoconsciousness by means of quantum events in non-human life forms, plants in particular.47,58 According to the survey by Gardiner and Marc59 of the correspondences between the macromolecular composition and developmental modifications of MTs of both neurons and plant cells, there seems to be no structural feature that would preclude plant MTs from participating in putative Orch OR events. However, the total mass of tubulin in plant cells may be a limiting factor for these events to occur with a meaningful frequency, although as will be seen (section 14), there may be situations in which this limitation is invalidated.
A structured view of consciousness
Before discussing either quantum-derived protoconscious events or the consciousness of living forms (animals and plants, especially), it is necessary to prepare a structured scheme of consciousness for use in further discussion. First, however, the remarks of Bohm and Peat (ref. 60, p. 209) are noted, that “whatever we say a thing is, it is something more and also something different … If we say that consciousness is a material process, this may well be fairly accurate … But it is also more. Its ground is in the infinite depths of the implicate and generative orders, going from the relatively manifest on to ever greater subtlety, the totality of which will always elude the grasp of science.” The authors introduce the idea that all potentialities are enfolded within a source, the implicate order, which is, roughly speaking, the quantum world; the potentialities referred to are organized and made substantial by information present in the generative order, the world in which forms of material existence unfold. This view is in keeping with what Hameroff and Penrose propose, that consciousness can be understood as an unfolding into the material world of some potentiality contained within the quantum world via an OR event. The more subtle aspects of consciousness mentioned by Bohm and Peat60 reside further ‘down-stream’, within the generative order, and may take the form of some rarefied, transcendent form of consciousness.
Although Bohm and Peat (ref. 60, p. 210) also make the point “that there is no absolutely sharp ‘cut’ or break between consciousness, life, and matter, whether animate or inanimate,” a remark which seems to align the authors with panpsychism, an area which Hameroff and Penrose1 assume is beyond the reach of scientific analysis, most people would suppose (for heuristic purposes) that breaks do indeed occur: firstly, between inanimate mineral materials (e.g., stones, mountains, seas) and animate organic material (e.g., prokaryotes, eukaryotes) and, secondly, that another break occurs within the animate group, dividing eukaryotes into 2 major kingdoms, the Plantae and Animalia; and then, thirdly, that there is a further break between the consciousness of non-human animals and humans. It becomes a matter of conjecture whether, with the advent of ‘mind’, human consciousness has become something radically new in evolution (ref. 61, p. 165 et seq.), setting human consciousness apart from non-human forms of ‘conscious’ neuro-cerebral activity.
It is proposed that living organisms, animals and plants, have potentialities for consciousness distributed between 3 or 4 apparatuses, or recognizable states:
– Sensorium,
– Brain,
– Mind,
– Over-mind.
This may be a simplistic approach, and certainly it is less elaborate than descriptions given, on the one hand, by Vaneechoutte62 (in everyday language) and, on the other hand, by Edelman63 (in the sophisticated language of neuroscience). Nevertheless, these 4 states provide the substratum for consciousness, as it has been generally studied up to the present, and may be sufficient to comprehend Orch OR in relation to both animal and the conjectured plant consciousness.
The distribution and combinations of these states 1–4 among living forms are as follows:
1: Prokaryotes, Protozoa.
1 + 2: Eukaryotes − Animals and Plants.
1 + 2 + 3: Humans (psychostatic).
1 + 2 + 3 + 4: Humans (psychokinetic).
The four states can be described in more concrete terms:
1 – Sensorium. That portion of a living organism, irrespective of whether plant or animal, which exists in a state of ‘sensitive awareness’ (ref. 60, p. 211) or of ‘experiencing’.62 Furthermore, the corresponding sensitive material may be enclosed within entities which characterize the hierarchically distributed organizational levels of a Living System,64 each level enclosing 3 canonical sets of sub-systems. The set of sub-systems that process matter and information are particularly relevant to the sensorium.
The first 3 organizational levels, n, n + 1, n + 2, correspond to Cell, Organ, and Organism. All living beings are composed of entities at these levels though, in the case of unicells (Protozoa), the Organism and Organ are enfolded within the Cell. Moreover, the Organism is itself an entity within the next-higher level (n + 3), in a continuation of the hierarchical system wherein there are further levels of increasing complexity (Demes, Clans, Societies …).64,65
The mentioned ‘sensitive awareness’ is an attribute of entities at every level of a Living System. Moreover, as Vaneechoute62 points out, experience (in his terminology), or awareness, may extend downwards, to the molecular level of enzymes, or any item which has the potentiality of connecting with another item, thereby forming a new unit. Awareness is thus a mediator between 2 potential partners and leads to the creation of a third item, as in a chemical reaction. Awareness may be accompanied by the passage of time, often represented as an aging process, and accompanied by some biological marker, or counting mechanisms, such as the step-wise loss of telomeres from chromosomes of animals and the consequent limitation that this places upon the proliferation of their cells.66 It is not known whether the aging of plants is regulated in this way. Another aspect of temporal awareness could be due to regular quantized variations of cell mass (see the later section ‘Moments of conscious awareness’).
‘Awareness’ applies not only to the perception of the inner environment of a living entity, at the level of the cellular interior and that of the levels immediately above, but also to the perception of the outer environment of each level. For example, the Cell may be aware not only of its internal ‘self’ but also of its immediate environment, that is, of the Organ of which the reference Cell is a part. At these levels of Cell and Organ, the means of environmental perception may involve, respectively, stretch receptors located in membranes and be coupled with ion channels and actin filaments (elements of the Cytostructural Code), and ionic gradients and electro-potential differences (elements of the Bioelectric Code 67). Similarly, the Organism is aware not only of its component unitary Organs but also of its ‘Umwelt’ (using this term in the sense of the local environment, which may include the artificial, or built, environment produced by certain animals to protect themselves), as well as its external natural environment, or biotic complex of soil, vegetation fauna and climate (‘Welt’ or ‘Nature’). From this perspective, awareness and consciousness evidently have behavioral, ecological and evolutionary implications.
One aspect of sensitive awareness is the question of whether it is related to the phenomenon of ‘primary perception’,68 of which more will be said (section 11). That is, the perception, at the level of Organism (say), whereby one organism can sense and respond to the presence of another organism (as in swarming of insects, for example). The range of such perception is said to extend between bodies as far as 143 Km apart1 and may depend upon a quantum coherence effect due to non-local entanglement between the individuals within swarms. Such primary perception may also exist between the different parts of a single body, also by means of quantum, non-local entanglement, or by coherence of neighboring electromagnetic field over relatively short distances. Such perceptions may lead to a sense (in humans) of contentment and completeness or, contrariwise, to a sensation of malaise and apathy.
Because the Orch OR theory is centered on human consciousness (though, as mentioned, this may not be the only consciousness to which the theory could apply), the sensorium (state 1) can, in this context, be separated into 2 parts: (1a) soma (body and locomotory apparatus) together with the somatic nervous system, and (1b) emotional, affective center responsible for bodily ‘feelings’, which are dependent upon the sympathetic and para-sympathetic divisions of the autonomic nervous system, and which are supported by a neuro-endocrine system.69 Links have been shown to exist between particular emotional states and somatic muscle systems.70 Results of the survey made by Beedie et al.71 suggest that ‘feelings’ are distinct from ‘moods’; the former reside within the body whereas the latter reside within the brain and result from its changeable chemical composition (see next part, ‘Brain’).
In higher plants, ‘soma’ usually refers to the vegetative body (sub-state 1a) of the sporophyte generation. Then, the plant counterpart of the ‘emotional center’ of humans and animals (sub-state 1b) is comprised of the plant phytohormonal system which, together with the cells of the phytoneural system that transport phytohormones, is embedded within the soma,36,72 In both animals and plants, the hormonal/phytohormonal systems prepare the soma for movement.
Soma, sub-state 1a of Sensorium, can participate in either a state of motion (1a+; activity being indicated by ‘+’) or in a propioceptive state of meditative quietude (1a−; passivity being indicated by ‘−’). Similarly for the emotional center (sub-state 1b): it can adopt 2 alternative aspects, 1b+ or 1b−. The same may be said of Brain (state 2 – see below): it may be occupied by meditative quietude (2−) or be alerted to activity (2+), whereupon it would be occupied by thoughts and associations.
2 – Brain. In the animal kingdom, this apparatus may be localized at a particular anatomical site and constructed of the conventional, animal-type neurons. By contrast, in the plant kingdom, ‘brain’ is not a word commonly used, and it is doubtful whether there is a single site where a ‘brain’ is located (see below). Comparative anatomy of animals suggests that brains, neurons and synapses have arisen many times during evolution.73,74 This leaves open the possibility that a plant ‘brain’ could be one of a number of parallel evolutionary innovations in organisms in toto, the significance of which is to perceive and coordinate responses to disturbances within the immediate environment of the organism in question. In addition, during animal evolution, there may have been more than one route to consciousness. For the moment, however, we focus upon MTs as providing this route, at least to momentary protoconsciousness, in accordance with the Orch OR hypothesis of Hameroff and Penrose.1 Thus, it is likely that an analog of the animal brain exists in plants, though it may have a dispersed location among the numerous primary growing points of plant organs where MTs are numerous. Another type of dispersed brain may be located within the secondary growth systems of trees, within cambium and secondary vascular tissues (see the later section ‘Does Orch OR apply to plants…’). Furthermore, both plant growth systems, primary and secondary, have access to a phytoneural/hormonal system, as does the proliferative system of animals with its neuronal and hormonal components.
The proposed dispersed brain of plants is a consequence of their ‘open’ manner of construction, which traces to the continual production of elongating and ramifying primary growing points from the meristematic termini of the organs composing the root and shoot systems. This contrasts with the ‘closed’ plan of animals, where growth and ramification (more evidently of cell lineages rather than of organs) are internal, within the soma, first within the embryo and later continuing into the adult organism, where a constant kinetic balance is maintained between cell proliferation, differentiation, and aging accompanied by cell loss. However, a ‘closed’ constructional plan is also a feature of the internalized secondary growth of plant organs (in trees, for example) though, here, dead cells (woody fibers and tracheids, bark) are retained over long periods within the plant body.
One element in the open type of growth of plants is the apical meristem. It is responsible for cell proliferation and the provision of cells for elongation growth. The ‘transition zone’, a short region interposed between primary root meristem and elongation zone, has been proposed as having a brain-like status and thus to act as a center both for the exchange of information between root and shoot systems and for initiating appropriate growth and physiological responses.36,75,76 The multitude of primary root meristems contained within a root system, each meristem with a putative brain-like zone, would loosely satisfy Seth et al.'s11 criterion no. 3 of consciousness, ‘widespread brain activity’.
Although the Brain of animals may, anatomically, be a hierarchically organized structure due to its phylogeny (Feinberg, 2013),77 the whole organ consists of 2 major compartments. One is of cellular matter, the other of cerebrospinal fluid, the latter being more than simply a matrix for the former, the fluid component being distributed between several distinct ventricles. The cellular and fluid compartments support their respective types of communication system,78,79 the former providing wiring transmission (WT) – comprised of electrical impulses associated with intercellular gap junctions and synapses, the latter providing volume transmission (VT) – with the diffusion of chemicals and vesicles within the cerebrospinal fluid. Together, the 2 compartments may be considered to constitute a ‘Global Molecular Network’.80 The chemical make-up and chemical exchanges between fluid and tissues of this network may account for ‘moods’. By analogy with human ‘moods’ based on chemical status of particular parts of the brain, plants may also be considered to experience ‘moods’: for example, when a root system is water-logged and deprived of oxygen, CO2, ethylene and abscisic acid (the latter 2 being phytohormones) accumulate within, and exchange between, the apoplasm and symplasm of the affected tissues81,82 and bring about a ‘wilting’ response in the shoot system, which could be considered analogous to a human VT mood response. Thus, plants, too, may have their own type of Global Molecular Network.
3 – Mind. This entity is, arguably, a property of humankind; but whether Mind is unique to humans is not known. Mind can comprehend the whole (holon) with which it is associated. It is sometimes described as the silent, reflective ‘observer’ of the human condition. Comprehension of the holon is by means of a binding together of (a) a brain-derived ‘knowingness’, which is the recognition of external, sensorially derived impressions received from states 1 and 2, and (b) an internal impression of self received from the soma and the emotional/hormonal component of Sensorium. Knowingness includes the products of impressions learned and committed to memory formed from both abstract, learned knowledge and concrete, innate knowledge, the latter giving rise to skills, fashioned initially from unlearned instinctive responses. The state of self-awareness, or ‘presence’ of the holon, comprehended by the state 3 of Mind, may, fundamentally, be one in which there is also introduced a subliminal awareness, or consciousness, of the passage of time. We shall discuss this temporal aspect of awareness (see section 10). The ability to contemplate and reflect upon the condition of the self and the selfhood of others5 derives from the relationship which the organism bears with respect to its next higher organisational levels: the pair, group, or society of which it is a part (the ‘Umwelt’ of the organism).
The reflective aspect of Mind may be summarized by deconstructing the compound sentence “I am sensing; I am feeling; I am thinking” into 3 grades. “Sensing” and “feeling” are the respective attributes of sub-states 1a (soma) and 1b (affective center) of the Sensorium. Thus, deconstructing to the first grade, to give “_ _ sensing; _ _ feeling,” would apply to Sensorium (state 1) only. Then, the second grade, “_ am sensing; _ am feeling; _ am thinking,” would apply to states 1 and 2, because of the inclusion of experience from the Sensorium and Brain, and can be alternatively notated as state 2[1]. Finally, the complete sentence, at the third grade, “I am sensing, feeling and thinking,” would apply to the sensory binding of the self-reflective Mind, more comprehensively notated as state 3[1,2]. This third grade is due to a Mind that knows that it is sensing, feeling, etc. With the emergence of this state, a ‘core’ of self-consciousness may be said to be present. Nevertheless, there may be a deeper, more prototypical, consciousness where there is simply a sensation of presence, of ‘am-ness’. This might be considered to be state 3[1a−,1b−,2]: one of protoconsciousness6 emphasized within state 3 by directing the attention of the Mind specifically upon sensation of the soma in its passive, or meditative, aspect, here indicated by the subscript 1a−, and where the very small subscript characters indicate the diminution or extinguishment of sub-states 1b and state 2. It is within this state 3[1a−,1b−,2] of Rosenbergian protoconsciousness that Hameroff and Penrose's Orch OR protoconsciousness1 can be experienced most directly – as a state of pure experience without cognitive awareness. As Rosenberg (ref. 6, p. 241) asserts, “Any natural individual is at least protoconscious: It is an experiential nexus even if it does not support thought.” Such a state would be represented as 3[1a−,1b−,2].
Mind (state 3) may sometimes be separable from the first 2 states, 1 and 2, as evidenced both by near-death, out-of-body experiences.83 This condition can be notated as 3[1,2]. Mind may also be uncoupled from Sensorium and Brain during journeying to the ‘spirit realm’, in the case of shamans and modern-day shamanic practitioners.84 Interestingly, near-death and, more specifically, out-of-body experiences, have been discussed in terms of quantum entanglements that link the consciousness of the material body and the out-of-body entity.85,86
4 – Over-mind. This elevated type of human consciousness emerges only when state 3 of Mind is accompanied by sensation of states 1 and 2 (either in their active +, or their passive −, aspects) together, even if only for a moment. The ‘togetherness’ of states accompanying the attainment of Over-mind is brought about by a voluntary exercise of ‘attention’.87 Linkage of states by means of attention can be notated using the undertie ‘ᴗ’ in place of the comma ‘,’, the latter having formerly denoted the separateness of states (as contrasted with their togetherness due to attention). Thus, Over-mind is notated 4[1ᴗ2ᴗ3]. It is a precondition for insight or intuition,60,88 or direct perception, in the sense used by Goethe.89,90 The fact that it can be achieved marks the development of a psychokinetic aspect of humanity, as contrasted with the previous psychostatic aspect which characterizes core consciousness of state 3 (ref. 61, p. 249).
For completeness with respect to the natural history of consciousness, ’higher’ states of consciousness should be mentioned, thereby recognizing the mentioned psychokinetic, or spiritual, nature of Man. Fortunately, this aspect of Man is no longer viewed as a source of tension with respect to the science of consciousness23,91 (see also volume 8 part 4 of the journal ‘NeuroQuantology’ (2010), an issue that contains 7 papers dealing with ‘Experimental NeuroTheology’), even though the relationship between these ‘higher’ states and the Minds of states 3 and 4 are not well defined and have not yet been brought within the domain of experimental psychology.
5 – Superconsciousness. It is possible that, beyond state 4, further conscious states exist – one of which was earlier referred to as transcendent consciousness. It is the state known as Samadhi. When realized, this state, state 5, completes the holon of human existence. According to Sadhu (ref 92, p. 45), Samadhi is not exactly a state of mind because it exists in the absence of mental activity and thought, provisionally notated as state 5[1a−ᴗ1b−ᴗ2ᴗ3ᴗ4]. Its nature is difficult to express in words. Samadhi, also referred to as a state of ‘enstasis’, is complete when state 4 has become sustainable. It may endure for only seconds of time; but three weeks is said to be the limit of endurance for this enstatic state.93
Samadhi is said to consist of 6 stages, the first of which (state 5.1) is described in Hindu terminology as ‘Kevala Nirvikalpa Samadhi’ (temporary formless Samadhi) appears to overlap with state 4 (Over-mind), except for the fact that the state of Samadhi exists with the absence of any evident thought, i.e., state 5.1 = state 4[1a−ᴗ1b−ᴗ2ᴗ3], which also would be present in state 4. The ultimate state, state 5.6, ‘Sahaja Nirvikalpa Samadhi’ (uninterrupted formless Samadhi), is one in which the mind is completely transcended (ref. 92, p. 55), as at the moment of death of the physical body, states 1 and 2, i.e., state 5.6 = 5[1a−ᴗ1b−ᴗ2ᴗ3ᴗ4] In fact, this final state may be postulated as the 6th attainable state of consciousness and to be incomprehensible to ordinary scientific knowledge
State transitions
Activity of the first 2 states, (1) Sensorium and (2) Brain, define, for practical purposes, the condition of the ordinary, non-reflective human being. The integration of these 2 states to produce a ‘core’ consciousness5 of state 3 (or state 3[1,2]) is rarely complete. Humans commonly live only with glimpses of reflective self-awareness (the feature of Mind – state 3), and often are only dimly aware of their own states 1 and 2, unless suffering from some pathological malady, the extreme sensation of which draws attention to a particular locality of the Sensorium. Only infrequently does state 3 make the transition to state 4. Then, as poetically described by James Joyce (ref. 94, p. 180), a ‘flash’ of (human) consciousness comes about when the usual state of sleeping-wakefulness (the psychostatic state 3[1,2]) switches, apparently spontaneously, to wakeful-wakefulness (the psychokinetic state 4[1ᴗ2ᴗ3]), but then, just as spontaneously, reverts to the former, more usual, mundane level:
“His thinking was a dusk of doubt … lit up at moments by the lightnings of intuition, but lightnings of so clear a splendour that in those moments the world perished about his feet as if fireconsumed: and thereafter … he felt that the spirit of beauty had folded him round like a mantle. But, when this brief pride of silence upheld him no longer, he was glad to find himself still in the midst of common lives …”
Dumit,95 in his description of ‘Sensorium’, sees it as being more in line with Mind, of states 3 and 4, as “a sensing package that constitutes our [human] participation in the world … [its] assemblage of sensory inputs defines our boundaries, making the world present to us and by subtraction making us present to ourselves as beings in the world.” By ‘subtraction’, this author is understood to mean a lapse of entrapment by internal distractions (a property of an active Brain, state 2+), or ‘letting go’. By this action, the internal world of state 1 and the stream of impressions from the external world (Umwelt and Welt) can be brought together, through the agency of the binding-factor property of the Brain, into the embrace of Mind (state 3). We can take this further to mean that, by ‘subtraction’, the Sensorium of Dumit enables the linkage of states that lead to authentic self-consciousness, state 4: a state of presence and sensitivity to both the quantitative and qualitative aspects of being, which together form the essence of livingness: in other words, the ‘letting go’ pertains to all those sensory inputs and consequent displays of knowingness which entrap the Mind (state 3) and screen it from apprehending the self of which Mind is a part.
