

Abstract
Liver fatty acid-binding protein (L-FABP), also known as fatty acid-binding protein 1 (FABP1), is a key regulator of hepatic lipid metabolism. Elevated FABP1 levels are associated with an increased risk of cardiovascular disease (CVD) and metabolic syndromes. In this study, we examine the association of FABP1 gene promoter variants with serum FABP1 and lipid levels in a Chinese population. Four promoter single-nucleotide polymorphisms (SNPs) of FABP1 gene were genotyped in a cross-sectional survey of healthy volunteers (n = 1,182) from Fuzhou city of China. Results showed that only the rs2919872 G>A variant was significantly associated with serum TG concentration(P = 0.032).Compared with the rs2919872 G allele, rs2919872 A allele contributed significantly to reduced serum TG concentration, and this allele dramatically decreased the FABP1 promoter activity(P < 0.05). The rs2919872 A allele carriers had considerably lower serum FABP1 levels than G allele carriers (P < 0.01). In the multivariable linear regression analysis, the rs2919872 A allele was negatively associated with serum FABP1 levels (β = —0.320, P = 0.003), while serum TG levels were positively associated with serum FABP1 levels (β = 0.487, P = 0.014). Our data suggest that compared with the rs2919872 G allele, the rs2919872 A allele reduces the transcriptional activity of FABP1 promoter, and thereby may link FABP1 gene variation to TG level in humans.
Introduction
Clinical lipid disorders are associated with enormous public health significance and increasing societal burden in many countries [1]. Epidemiological evidence supporting raised plasma triglycerides (TG) is emerging as an independent risk factor for Type 2 diabetes, fatty liver, metabolic syndrome and atherosclerotic cardiovascular disease (CVD) [2,3,4]. The concentration of TG in an individual depends on the interplay between genetic and environmental factors. Although environmental factors such as smoking and alcohol consumption are triggers, hypertriglyceridemia has a tendency to cluster within families, suggesting that genetic factors also contribute to the risk of developing this disorder. Although many genetic candidates have been discovered to date, these only explain a small fraction of the total inter-individual variation in plasma TG levels [5,6,7]. Therefore, the search for the genetic factors that explain the increased susceptibility to hypertriglyceridemia is a current focus of research.
The Liver fatty acid-binding protein (L-FABP), also known as fatty acid-binding protein 1 (FABP1), is a member of the FABP family that is found in abundance in the cytosol of liver parenchymal cells [8]. It serves as an intracellular acceptor of long-chain fatty acids (LCFA), following their cellular uptake, trafficking, and mitochondrial oxidation [9,10]. Studies performed in vitro and in vivo indicate that FABP1 plays a role in the incorporation of fatty acids into TGs [11]. For example, murine FABP1 overexpression increased LCFA uptake and increased hepatic TG, while decreased LCFA uptake and reduced hepatic TG levels have been found in FABP1 knockout mice [12,13,14]. Given the key role of FABP1 in lipid metabolism, it is conceivable that variation in the FABP1 gene, either in the coding region or regions that regulate FABP1 expression, could directly influence plasma TG levels or others lipid-related phenotypes. In fact, a highly conserved c.340A>G missense mutation in exon 3 of the human FABP1 gene alters a threonine (T) to alanine (A) at position 94 (T94A) and is thought to contribute negatively to FA binding. This variant is associated with increased serum triglycerides and LDL cholesterol levels [15], reduced response to lipid-lowering therapy with fenofibrate [16], as well as the development of nonalcoholic fatty liver disease (NAFLD)[17]. However, little is known about the association of polymorphisms in the promoter region of the FABP1gene with lipid metabolism.
In the present study, common mutations in the promoter of the FABP1 gene responsible for alterations in serum TG levels was identified. Furthermore, the association of serum TG levels with altered FABP1 levels attributed to this mutation in the promoter region was analyzed.