The opinion of Hoeller96 points in a similar direction, to the conditions for the transition from state 3 to state 4. For him, “subtraction” is a precondition for the acquisition of ‘true’ knowledge, or the state of gnosis, a property of Over-mind, state 4. He says of this state that it “does not consist so much in adding some miraculous external element to our consciousness; rather, it requires a subtracting from our minds and lives of much material that serves to obstruct insight.” In other words, these obstructions are due to certain regenerative or self-perpetuating neuronal circuits generated by the Brain (state 2). These are ‘associations’ based on memory rather than true ‘thoughts’; they participate in the development of the ‘ego’ and personality (components of Mind, state 3). There are also obstructions due to emotional prejudices and irrational expectations – all things that comprise personality – which hinder attainment of state 4. It is probable that each of these state transitions, as well as other transitions not mentioned in this section (i.e., states 4 to 5 to 6), are accompanied by particular alterations in the EEG profiles of the individuals accomplishing these transitions.97 Thus, each of the proposed states may have its own electroencephalographic signature.
A brief comment on ‘Umwelt’ and ‘Welt’ is appropriate here. They are implicit in the title of Arthur Schopenhauer's major work, “World as Will and Representation” (Die Welt als Wille und Vorstellung),98 ‘World’ being that particular representation (which Schopenhauer called ‘The Veil of Maya’, taking this term from Hindu philosophy) of Welt and Umwelt produced by the Mind (state 3). Friedrich Nietzsche99 expressed a similar view, as indeed did Descartes before him, though perhaps not quite so forcefully (Seager3). Using the analogy of the Apollonian world of dreams and the Dionysian world of action, Nietzsche (ref. 99, p. 15) writes, “Men of philosophy even have a sense that beneath the reality in which we live there is a second, quite different world, and that our own world is therefore an illusion.” For parity with the term ‘state’, the word ‘realm’ is used to denote the illusory external world; thus, this representational (or illusory) world is called realm 3. At the level of higher states of Mind, it can be presumed that other representational worlds become evident, but at the same time more authentic: realms 4 and 5 could, for example, be representations of a more authentic reality due to Minds that have attained states 4 and 5. It is possible that these realms may be partially accessible during episodes of non-ordinary consciousness in which the intensity of sensations are preternaturally heightened (though the spectrum of sensations is apparently not widened100,101), as described by Harner84 in relation to trance- or drug-induced episodes. These realms exist in parallel, as para-realm 4 and para-realm 5, with the realms of ordinary or enhanced consciousness, of states 4 and 5. It seems that increased vividness of the representations of these para-realms result from an enlarged range of electrical connections within the brain.102
From the secularized Christian position, Theilhard de Chardin103 formed the idea of an ‘Omega Point’ at which consciousness merged with a cosmic field of energy – one which Bennett (ref. 61, p. 130) referred to as an ‘Eternal Potential Energy Field’, and which theoretical physics posits as the ‘Zero-point-fluctuation State’.104 This state (state 7, which we shall mention later), is probably equivalent to the Buddhist state of Nirvana. It may be associated with an ultimate representational realm 7, and is reached upon transition from state 5.6 (=state 6), Sahaja Nirvikalpa Samadhi.
Bennett61 believed that each conscious state transition was associated with the liberation of a different type, or quality, of energy. However, in this notion, he was pre-empted 64 years earlier by Charles S. Minot: in his presidential address to the American Association for the Advancement of Science, Minot wrote that “if it has any real power consciousness must be able to change the form of energy …,” and further, “it may be that without consciousness the universe would come to absolute rest.”105
Sensitivity of plants toward their environment
The sensory faculties of plants and animals are attuned to physico-chemical properties of the wider natural environment (Welt). Disturbance or change of these properties automatically initiates biological response in conformity with physical laws operating on biochemical reactions. In the case of plants, responses are commonly expressed as differential growth movements (tropisms) toward or away from particular stimuli. Differentials of light intensity and humidity, shear forces at organ boundaries, are some of the stimuli which evoke such tropic responses. The transducers of the responses are often macromolecules sensitive to weak mechanical forces (e.g., MTs, actin filaments, intracellular motors). Organisms are also sensitive to stressors, and these can elicit adaptive responses: for example, responses to temperatures outside the optimal range for growth can lead to fabrication of adaptors such as heat-shock proteins, natural anti-freeze molecules, and desiccation responses. There can also be responses in the form of intracellular modifications linked to the organism's life-cycle, such as the seasonally dependent conversion of rough endoplasmic reticulum from a fluid form to a quasi-crystalline form.106,107 DNA itself is also sensitive to environmental stressors, enabling this molecule to accept structural, epigenetic modifications which have consequences for future generations.26,58,108
Departures from the environmental optimum may modify organismic form. Plant morphology is especially plastic, as evidenced by the response to external conditions: shade (relatively low light intensity) gives rise to etiolation of herbs and to the self-pruning of tree branches, to mention 2 examples; and, at the organ level, leaves, especially, show plastic development in response to varying light quantity and quality.109 Form can also be modified by the internal environment: the aging of the organism is often accompanied by a ‘phase transition’, with a change from a juvenile to an adult form, probably as a result of a corresponding change from a juvenile to an adult physiology, which may be referrable to epigenetic modification (viz. the altered leaf form of ivy, Hedera helix, which contributes to an adult climbing habit, or the cephalia of Melocactus spp., stem modifications that support floral apices). At least some of these situations are reversible. Etiolation is ameliorated by increased light; and a juvenile plant form can be restored when old tissue is removed.
There is a broad range of commonly encountered physico-chemical stimuli to which roots will react, but they do so with a relatively restricted repertoire of responses. As discussed elsewhere,26 the growth responses that are directed from a ‘root-brain’ zone are innate and, thus, deterministic. The type of response can nevertheless be prioritized according to thresholds: that is, from the diverse in-coming physico-chemical stimuli, the root sums up the respective units by which the stimulus can be expressed (as composite units, such photons per unit time, or degree hours, for example) to some critical threshold value.110 The summation of units in the lead-up to a response is considered to be a type of ‘facilitation of learning’, one of the criteria of consciousness (no. 14) of Seth et al..11 Whichever composite unit first exceeds a threshold initiates a response in accordance with the physical signal whose units were summed. There can also be a decay of units, which can modify the rate at which a threshold value is achieved. In fact, there is evidence concerning how one stimulus can take precedence over another. For example, when a plant root is presented simultaneously with 2 different physical conditions which, individually, would elicit either a) hydrotropism or b) gravitropism, it is hydrotropism which prevails.111 It appears that the water-stress related to the conditions for hydrotropism brings about the regression of the starch-bearing amyloplasts,112 these being the sensors which, otherwise, would have initiated gravitropism. Some similar physiological competition between gravisensing and phosphate content of soil may be responsible for the modification of the gravitropic liminal angle of roots.113 In each case, the particular physiological response brings about a correspondingly re-directed flow of auxin which accordingly modifies the growth differential between opposing sides of the root.114 The ability to discriminate between 2 different and competing stimuli is in accord with the consciousness criterion no. 7, ‘internal consistency’, of Seth et al.11
It is proposed that such summations of stimulus-related composite units are bases for plant memory, this being a faculty generally considered to be a feature of consciousness.5 The ability to memorize is a faculty that is attributed to plants.24,26 The decay of developmental units (mentioned above) is analogous to memory loss, or forgetting, and the tropic response to the first threshold reached of 2 alternative summations may be a marker of the consciousness criterion ‘internal consistency’. These threshold responses are nevertheless predetermined (instinctive), and are not evidence of ‘intelligence’, in the sense that a root (say) has foresight of the consequences of its response to one signal or to another, and then chooses between them. The word ‘intelligence’ is often misused, largely because of the ease with which patterns of plant behavior can be extrapolated from a template of animal or human behavior (anthropomorphism). A reasonable defining attribute of intelligence is, to paraphrase the carefully chosen words of social theorist, Harvey Jackins (quoted by Garrard,115), “the ability to create new, exact responses by comparing and contrasting new information [with that] already ‘on file’ from past experience, and [then] constructing a response based on similarities to past situations but modified to allow for the difference.”
The above definition of intelligence also aligns with the view of the philosopher, Henri Bergson.116 For him, intelligence is more than mere information gathering, accompanied by an appropriate response: he remarks, “there is intelligence wherever there is inference …, which consists in an inflection of past experience in the direction of present experience.” There is no projection of future intention; nevertheless, some element of memory is implied in both Jackins's and Bergson's statements. Or, more succinctly, as paraphrased by Gunter,117 “Instinct involves the use of tools which are parts of the body; intelligence involves the use of tools distinct from the body. Instinct involves knowledge – an unlearned awareness – of living things. Intelligence, by contrast, involves the capacity to shape non-living matter.” Bergson116 concludes that “there is no intelligence in which some traces of instinct are not to be discovered … that is not surrounded with a fringe of intelligence. The two are complementary and mutually antagonistic. In plants, consciousness and mobility lie dormant; the main features are vegetative “torpor” and instinct. Animals by contrast are endowed with greater consciousness and mobility.” Plant behavior is, in other words, largely instinctive (deterministic), although located beyond the fringe of instinctive responses there is a brain-in-waiting – the plant's torpid behavior and its limited movement are indicative of a dormancy of consciousness: for Bergson,116 movement is always an indicator of consciousness, an idea rediscovered by Sheets-Johnstone.15 However, if intelligence “consists in an inflection of past experience” (i.e., memory), then it would seem that intelligence is fed by one of the aspects of consciousness, even though intelligence is not a necessary attribute of consciousness itself.
In relation to the structured scheme of consciousness (section 5) composed of 5 states of development, plants may, firstly, be said to be characterized by ‘torpor’ of their Sensorium (state 1); they do, nevertheless, also show a small degree of bodily mobility, which is the reactive, instinctive response to sensation. This latter attribute corresponds to the Bergsonian surrounding ‘fringe’ of brain-like activity (state 2). However, the property of mobility is one of degree. It is the nature of plants to be sessile, not mobile. Even though plants and animals share many fundamental genetic and biochemical properties, their respective body plans and modes of development differ, thus permitting only analogies, not absolute comparisons, to be made between plant and animal processes. Plants are of a class sui generis within the realm of living organisms. Secondly, brain-like activity in plants may be more developed than is generally thought. If this activity is supported by a phytoneural system (see section 13) capable of MT-derived OR events, then states 1 and 2 could be integrated and brought to a focus by some type of sensory binding within an emergent state 3, a state that may already be receiving impulses of protoconsciousness from OR events or, more directly, from a sensation of self, state 3[1,2]. It follows that the MTs of plants and their facilitation of OR events could serve as the bases for self-awareness, as proposed by Hameroff and Penrose1 for the human mind (state 3). And, as will be mentioned in section 11, quantum events that impinge upon neural activity may be responsible for occasions of ‘primary perception’ whereby 2 separate sensoria become aware of each other at a distance. These, too, might be brought to a focus within a mind at state 3.
With all these factors in view, we can now glimpse not only the circumstances under which plant ‘sensitive awareness’ exists and might contribute to the evolutionary conditions that develop and construct consciousness, but also how the Hameroff-Penrose Orch OR theory of consciousness might relate to this, especially when it is considered how much of plant development relies on MTs, the very items presumed crucial to OR events. However, the MTs that contribute to growth and development may be sites of other quantum events, such as quantum computations from which there is the possibility of initiating certain events deemed improbable as, for example, those described in the context of the development flower structure,118 and in relation to anomalous patterns of cell division.119
The central point of Orch OR theory
MTs have been found to be excellent electronic conductors, conduction taking various routes through the MTs, along their axes, and along and around their surfaces. The tubular structure itself promotes greater conductance than is possible through single tubulins; and conductance increases with MT length.1 Importantly, MTs provide a vector for the electronic conduction. Orch OR theory proposes that MT quantum vibrations (in megahertz, MHz) interfere and produce much slower ‘beat frequencies’, and that these can be recorded by electroencephalography (EEG), a technique which records the collective pattern of neuronal activity in the brain and separates the pattern into bands (α, β, theta, …) of different frequencies. The mentioned beat frequencies are in the gamma range and are typically of 40 Hz frequency.40 The implication is that this 40 Hz gamma frequency is produced somewhere within the brain by its complement of neuronal MTs. Moreover, the beat frequencies, which apparently arise as a consequence of the disposition of tubulin as MTs, result in slower and, thus, more realistic, timescales for the repetitive OR events to be ordered as a conscious substratum than would be possible if the tubulin primary energetic vibrations of around 10 MHz were solely responsible for OR events. Thus, it seems that the organization of tubulin at specific sites into macromolecular MTs, such as occurs at the synaptic inputs1 of animal neurons and the MT-organizing centers of plant cells (see later), assist in the orchestration of OR events as well as the generation of beat frequencies, such as those of 40Hz recorded by EEG.
There is one known post-translational modification of MTs – the acetylation of α-tubulin by acetyl-transferase120 – which may be important in this respect. It is thought that the acetylated sites of neighboring MTs are bridged by tau, a microtubule-associated protein (MAP), and that MT bundles are stabilized thereby.121,122 Tau is highly expressed in neocortical regions and the hippocampus, which suggests that tau has a role in neuronal activity, and that its presence in MTs underlies both fast and slow brain oscillations as well as long-range co-ordination of electrical signaling between the neocortex and hippocampus. Displacement of tau from brain MTs not only destabilizes the MTs and intracellular architecture but also impairs cognitive function, as evidenced from Alzheimer's disease.123 Lack of tau protein also compromises neuronal functioning.124 Furthermore, it is probably significant that genetic knock-out of tau in Tau −/− mice brings about reduced levels of gamma synchrony involving the frontal cortex.125 The last-mentioned result suggests that bundles of MTs containing tau MAPs might be effective in promoting the beat frequencies at 40 Hz.
From the point of view of consciousness, the modulation of the gamma EEG frequency is of interest in another context. Buddhist monks during their meditational practice are able to self-induce high-amplitude gamma-band oscillations and phase-synchrony, particularly in the neocortical region of the brain,126 an effect not seen in novice meditators who had only a week's training in meditational practice. These results concerning enhanced gamma frequencies were confirmed with a different group of experienced meditators.127 Moreover, a recent development in the study of gamma EEG has shown that, by applying an AC current with a frequency of 40 Hz to subjects during a period of REM sleep, an interval of ‘lucid’ dreaming could be interposed.128 In this state, the subject reportedly appears as an observer within his/her own dream, and is even able to intervene and comment on the action of the dream (ref. 129, p. 324). Stimulation with lesser (2 – 25 Hz) or greater (75 – 100 Hz) frequencies during the REM period failed to affect the content of the dream, which remained ‘non-lucid’. Together, these 2 sets of results suggest that the 40 Hz frequency is associated with elevating the Mind of state 3 into state 4 (Over-mind) whereby prolonged periods of self-observation would be possible. Interestingly, a flicker pattern of 50 Hz presented to the eye can be perceived subliminally by the human mind and is also able to induce moments of awareness.130
A further reason why the 40 Hz gamma frequency is of interest is that this frequency is believed by Hameroff and Penrose1 to be implicated in an OR event. The clue to this relationship lies in the equation τ ≈ ħ/EG, where EG is the gravitational self-energy, ħ is the Dirac-Planck constant, and τ approximates to the duration of successive OR events. EG may also be regarded as the mass difference between 2 states of tubulin in superposition; it is the superposition of the masses of these molecules in the form of quantum ‘bits’ which entangle, compute, and then collapse to thereby elicit an OR event.1,20,21
Evidently, if the value of one of the variables in the equation τ ≈ ħ/EG is known, the value of the other variable can be estimated. Thus, when τ has a value of 25 ms, as given by the 40 Hz gamma frequency, the estimated EG is equivalent to a mass difference occurring during superposition within a population of an estimated 2 × 1010 tubulin molecules. However, each cerebral neuron, instead of containing 2 × 1010 tubulin molecules, is estimated to contain only about 109 such molecules, a tubulin mass which would result in τ = 500 ms (2 Hz). Moreover, it is presumed that only a fraction of these 109 molecules, when assembled into MTs, as well as being in superposition, would be directly involved in a coherent event of τ = 25 ms duration. Thus, in order to amplify the self-energy mass to the level necessary to elicit 40 Hz gamma synchrony, it follows that an aggregate of approx. 20,000 neurons would be required. It is then supposed that the frequency of such 25 ms OR events within the brain is such that they produce a continuous stream of protoconsciousness.
How do such estimates of tubulin mass and neuronal numbers apply to non-human animals and to plants? It is known that the nematode, Caenorhabditis elegans, has exactly 302 neurons.131 From the quantitative considerations above, it seems that MTs of the nematode organism would barely achieve the threshold energy-mass required for gamma-synchrony and, thus, are unlikely to be continuously conscious. Animals (and plants) with inherently fewer tubulin molecules may achieve conscious moments at a much reduced rate (due to the larger τ values associated with an OR event). In the case of C. elegans, τ would be estimated, on the basis of the data to hand, to have a value of 500ms.132 A similar rate may apply to plants, as evidenced by their spontaneous action potentials (see later). This would mean that protoconscious events are less frequent and, hence, consciousness is discontinuous.
Because it follows that, when EG has a smaller value and the value of τ is consequently increased, indicating a longer time between successive protoconscious events, different values of τ may be taken to represent a sliding scale, or grade, of consciousness. That is, larger values of τ would indicate a lower grade of consciousness, whereas smaller values would indicate a higher grade with more continuous spells of consciousness. A similar consideration applies to EG: its value, which is purportedly related to the density of MTs and their tubulin dimers within certain critical cells, would also bear some relation to the intensity of conscious experience. Given these considerations concerning τ and EG, it may be possible to set criteria for the grade of Orch OR-related consciousness in non-human organisms, including plants: these criteria would hinge upon the number and density of MTs.
There are a few qualifications regarding the Orch OR hypothesis which should be mentioned. Firstly, monks meditating may attain a ‘higher state’ of consciousness without displaying higher frequency EEG waves. Meditation may therefore act to enhance a prior-existing gamma-synchrony, perhaps achieved as a result of their previous, frequently repeated experiences of consciousness. Secondly, the Orch OR hypothesis treats the mass of tubulin as one continuous entity in the form of MTs. In reality, MTs in axons and dendrites in toto provide the requisite mass (EG in the Orch OR equation), and do so by means of fabricating, out of a labile pool of tubulin dimers, a higher-level structure – the microtubular bundles which arise due to the association of MTs with MAPs. The Orch OR hypothesis, therefore, may depend as much upon a critical number of linker molecules, such as tau, as it does upon the number of MTs. Thirdly, recent research has demonstrated that glial cells are required for EEG gamma synchrony (Lee et al., 2014),133 a discovery that again falls outside the Orch OR scenario. But it may be that glial cells simply provide trophic or metabolic support to the neurons with which they connect due to the proposed Global Molecular Network.80
Relevance of Buddhist psychology to the study of consciousness
Hameroff and Penrose1,134 showed interest in Buddhist writings on consciousness because these texts not only reveal the results of 2500 years of practical investigation of consciousness by means of intense meditation and contemplation135 but they also comment upon human cognition and human nature. These results help draw a map of human psychological structures. The same goal has been pursued by Hindu yogins, whose discoveries are recorded in the Vedas and Upanishads.136
The Buddha, Siddhārta Gautama (b. ca. 563 BCE), specified a number of causal phenomena (the 12 Nidānas) associated with the arising of consciousness. One of these relates to the sensations perceived through the eye, ear, nose, tongue, body and mind. There are also the 5 Skandhas (the active and reactive functions of consciousness) consisting of touch, feeling, perception, volition and awareness.135 These causal conditions flare up in the brain moment by moment, serving as triggers for moments of consciousness, but happening so quickly that the mind is barely aware of them. The Buddha also touched on ‘ignorance’, which represents evidence of a disconnectedness from consciousness (in the Buddhist sense) or, in terms of Orch OR, a disconnection from quantum protoconsciousness.; or, in terms of Rosenberg's use of the term ‘protoconsciousness’,6 a dissociation between this property and cognition. The Buddhist practice of meditation is designed to pacify thought activities (thereby reaching state 3[1a−ᴗ1b-ᴗ2]) and to decrease ignorance, and thus produce perception of an unbroken sequence of conscious moments linking protoconscious events (the protoconsciousness of Rosenberg, mentioned immediately above) with consciousness via cognition. These moments prepare the Mind for entry into consciousness of state 4 (4[1a-ᴗ1b-ᴗ2ᴗ3]); here, it will be noticed, thought is not a requirements for consciousness.129 Ultimately, one aim of this preparation is to connect the mind with the Universal Mind, leading to its blending with the ultimate oneness of the Universe at the transition from state 5 to state 6.