Materials and Methods
Subjects
Population Selection
The association between the promoter polymorphism and the risk for dyslipidemia in the Han population was assessed using a cross-sectional survey. A total of 1,182 subjects (male/female: 817/365, aged 18–72 years) were recruited from among individuals visiting the Union Hospital of Fujian Medical University for regular medical check-ups between August 2012 and January 2013. Participants were selected according to the following criteria, 1) absence of prior CVD, severe acute disease, Type 1 diabetes or pregnancy; 2) had not received lipid-lowering treatment or any other drug-modifying lipid measures. In addition, secondary causes of lipid disorders were excluded in all subjects based on clinical history and blood tests. All subjects enrolled were of Chinese Han ethnicity. This study was conducted in accordance with the guidelines of the 1975 Declaration of Helsinki and was approved by the Ethics Committee of the Fujian Medical University. Each subject gave written informed consent before participation in the study.
Data Collection
Interview
A pilot-tested structured questionnaire was administered by trained interviewers to collect information on demographic characteristics (e.g. age, education level, job, marital status, sex), as well as relevant information on smoking habits, tea and alcohol drinking habits, physical exercise, personal medical history, and potential others risk factors. Individuals reporting regular tobacco use in the previous 6 months were classified as smokers. Alcohol and tea drinkers were defined as those who had drunk alcohol/tea at least once per week for more than 6 months. Physical activity was ascertained by asking participants about their workplace physical activity and how often in the previous month they had engaged in any number of different activities, including walking, jogging, cycling and swimming. The participants were grouped into two broad categories based on their total score: inactive (lower physical activity) and active (higher physical activity).
Anthropometric Measurements and Laboratory Evaluation
Weight and standing height were measured in a standardized fashion by a trained examiner. Body mass index (BMI) was calculated as the ratio of weight (kilograms) to the square of height (meters). Systolic and diastolic blood pressure was determined in two consecutive measurements on the upper arm using an automated sphygmomanometer (HEM- 907, Omron, Kyoto, Japan) with subjects in a seated, resting position; The mean of blood pressure was used for analysis. An overnight fast venous blood samples were drawn after an anthropometric examination. The levels of total cholesterol (TC), high-density lipoprotein cholesterol (HDL-C) and low density lipoprotein cholesterol (LDL-C), triglycerides (TGs), fasting plasma glucose (FPG), and uric acid (UA), in addition to liver function tests, were measured by standard clinical laboratory techniques, which have been described previously [17].
Genotype Analysis
The genomic region harboring FABP1 promoter was examined to select single nucleotide polymorphisms (SNPs) based on the linkage disequilibrium (LD) patterns of Chinese Han population in Beijing (CHB) from Hapmap database. Then we used the pairwise tagging method of the Haploview v4.2 software (Broad institute, Cambridge, MA, USA) to capture SNPs with a minimum minor allele frequency (MAF) of >0.05 and a minimum r2 of >0.8. As result, four SNPs (rs2919872, rs2970901, rs2970902, and rs2970903) were selected from unrelated Han Chinese individuals in Beijing. Finally, widely used software for TF binding site prediction MatInspector and CHIP Mapper[18], were used to predict possible transcription changes due to the presence of a variant.
DNA was isolated from EDTA-treated blood samples following standard procedures. These polymorphisms were genotyped by iPlex technology based on a MassARRAY platform in all subjects. Primers for the amplification and extension reactions were designed using the Mass Array Assay Design Version 3.1 software (Sequenom, San Diego, CA, USA), and SNP genotypes were determined according to the iPLEX protocol provided by the manufacturer. Genotyping assays were performed by laboratory personnel who were blinded to the lipid status of each subject. The genotyping quality was examined in a detailed QC procedure consisting of a >95% successful call rate, duplicate calling of genotypes, internal positive control samples and Hardy–Weinberg Equilibrium (HWE) testing.
FABP1 Measurement
Serum FABP1 levels were measured in a subset of subjects with different FABP1 rs2919872 genotypes. Subjects with impaired glucose metabolism (indicated by an oral glucose tolerance test), abnormal liver function, a history of smoking, alcohol drinking, or tea drinking were excluded. There were no significant differences in the age and sex distributions of the subjects in this subset compared with those of the total subject population. All evaluations were performed on an aliquot of serum collected after overnight fasting and stored at −80°C. Serum FABP1 levels were evaluated in 95 subjects with different rs2919872 genotypes using the commercially available FABP1 ELISA kit (GWB-HFABP1, GeneWay Biotech, San Diego, CA, USA) according to the manufacturer’s instructions.