As recorded by Hameroff and Penrose,1,134 Buddhists see consciousness as “momentary collections of mental phenomena” and as “distinct, unconnected and impermanent moments,” occurring so rapidly that they seem to form a continuum. At the heart of this proposal is the Buddhist concept of ‘momentariness’.32 As outlined by von Rospatt (ref. 32, pp. 21, 99), it is believed that there are 6,480,000 impermanent moments (Kṣaṇas) per day. Each moment would therefore be attributed a duration of 13.3 ms (≈75 Hz). Similarly, moments of 20 ms (i.e., 50 Hz) are proposed in some Chinese Buddhist texts. It is of significance that, as mentioned, EEG has recorded comparable frequencies of brain activity, and these have been correlated with a conscious state achieved by intense meditation.126,137 Hence, if we take the EEG 40 Hz gamma wave as an indicator of imminent quantum protoconscious events due to MT beat frequencies, then, as suggested by Orch OR, this would result in 3,456,000 moments of protoconsciousness in one day, each moment having a duration of 25 ms.
Another form of meditation, Zazen, practiced by adepts of Zen Buddhism, was one of the first to be studied by EEG.138 When the EEG outputs from Zen priests and disciples, who had varying degrees of meditative experience and proficiency, were compared, differences were found with respect to both their α and theta rhythms. As the ‘conscious’ state of the experienced meditators unfolded, the amplitude of their α rhythm increased while its frequency decreased (8–12 Hz falling to 7–8 Hz). At a later stage in the meditation period there was often the presence of a theta rhythm (6–7 Hz). These results were corroborated by Banquet137 and Murata et al. 139 and similar findings have been reviewed by Austin and others129,140,141 Probably, gamma EEG rhythms were not technically accessible at the time of some of these Zen meditational studies. Although α rhythms were not mentioned in the study of Lutz et al.126 on gamma rhythms, it may be that, within the meditators who participated in this study, these comparatively low-frequency rhythms were replaced by higher-frequency theta rhythms and then by gamma rhythms as the meditation proceeded. Indeed, Banquet137 recorded EEG rhythms of 40Hz from meditators who had entered a deeper state of consciousness, using transcendental meditative techniques, and Murata et al.139 reached a similar conclusion. An added possibility is that states of consciousness can be influenced by day-to-day variations of the Earth's geomagnetic field, as suggested by the results of Persinger and others142-144 who showed that this field affects the results of EEG and the quality of meditation. It is also possible that the various Schumann resonance frequencies of 8, 14, and 20 Hz within the Earth's ionosphere cavity145 may influence consciousness-related electrical activity within the brain, which itself generates similar frequencies, as recorded by EEG.
Coming now to a consideration of plants, Buddhists do not deny the possibility of plant consciousness. Indeed, the Buddhist view is that, there are (within the fifth Skandha of ‘awareness’, Vijñāna-skandha) 6 kinds of, or potentialities for, consciousness.135 On this basis, plants would be admitted to possess 5 of these attributes – of hearing, sight, smell, taste, and touch – but that they do not possess the sixth attribute of awareness, unless quantum computation, or some form of electrical activity, is considered as evidence of the sixth potentiality of consciousness, as will be elaborated when the idea of ‘oneness’ is discussed later. Nevertheless, the mentioned 5 attributes may be sufficient to qualify plants for protoconsciousness sensu Rosenberg.6
Moments of conscious awareness
The impermanent moments of protoconsciousness can be examined from another point of view. Entirely independently, and based on a study of time in relation to the Earth's movements around the Sun, Gerhard Dorda2 estimated a quantal unit of Earthly time to have a duration of 49.6 ms (20.2 Hz). The threshold values for the uncertainty of the duration and for the ordering of these time quanta were estimated as 24.9 and 30 ms, respectively. The uncertainty associated with the duration of each time-quantum results in frequencies of temporal duration ranging from 13.4 Hz to 40.5 Hz. When expressed in ‘moments’ of temporal perception during a 24-h day, the quantal-time durations lead to an estimate of 1,157,760 such moments, with an upper limit of 3,499,200 moments per day. These values are of the same order of magnitude as the ‘moments’ of consciousness described in Buddhist texts,32 the last-mentioned value being close to the number of quantal protoconscious moments (3,456,000) that correlate with the 40 Hz gamma EEG frequency. Given the correspondence between the lower limit of the frequency of quantal moments of time (13.4 Hz) and the rhythms of α waves from EEG (8 − 12 Hz) during Zazen meditation, it is suggested that meditative states allow an ‘awareness of the flow of time’, one moment of meditative self-awareness (or protoconsciousness) corresponding to one unit of quantal time, and occurring sufficiently frequently for this ‘self-awareness’ to be perceived as a continuous process. The upper limit of quantal time frequency (40.5 Hz) is similar to the 40 Hz frequency associated with meditative state of mind.126
Thus, a succession of quantal moments of time, when processed during meditation, deposits an impression of continuous self-awareness or ‘moments of consciousness’. ‘Sensitive awareness’ of state 3 (Mind) is therefore a state in which the passage of time is sensed. This state corresponds to what Bergson116 called ‘duration’ (durée). He says, “Real duration is what we have always known as time, but time perceived as indivisible … I am aware that we normally … have no interest in listening to the uninterrupted humming of life's depths. And yet, that is where real duration abides.” The state of “no interest,” which he mentions, is the usual characteristic of states 1 and 2 of Man. In state 3 and, more likely, in state 4, the “uninterrupted humming” of time (Bergson's durée or duration) is perceived as an uninterrupted ‘moment’ of authentic self-consciousness. It is a defining feature of these 2 states. As Michael Foley (ref. 146, p. 27) remarks, although “Bergson offered no practical advice on how to experience real duration – attendance to the humming of the depths sounds remarkably like the Buddhist practice of Ānāpāna-sati, respiration-mindfulness,” i.e., the rhythmic breathing and self-sensing of meditative practice, which can lead to the deeper meditative state of Samadhi (ref. 135, p. 152).
How could awareness of the passage of time come about and bring about a continuity of moments that construct Bergson's ‘duration’, or protoconsciousness? Dorda's estimation of the duration of a quantal time unit is linked to the gravitational force experienced on Earth in conjunction with the gravitational effects of Sun and Moon.2 Hence, mass was also treated by Dorda in a quantum manner; indeed, Hameroff and Penrose1,20,21 also introduced gravity into their Orch OR equation and thereby derived an estimate of 25 ms for the duration of a moment of protoconsciousness, which is very similar in value to Dorda's minimal estimate (24.9 ms) of a quantum of time.
Quantum units of both mass and time were considered by Dorda2 in relation to cellular growth in plants, a process dependent upon the movement of water into and out of cells. Cellular water was treated from the quantum perspective as providing quantal aggregates of mass. The temporal regulation of growth was hypothesized to involve quantum aggregates of water-mass being added to or subtracted from the main mass of cellular water during the passage of quantised time, this process being regulated by the orbits of Earth and Moon around the Sun. Recognition of the Moon's orbit and the concomitant variation of the gravity field, in particular, accounted for rhythmic increases and decreases of cellular volume in the plant system (tree-stem dilatation growth) which Dorda2 took for his working example. In multicellular systems, synchronous rhythmic cellular growth becomes amplified and is seen as rhythmic organ growth. Such rhythms are, in turn, manifestations of a lunisolar ‘clock’, evidence for which has also been adduced from the diurnal movements of leaves and other plant systems where rhythms of cellular water movement are involved.147,148 Nevertheless, these rhythms of movement trace back to quantal time units and quantal aggregates of cellular water mass; and it is the succession of these events – passage of quantal time coupled with movements of quantal water mass – which we postulate to be the clue to human (and other animals) and plant examples of sensitive awareness at the state 3 of Mind (or, more exactly, of state 3[1a−ᴗ1bᴗ2], reducing here the state of awareness to that of the soma) – fundamentally, it is the sensitive awareness of Sensorium (particularly the quiescent sub-state 1a− as interpreted by the Brain, state 2) to the passage of time. How this rhythmic quantal movement of water-mass could be translated, in terms of cellular biophysics, into neural impulses is an intriguing question. Could it be that it works in a similar way to what was postulated for plants – that it is related to ventricular volume changes of the cerebrospinal fluid? The volume of this component in various regions of the brain has been estimated149 but diurnal variations have not yet been studied (C. Nicholson, personal communication). It is through this volume that a second type of signaling (volume transmission, VT) takes place, which contrasts with signaling due to ‘wiring transmission’ (WT) by neurons. Like the phytoneural transmission of plants and the hypothesized movement of water aggregates, VT depends on exocytotic vesicles,150 and these vesicles themselves may transport water aggregates. It should also be remarked that Dorda (ref. 2, p. 102) estimated that, with the passage of each unit of quantized time, an aggregate of 6313 molecules of water are either added to or subtracted from an intracellular water aggregate of approx. 1 × 1013 molecules at a frequency of 73 Hz (every 14.7 ms), this passage being regulated by the timing of the Moon's orbit within the Earth-Sun complex.
Sensitivity to the passage of lunisolar time and gravity can be demonstrated by means of 3 examples of the activity patterns of a plant (bean leaf), a crustacean (crab), and a human recorded over long stretches of time in free-running conditions, where there was no contact with the external natural environment. Evident in Figures 1–3 is that, in each case, the temporal interfaces between 2 alternating periods – of movement and rest (crab) (Fig. 1), and of sleep and wakefulness (human, plant) (Fig. 2 and 3) – coincide with the times at which the lunisolar tide ‘turns’ (i.e., the lunisolar tidal force diminishes from a maximum high tide, or increases from a minimum low tide). This general pattern of a response to a turning point of the lunisolar tide is characteristic of many examples of both plant147 and animal (Barlow, unpublished) movements. The lunisolar gravity turning points correspond to times of day when the flow of water, either into or out of cells, is reversed.148 In all 3 examples of Figures 1–3, the turning points of lunar gravity have dramatic effects because they associate with abrupt changes of motor or growth activity of the experimental subjects. Nevertheless, it may be presumed that there is a constant subliminal awareness of this postulated inflow and outflow of water, and that a physical change in the direction of the flow, coincident with the lunisolar tidal change, corresponds with a change in biological activity. Each physical change, at the level of the cell, may be translated into a bioelectrical impulse. We propose that this sensation of the flow of time (duration), which may run in parallel with Orch OR events, is perceived as protoconsciousness of self during the integrated states 3 and 4 of Mind, when all other mental and physical preoccupations are quietened – at state 4 this would be notated 4[1a−ᴗ1b−ᴗ2ᴗ3] – by self-awareness and/or meditation.
Figure 1.
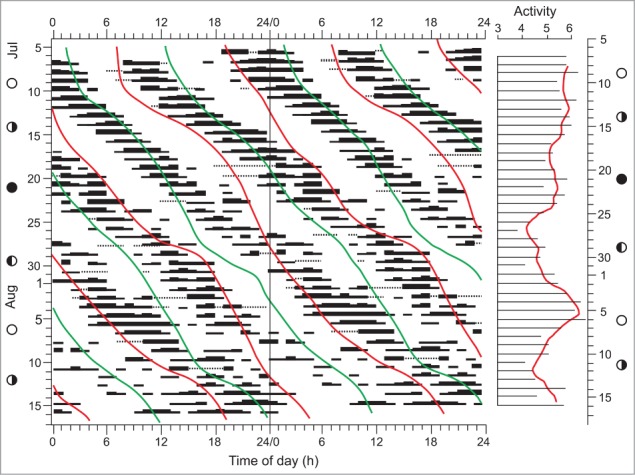
The perception of the passage of lunisolar time by a crab. Activity pattern (active period - black bars), recorded on an hourly basis over a period of 43 days during July-August 1963, of a single crab (Uca sp.) kept in constant low-level light in a laboratory at Woods Hole, CT, USA. The activity pattern drifts to the right, as though the activity time-keeper adheres to a 25-hour day. Superimposed on the activity pattern are the times of the contemporaneous lunisolar gravimetric high tides (red lines) and lunisolar low tides (green lines) for the location and dates in question. The transitions from inactivity to activity and vice versa are bounded by the times of high and low gravimetric tide (i.e., when each tide turns - commences descent from high tide or ascent from low tide). Lunar phases are shown on the left- and right-hand axes. Panel at the right-hand side is the √hours of cumulative daily activity of the crab, using a 3-d moving average. Activity is greater at Full and New Moon than at Quarter Moon. (Activity data from Fig. 4 of Barnwell151).
‘Primary perception’ and its link with consciousness
In the study of Lutz et al.,126 EEG gamma synchrony appeared in the course of meditation which directed the mind toward “unconditional loving-kindness and compassion” and “unrestricted readiness and availability to help living beings.” In the terminology of religious prayer, this meditative state might be regarded as an intercessionary state: that is, a state directed toward some image or memory. Early results of Harris et al.154 showed that prayers directed from Christian intercessors were effective in shortening the recovery period of hospitalized patients suffering from disabling coronary conditions. Since that time, other, generally positive, responses from intercessionary prayers have been documented.155 Positive results have also been reported concerning prayer directed at injured non-human primates (bush babies).156 More controversial (because of a lack of follow-up experiments) have been reports of plants being injured as a consequence of negative or malicious intentions emanating from human minds, their moods and their feelings.68,157
Results of intercessory activity may give evidence of ‘primary perception’ in the recipients of such intercessions or directed intentions. However, it is unknown whether an explicitly religious context (e.g., invocation of a deity) is a necessary condition for a successful intervention into a disturbed state. A state 3 consciousness of Mind may be sufficient to impart an at-a-distance quantum entanglement linking the intention of the directed, intercessionary Mind (of the healer) with the body and mind of the recipient (the one to be healed). Entanglements that resolve a bodily healing (or harming) process within the soma (sub-state 1a) of the recipient may be the means by which primary perception takes place, and could also be the route taken for healing through shamanic practice.158
Prayers for altered interpersonal relationships may be another area where state 3 consciousness of the intercessor can effect an emotional or psychological response.159 But in this case, where, for example, forgiveness of one party (the violator) by another (the violated) is prayed for by the latter party (who is also the intercessor), it is more likely that the reconciliation occurs by primary perception at the level of emotional self-awareness (sub-state 1b) and would pertain to the intercessor (the violated) him/herself. Whether intercessionary transfers by entanglements of consciousness would be more successful if gamma-synchronized, self-conscious state-4-embodied Minds were involved is an interesting question.
The above examples referred to perception at-a-distance. However, with regard to plants,68,157 the described cases of primary perception were ‘at-close-quarters’. Here another possibility presents itself: that of Mind as a force-field.160 A possible effector of this mode of perception is the electromagnetic field generated by the vast assembly of electrically conducting neurons of the brain. As pointed out by Nagel,24 who reviewed a number of relevant publications, plants are susceptible to intercessory prayers, or ‘friendly thoughts’. She suggests that “prayer can be considered a form of mind power,” in the sense discussed by Lindahl and Århem,160 although she does not mention this particular publication. She also comments that “if ‘primary perception’ did exist in plants, this would not necessarily prove that plants are conscious.” The results of prayer would simply be evidence of a susceptibility of some sort toward the mind that formed the prayer. However, for susceptibility of either plant or animal to be recognized by an observer, there would be the need for its translation into an observable movement (growth) and/or physiological change. A directed meditational technique (‘thought transaction’) invoking “wellness of the crop and good yield” was reported to bring about an enhancement of okra (Abelmoschus esculentus) plant growth161 and resistance to nematode infection. The transaction involved the preparation of a mental state at which theta frequency (4 – 8 Hz) EEG waves became manifest. It was maintained, in the presence of the plants, for 5 min each day for 60 d. At harvest, okra crop yield, in kg, of the ‘thought transaction’ group of plants was enhanced by approx. 20% compared to ‘non-transacted’ controls.
Even more intriguing are the observations of JC Bose162 who recorded minute changes in the growth of wheat coleoptiles (using a ‘crescograph’ of his own invention, which magnified growth by × 50 × 106) each time a controlled electrical discharge took place. The discharges took place at a location 200m distant from the test plants. Discharges were fed to a transmitting aerial and were recorded by the plants, which were linked to a receiving aerial. The claimed ‘wireless’ communication indicated that plants could theoretically “respond to the long æther waves, including those employed in signaling through space.”162 Bose was careful to distinguish between the internal and external effects of these wireless waves on the receiving plant, the former effects manifesting as growth movements whereas the latter had no such outward manifestation but consisted of, so Bose believed, a chemical change with a concomitant increase of potential energy.
The occupancy of Mind at state 3 may be a means by which humans can ‘communicate’ with plants, and vice versa. It may also play a part in inter-plant communication as, for example, in phenomena such as kin recognition,163-166 one of topics which has also been central to discussion of plant cognition.29 Kin recognition may trace to the consciousness criterion no. 12, ‘subjectivity’, where kinship is the source of a sensation private to a given plant. It, too, may be a situation whereby quantum or electromagnetic effects achieve communication over long- or short-range distances, with observable consequences. Primary perception may also be the means by which animals communicate and show their concern for one another, and how plants communicate warning signals to each other, though this latter may also involve volatile chemical signals.167 Such examples seem to be forms of sensitive awareness of one being toward another that transcend mere instinctive responses.
The idea of plant ‘oneness’
Plants, by virtue of their open and branched pattern of primary growth, are continually emitting new parts and, hence, fragmenting their individuality, particularly when engaged in vegetative reproduction – by emitting suckers, for example, which eventually assume their own individuality. Branching occurs not only externally by budding but sometimes also internally, by splitting (schizogeny) of tissues as well as of organs. Organ splitting affects the form of trees when, for example, an aged trunk splits and forms second-order trunks. In this way, multiple copies of new individuals (clones) are formed, which eventually supplant the original first-order trunk. Nevertheless, in plants, there may be a contrary striving: for integration, or ‘oneness’, to compensate for this branching of organs and the tendency to fragment. The fusion of aerial roots, typical of some trees (those Ficus spp, for example, whose intertwined roots form an aerial pseudo-trunk), may be an expression of such a tendency toward oneness, and which follows on from an earlier phase of root and shoot branching. The gathering together of parts that were once separate is similar to what occurs in swarming or flocking events among insects and birds.168
It is as though, by means of an internal act of primary perception, the plant or tree has a sense (or consciousness overseen by a Mind at state 3), not only of its parts, but also of its form; and through this self-awareness of form arises the means whereby a plant's own oneness can be attained. An aspect of this tendency toward oneness derives from the property of whole-plant ‘correlation’.169-171 It is the means by which not only the fundamental architectural form (or model) of the whole is realized during undisturbed development172 but also is regained after partial dismemberment. In each case, information is communicated within and between the various entities at their respective organizational levels. However, the correlative process also operates internally, and also at more restricted levels, within organs and tissues, enabling the maintenance of their respective integrities.173 The necessary communicative processes are due, in part, to the movement of plant growth regulators – auxin, for example, which has neurotransmitter-like properties, and is probably also assisted, or mediated, by MTs and the cytostructural code. Presumably, in order to re-establish the correct architecture (after an injury, say) the plant titrates the entire system for hormonal composition and quantities, as well as making use of the bioelectrical code,67 part of which involves electropotential difference (EPDs) between growing parts. These procedures re-establish the balance of the various morphogenetic components until equilibrium is reached in the meristems and their immediate derivatives: that is, there is a restoration of the optimum sense of ‘oneness’.
A second aspect of ‘oneness’ is that plants are only partially compartmentalized into cells and organs; the totality of their cells in fact forms one continuous, cytoplasmic network.174-176 This intercommunicating, symplasmic system has few if any insuperable barriers to an internal ‘oneness’.
Another aspect of ‘oneness’, and one which includes the plant's discovery of its sense of ‘sameness’ as well as its sense of ‘otherness’, may arise, not from the plant's interior but from the organisms which surround it. This could be through inter-organ primary perception, or by means of communication mediated by electrical fields and possible quantum entanglements, as well as by chemical means – elements of a type of ‘kin recognition’, but a type that operates at the level of the organism and, again, utilizing the cytostructural and bioelectrical codes. This possibility, that use is made of internally generated bioelectromagnetic properties, has many ramifications in relation to plant communities of small size, and has been suggested as a means facilitating seed germination and seedling growth.177,178 It is anticipated that other unknown but mutually influential factors may also operate between plants.164 If so, then plants may be able to direct their own ecologies through plant-plant interactions more subtle than those due to either allelopathy via secreted chemicals or restrictions on living space brought about by nutritional preferences.
Do plants have a nervous system?
Nervous systems in many ‘higher’ animals are characterized by specialized cells which propagate action potentials (APs) as a consequence of membrane depolarization and the movement of ions into and out of neurons. Such systems involve electrical synapses (gap junctions) and chemical synapses facilitated by neurotransmitter substances. There are approx. 60 types of neurotransmitter molecules in humans, including acetylcholine, amino acids, and biogenic amines; MTs support and guide the vesicles in which these molecules are contained toward the inter-neuronal synaptic junctions.
Action potentials in plants were first recorded in a higher plant (Mimosa)179 and in a green alga (Nitella),180 long before their discovery in animals. If algae and higher plants have a ‘phytoneural’ system which is similar to any of the various animal nervous systems, then not only should APs be expected, but plant anatomy might also reveal cellular structures and ultrastructures (such as exocytotic vesicles and MTs), as well as specific molecules which have been adapted to participate in the development of APs at cell boundaries. It is believed that plants do indeed possess ‘plant chemical synapses’, and that these share many of the features of chemical synapses found in animals.181 As for the identity of the phytoneural cells in higher plants, anatomical and physiological evidence indicates that they correspond to the phloem strands, and that these strands propagate APs over long distances,72,182,183 as also occurs within the giant cells of green algae. Also, numerous molecules have been discovered in plants36,184 – for example, gamma-aminobutyric acid (GABA), melatonin, serotonin – which, in animals, serve as neurotransmitters. One of the roles of these molecules in plant cells is to modulate MT stability;59 and it because of this property that they affect neurotransmission in animals.185 Certain other molecules found in plants can also modulate animal neurotransmission – for example, hyperforin, which increases acetylcholine release from cerebral tissue,186 and some secondary plant products are potent animal neurotoxins and hallucinogens – nicotine, curare and psilocybin, for example,187 all of which affect the acetylcholine receptors and the passage of electrical currents via chemical synapses. Inhibition of these last-mentioned receptors is also a feature of certain anesthetics.188,189 Thus, plants do indeed possess structural and chemical elements which could participate in a putative phytoneural system, though at present there are no details of how they do so.
Plants, because they are encased in cellulosic walls, are typically sessile organisms. Their physical movements are measured in days rather than in seconds, as would apply to animals. If plants possess an analog of the animal-type nervous system, it may either function slowly – to alter the orientation of an organ by differential growth, or more rapidly – by activating molecular transport mechanisms which can then lead to an alteration of physiological state. The nervous reactions involved initiate an immediate electrical response, ion exchange being followed by cascades of other signals; the latter then propagate relatively slowly throughout the organism. The animal nervous system which perhaps most closely resembles that of a plant is the one expressed in sponges,190 where GABA is used as neurotransmitter.191 Sponges belong to a basal phylum of the Animalia and possess ‘protosynapses’;74 these seem to be similar in function and structure to the plant chemical synapses proposed by Baluška et al.181
The different nervous systems found in sponges, animals and, putatively, plants, indicate that nervous systems have probably evolved many times during evolution, as was also proposed for brains73 and, indeed, was also proposed for consciousness itself.12 What critics of the notion of plant neurobiology have overlooked (due, probably, to an anthropomorphic orientation) is the diversity of nervous systems and neurons within the animal kingdom. Furthermore, Edelman et al.12 draw attention to the finding that the octopus (which these authors credit with both cognition and a precursor of consciousness) has more neurons in all its tentacles than it has in its brain, suggesting that some attributes of ‘consciousness’ might reside elsewhere than within a brain. As Moroz73 remarks in responding to his own question “What is a neuron?,” it is that they “can make polarized and specialized (synapses) connections, but do not necessarily do so in all animals and nervous circuits.” Moroz74 continues, “Hormonal-like volume transmission [see earlier] can serve many true integrative and neuronal functions without a specialized synapse … if targets are localized within a few micrometers from the transmitter release points.” The author is referring here to nervous systems which are used by “sessile animals with limited motor reactions or for vegetative processes.” Such a description for certain animal nervous systems corresponds with the situation in plants. The mention of volume transmission suggests that plants, too, may have a ‘Global Molecular Network’,80 which could be constituted of both a WT system (bioelectrical circuitry) and a VT system (phytohormonal and growth regulator mobility, including microRNAs), which together would help sustain the global inter-cellular and inter-organ communication system needed to maintain a plant's sense of ‘oneness’ (see the preceding section). Thus, no special pleading is needed for a ‘phytoneural system’. What has been discovered so far is that its features lie within the range of diversity of neural systems in toto.
One specialized site on the cell membrane of animals across which an electrical current can flow is the gap junction. These may be important in regulating bioelectric fields, which, in turn, appear to regulate the correct spatial distribution of cellular groups in embryos and elsewhere in the soma.192 Possible plant analogs of the gap junctions of animal cell membranes are the cell-cell channels known as plasmodesmata.193,194 These structures are inserted, in the first instance, into new cell division walls. Secondary plasmodesmata are sometimes inserted at a later time into particular pre-existing cell walls of differentiated tissues.195 These secondary plasmodesmata may be analogs of the recently discovered tunneling nanotubes which link cells together via adventitious cytoplasmic bridges.175,195 Plant parenchyma cells have the ability to propagate electrical impulses through the plasmodesmata and, hence, through the symplasm until they reach non-excitable neighboring cells.196,197 However, the current so propagated is strongly damped due to a bottleneck effect exerted by the plasmodesmata themselves, thus limiting the range of the electrical signals to only a few cells.196 This result was obtained from root apical meristems of the fern, Azolla pinnata, in which plasmodesmatal abundance, densities, and cell-cell distributions have been carefully mapped. Even in giant shoot apical meristems, such as those of the Cactaceae, which have possibly tens of thousands of cells, the scale of coherent electrical signaling would be limited to a small number of cells at any one time. It follows that, in both shoot and root apices, Orch OR events and the subsequent propagation of bioelectrical impulses would occur only slowly and possibly infrequently.
Orch OR suggests that EEG gamma synchrony occurs by means of the coordinated activities of MTs, synapses and gap junctions.198 Halothane and ethylene, anesthetics which work as such on animals, destabilize MTs in growing plant cells199 although, unfortunately, little is known of any putative phytoneurological effects of anesthetics save their ability to alter the sensitivity of plant cells to cold stress,200 an effect probably mediated by MTs. Interestingly, ethylene brings about the rearrangement of MTs in growing cells of dicotyledonous plants, thus enhancing the lateral expansion of the respective plant organs.201,202 The newly formed growing portions of an ethylene-treated shoot or root therefore remain squat, as though ‘resting’ and waiting for conditions to ameliorate before resuming rectilinear growth. Because ethylene is released from plant organs following wounding, this ‘resting’ response may be some form of self-anesthesia.
Does Orch OR apply to plants and, if so, could plants be conscious?
The Orch OR hypothesis, with its reliance upon MTs to facilitate the firing of electrical nervous impulses, has many favorable points for an understanding of the origin of human consciousness and, as has been described in earlier sections, recent experimental results have provided circumstantial evidence for its applicability to consciousness processes. In animals, MT arrays are crucial for electrical signaling within and between neurons whereas, in plants, the corresponding electrical properties and their role in intercellular signaling are imperfectly known. Although Baars and Edelman47 were doubtful that MTs, whether in animal or plant cells, could be a sufficient basis for ‘quantum consciousness,’ the Hameroff-Penrose Orch OR hypothesis does nevertheless suggest that quantum OR events provide a potentiality for moments of protoconsciousness. It should follow that all such moments are liberated by OR events, and that their frequency would depend on the number and density of MTs. On the grounds of this last-mentioned prediction, plants, which have no particular type of cells that are directly equivalent animal neurons in terms of their degree of enrichment with MTs, might in this respect be disadvantaged in their attainment of consciousness.
Actin microfilaments which, in addition to MTs, are the other major class of structures of the plant cytoskeleton, should also be considered with respect to their electrical capabilities. In a recent study where a 50Hz oscillating electrical field was imposed on plant tissue, permeabilization of the tissue was brought about in a manner dependent upon the presence of actin, suggesting that actin filament oscillation and/or cytoplasmic streaming may be allow the opening of the plasma membrane,203 which possibly may allow the initiation of APs.
In the preceding section, primary meristems were considered to be rather poor candidates for OR events on account of the lability of their MT population, and then the short cell-to-cell distances over which electrical impulses could be transmitted via plasmodesmata, to promote moments of protoconsciousness in some brain-like center. The situation is different in tree trunks, however, where there is a secondary cambial meristem from which secondary, or widening, growth is established. Cambial cells are not only extremely numerous, covering the entire cylindrical surface immediately beneath the bark, but they and their derivatives are also replete with MTs which participate in the development of both woody tissue204 and secondary phloem. The MTs of these cells have more stable configurations than those of meristems. There are also groups of parenchymatous ray cells, which intrude into secondary phloem and secondary xylem (Fig. 4A) and which are rich in MTs. Unlike the immediate derivative cells of primary meristems, derivatives of ray cells remain for a long time close to their site of origin, do not divide, and live for at least 4–5 years. MTs of ray cells appear bound together as microtubular cables205 (Fig. 4B). Ray cells also contain prominent cables of actin filaments (Fig. 4C and D). Additionally, there are abundant, closely bound helices of MTs lying against the plasma membrane of developing secondary xylem cells, as well as circles of MTs at the pit borders of xylem vessels.206 Similar findings apply to the secondary cambium and vascular tissues of woody root systems.207 The numbers of MTs in such cells and tissues has not been estimated, but considering the total volume of tissue present in a mature tree, these numbers would be prodigious. However, an approximate number of MTs can be reckoned, as shown below.
Figure 4.
Ray parenchyma cells within secondary stem tissue of trees. (A) Vertical columns of rays (spindle-shaped groups of small cells) located amongst elongated fusiform cells within the stem cambium of a two-year old hybrid poplar tree. Toluidine blue-stained semi-thin tangential section. Mag. × 280. (B) Radially oriented bundles of MTs in vertical stacks of ray parenchyma cells from Abies sachalinensis. Anti-tubulin immunofluorecent image from a radial section visualised by confocal microscopy. Mag. × 1500. (Micrograph modified from Begum et al.205). (C, D) Radially oriented cables of actin within rays of a two-year old hybrid poplar tree. Anti-actin immunofluorescent image from a radial section visualised by confocal microscopy. Fluorescent images of actin filaments running vertically in c are due to their presence in overlying fusiform cells of the cambium. Mag. × 1200 (C), × 450 (D). (Micrograph modified from Chaffey et al.206).
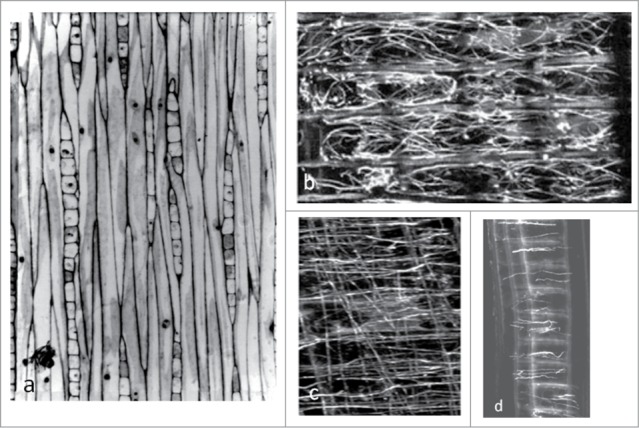
Ray parenchyma tissue is composed of up to 8–16 or more tiers of cells (Fig. 4A). Tangential cuts through the cambium also reveal that the ray complexes are organized in a helical pattern along prominent parastichies winding around the cylindrical cambial surface. Thus the secondary xylem and phloem tissues beneath the bark of a tree trunk have embedded within them a lattice of radially intrusive rays (Fig. 5). Because most of the ray cells no longer divide and their differentiation is rapidly completed, their MTs would be available for new functions and might provide the conditions for Orch OR events. Moreover, all ray complexes traverse the cambium and could thus feed signals transmitted radially, by the rays, into the longitudinally oriented cambial system which extends into the apical and basal extremities of a tree. The density of ray clusters shown in Fig. 5 is estimated as approx. 70 per mm2, a value which might offer possibilities for amplification of MT-based OR events, particularly since the lattice configuration of the rays might itself provide a basis for the mentioned amplification by introducing an extra dimension for the postulated quantum computing required by Orch OR. The large total number of MTs collected into such a lattice encircling the trunk of a large tree could therefore be a powerful structure, analogous to a population of MT-bearing neurons within a brain, for the triggering OR events. Hence, ray tissue could be a location for propagating moments of protoconsciousness. In this way – through MT density and large number of ray cells within a lattice arrangement, where quantum computing and quantum entanglement could take place – the potential limitation would be overcome of not having one particular cell type, like a neuron, or tissue especially enriched by MTs, like a brain. The entire trunk of a tree filled with rays and their MTs could be regarded as a candidate plant brain (on a large scale), where protoconscious events might be produced by Orch OR.
Figure 5.
Distribution of rays, such as shown in Fig. 4A, recorded in a large photomontage of tangential sections of hybrid poplar cambium. The montage was scanned and the vertical height was foreshortened. The spindle-shaped ray areas were then traced (stippled regions). Clear areas contain fusiform cambial initials. Sometimes rays had recently undergone binary fission due to intrusive growth of fusiform cells; the two sister rays are outlined within a single oval. Ray cell complexes align on 3 evident parastichies (red lines). Occasionally, a parastichy terminates due to the introduction of a new ray complex arising from fission of a pre-existing ray complex. Circumferential expansion of the cambium accompanied by growth and division of the fusiform cells lying between the sister ray complexes and elsewhere causes the ray complexes to move apart.
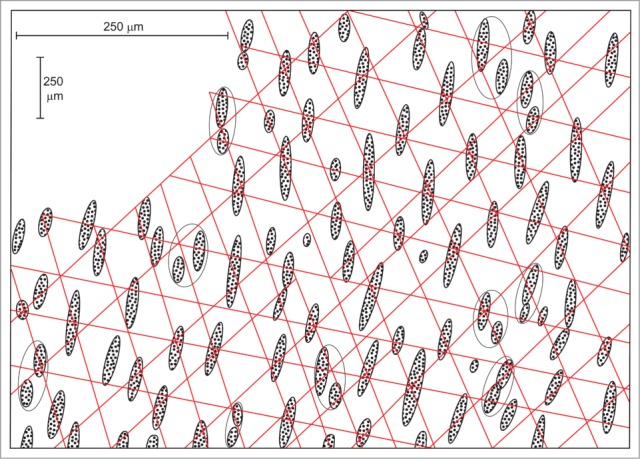
MT-bearing rays embedded within tree trunks with their potentiality for OR events may be the reason, at a subliminal level, why sensitive persons wish to ‘hug a tree’: it is an indication of the resonance, or quantum entanglement-at-a-distance,45 of tree and human MT-beat frequencies emanating from both the human neuronal and the putative ‘phytoneuronal’ channels of the ray system, perhaps coupled also to bioelectromagnetic field effects between the tree and the human brain and heart. The dual tree ray-human MT system could be another situation for primary perception, and thus be the site of something similar to kin recognition mentioned earlier. Although it is not known whether the gamma synchrony of the type recorded by EEG from human subjects, and which is associated with consciousness, is present in plants, the cambium and its ray and secondary tissues of trees may be a good place to start to search for such synchrony.
Quantitative estimates of tubulin and MT characteristics in the trunks of young trees of hybrid poplar clarify the potentiality of rays for generating Orch OR events leading to moments of “tree protoconsciousness.” Data on ray density were obtained from images, such as shown in Figure 5, and additional quantitative values for the volumes of ray tissue in secondary xylem and phloem are taken from Barlow et al.208 The estimates are as follows:
Figure 5, which is typical of a poplar tree trunk seen in tangential section through the cambial zone, reveals that there are 85 ray complexes inserted within a cambial surface of 1.2 × 106 μm2.
Single, uniseriate ray complexes are approx. 250 μm high (in the vertical dimension), 14 μm wide and, in one annual growth ring of xylem and phloem, approx. 4610 μm long (in the radial dimension). Rays complexes of poplar are viable for about 4 years;209 in the fifth year following their birth, the cells die. All told, therefore, the volume of one typical complete living ray complex is approx. 6.5 × 107 μm3.
A microtubule of length 1 μm consists of 1600 tubulin dimers.210 Therefore, a single MT, 4610 μm long (i.e., the total length of one typical living ray complex), consists of 7.4 × 106 dimers. A conservative estimate that 100 MTs extend the length of such a ray complex yields a total of 7.4 × 108 dimers.
Ray complexes are multicellular, composed of one vertical file of approx. 8 cells. Files of each of these 8 cells extends in the radial direction. Hence, there would be approx. 5.9 × 109 tubulin dimers in the set of 8 radial files of one typical single ray complex.
Hameroff and Penrose state that a single cerebral neuron contains approx. 109 tubulin dimers. In terms of tubulin mass, one complete ray complex is equivalent to a single neuron.
According to the Orch OR hypothesis, a quantum superposition of duration τ ≈500 ms requires 109 tubulin dimers. Thus, a single, 3-dimensional ray complex may be able to supply the necessary tubulin mass for an OR event at intervals of 500 ms (2 Hz). Many ray complexes working together in their lattice arrangement may increase this frequency.
One proviso with respect to the ray system is that in order for the ray MTs to behave analogously to a single animal neuron with 109 tubulins, continuity of MTs should be maintained throughout all the cells of the ray complex, both vertically and radially. MAPs such as tau associated with MTs, as well as plasmodesmata which support the cellular symplasm, may provide the necessary degree of quantum coherence.
If such a system were capable of orchestrating OR events in the way proposed by Hameroff and Penrose,1 then there is the question of how the resulting moments of protoconsciousness would be registered in the ‘mind’ of the tree. Of course, this is also a problem for bona fide systems of animal consciousness. Furthermore, one might ask whether the stream of sensory inputs to the plant from both internal and external sources can be integrated by means of Orch OR. Thus, for a plant or tree, there may also be the ‘binding problem’ of how to establish a unification of conscious perception.35,37 But this might not be such a vital problem for a tree, as it is for animal/human systems, where it is essential for their survival, for example. And, as was mentioned earlier, thought (as supported by a brain) is not a necessary component of consciousness.