FABP1 Promoter Luciferase Reporter Constructs
The pGL3-rs2919872G plasmid harboring the human FABP1 promoter with the rs2919872 G allele was constructed by ligation of the PCR-generated full length human FABP1 promoter (nucleotides –2125 to +50, relative to the transcription start site) into the pGL3-Basic luciferase reporter plasmid (Promega, Madison, WI, USA), as described previously in our study (also designated pGL3B–2125) [19]. Fusion PCR was performed to create the rs2919872 A allele of the FABP1 promoter (designated pGL3- rs2919872A) using pGL3-rs2919872G as a template. Briefly, two pairs of primers, i.e., P1/P3 and P2/P4, were used to amplify the fragments flanking the site of rs2919872A. The two amplified fragments were annealed and fused by PCR with P1 and P4. The fused fragment was digested with Tth111I and BstAP1and used to replace the Tth111I-BstAP1 fragment of pGL3-rs2919872G. The amplified sequences were confirmed by DNA sequencing. The primers used were: P1 (forward): 5′ TGCCCGCTGTTCAGGTAGTC 3′, P2 (forward): 5′ TGAGGGGGTGCTTGTAAAGAGCTGCCTCAGAGGCAG 3′, P3 (reverse): 5′ TCTTTACAAGCACCCCCTCA 3′, P4 (reverse): 5′ ACAAGTGTGT GGGTGCATGT G 3′.
Cell Culture and Transfection and Dual-luciferase Reporter Assay
The human hepatoblastoma cell line HepG2 (HB–8065, ATCC, VA, USA) and the hepatoma cell line Huh7 (JCRB0403, Japan) were maintained in Dulbecco’s modified Eagle medium (DMEM, Invitrogen) supplemented with 10% (v/v) fetal bovine serum (FBS, Invitrogen) at 37°C in a humidified atmosphere containing 5% CO2. Cells were seeded (2×105 cells/well) in 12-well plates and transfected with the empty pGL3-Basic vector (a promoterless control) or with pGL3- rs2919872G or pGL3- rs2919872 A constructs using Lipofectamine 2000 (Invitrogen). The pRL-SV40 plasmid (Promega), containing Renilla reniformis luciferase, was cotransfected into cells as a normalizing control. For each plasmid, triplicate transfections were performed in three independent transfection experiments. After 48 h, intracellular luciferase activity was determined in a total of 20 μg of cell lysate using the Dual-Luciferase Reporter Assay System (Promega) according to the manufacturer’s recommendations. Luminescence measurement was carried out on a luminometer (Orion II Microplate Luminometer, Berthold Detection Systems, Germany). The relative luciferase units (RLU) were determined by comparison with the promoterless pGL3-Basic plasmid, which was assigned an arbitrary value of 1.
Statistical Analysis
Coefficients of skewness and kurtosis were calculated to test deviation from a normal distribution. Logarithmic transformation was performed on the individual values of skewed variables, and a normal distribution of transformed values was confirmed before statistical analysis and significance testing. Unless otherwise stated, data are presented as means ± standard deviation (SD) for continuous variables and as frequencies or percentages for categorical variables. Differences in mean values were assessed using unpaired t-tests, one-way ANOVA or the Mann–Whitney U-test, as appropriate. Categorical variables were compared using χ 2 test. For each SNP, Hardy–Weinberg equilibrium (HWE) was checked in subjects using the χ 2 test. All tests were performed on the basis of an additive, dominant or recessive model. Previous studies have suggested that the multiple testing correction may hid many true-positive loci[20], and reduce the number of tests may be advantageous for finding true-positive loci [21,22]. Therefore, P-values were not adjusted for multiple tests in the present study.
In addition, multiple linear regression was performed to examine the independent predictors of serum TG or FABP1 concentrations with adjustment for age, sex, income, marital status, education, smoking, tea drinking, BMI and other clinical features. Before statistical analysis, raw TG and FABP1 level phenotype data were transformed into natural log (ln) values to overcome high-value deviation. All analyses were performed using the SPSS for Windows statistical package, Version 18.0 (SPSS). For all tests, P-values less than 0.05 were considered to indicate statistical significance.