Alternative modes of consciousness
Because there are different types of brains and nervous systems among animals, as well as different modes of consciousness,13 the approach to the question of whether plants can be considered ‘conscious’ may be made only in an analogical way using animal systems as a reference. However, the way becomes clearer when consistent attributes of consciousness are recognized – albeit using attributes derived from studies of animals, both human and non-human. Earlier, a structured series of conscious states was provided, and it was deduced that, in the human situation, and also in the light of Hameroff and Penrose's Orch OR theory of protoconsciousness,1 whenever neuronal vibrations of lower frequency were, even if momentarily, replaced by vibrations of a higher frequency (theta and α frequencies of 4–7 Hz and 8–15 H, respectively), and thence to gamma frequencies (of 40–100Hz, for example), there may literally be a ‘quantum’ leap into a deeper level of reflective-, or self-, consciousness. This might be due to OR events in the Brain apparatus (state 2) somehow becoming registered within a Mind (states 3 and 4). For plants, the corresponding situation, especially the final step of registering consciousness, is nearly unknown, but some clues regarding the preceding steps can come from studies of action potentials (APs) as indicators of electrical activity.211 Quantum effects at ion channels have been proposed to induce APs,19 and the electromagnetic field generated by the multiple APs of cerebral neurons has been advanced as a component of conscious awareness.212 Action potentials and their relevance to possible plant consciousness will now be explored further.
Interior protoconsciousness: An electrical basis
In plants, besides the presence of electropotential differences (EPDs) across organs, there is continual transmission of electrical action potentials (APs) via vascular tissues. For example, whenever sap flows within living phloem cells,213 or whenever there are changes in hydrostatic pressure within xylem vessels,214 there is excitation of membranes which then generates APs and sets up EPDs between reference sites along the organs concerned. Moreover, the arrival of APs at certain destinations, or target zones, is likely to promote distinct patterns of gene activity,215,216 as well as cascades of secondary messengers, such as Ca+ ions, and the stimulation of phytohormonal transport, and thence lead to various motor activities. Actin microfilaments and actin cables within ray cells (Fig. 4C and D) and elsewhere may also convey sufficient kinetic energy to disturb cell plasma membranes and induce APs.
Internally, plants are rarely quiet, except at night and under relatively still conditions.213 Environmental stimuli – light, temperature, salinity, drought, waterlogging, contact with predators, gaseous signals released from sites of infection in neighboring plants, even the passage of clouds across the sky – can all modulate phytoelectrical activity. Because of the incessant input of signals from the environment, as well as the widespread internal signaling between organs and modules, plant systems are ‘alive’ with electrical activity. Thus, in addition to local brain-like areas (root apex transition zones, e.g.,), the whole plant, and especially the trunks of trees, can be viewed as a distributed ‘brain’, in the sense of ‘brain’ being a center for the integration of electrical activity. This distributedness contrasts with the situation in higher animals (but may correspond to the already mentioned situation in sponges) where electrical signaling is channeled toward a localized and integrating central nervous system and brain. Then, the phytohormonal correlative system, which is able to initiate appropriate morphogenetic responses to impulses originating from a diffuse ‘brain’ or some other localized command center75,76 and thereby regulate the structural form of the organism, provides the plant with a sense of self, or of ‘oneness’ (state 3). This may be the nearest we can come to resolving the question of whether (or not) plants are conscious, with an awareness of self and of others.
One further aspect of plant electrical activity should also be mentioned. This continual activity, due to the plants’ continual responsiveness to their environments, masks another, more basic aspect of phytoelectrical activity inherent to the structure of plants. Such an intrinsic and basal electrical state should be discoverable when the environment is totally stable. Then, quantum reduction events, perhaps due to MTs, or events due to actin filament kinetics, should become apparent. Hence, it is of interest to estimate the frequency (in Hz) of APs during resting periods, when plants are free from perturbation. A survey of APs recorded from base-line, unstimulated states approximating to ‘free-running’ conditions allows the following estimates of their resting frequencies to be made.
The simplest case in this survey is the trichome of the blue-green alga, Phormidium. These cells show no electrical activity (0 Hz) except when they are transferred from light to darkness; then an impulse is recorded, coincident with the transfer.217 From the giant unicellular green alga, Acetabularia, spontaneous APs with frequencies of 8 × 10−4 Hz and approx 2.3 × 10−3 Hz were recorded from cells in a quiescent state by Thavarungkul et al. and by Saddler, respectively.218,219 At the developmental stages studied, MTs would have been either absent or sparse.220 Similarly, APs of 3 × 10−2 Hz were recorded from the giant cells of the green alga, Nitella mucronata.180
In non-vascular and vascular plants, spontaneous APs, sometimes in long trains of activity, arise without any known cause. Records from thalli of the liverwort, Conocephalum conicum, showed electrical discharges with frequencies of 2–3 × 10−3 Hz.221 Although these signals were recorded sometime after a wounding event, they had settled to a constant frequency and amplitude and the tissue appeared to have recovered from the trauma when these estimates were made. From growing stems of Helianthus annus, spontaneous impulses with frequencies of 1 × 10−2 Hz were consistently found;222 and from growing portions of stems of Dianthus sp. (where cytoskeletal MTs and actin filaments would be abundant) spontaneous APs with frequencies of 0.2–0.25 Hz were recorded by Glębicki et al.223 However, a frequency of only 0.08 Hz was found in the non-growing, basal zone of the stem, where the cytoskeleton might be expected to be less developed. From various zones of a small tree of Ficus elastica, the highest electrical impulse rate, recorded during a period of many weeks, was 3.3 Hz.224 Unfortunately, the exact locations of the recording zones were not specified and, moreover, the frequencies varied from minute to minute: one zone had a frequency of 0.8 Hz, another of 1.7 Hz, while a third zone showed 1.7 Hz one minute and then 0.3 Hz during the next minute. Nevertheless, the impulses were a constant, unprovoked feature of recordings from the whole tree. Pickard225 studied APs from petioles and lamina of cotyledons of Ipomoea hederacea. Successions of spontaneous electrical impulses were found, often persisting over long periods of time, the longest period recorded lasting 44 min; the average frequency of these impulses was 0.95 Hz. Williams and Pickard226 recorded impulses of approx. 0.12 Hz frequency from tentacles of Drosera intermedia, when the material was apparently in a stable, free-running condition. It is interesting that in both the study of Karlsson224 and Zawadski et al. 222 diurnal rhythms of the impulses were noted, but these were probably related to the daily changes from light to dark conditions. Nevertheless, they indicate that plants can be aware of the time of day through the frequency and amplitude of their APs.
From roots of Lepidium sativum, Hejnowicz et al.227 registered electrical signals of 2–5 Hz during the first minute of one typical recording session, and which diminished to 0.3 Hz in the next minute. The most coherent signals were found in the apical meristem and just behind it, where the transition zone would be located and where the cytoskeletal MTs would be densest. According to the authors, gravistimulation of the root did not affect the fluctuations of the APs “in any reproducible way.” However, exposure of the roots to both N2 gas (anoxia) and ether vapor reduced the frequency of the impulses. The effect was reversed by flushing the gases from the system with clean air. Both these gaseous agents disassemble MTs in a reversible way.228
Notable is that all AP frequencies were relatively low. Furthermore, APs in plants have relatively long refractory periods; and if multiple impulses were to arrive from different locations, this would tend to confuse any evaluation of an average frequency at the recording site. Nevertheless, the consistency with which a low level of electrical activity is encountered in non-stimulated plants is probably significant. It suggests a degree of torpor, or baseline sensitive awareness; nevertheless, the plant can be aroused from this state when appropriate stimuli are provided.
Other types of electrical signal214 are more frequent when plants are appropriately stimulated – by wind action, for example, which is able to induce volume potentials due to hydrostatic variations;229 and EPDs become evident when trees photosynthesise, transpire, and translocate sugar solutes.213 Thus, there may be 2 periods of protoconsciousness for trees and plants: one during rest within the hours of darkness, the other during wakefulness and consequent upon environmental stimuli, which are more abundant during hours of daylight. Activity patterns in the form of EPDs are also modulated by lunar phase230,231 in the same way that activity patterns of animals are affected (see Fig. 1 and 2).
Exterior protoconsciousness
One topic not discussed by Hameroff and Penrose, believing it to be outside the scope of science, is the possibility that protoconsciousness might occur in non-biological systems, including the cosmos itself.1,45 This they did not discuss. However, as JBS Haldane wrote, “I have no doubt that in reality the future will be vastly more surprising than anything I can imagine … my own suspicion is that the Universe is not only queerer than we suppose, but queerer than we can suppose.” Thus, it can be surmised that whatever topic has been considered outside the scope of science in one era (e.g., theories of planetary motion and of gravity, wave-particle duality, dark matter) might enter the scope of science in a succeeding era. Therefore, one might touch on this unknown issue of a non-biological origin of protoconsciousness,1,45 partly because Hameroff and Penrose entitle their article, “Consciousness in the Universe” and then continue (see their abstract) “we conclude that consciousness plays an intrinsic role in the universe” (our emphasis). While one may puzzle about what this “intrinsic role” may be, protoconsciousness can be imagined to exist independently of a biological context (as indeed Hameroff and Penrose speculated,1 but then in the context of massive neutron stars), and that it is an imminent property within the Universe due to physical laws; it is as though scintilla of protoconscious could theoretically exist and await capture by biological organisms who have the ability (or wish) to perceive them. For example, the Schumann resonance frequencies145 within the Earth's ionosphere correspond with frequencies recorded from the brain by EEG, but whether the Schumann frequencies intervene in such electrical activity and, hence, in consciousness, is not known.
Three vital questions surround the OR event associated with consciousness in living forms: 1) Is a protoconscious event generated only by MTs? 2) Can it be that certain groups of MTs within a biological organism attract to themselves a precursor (scintilla) of protoconsciousness from a universal pool of such events which reside in the cosmos, and then relays and amplifies it to consciousness via the neurons? 3) Are biological/quantum-physical protoconscious events simulations of analogous events within the cosmos?
As mentioned in section 10, a perception of self and an awakening of activity in the soma emerge at the turning of high and low lunar tides (Fig. 2). This awareness, this subliminal consciousness of self, is thus the consequence of the soma and brain being touched by an interaction with the cosmos – the relationship between the organism and the gravitational attraction between Earth, Moon and Sun. In this regard, where Organism-Earth-Moon-Sun forms a unit, a fully developed, continuous awareness of organismal self might be considered to be an answer to a further, fourth question: Can consciousness in a living organism arise from some physical feature of the cosmos?
Figure 2.
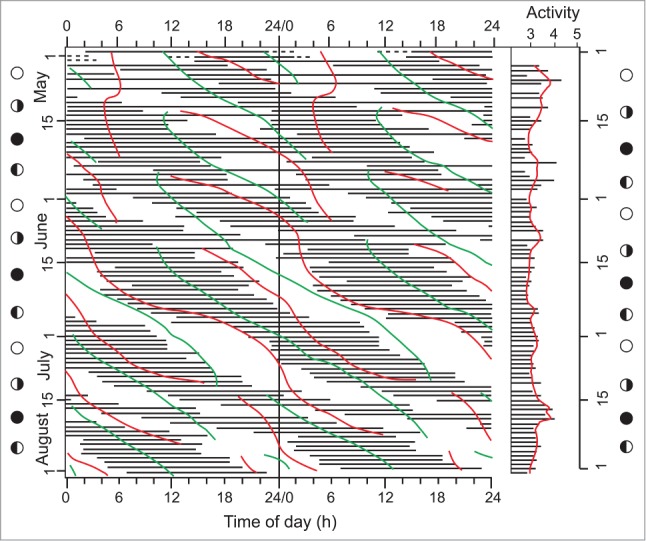
The perception of the passage of lunisolar time by a human. Activity pattern from a human subject in the total darkness of a cave at Cheddar, UK, over a period of 127 days during April-July 1966, and without any knowledge of the chronological time in the outside world. The sleep (black bars) and wakefulness periods were recorded each day with a resolution of 1 h. Superimposed is the contemporaneous lunisolar tidal pattern (red and green lines, as in Fig. 1). The sleep-wakefulness periods are bounded by the turnings of the gravimetric tide. Lunar phases are shown on the right-hand axis, together with the √hours of wakeful activity (right-hand panel, as in Fig. 1). (Sleep/awake pattern data from Fig. 8 of Mills et al.152).
At present, however, the quantum-physical protoconscious events of Orch OR can be usefully discussed only at the human/biological level, mainly because discussions of consciousness center upon human consciousness. But a more general, or even a more universal, aspect is suggested by questions 2 and 3 above. The topics to which these 2 questions relate can be analogized to the perception of Cherenkov radiation by astronauts.232,233 When astronauts of both the Apollo and Soyuz missions were in darkness or with eyes closed, and especially when they were in a relaxed state, they experienced flashes of light, at the intra-ocular location where vision would be normally experienced. One explanation is that radiation (probably muons and pions) from outer space penetrated the space capsule, and traveled through the astronaut's eyeball and interacted with the retina, triggering an event registered in the brain as a flash, or streak, of light. Continuing from this example, so it may be with moments of protoconsciousness: perhaps neurons of the human brain are from time to time penetrated by precursors of protoconsciousness as a result of which sensitive sites, say among the arrays of MTs, are caused to activate a neural mechanism, the transduction of which is a ‘flash’ of consciousness. Furthermore, and in keeping with this speculative view of the induction of consciousness, the duration of the Cherenkov flash within the eye is short (≤ 10−10s) compared with the integration time of the eye (approx. 50 ms).232 Similarly with quantum protoconscious moments: an OR event occurs within a moment of time lasting about 13–20 ms; longer times (25 and 30 ms) are required to bring about, respectively, the recognition of the event and to set it in the correct place within the sequence of such events.
Concluding remarks
The evolution of animals and plants has allowed each to develop a Sensorium (state 1) and a Brain (state 2). An emergent state 3 (Mind), accompanied by consciousness, has appeared in some animals, including Man. The ordinary state of Man is characterized by an EEG record exhibiting a frequency of 4 Hz. In plants, spontaneous APs with frequencies one or 2 orders of magnitude less than this have been recorded. Nevertheless, both animals and plants exhibit a type of protoconsciousness in which, even if there is no reasoning or thought, there is sensitive awareness and cognizance of their respective internal, physiological environments and their external, physical environments. Man, however, has developed further capabilities and is able to experience a ‘higher’ state of consciousness. In this case, it may be that the bundling of MTs in neurons has provided the critical mass for a more intense vibration of 40 Hz (recorded from EEG), allowing the emergence of state 4, of self-reflective consciousness, and sometimes also a state 5 (Samadhi). Unfortunately, it is not known how this advancement of consciousness – the steps forward from states 2 and 3 to state 4, and thence to state 5 – have taken place, but it might, on the basis of Orch OR hypothesis, relate to the increasing mass and deployment of MTs in specific cells. The formula developed for the Orch OR hypothesis, τ ≈ Ћ /EG, predicts that, as the self-energy mass of MTs increases, τ, the frequency of quantum reduction events leading to protoconscious impulses, increases. Therefore, a larger EG would be associated with a larger complement of MTs and would lead to a higher rate of EEG-registered vibration (in Hz). But this raises certain questions: Is energy mass EG evaluated in terms of MT number per cell or per tissue? Is the value of EG associated with the spatial arrangement of MTs? Furthermore, is the number of cells bearing parallel and closely arranged MTs, a feature which is characteristic of growing plant cells, also a factor in the evaluation of EG? It is not known to what degree the numbers and arrangements of MTs in animal neurons and plant cells differ, but although individual plant cells mostly have relatively few MTs, bundled or otherwise, when compared to human neurons, certain specialized plant cells, such as the ray parenchyma cells of trees have considerably more. MT density might therefore influence the frequency of OR events but, in plants, MT density may limit the production of self-aware consciousness to trees where MTs are most abundant in cells involved with secondary growth. A Mind (state 3) seems to be a prerequisite for the recognition and acknowledgment of the conscious state in humans. But Mind appears to be an emergent property. Whether or not plants have a ‘mind’ is unanswerable: but plants, especially trees, may have a ‘phyto-mind’, sui generis, to which only they themselves can be present and can comprehend.
A state of conscious mindfulness, or awareness, emerges as an unfolding of the implicate quantum order into the explicated material aspect of existence through the presence of MTs. It is the ability to be consciously aware of the electrical impulses initiated by MTs, and whose passage the MTs direct toward the brain, that releases, or realizes, this potentiality for awareness. Thus, plants probably lag one step behind the experiential world of animals, for although both plants and animals may have attained state 2 (of Brain) and maybe state 3 (of Mind), animals (and humans in particular) have achieved the possibility of self-reflective consciousness (state 4). It is from this last-mentioned auto-generated desire for self-reflection, the property of state 4, that psychological, or psychokinetic, evolution in Man will continue, and will be accompanied by a parallel cultural evolution, as proposed by Gebser.234 On the other hand, Bergson116 was more comprehensive in considering not just human life, but life as a whole: “Now the more we fix our attention on the continuity of life, the more we see that organic evolution resembles the evolution of consciousness, in which the past pushing into the present causes a new form of awareness, incommensurable with what went before.” Acknowledgment of the displacement of the degree of conscious between plants and animals should enable humanity with its higher-grade consciousness and rapid mobility not only to nurture, comprehend and value the relative stillness of the lower-grade plant protoconsciousness, but also for both animals and plants to share mutual experiences through primary perception. This displacement of states of consciousness between plants and animals may be lawful in another way: to maintain the balance of Nature. To paraphrase Bergson (replacing in the following passage, the word ‘cell’ by ‘plant’, and ‘organism’ by ‘ecosystem’), “The component [plant] of an [ecosystem], on becoming momentarily conscious, would barely have outlived the wish to emancipate itself when it would be recaptured by necessity.”235
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
Gratitude is expressed to Dr John Gardiner for sharing many of his ideas on microtubules in relation to Orch OR hypothesis, and to Prof F. Baluška for useful information. Also thanked are Prof L.F. Agnati, Dr G. Dorda and Prof E. Zűrcher for their useful comments on early drafts of the typescript. Photomicrographs shown in Figure 4B and C were kindly provided by Prof Ryo Funada and Dr Shahanara Begum, and by Dr Nigel Chaffey, respectively. Mr Tim Colborne prepared all the material shown in Figure 1, 2, 3, and 5.
Figure 3.
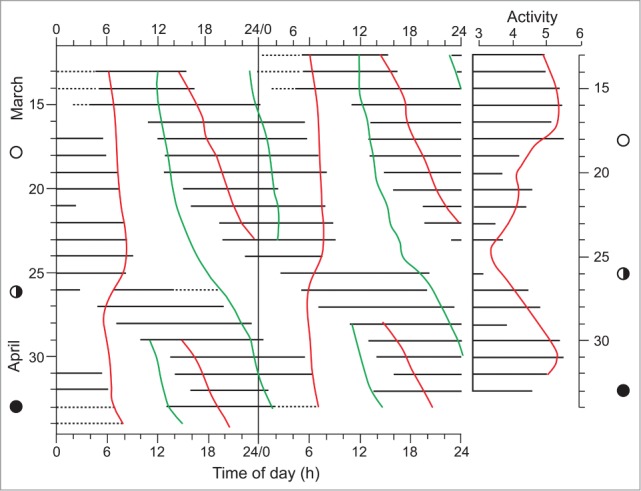
The perception of the passage of lunisolar time by a bean leaf. Diurnal movements of a bean leaf (Canavalia ensiformis) recorded at Delft, Netherlands, during March-April 1927. The leaf lamina, when raised to the ‘up’ position during the day, is potentially able to photosynthesise when in light. This is the position of ‘wakefulness’. The leaf descends to the ‘sleep’ position during the night (black bars). This oscillation continues even when the leaf is kept in constant light or darkness (as here). The changes in leaf position, from its lowest ‘down’ position to the ‘up’ position, occurs when there is a turning point in the lunisolar gravimetric tide (red and green lines, as in Fig. 1). The activity pattern - here the √amplitude of the leaf movement, using a 3-d moving average - is shown in the right-hand panel, together with the lunar phases (as in Fig. 1). During the first 3 d the leaf was in L:D 16 : 8 h (light commencing at 1600 h), for the next 12 d the leaf was in constant light (L:L). It was then moved to constant darkness (D:D) for the remaining 4 d (commencing at 0900 h on 30 April). (Original leaf rhythm data from Fig. 35 of Kleinhoonte153).