Results
Subject Status and General Data
The study population included 817 males and 365 females, with a mean age of 45.06 ± 12.93 years (18–72 years). Clinical features, anthropometric variables and laboratory findings for all individuals are shown in Table 1. There were no significant differences in the distributions of LDL-C, AST, marital status, education or income between males and females included in the study (all P > 0.05). Compared with females, males showed significantly higher levels of BMI, SABP, DABP, ALT, and TG, and lower levels of TC, FPG, and HDL-C (all P < 0.05). In addition, the frequencies of active physical activity, smoker and alcohol drinkers were higher frequency among the male subjects than among the female subjects.
Table 1. Clinical characteristic of 1,182 participants.
| Variables | All cases (n = 1182) | Men (n = 817) | Women (n = 365) | P |
|---|---|---|---|---|
| Age(years; mean ± SD) | 45.06±12.93 | 43.56±13.36 | 48.42±11.22 | <0.01 |
| BMI(kg/m2; mean ± SD) | 23.80±3.00 | 24.08±3.03 | 23.20±3.10 | <0.01 |
| SABP(mmHg; mean ± SD) | 76.72±10.72 | 77.44±10.66 | 75.13±10.69 | <0.01 c |
| DABP(mmHg; mean ± SD) | 124.27±13.17 | 125.14±12.98 | 122.33±13.40 | <0.01 c |
| AST (IU/L; mean ± SD) | 24.58±25.94 | 25.56±30.32 | 22.38±10.77 | .051 c |
| ALT(IU/L; mean ± SD) | 31.57±58.13 | 35.05±67.88 | 23.78±23.41 | <0.01 c |
| TC (mmol/L; mean ± SD) | 5.23±1.08 | 5.18±1.06 | 5.34±1.11 | .024 c |
| TG (mmol/L; mean ± SD) | 1.54±1.02 | 1.62±1.03 | 1.35±1.00 | <0.01 c |
| FPG (mmol/L; mean ± SD) | 5.60±1.22 | 5.58±1.15 | 5.65±1.36 | <0.01 c |
| HLD-C (mmol/L; mean ± SD) | 1.49±0.61 | 1.43±0.68 | 1.62±0.40 | <0.01 c |
| LDL-C (mmol/L; mean ± SD) | 3.29±1.10 | 3.28±1.08 | 3.32±1.13 | 0.557 c |
| Per capita incoming | >0.05 b | |||
| Low, n(%) | 314 (26.54) | 192 (23.50) | 122 (33.33) | |
| Middle, n(%) | 486 (41.10) | 312 (38.19) | 174 (47.62) | |
| High, n(%) | 383 (32.36) | 313 (38.31) | 70 (19.05) | |
| Marriage(married), n(%) | 1017 (86.04) | 663 (81.15) | 354 (96.99) | >0.05 b |
| Smoking status(smoking) | 307 (26.00) | 302 (36.96) | 5 (1.45) | <0.01 |
| Tea drinking(drinking) | 589 (49.87) | 436 (53.37) | 153 (42.03) | |
| Alcohol drinking(drinking) | 279 (23.64) | 272 (33.29) | 7 (2.04) | <0.01 |
| Physical activity(active) | 316(38.68) | 105(28.77) | <0.01 | |
| Education, n(%) | >0.05 b | |||
| ≤6 years | 548 (46.35) | 405 (49.57) | 143 (39.13) | |
| ≤9 years | 506 (42.80) | 326 (39.90) | 180 (49.28) | |
| >9 years | 128 (10.86) | 86 (10.53) | 42 (11.59) |
Continuous variables are expressed as mean ± SD; Categorical variables are expressed as frequency (percent). n: number of individuals; SD: standard deviation.
P-values a: obtained by Student's t-test for normal distributed continuous variables
b: obtained by Pearson Chi square test for categorical variables
c: obtained by Mann-Whitney U-test for continuous variables deviation from a normal distribution.