Funding
Financial support from School of Biological Sciences, Bristol University, is gratefully acknowledged.
References
- 1.Hameroff S, Penrose R. Consciousness in the universe. A review of the ‘Orch OR’ theory. Phys. Life Rev. 2014; 11:39-78; PMID:24070914; http://dx.doi.org/ 10.1016/j.plrev.2013.08.002 [DOI] [PubMed] [Google Scholar]
- 2.Dorda G. Quantisierte Zeit und die Vereinheitlichung von Gravitation und Elektromagnetismus. Göttingen, Cuvillier Verlag, 2010 [Google Scholar]
- 3.Seager W. Theories of Consciousness An Introduction and Assessment. London, Routledge, 1999 [Google Scholar]
- 4.Damasio AR. How the brain creates the mind. Sci. Am. 1999; 281(6):112-7; PMID:10614073; http://dx.doi.org/ 10.1038/scientificamerican1299-112 [DOI] [PubMed] [Google Scholar]
- 5.Damasio AR. The Feeling of What Happens: Body and Emotions in the Making of Consciousness New York, Harcourt Brace, New York, 1999 [Google Scholar]
- 6.Rosenberg G. A Place for Consciousness Probing the Deep Structure of the Natural World. Oxford, Oxford University Press, 2004 [Google Scholar]
- 7.Tart CT. The basic nature of altered states of consciousness: a systems approach. J. Transpersonal Psychol. 1976; 8:45-64 [Google Scholar]
- 8.Tart CT. (ed.). Altered States of Consciousness London, Harper Collins, 1990 [Google Scholar]
- 9.Vaitl D, Birbaumer N, Gruzweiler J, Jamieson GA, Kotchoubey B, Kűbler A, Lehmann D, Miltner WH, Ott U, Pütz P, et al.. Psychobiology of altered states of consciousness. Psychol. Bull. 2005; 131:98-127; PMID:15631555; http://dx.doi.org/ 10.1037/0033-2909.131.1.98 [DOI] [PubMed] [Google Scholar]
- 10.Nagel T. What is it like to be a bat? Phil. Rev. 1974; 73:435-50; http://dx.doi.org/ 10.2307/2183914 [DOI] [Google Scholar]
- 11.Seth AK, Baars BJ, Edelman DB. Criteria for consciousness in humans and other animals. Conscious. Cogn. 2005; 14:119-39; PMID:15766894; http://dx.doi.org/ 10.1016/j.concog.2004.08.006 [DOI] [PubMed] [Google Scholar]
- 12.Edelman DB, Baars BJ, Seth AK. Identifying hallmarks of consciousness in non-mammalian species. Conscious. Cogn. 2005; 14:169-87; PMID:15766896; http://dx.doi.org/ 10.1016/j.concog.2004.09.001 [DOI] [PubMed] [Google Scholar]
- 13.Edelman DB, Seth AK. Animal consciousness: a synthetic approach. Trends Neurosci. 2009; 32:476-84; PMID:19716185; http://dx.doi.org/ 10.1016/j.tins.2009.05.008 [DOI] [PubMed] [Google Scholar]
- 14.Feinberg TE, Mallatt J. The evolutionary and genetic origins of consciousness in the Cambrian period over 500 million years ago. Front. Psychol. 2013; 4(article 667):1-27; PMID:23382719; http://dx.doi.org/ 10.3399/fpsyg.2013.000667 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sheets-Johnstone M. Consciousness: a natural history. J. Consciousness Stud. 1998; 5:260-94; [Google Scholar]
- 16.Fischer-Friedrich E, Friedrich BM, Gov NS. FtsZ rings and helices: physical mechanisms for the dynamic aliginment of biopolymers in rod-shaped bacteria. Physical Biol. 2012; 9:016009; http://dx.doi.org/ 10.1088/1478-3975/9/1/016009 [DOI] [PubMed] [Google Scholar]
- 17.Vannini A. Quantum models of consciousness. Quantum Biosystems 2008; 2:165-84; [Google Scholar]
- 18.Walker E. 1970 The nature of consciousness. Math. BioSci. 1970; 7:131-78; http://dx.doi.org/ 10.1016/0025-5564(70)90046-5 [DOI] [Google Scholar]
- 19.Bernroider G, Summhammer J. Can quantum entanglement between ion transition state effect action potential initiation? Cognitive Comput. 2012; 4:29-37; http://dx.doi.org/ 10.1007/s12559-012-9126-7 [DOI] [Google Scholar]
- 20.Penrose R. Quantum computation, entanglement and state reduction. Phil. Trans. R. Soc. Lond. A, 1998; 356:1927-39; [Google Scholar]
- 21.Penrose R. On the gravitization of quantum mechanics 1: Quantum state reduction. Found. Phys. 2014; 44:557-75; http://dx.doi.org/ 10.1007/s10701-013-9770-0 [DOI] [Google Scholar]
- 22.Crick F, Koch C. Consciousness and neuroscience. Cereb. Cortex 1998; 8:97-107; http://dx.doi.org/ 10.1093/cercor/8.2.97 [DOI] [PubMed] [Google Scholar]
- 23.Sperry RW. Psychology's mentalist paradigm and the religion/science tension. Am. Psychol. 1988; 43:607-13; http://dx.doi.org/ 10.1037/0003-066X.43.8.607 [DOI] [Google Scholar]
- 24.Nagel AHM. Are plants conscious? J. Consciousness Stud. 1997; 4:215-30 [Google Scholar]
- 25.Trewavas A. Plant intelligence. Naturwissenschaften 2005; 92:401-13; PMID:16142448; http://dx.doi.org/ 10.1007/s00114-005-0014-9 [DOI] [PubMed] [Google Scholar]
- 26.Barlow PW. Plastic, inquisitive roots and intelligent plants in the light of some new vistas in plant biology. Plant Biosystems 2010; 144:396-407; http://dx.doi.org/ 10.1080/11263501003718570 [DOI] [Google Scholar]
- 27.Hodge A. Root decisions. Plant Cell Environ. 2008; 32:628-40; PMID:18811732; http://dx.doi.org/ 10.1111/j.1365-3040.2008.01891.x [DOI] [PubMed] [Google Scholar]
- 28.Calvo P, Keijser F. Cognition in plants In: Baluška F. (ed.), Plant Environment Interaction. Berlin, Springer-Verlag, 2009; 247-66 [Google Scholar]
- 29.Calvo P, Keijser F. Plants: Adaptive behaviour, root-brains, and minimal cognition. Adapt. Behav. 2011; 19:155-71; http://dx.doi.org/ 10.1177/1059712311409446 [DOI] [Google Scholar]
- 30.Chew MK, Laubichler MD. Natural enemies–Metaphor or misconception? Science 2003; 301:52-3; PMID:12846231; http://dx.doi.org/ 10.1126/science.1085274 [DOI] [PubMed] [Google Scholar]
- 31.Baars BJ. Metaphors of consciousness and attention in the brain. Trends Neurosci. 1998; 21:58-62; PMID:9498298; http://dx.doi.org/ 10.1016/S0166-2236(97)01171-5 [DOI] [PubMed] [Google Scholar]
- 32.Von Rospatt A. The Buddhist Doctrine of Momentariness. A Survey of the Origins and Early Phase of this Doctrine up to Vasubandhu. Stuttgart, Franz Steiner Verlag, 2005. Also available at: http://ahandfulofleaves.files.wordpress.com/2011/12/the-buddhist-doctrine-of-momentariness_vonrospatt.pdf [Google Scholar]
- 33.Gunter PAY, Papanicolaou AC. Bergson and Modern Thought: Towards a Unified Science. London, Routledge, 1987 [Google Scholar]
- 34.Mullarky J. (ed.). The New Bergson. Manchester, Manchester University Press, 1999 [Google Scholar]
- 35.Varela F, Lachaux J-P, Rodriguez E, Martinerie J. The brainweb: Phase synchronization and large-scale integration. Nature Rev. Neurosci. 2001; 2:229-39; http://dx.doi.org/ 10.1038/35067550 [DOI] [PubMed] [Google Scholar]
- 36.Baluška F, Mancuso S, Volkmann D, Barlow PW. Root apices as plant command centres: the unique “brain-like” status of the root apex transition zone. Biologia (Bratislava) 2004; 59:7-19 [Google Scholar]
- 37.Zeki S. The disunity of consciousness. Trends Cogn. Sci. 2003; 7:214-8; PMID:12757823; http://dx.doi.org/ 10.1016/S1364-6613(03)00081-0 [DOI] [PubMed] [Google Scholar]
- 38.Horio T, Murata T. The role of dynamic instability in microtubule organisation. Front. Plant Sci. 2014; 5(article 511):1-10; http://dx.doi.org/ 10.3389/fpls.2014.00511 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hameroff S, Penrose R. Conscious events as orchestrated space-time selections. NeuroQuantol. 2003; 1:10-35 [Google Scholar]
- 40.Sahu S, Ghosh S, Ghosh C, Aswani K, Hirata K, Fujita D, Bandyopadhyay A. Atomic water channel controlling remarkable properties of a single brain microtubule: correlating single protein to its supramolecular assembly. Biosens. Bioelectron. 2013; 47:141-6; PMID:23567633; http://dx.doi.org/ 10.1016/j.bios.2013.02.050 [DOI] [PubMed] [Google Scholar]
- 41.Priel A, Ramos AJ, Tuszynski JA, Cantiello HF. A biopolymer transistor: electrical amplification by microtubules. Biophys. J. 2006; 90:4639-43; PMID:16565058; http://dx.doi.org/; http://dx.doi.org/ 10.1529/biophysj.105.078915 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Cifra M, Pokorný J, Havelka D, Kučera O. Electric field generated by axial longitudinal vibration modes of microtubule. BioSystems 2010; 100:122-31; PMID:20178826; http://dx.doi.org/ 10.1016/j.biosystems.2010.02.007 [DOI] [PubMed] [Google Scholar]
- 43.Havelka D, Cifra M, Kučera O. Multi-mode electromechanical vibrations of a microtubule: In silico demonstration of electric pulse moving along a microtubule. Appl. Phys. Lett. 2014; 104:243702-1-4; http://dx.doi.org/ 10.1063/1.4884118 [DOI] [Google Scholar]
- 44.Gagliadi LJ, Shain DH. Polar electrostatic force drives poleward chromosome motions. Cell Division 2014; 9(5):1-14; PMID:25024738; http://dx.doi.org/ 10.1186/s13008-014-0005-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Penrose R, Hameroff S. Consciousness in the universe: Neuroscience, quantum space-time geometry and Orch OR theory. J. Cosmol. 2011; 14:1-17 [Google Scholar]
- 46.Hameroff S, Penrose R. Replies to seven commentaries on “Consciousness in the universe: Review of the ‘Orch OR’ theory”. Phys. Life Rev. 2014; 11:94-100; http://dx.doi.org/ 10.1016/j.plrev.2013.11.013 [DOI] [PubMed] [Google Scholar]
- 47.Baars BJ, Edelman DB. 2012. Consciousness, biology and quantum hypotheses. Phys. Life Rev. 2012; 9:285-94; PMID:22925839; http://dx.doi.org/ 10.1016/j.plrev.2012.07.001 [DOI] [PubMed] [Google Scholar]
- 48.Al-Khalili J, McFadden J. Life on the Edge. The Coming of Age of Quantum Biology. London, Bantam Press, 2014 [Google Scholar]
- 49.Panitchayangkoon G, Hayes D, Fransted KA, Caram JR, Harel E, Wen JZ, Blankenship RE, Engel GS. Long-lived quantum coherence in photosynthetic complexes at physiological temperatures. Proc. Natl. Acad. Sci. USA 2010; 107:12766-70; http://dx.doi.org/ 10.1073/pnas.1005484107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Turner DB, Dinshaw R, Lee K-K, Belsley MS, Wilk KE, Curmi PMG, Scholes GD. Quantitative investigations of quantum coherence for a light-harvesting protein at conditions simulating photosynthesis. Phys. Chem. Chem. Phys. 2012; 14:4857-74; PMID:22374579; http://dx.doi.org/ 10.1039/c2cp23670b [DOI] [PubMed] [Google Scholar]
- 51.Lambert N, Chen Y-H, Cheng Y-C, Li C-M, Chen G-Y, Nori F. Quantum biology. Nature Phys. 2012; 9:10-8; http://dx.doi.org/ 10.1038/nphys2474 [DOI] [Google Scholar]
- 52.Emerson DJ, Welser BP, Psonis J, Liao Z, Taratula O, Flamengo A, Wang X, Sugasawa K, Smith AB 3rd, Eckenhoff RG, et al.. Direct modulation of microtubule stability contributes to anthracene general anaesthesia. J. Am. Chem. Soc. 2013; 135:5389-98; PMID:23484901; http://dx.doi.org/ 10.1021/ja311171u [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Ledbetter MC, Porter KR. A ‘microtubule’ in plant fine structure. J. Cell Biol. 1963; 19:239-50; PMID:19866635; http://dx.doi.org/ 10.1083/jcb.19.1.239 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Baluška F, Volkmann D, Barlow PW. Nuclear components with microtubule-organizing properties in multicellular eukaryotes: Functional and evolutionary considerations. Int. Rev. Cytol. 1997; 175:91-135; http://dx.doi.org/ 10.1016/S0074-7696(08)62126-9 [DOI] [PubMed] [Google Scholar]
- 55.Shibaoka H. Microtubules and the regulation of cell morphogenesis by plant hormones In: Lloyd CW. (ed.), The Cytoskeletal Basis of Plant Growth and Form. London, Academic Press, 1991; 159-68 [Google Scholar]
- 56.Baluška F, Volkmann D, Barlow PW. Hormone-cytoskeleton interactions in plant cells In: Hooykaas PJJ, Hall MA, Libbenga KR (eds), Biochemistry and Molecular Biology of Plant Hormones. Amsterdam, Elsevier Science BV, 1999; 363-90 [Google Scholar]
- 57.Nick P. (ed.). Plant Microtubules. Berlin, Springer-Verlag, 2001 [Google Scholar]
- 58.Gardiner J. Insights into plant consciousness from neuroscience, physics and mathematics: a role for quasicrystals? Plant Signal. Behav. 2012; 7:1049-55; PMID:22899055; http://dx.doi.org/ 10.4161/psb.21325 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Gardiner J, Marc J. Arabidopsis thaliana, a plant model organism for the neuronal microtubule cytoskeleton? J. Exp. Bot. 2011; 62:89-97; PMID:20813785; http://dx.doi.org/ 10.1093/jxb/erq278 [DOI] [PubMed] [Google Scholar]
- 60.Bohm D, Peat FD. Science, Order, and Creativity. London, Routledge, 2011 [Google Scholar]
- 61.Bennett JG. The Dramatic Universe Volume 4 History. London, Hodder and Stoughton, 1966 [Google Scholar]
- 62.Vaneechoutte M. Experience, awareness and consciousness: suggestions for definitions as offered by an evolutionary approach. Found. Sci. 2000; 5:429-56; http://dx.doi.org/ 10.1023/A:1011371811027 [DOI] [Google Scholar]
- 63.Edelman GM. Naturalizing consciousness: A theoretical framework. Proc. Natl. Acad. Sci. USA 2003; 100:5520-4; http://dx.doi.org/ 10.1073/pnas.0931349100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Miller JG. Living Systems New York, McGraw-Hill, 1978 [Google Scholar]
- 65.Barlow PW. Living plant systems: How robust are they in the absence of gravity? Adv. Space Res. 1999; 23(12):1975-86; PMID:11710379; http://dx.doi.org/ 10.1016/S0273-1177(99)00340-3 [DOI] [PubMed] [Google Scholar]
- 66.Hayflick L. The illusion of cell immortality. Br. J. Cancer 2000; 83:841-6; PMID:10970682; http://dx.doi.org/ 10.1054/bjoc.2000.1296 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Tseng AS, Levin M. Cracking the bioelectric code. Probing endogenous ionic controls of pattern formation. Commun. Integr. Biol. 2013; 6:e22595. DOI: 10.4161/cib.22595; PMID:23802040; http://dx.doi.org/ 10.4161/cib.22595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Backster C. Primary Perception Biocommunication with Plants, Living Foods, and Human Cells. Anza CA, White Rose Millenium Press, 2003 [Google Scholar]
- 69.Ekman P. Emotions Revealed: Understanding Faces and Feelings London, Weidenfeld & Nicolson, 2003 [Google Scholar]
- 70.Ekman P, Levenson RW, Friesen WV. Autonomic nervous system activity distinguishes among emotions. Science 1983; 221:1208-10; PMID:6612338; http://dx.doi.org/ 10.1126/science.6612338 [DOI] [PubMed] [Google Scholar]
- 71.Beedie CJ, Terry PC, Lane AM. Distinctions between emotion and mood. Cognition Emotion 2005; 19:847-78; http://dx.doi.org/ 10.1080/02699930541000057 [DOI] [Google Scholar]
- 72.Barlow PW. Reflections on ‘plant neurobiology’. BioSystems 2008; 92:132-47; PMID:18336993; http://dx.doi.org/ 10.1016/j.biosystems.2008.01.004 [DOI] [PubMed] [Google Scholar]
- 73.Moroz LL. On the independent origins of complex brains and neurons. Brain Behav. Evol. 2009; 74:177-90; PMID:20029182; http://dx.doi.org/ 10.1159/000258665 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ryan TJ, Grant SGN. The origin and evolution of synapses. Nature Rev. Neurosci. 2009; 10:701-12; http://dx.doi.org/ 10.1038/nrn2717 [DOI] [PubMed] [Google Scholar]
- 75.Baluška F, Mancuso S, Volkmann D, Barlow PW. Root apex transition zone: a signalling-response nexus in the root. Trends Plant Sci. 2010; 15:402-8; http://dx.doi.org/ 10.1016/j.tplants.2010.04.007 [DOI] [PubMed] [Google Scholar]
- 76.Baluška F, Mancuso S. 2013 Root apex transition zone as oscillatory zone. Front. Plant Sci. 2013; 4(article 354):1-15; http://dx.doi.org/ 10.3389/fpls.2013.00354 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Feinberg TE. The nested neural hierarchy and the self. Conscious. Cogn. 2011; 20:4-15; PMID:20951608; http://dx.doi.org/ 10.1016/j.concog.2010.09.016 [DOI] [PubMed] [Google Scholar]
- 78.Agnati LF, Bjelke B, Fuxe K. Volume transmission in the brain. Am. Sci. 1992; 80:362-73 [Google Scholar]
- 79.Agnati LF, Guidolin D, Guescini M, Genedani S, Fuxe K. Understanding wiring transmission and volume transmission. Brain Res. Rev. 2010; 64:137-59; PMID:20347870; http://dx.doi.org/ 10.1016/j.brainresrev.2010.03.003 [DOI] [PubMed] [Google Scholar]
- 80.Agnati LF, Zunarelli E, Genedani S, Fuxe K. On the existence of a global molecular network enmeshing the whole central nervous system: Physiological and pathological implications. Curr. Prot. Peptide Sci. 2006; 7:3-15; http://dx.doi.org/ 10.2174/138920306775474086 [DOI] [PubMed] [Google Scholar]
- 81.Gunawardena AHLAN, Pearce DME, Jackson MB, Hawes CR, Evans DE. Rapid changes in cell wall pectic polysaccharides are closely associated with early stages of aerenchyma formation, a spatially localized form of programmed cell death in roots of maize (Zea mays L.) promoted by ethylene. Plant Cell Environ. 2001; 24:1369-75; http://dx.doi.org/ 10.1046/j.1365-3040.2001.00774.x [DOI] [Google Scholar]
- 82.Jackson MB, Hall KC. Early stomatal closure in waterlogged pea-plants is mediated by abscisic acid in the absence of foliar water deficits. Plant Cell Environ. 1987; 10:121-30 [Google Scholar]
- 83.Van Lommel P. Consciousness Beyond Life The Science of the Near-Death Experience. New York, HarperCollins, 2010 [Google Scholar]
- 84.Harner M. Cave and Cosmos Shamanic Encounters with Another Reality. Berkeley CA, North Atlantic Books, 2013 [Google Scholar]
- 85.Hameroff S, Chopra D. The “quantum soul”: A scientific hypothesis In: Moreira-Almeida A, Santos FS (eds.), Exploring Frontiers of the Mind-Brain Relationship. New York, Springer, 2012; 79-93; http://dx.doi.org/ 10.1007/978-4614-1_5 [DOI] [Google Scholar]
- 86.Bókkon I, Mallick BN, Tuszynski JA. Near death experiences: a multidisciplinary hypothesis. Front. Hum. Neurosci. 2013; 7(article 533):1-11; http://dx.doi.org/ 10.3399/fnhum.2013.00533 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Posner MI. Attention: the mechanisms of consciousness. Proc. Natl. Acad. Sci. USA 1994; 91:7398-403 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Bennett JG. The Dramatic Universe Volume 1 The Foundations of Natural Philosophy. London, Hodder and Stoughton, 1956 [Google Scholar]
- 89.Shotter J. Goethe and the refiguring of intellectual inquiry: From ‘aboutness’-thinking to ‘withness’-thinking in everyday life. Janus Head 2005; 8:132-58 [Google Scholar]
- 90.Wahl DC. “Zarte Empirie”: Goethean science as a way of knowing. Janus Head 2005: 8:58-76 [Google Scholar]
- 91.Dennis D. Humanistic neuroscience, mentality, and spirituality. J. Humanist. Psychol. 1995; 35:34-72; http://dx.doi.org/ 10.1177/00221678950352003 [DOI] [Google Scholar]
- 92.Sadhu M. Samhadi. London, Allen and Unwin, 1962 [Google Scholar]
- 93.Feuerstein G. The essence of yoga In: Feuerstein G, Miller J (eds.), A Reappraisal of Yoga. Essays in Indian Philosophy. London, Rider & Co, 1971; 1-47 [Google Scholar]
- 94.Joyce J. A Portrait of the Artist as a Young Man. London, Paladin, 1916/1988 [Google Scholar]
- 95.Dumit J. Neuroexistentialism In: Jones CA. (ed.), Sensorium. Embodied Experience, Technology, and Contemporary Art. Cambridge MA, MIT Press, 2006; 182-9 [Google Scholar]
- 96.Hoeller SA. Jung and the Lost Gospels. Wheaton IL, Theosophical Publishing House, 1989 [Google Scholar]
- 97.Hinterberger T, Schmidt S, Kamei T, Walach H. Decreased electrophysiological activity represents the conscious state of emptiness in meditation. Front. Psychol. 2014; 5(article 99):1-14; http://dx.doi.org/ 10.3389/fpsyg.2014.00099; PMID:24474945 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Schopenhauer A. The World as Will and Representation. Volume 1 New York, Dover Publications, New York, 1859/1969 [Google Scholar]
- 99.Nietzsche F. Birth of Tragedy. London, Penguin Books, 1872/1993 [Google Scholar]
- 100.Hartman AM, Hollister LE. Effect of mescaline, lysergic acid diethylamide and psilocybin on color perception. Psychopharmacologia 1963; 4:441-51; PMID:14048565; http://dx.doi.org/ 10.1007/BF00403349 [DOI] [PubMed] [Google Scholar]
- 101.Sinke C, Halpern JH, Zedler M, Neufeld J, Emrich HM, Passie T. Genuine and drug-induced synesthesia: a comparison. Conscious. Cogn. 2012; 21:1419-34; PMID:22521474; http://dx.doi.org/ 10.1016/j.concog.2012.03.009 [DOI] [PubMed] [Google Scholar]
- 102.Petri G, Expert P, Turkheimer F, Carhart-Harris R, Nutt D, Hellyer PJ, Vaccarino F. Homological scaffolds of brain functional networks. J. R. Soc. Interface 2014; 11:20140873; PMID:25401177; http://dx.doi.org/ 10.1098/rsif.2014.0873 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Theilhard de Chardin P The Future of Man. London, Collins, 1964 [Google Scholar]
- 104.Puthoff HE. Ground state of hydrogen as a zero-point-fluctuation-determined state. Phys Rev D 1987; 35:3266-9; http://dx.doi.org/; http://dx.doi.org/ 10.1103/PhysRevD.35.3266 [DOI] [PubMed] [Google Scholar]
- 105.Minot CS. The problem of consciousness in its biological aspects. Science 1902; 16:1-12; PMID:17801025; http://dx.doi.org/ 10.1126/science.16.392.1 [DOI] [PubMed] [Google Scholar]
- 106.Duckett JG. Pentagonal arrays of ribosomes in fertilized eggs of Pteridium aquilinum (L.) Kuhn. J. Ultrastruct. Res. 1972; 38:390-7; http://dx.doi.org/ 10.1016/S0022-5320(72)90013-5 [DOI] [PubMed] [Google Scholar]
- 107.Bouvier-Durand M, Dereuddre J, Côme D. Ultrastructural changes in the endoplasmic reticulum during dormancy release of apple embryos (Pyrus malus L.). Planta 1981; 151:6-14; PMID:24301664; http://dx.doi.org/ 10.1007/BF00384231 [DOI] [PubMed] [Google Scholar]
- 108.Bond DM, Finnegan EJ. Passing the message on: Inheritance of epigenetic traits. Trends Plant Sci. 2007; 12:211-6; PMID:17434332; http://dx.doi.org/ 10.1016/j.tplants.2007.03.010 [DOI] [PubMed] [Google Scholar]
- 109.Dengler NG. The influence of light on leaf development In: Iqbal M. (ed.), Growth Patterns in Vascular Plants. Portland OR, Dioscorides Press, 1994; 100-36 [Google Scholar]
- 110.Barlow PW, Powers SJ. Predicting the environmental thresholds for cambial and secondary vascular tissue development in stems of hybrid aspen. Ann. Forest Sci. 2005; 62:565-73; http://dx.doi.org/ 10.1051/forest:2005049 [DOI] [Google Scholar]
- 111.Takahashi H, Miyazawa Y, Fujii N. Hormonal interactions during root tropic growth; hydrotropism versus gravitropism. Plant Mol. Biol. 2009; 69:489-502; PMID:19083152; http://dx.doi.org/ 10.1007/s11103-008-9438-x [DOI] [PubMed] [Google Scholar]
- 112.Nakayama M, Kaneko Y, Miyazawa Y, Fujii N, Higashitani N, Wada S, Ishida H, Yoshimoto K, Shirasu K, Yamada K, et al.. A possible involvement of autophagy in amyloplast degradation in columella cells during hydrotropic response of Arabidopsis roots. Planta 2012; 236:999-1012; PMID:22532286; http://dx.doi.org/ 10.1007/s00425-012-1655-5 [DOI] [PubMed] [Google Scholar]
- 113.Bai H, Murali B, Barber K, Wolverton C. Low phosphate alters lateral root setpoint angle and gravitropism. Am. J. Bot. 2013; 100:175-82; PMID:23125433; http://dx.doi.org/ 10.3732/ajb.1200285 [DOI] [PubMed] [Google Scholar]
- 114.Spalding EP. Diverting the downhill flow of auxin to steer growth during tropisms. Am. J. Bot. 2013; 100:203-14; PMID:23284058; http://dx.doi.org/ 10.3732/ajb.1200420 [DOI] [PubMed] [Google Scholar]
- 115.Garrard B. The Ancient Problem with Men The Prehistoric Origins of Patriarchy and Social Oppression. Glastonbury, Unique Publications, 2011 [Google Scholar]
- 116.Bergson H. Creative Evolution. Minneola NY, Holt and Co, NY/Dover Publications, 1911/1998 [Google Scholar]
- 117.Gunter PAY. Bergson and the war against nature In: Mullarky J. (ed.), The New Bergson. Manchester, University of Manchester Press, 1999; 168-82 [Google Scholar]
- 118.Kitazawa MS, Fujimoto K. A developmental basis for stochasticity in floral organ numbers. Front. Plant Sci. 2014; 5(article 545):1-14; http://dx.doi.org/ 10.3389/fpls.2014.00545 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Besson S, Dumais J. Universal rule for symmetric division of plant cells. Proc Natl. Acad. Sci. USA 2011; 108:6294-9; http://dx.doi.org/ 10.1073/pnas.1011866108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Szyk A, Deaconescu AM, Spector J, Goodman B, Valenstein ML, Ziolkowska NE, Kormendi V, Grigorieff N, Roll-Mecak A. Molecular basis for age-dependent microtubule acetylation by tubulin acetyltransferase. Cell 2014; 157:1405-15; PMID:24906155; http://dx.doi.org/ 10.1016/j.cell.2014.03.061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Takemura R, Okabe S, Umeyama T, Kanai Y, Cowan NJ, Hirokawa N. Increased microtubular stability and alpha tubulin acetylation in cells transfected with microtubule-associated proteins MAP1B, MAP2 or tau. J. Cell Sci. 1992; 103:953-64; PMID:1487506 [DOI] [PubMed] [Google Scholar]
- 122.Sudo H, Baas PW. Acetylation of microtubules influences their sensitivity to severing by katanin in neurons and fibroblasts. J. Neurosci. 2010; 30:7215-26; PMID:20505088; http://dx.doi.org/ 10.1523/JNEUROSCI.0048-10.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Ballatore C, Lee V, M-Y, Trojanowski JQ. Tau-mediated neurodegeneration in Alzheimer's disease and related disorders. Nature Rev. Neurosci. 2007; 8:663-72; http://dx.doi.org/ 10.1038/nrn2194 [DOI] [PubMed] [Google Scholar]
- 124.Iqbal K, Alonso ADC, Chen S, Chohan MO, El-Akkad E, Gong CX, Khatoon S, Li B, Liu F, Rahman A, et al.. Tau pathology in Alzheimer disease and other tauopathies. Biochim. Biophys. Acta – Mol. Basis Dis. 2005; 1739:198-210; http://dx.doi.org/ 10.1016/j.bbadis.2004.09.008 [DOI] [PubMed] [Google Scholar]
- 125.Cantero JL, Moreno-Lopez B, Portillo F, Rubio A, Hita-Yañez E, Avila J. Role of tau protein on neocortical and hippocampal oscillatory patterns. Hippocampus 2011; 21:827-34; PMID:20607795 [DOI] [PubMed] [Google Scholar]
- 126.Lutz A, Greischar LL, Rawlings NB, Ricard M, Davidson RJ. Long-term meditators self-induce high-amplitude gamma synchrony during mental practice. Proc Natl. Acad. Sci. USA 2004; 101:16369-73; http://dx.doi.org/ 10.1073/pnas.0407401101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Cahn BR, Delorme A, Polich J. Occipital gamma activation during Vipassana meditation. Cogn. Process. 2010; 11:39-56; PMID:20013298; http://dx.doi.org/ 10.1007/s10339-009-0352-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Voss U, Holzmann R, Hobson A, Paulus W, Koppehele-Gossel J, Klimke A, Nitsche MA. 2014 Induction of self awareness in dreams through frontal low current stimulation of gamma activity. Nature Neurosci. 2014; 17:810-2; http://dx.doi.org/ 10.1038/nn.3719 [DOI] [PubMed] [Google Scholar]
- 129.Austin JH. Zen and the Brain Toward an Understanding of Meditation and Consciousness. Cambridge MA, MIT Press, 1998 [Google Scholar]
- 130.Bauer F, Cheadle SW, Parton A, Műller HJ, Usher M. Gamma flicker triggers attentional selection without awareness. Proc. Natl. Acad. Sci. USA 2009; 106:1666-71; http://dx.doi.org/ 10.1073/pnas.0810496106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.White JG, Southgate E, Thomson JN, Brenner S. 1986 The structure of the nervous system of the nematode Caenorhabditis elegans. Phil. Trans. R. Soc. Lond. B 1986; 314:1-340; http://dx.doi.org/ 10.1098/rstb.1986.0056 [DOI] [PubMed] [Google Scholar]
- 132.Hameroff S, Penrose R. Conscious events as orchestrated space-time selections In: Gordon BL, Dembski WA (eds.), The Nature of Nature: Examining the Role of Naturalism in Science. Wilmington DE, ISI Books, 2011 [Google Scholar]
- 133.Lee HJ, Ghetti A, Pinto-Duarte A, Wang X, Dziewczapolski G, Galimi F, Huitron-Resendiz S, Piña-Crespo JC, Roberts AJ, Verma IM, et al.. Astrocytes contribute to gamma oscillations and recognition memory. Proc. Natl. Acad. Sci. USA 2014; 111:E3343-52 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Hameroff S. How quantum brain biology can rescue conscious free will. Front. Integrative Neurosci. 2012; 6(article 93):1-17; http://dx.doi.org/ 10.3389/fnint.2012.00093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Govinda A. Foundations of Tibetan Mysticism. London, Rider, 1969 [Google Scholar]
- 136.Feuerstein GA. The Essence of Yoga A Contribution to the Psychohistory of Indian Civilization. London, Rider, 1974 [Google Scholar]
- 137.Banquet J. Spectral analysis of the EEG of meditation. Electroen. Clin. Neurophysiol. 1973; 35:143-51; http://dx.doi.org/ 10.1016/0013-4694(73)90170-3 [DOI] [PubMed] [Google Scholar]
- 138.Kasamatsu A, Hirai T. An electrocephalographic study on the Zen meditation (Zazen). Folio Psychiat. Neurolog. Jap. 1996; 20:315-36 [DOI] [PubMed] [Google Scholar]
- 139.Murata T, Koshino Y, Omori M, Murata I, Nishio M, Sakamoto K, et al.. 1994 Quantitative study on Zen meditation (Zazen). Jap. J. Psychiat. Neurol. 1994; 48:881-90 [Google Scholar]
- 140.Cahn BR, Polich J. Meditation states and traits: EEG, ERP, and neuroimaging studies. Psychol. Bull. 2006; 132:180-211; PMID:16536641; http://dx.doi.org/ 10.1037/0033-2909.132.2.180 [DOI] [PubMed] [Google Scholar]
- 141.Berman AE, Stevens L. EEG manifestations of nondual experiences in meditators. Conscious. Cogn. 2015; 31:1-11; PMID:25460236; http://dx.doi.org/ 10.1016/j.concog.2014.10.002 [DOI] [PubMed] [Google Scholar]
- 142.Persinger MA. Increased geomagnetic activity and the occurrence of bereavement hallucinations: Evidence for melatonin-mediated microseizuring in the temporal lobe? Neurosci. Lett. 1988; 88:271-4; PMID:3386872; http://dx.doi.org/ 10.1016/0304-3940(88)90222-4 [DOI] [PubMed] [Google Scholar]
- 143.Krippner S, Persinger M. Evidence for enhanced congruence between dreams and distant target material during periods of decreased geomagnetic activity. J. Scient. Explor. 1996; 10:487-93 [Google Scholar]
- 144.Palmer SJ, Rycroft MJ, Cermak M. Solar and geomagnetic activity, extremely low frequency magnetic and electric fields and human health at the Earth's surface. Surv. Geophys. 2006; 27:557-95; http://dx.doi.org/ 10.1007/s10712-006-9010-7 [DOI] [Google Scholar]
- 145.Sátori G, Zieger B. Spectral characteristics of Schumann resonances observed in Central Europe. J. Geophys. Res. 1996; 101(D23):29663-9; http://dx.doi.org/ 10.1029/96JD00549 [DOI] [Google Scholar]
- 146.Foley M. Life Lessons from Bergson. London, Macmillan, 2013 [Google Scholar]
- 147.Barlow PW, Fisahn J. Lunisolar tidal force and the growth of plant roots, and some other effects on plant movements. Ann. Bot. 2012; 110:301-18; PMID:22437666; http://dx.doi.org/ 10.1093/aob/mcs038 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Barlow PW. Leaf movements and their relationship with the lunisolar gravitational force. Ann. Bot. 2015; 116:149-187; http://dx.doi.org/ 10.1093/aob/mcv096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Nicholson C, Syková E. Extracellular space structure revealed by diffusion analysis. Trends Neurosci. 1988; 21:207-15; http://dx.doi.org/ 10.1016/S0166-2236(98)01261-2 [DOI] [PubMed] [Google Scholar]
- 150.Agnati LF, Fuxe K. Extracellular-vesicle type of volume transmission and tunnelling-nanotube type of wiring transmission add a new dimension to brain neuro-glial networks. Phil. Trans. R. Soc. Lond. B 2014; 369(1652): 20130505; http://dx.doi.org/ 10.1098/rstb.2013.0505 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Barnwell FW. Daily and tidal patterns of activity in individual Fiddler Crab (Genus Uca) from the Woods Hole region. Biol. Bull. 1966; 130:1-17; PMID:5907300; http://dx.doi.org/ 10.2307/1539948 [DOI] [PubMed] [Google Scholar]
- 152.Mills JN, Minors DS, Waterhouse JM. The circadian rhythms of human subjects without timepieces or indication of the alternation of day and night. J. Physiol. 1974; 240:567-94; PMID:4416124; http://dx.doi.org/ 10.1113/jphysiol.1974.sp010623 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Kleinhoonte A. 1929 Űber die durch das Licht regulierten autonomen Bewegungen der Canavalia-Blätter. Arch. Néerland. Sci. Exactes Naturelles Ser. B 1929; 5:1-110 [Google Scholar]
- 154.Harris WS, Gowda M, Kolb JW, Strychacz CP, Vacek JL, Jones PG, Forker A, O'Keefe JH, McCallister BD. A randomized, controlled trial of the effects of remote, intercessory prayer on outcomes in patients admitted to the coronary care unit. Arch. Intern. Med. 1999; 155:2273-8 [DOI] [PubMed] [Google Scholar]
- 155.Hodge DR. A systematic review of the empirical literature on intercessionary prayer. Res. Social Work Pract. 2007; 17:174-87; http://dx.doi.org/ 10.1177/1049731506296170 [DOI] [Google Scholar]
- 156.Lesniak KT. The effect of intercessionary prayer on wound healing in nonhuman primates. Altern. Therap. Health Med. 2006; 12:42-8 [PubMed] [Google Scholar]
- 157.Vogel M. Man–plant communication In: White J. (ed.), Psychic Exploration. A Challenge for Science. New York, GP Putnam's Sons, 1974; 289-312 [Google Scholar]
- 158.Lawlis F. Shamanic approaches in a hospital pain clinic In: Doore G. (ed.), Shamanic Path Healing, Personal Growth and Empowerment. Boston, Shambala, 1988; 139-49 [Google Scholar]
- 159.Vasiliauskas SL, McMinn MR. The effects of a prayer intervention on the process of forgiveness. Psychol. Religion Spirituality 2012; 5:23-32; http://dx.doi.org/ 10.1037/a0029324 [DOI] [Google Scholar]
- 160.Lindahl BIB, Århem P. Mind as a force field: Comments on a new interactionist hypothesis. J. Theor. Biol. 1994; 171:111-22; PMID:7844991; http://dx.doi.org/ 10.1006/jtbi.1994.1217 [DOI] [PubMed] [Google Scholar]
- 161.Letchoumanane S, Gupta VK. Effect of thought transaction on ‘okra’ yield. Ancient Sci. 2014; 1:49-56; http://dx.doi.org/ 10.14259/as.v1i2.163 [DOI] [Google Scholar]
- 162.Bose JC. The response of plants to wireless stimulation. Nature 1919; 104:172-4; http://dx.doi.org/ 10.1038/104172c0 [DOI] [Google Scholar]
- 163.Murphy GP, Dudley SA. Kin recognition: Competition and cooperation in Impatiens (Balsaminaceae). Am. J. Bot. 2009; 96:1990-6; PMID:21622319; http://dx.doi.org/ 10.3732/ajb.0900006 [DOI] [PubMed] [Google Scholar]
- 164.Gagliano M, Renton M. Love thy neighbor: facilitation through an alternative signalling modality in plants. BMCEcol. 2013; 13:19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Depuydt S. Arguments for and against self and non-self root recognition in plants. Front. Plant Sci 2014; 5(article 614):1-7; http://dx.doi.org/ 10.3389/fpls.2014.00614 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Mercer CA, Eppley SM. Kin and sex recognition in a dioecious grass. Plant Ecol. 2014; 214:845-52; http://dx.doi.org/ 10.1007/s11258-014-0336-9 [DOI] [Google Scholar]
- 167.Karban R, Wetzel WC, Shiojiri K, Ishixaki S, Ramirez SR, Blande JD. Deciphering the language of plant communication: volatile chemotypes of sagebush. New Phytol. 2014; 204:380-5; PMID:24920243; http://dx.doi.org/ 10.1111/nph.12887 [DOI] [PubMed] [Google Scholar]
- 168.Barlow PW, Fisahn J. 2013 Swarms, swarming and entanglements of fungal hyphae and of plant roots. Commun. Integr. Biol. 2013; 6, e252991.; http://dx.doi.org/ 10.4161/cib.25299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Champagnat P. Les corrélations sur le rameau d'un an des végétaux ligneux. ΦYTON 1954; 4:1-102 [Google Scholar]
- 170.Champagnat P. Les correlations entre feuilles et bourgeons de la pousse herbacée du lilas. Rev. Gén. Bot 1955; 62:325-71 [Google Scholar]
- 171.Nozeran R. Integration of organismal development In: Barlow PW, Carr DJ (eds.), Positional Controls in Plant Development. Cambridge, Cambridge University Press, 1984; 375-401 [Google Scholar]
- 172.Barlow PW. From cell to system: Repetitive units of growth in the development of roots and shoots In: Iqbal M. (ed.), Growth Patterns in Vascular Plants. Portland OR, Dioscorides Press, 1994; 19-58 [Google Scholar]
- 173.Blomme J, Inzé D, Gonzalez N. The cell-cycle interactome: a source of growth regulators? J. Exp. Bot. 2014; 65:2715-30; http://dx.doi.org/ 10.1093/jxb/ert388 [DOI] [PubMed] [Google Scholar]
- 174.Gamalei Yu V. Supercellular plant organization. Russ. J. Plant Physiol. 1997; 44:819-46 [Google Scholar]
- 175.Baluška F, Volkmann D, Barlow PW. Cell-cell channels and their implications for cell theory In: Baluška F, Volkmann D, Barlow PW (eds), Cell-Cell Channels. Georgetown TX, Landes Bioscience, and New York, Springer Science, 2006; 1-18 [Google Scholar]
- 176.Spicer R. 2014. Symplasmic networks in secondary vascular tissues: parenchyma distribution and actively supporting long-distance transport. J. Exp. Bot. 2014; 65:1829-48.; PMID:24453225; http://dx.doi.org/ 10.1093/jxb/ert459 [DOI] [PubMed] [Google Scholar]
- 177.Gagliano M, Renton M, Duvdevani N, Timmins M, Mancuso S. Acoustic and magnetic communication in plants. Is it possible? Plant Signal. Behav. 2012; 7:1346-8; PMID:22902698; http://dx.doi.org/ 10.4161/psb.21517 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Gagliano M, Renton M, Duvdevani N, Timmins M, Mancuso S. Out of sight but not out of mind: alternative means of communication in plants. PLoS ONE 2012; 7(5):e37382; PMID:22629387; http://dx.doi.org/ 10.1371/journal.pone.0037382 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Bose JC. The Nervous Mechanisms of Plants London, Longmans, Green and Co, 1926. (http://www.drmichaellevin.org/Nervous%20mechanism%20of%20plants.pdf) [Google Scholar]
- 180.Umrath K. Untersuchungen űber Plasma und Plasmastrőmungen an Characeen IV. Potentialmessungen an Nitella mucronata mit besonderer Berűcksichtigung der Erregungserscheinungen. Protoplasma 1930; 9:576-97; http://dx.doi.org/ 10.1007/BF01943373 [DOI] [Google Scholar]
- 181.Baluška F, Volkmann D, Menzel D. Plant synapses: actin-based domains for cell-cell communication. Trends Plant Sci. 2005; 10:106-11; http://dx.doi.org/ 10.1016/j.tplants.2005.01.002 [DOI] [PubMed] [Google Scholar]
- 182.Baluška F, Volkmann D, Hlavacka A, Mancuso S, Barlow PW. Neurobiological view of plants and their body plan In: Baluška F, Mancuso S, Volkmann D. (eds), Communication in Plants. Neuronal Aspects of Plant Life. Berlin, Springer-Verlag, 2006; 19-35 [Google Scholar]
- 183.Fromm J, Lautner S. Electrical signals and their physiological significance in plants. Plant Cell Environ. 2007; 30:249-57; PMID:17263772; http://dx.doi.org/ 10.1111/j.1365-3040.2006.01614.x [DOI] [PubMed] [Google Scholar]
- 184.Murch SJ. Neurotransmitters, neuroregulators and neurotoxins in plants In: Baluška F, Mancuso S, Volkmann D (eds.), Communication in Plants. Neuronal Aspects of Plant Life. Berlin, Springer, 2006; 137-51 [Google Scholar]
- 185.Gardiner J, Overall R, Marc J. The microtubule cytoskeleton acts as a key downstream effector of neurotransmitter signalling. Synapse 2011; 65: 249-56; PMID:20687109; http://dx.doi.org/ 10.1002/syn.20841 [DOI] [PubMed] [Google Scholar]
- 186.Kiewert C, Bucholzer M-L, Hartmann J, Chatterjee SS, Klein J. Stimulation of hippocampal acetylcholine release by hyperforin, a constituent of St. John's Wort. Neurosci, Lett. 2004; 364:195-8; http://dx.doi.org/ 10.1016/j.neulet.2004.04.046 [DOI] [PubMed] [Google Scholar]
- 187.Daly JW. Nicotinic agonists, antagonists, and modulators from natural sources. Cell. Mol. Neurobiol. 2005; 25:513-52; PMID:16075378; http://dx.doi.org/ 10.1007/s10571-005-3968-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Neher E, Steinbach JH. 1978 Local anaesthetics transiently block currents through single acetylcholine-receptor channels. J. Physiol. 1978; 77:153-76; http://dx.doi.org/ 10.1113/jphysiol.1978.sp012267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Weber M, Motin L, Gaul S, Beker F, Fink RHA, Adams DJ. 2005. Intravenous anaesthetics inhibit nicotinic acetylcholine receptor-mediated currents and Ca2+ transients in rat intracardiac ganglion neurons. Br. J. Pharmacol. 2005; 144:98-107; PMID:15644873; http://dx.doi.org/ 10.1038/sj.bjp.0705942 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Leys SP, Mackie GO, Meech R. Impulse conduction in a sponge. J. Exp. Biol. 1999; 202:1139-50; PMID:10101111 [DOI] [PubMed] [Google Scholar]
- 191.Elliott GRD, Leys SP. Evidence for glutamate, GABA, and NO in coordinating behaviour in the sponge, Ephydatia muelleri (Desmospongiae, Spongillidae). J. Exp. Biol. 2010; 213:2310-21; PMID:20543130; http://dx.doi.org/ 10.1242/jeb.039859 [DOI] [PubMed] [Google Scholar]
- 192.Levin M. Morphogenetic fields in embryogenesis, regeneration, and cancer: Non-local control of complex patterning. BioSystems 2012; 109:243-61; PMID:22542702; http://dx.doi.org/ 10.1016/j.biosystems.2012.04.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Gunning BES, Overall RL. Plasmodesmata and cell-to-cell transport in plants. BioSci. 1983; 33:260-5; http://dx.doi.org/ 10.2307/1309039 [DOI] [Google Scholar]
- 194.Sager R, Lee JY. Plasmodesmata in integrated cell signalling: Insights from development and environmental signals and stresses. J. Exp. Bot. 2014; 65:6337-58; PMID:25262225; http://dx.doi.org/ 10.1093/jxb/eru365 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Lee JY. New and old roles of plasmodesmata in immunity and parallels to tunnelling nanotubes. Plant Sci. 2014; 221–222:13-20; PMID:24656331 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Overall RL, Gunning BES. Intercellular communication in Azolla roots: II. Electrical coupling. Protoplasma 1982; 111:151-60; http://dx.doi.org/ 10.1007/BF01282072 [DOI] [Google Scholar]
- 197.Holdaway-Clarke TL, Walker NA, Hepler PK, Overall RL. Physiological elevations in cytoplasmic free calcium or ion injection results in transient closure of higher plant plasmodesmata. Planta 2000; 210:329-35; PMID:10664140; http://dx.doi.org/; http://dx.doi.org/ 10.1007/PL00008141 [DOI] [PubMed] [Google Scholar]
- 198.Traub RD, Kopell N, Bibbig A, Buhl EH, LeBeau FEN, Whittington MA. Gap junctions between interneuron dendrites can enhance synchrony of gamma oscillations in distributed networks. J. Neurosci. 2001; 21:9478-86; PMID:11717382 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Nunn JF, Lovis JD, Kimball KL. Arrest of mitosis by halothane. Br. J. Anaesth. 1971; 43:524-30; PMID:5089930; http://dx.doi.org/ 10.1093/bja/43.6.524 [DOI] [PubMed] [Google Scholar]
- 200.Saltveit ME., Jr Effect of high-pressure gas atmospheres and anaesthetics on chilling injury of plants. J. Exp. Bot. 1993; 44: 1361-8; http://dx.doi.org/ 10.1093/jxb/44.8.1361 [DOI] [Google Scholar]
- 201.Roberts IN, Lloyd CW, Roberts K. Ethylene-induced microtubule reorientations: mediation by helical arrays. Planta 1985; 164:439-47; PMID:24248216; http://dx.doi.org/ 10.1007/BF00395959 [DOI] [PubMed] [Google Scholar]
- 202.Baluška F, Brailsford RW, Hauskrecht M, Jackson MB, Barlow PW. Cellular morphogenesis in the maize root cortex: Involvement of microtubules, ethylene and gibberellins in the differentiation of cellular behaviour in postmitotic growth zones. Bot. Acta 1993; 106:394-403 [Google Scholar]
- 203.Asavasanti S, Stroeve P, Barrett DM, Jernstedt JA, Ristenport WD. Enhanced electroporation in plant tissues via low frequency pulsed electric fields: Influence of cytoplasmic streaming. Biotechnol. Progr. 2012; 28:445-53; http://dx.doi.org/ 10.1002/btpr.1507 [DOI] [PubMed] [Google Scholar]
- 204.Funada R. Control of wood structure In: Nick P. (ed.), Plant Microtubules. Potential for Biotechnology. Berlin, Springer, 2001; 51-81 [Google Scholar]
- 205.Begum A, Shibagaki M, Furusawa O, Nakaba S, Yamagishi Y, Yoshimoto J, Jin HO, Sano Y, Funada R. Cold stability of microtubules in wood-forming tissues of conifers during seasons of active and dormant cambium. Planta 2012; 235:165-79; PMID:21861112; http://dx.doi.org/ 10.1007/s00425-011-1500-2 [DOI] [PubMed] [Google Scholar]
- 206.Chaffey N, Barlow P, Sundberg B. Understanding the role of the cytoskeleton in wood formation in angiosperm trees: Hybrid aspen (Populus tremula x P. tremuloides) as the model species. Tree Physiol. 2002; 22:239-49; PMID:11874720; http://dx.doi.org/ 10.1093/treephys/22.4.239 [DOI] [PubMed] [Google Scholar]
- 207.Chaffey NJ, Barnett JR, Barlow PW. Visualization of the cytoskeleton within the secondary vascular system of hardwood species. J. Microscop. 1997: 187:77-84; http://dx.doi.org/ 10.1046/j.1365-2818.1997.2270786.x [DOI] [PubMed] [Google Scholar]
- 208.Barlow PW, Brain P, Powers SJ. Estimation of directional division frequencies in vascular cambium and in marginal meristematic cells of plants. Cell Prolif. 2002; 35:49-68; PMID:11856178; http://dx.doi.org/ 10.1046/j.1365-2184.2002.00225.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Nakaba S, Begum S, Yamagishi Y, Jin H, Kubo T, Funada R. Differences in the timing of cell death, differentiation and function among three different types of ray parenchyma cells in the hardwood Populus sieboldii × P. grandidentata. Trees 2012; 26:743-50; http://dx.doi.org/ 10.1007/s00468-011-0640-0 [DOI] [Google Scholar]
- 210.Max-Planck-Institute, Mitchison Lab protocol at http://hymanlab.mpi-cbg.de/hyman_lab/wp-content/uploads/2012/08/Tubulin-Protocols-Mitchison.pdf. [Google Scholar]
- 211.Dziubińska H. Ways of signal transmission and physiological role of electrical potentials in plants. Acta Soc. Bot. Polon. 2003; 72:309-18; http://dx.doi.org/ 10.5586/asbp.2003.040 [DOI] [Google Scholar]
- 212.McFadden J. Conscious electromagnetic theory. NeuroQuantol. 2007; 5:262-70; http://dx.doi.org/ 10.14704/nq.2007.5.3.135 [DOI] [Google Scholar]
- 213.Gibert D, Le Mouël J-L, Lambs L, Nicollin F, Perrier F. Sap flow and daily electric potential variations in a tree trunk. Plant Sci. 2006; 171:572-84; http://dx.doi.org/ 10.1016/j.plantsci.2006.06.012 [DOI] [Google Scholar]
- 214.Stahlberg R, Cleland RE, Van Volkenburgh E. Slow wave potentials – a propagating electrical signal unique to plants In: Baluška F, Mancuso S, Volkmann D (eds.), Communication in Plants. Neuronal Aspects of Plant Life. Berlin, Springer, 2006; 291-308 [Google Scholar]
- 215.Davies E, Stankovic B. Electrical signals, the cytoskeleton, and gene expression: a hypothesis on the coherence of the cellular responses to environmental insult In: Baluška F, Mancuso S, Volkmann D (eds.), Communication in Plants. Neuronal Aspects of Plant Life. Berlin, Springer, 2006; 309-20 [Google Scholar]
- 216.Stanković B, Davies E. Intercellular communication in plants: electrical stimulation of proteinase inhibitor gene expression in tomato. Planta 1997; 202:402-6; http://dx.doi.org/ 10.1007/s004250050143 [DOI] [Google Scholar]
- 217.Häder D-P. Evidence of electrical potential changes in photophobically reacting blue-green algae. Arch. Microbiol. 1978; 118:115-9; http://dx.doi.org/ 10.1007/BF00406083 [DOI] [Google Scholar]
- 218.Thavarungkul P, Lertsithichai S, Sherlock RA. Spontaneous action potential initiation and propagation in regenerating cell segments of Acetabularia mediterranea. J. Exp. Bot. 1987; 38:1541-56; http://dx.doi.org/ 10.1093/jxb/38.9.1541 [DOI] [Google Scholar]
- 219.Saddler HD. Spontaneous and induced changes in the membrane potential and resistance of Acetabularia mediterranea. J. Membrane Biol. 1971; 5:250-60; http://dx.doi.org/ 10.1007/BF01870553 [DOI] [PubMed] [Google Scholar]
- 220.Liddle LB, Gleissberg S, Giese G, Berger S. Microtubule formation on the nuclear surface during meiosis in Acetabularia acetabulum. Protoplasma 1997; 200:10-16; http://dx.doi.org/ 10.1007/BF01280730 [DOI] [Google Scholar]
- 221.Paszewski A, Dzubińska H, Trębacz K, Zawadski T. Electrical activity of the liver-wort Conocephalum conicum: Method of investigation and general characteristics of excitation. Physiol. Plant. 1982; 54:83-7; http://dx.doi.org/ 10.1111/j.1399-3054.1982.tb00582.x [DOI] [Google Scholar]
- 222.Zawadski T, Dziubińska H, Davies E. Characteristics of action potential generated spontaneously in Helianthus annuus. Physiol. Plant. 1995; 93:291-7; http://dx.doi.org/ 10.1034/j.1399-3054.1995.930213.x [DOI] [Google Scholar]
- 223.Glębicki K, Hejnowicz Z, Pijanowski A. Localized spontaneous fluctuations of electrical potential in shoots of different plants. Acta Soc. Bot. Polon. 1986; 55:67-76; http://dx.doi.org/ 10.5586/asbp.1986.008 [DOI] [Google Scholar]
- 224.Karlsson L. Nonrandom bioelectrical signals in plant tissue. Plant Physiol. 1972; 49:982-6; PMID:16658096; http://dx.doi.org/ 10.1104/pp.49.6.982 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Pickard BG. Spontaneous electrical activity in shoots of Ipomoea, Pisum, and Xanthium. Planta 1972; 102:91-114; http://dx.doi.org/ 10.1007/BF00384863 [DOI] [PubMed] [Google Scholar]
- 226.Williams SE, Pickard BG. Receptor potentials and action potentials in Drosera tentacles. Planta 1972; 103:193-221; PMID:24481555; http://dx.doi.org/ 10.1007/BF00386844 [DOI] [PubMed] [Google Scholar]
- 227.Hejnowicz Z, Krause E, Glebicki K, Sievers A. Propagated fluctuations of the electric potential in the apoplasm of Lepidium sativum L. roots. Planta 1991; 186:127-34; PMID:24186585; http://dx.doi.org/ 10.1007/BF00201508 [DOI] [PubMed] [Google Scholar]
- 228.Allison AC, Hulands GH, Nunn JF, Kitching JA, Macdonald AC. The effect of inhalational anaesthetics on the microtubular system in Actinosphaerium nucleofilum. J. Cell Sci. 1970; 7:483-99; PMID:5530568 [DOI] [PubMed] [Google Scholar]
- 229.Chu CR, Hsieh C-I, Wu S-Y, Phillips NG. Transient response of sap flow to wind speed. J. Exp. Bot. 2009; 60:249-55; PMID:19022910; http://dx.doi.org/ 10.1093/jxb/ern282 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Burr HS, Moon madness. Yale J. Biol. Med. 1944; 16:249-56; PMID:21434145 [PMC free article] [PubMed] [Google Scholar]
- 231.Barlow PW. Moon and cosmos: Plant growth and plant bioelectricity In: Volkov AG. (ed.), Plant Electrophysiology. Signaling and Responses. Berlin, Springer-Verlag, 2012; 249-280 [Google Scholar]
- 232.Fazio GG, Jelley JV, Charman WN. Generation of Cherenkov light flashes by cosmic radiation within the eyes of the Apollo astronauts. Nature 1979; 228:260-4; http://dx.doi.org/ 10.1038/228260a0 [DOI] [PubMed] [Google Scholar]
- 233.Sannita WG, Narici L, Picozza P. Positive visual phenomena in space: A scientific case and a safety issue in space travel. Vision Res. 2006; 46:2159-65; PMID:16510166; http://dx.doi.org/ 10.1016/j.visres.2005.12.002 [DOI] [PubMed] [Google Scholar]
- 234.Gebser J. The Ever-Present Origin: The Foundations and Manifestation of the Aperspectival World. Athens OH, Ohio University Press, 1986 [Google Scholar]
- 235.Bergson H. The Two Sources of Morality and Religion. Notre Dame IN, University of Notre Dame Press, 1935/1977 [Google Scholar]