Analysis of the Association of Various Quantitative Phenotypes with the FABP1 Gene Promoter SNPs
The promoter SNPs information of FABP1 gene was presented in Fig 1[19,23]. These SNPs were genotyped in all 1,182 individuals using iPlex technology based on a MassARRAY platform, and were commonly distributed in the study samples. The corresponding frequencies of the rs2970902G, rs2919872A, rs2970903A, and rs2970901A alleles among the participants were 0.160, 0.234, 0.053 and 0.232, respectively, and all of the tested SNPs were in Hardy–Weinberg equilibrium (P > 0.05).
Fig 1. SNPs in the FABP1 gene promoter.

A. Schematic diagram showing the positions of SNPs and known binding sites for nuclear factors. Four SNPs (SNP1 to SNP4), located at -2,055 nt, -1,647 nt, -1,298 nt, and -1,036 nt respectively, were genotyped within the FABP1 promoter (-2,125 nt/+51 nt). The known binding sites for C/EBPα (CCAAT-element binding protein alpha), HNF3β (human nuclear factor beta), DR1-C/EBP (direct repeat with 1 base spacing/CCAAT-element binding protein), PPARα (peroxisome proliferator-activated receptor alpha) in the FABP1 gene promoter have been reported previously [19,23]. B. SNP information. SNP1 to SNP4 are listed from 5′–3′ of the FABP1 promoter. “CHR-ID” indicates the chromosome ID of the four SNPs in the FABP1 gene promoter. “RS ID”, reported in UCSC or NCBI website. “Function” indicates the altered genetically-coded function of the SNPs. “Ref SNP” indicates SNP form in the original reference sequence. “mRNA location” corresponding mRNA positions are shown for each SNP. NCBI db is available from http://www.ncbi.nlm.nih.gov/SNP.
Given that the involvement of FABP1 in lipid metabolism, we investigated the association of variants in the promoter sequence of this gene with altered serum lipid levels. The single allelic analysis showed that rs2919872 was associated with serum TG levels (P = 0.032, Table 2). In allele-specific phenotype analysis, the mean serum TG levels associated with rs2919872 G>A suggested that the A allele decreases the levels of serum TG, with AA homozygotes having a 14.47% decrease in serum TG levels (1.36 mmol/L vs. 1.59 mmol/L) relative to homozygotes for the GG allele and this effect was apparently recessive. We found no significant associations between rs2919872 and other lipid characteristics including TC, HDL-C and LDL-C. Among the other three tested polymorphisms (rs2970901, rs2970902, and rs2970903), no significant associations were identified with either serum TG levels or other clinical characters (data not shown). It was noteworthy that FABP1 SNPs (rs2919871, rs1441644, rs2241883, rs2970902, rs2970903 and rs2919867) were previously reported not to be associated with lipid levels in the European population [24]. However, these SNPs are not in high linkage disequilibrium (LD) with the SNP rs2919872.
Table 2. Associations of the FABP1 gene promoter variant rs2919872G>A with investigated parameters.
| Quantitative phenotype | GG (n = 705) | GA (n = 401) | AA (n = 75) | P c add | P d dom | P e Recessiv |
|---|---|---|---|---|---|---|
| Age (years) a | 44.77±12.61 | 45.49±13.31 | 45.31±14.15 | 0.667 | 0.372 | 0.855 |
| SABP (mm Hg) b | 124.41±13.43 | 123.46±12.59 | 127.23±13.65 | 0.065 | 0.688 | 0.051 |
| DABP (mm Hg) b | 76.82±10.44 | 76.14±11.02 | 78.94±11.55 | 0.103 | 0.728 | 0.061 |
| BMI (kg/m2) a | 23.77±2.98 | 23.86±3.23 | 24.09±3.28 | 0.639 | 0.461 | 0.423 |
| HDL-C (mg/dL) b | 1.49±0.54 | 1.49±0.76 | 1.48±0.34 | 0.993 | 1.000 | 0.911 |
| LDL-C (mg/dL) b | 3.29±1.13 | 3.30±1.03 | 3.31±1.16 | 0.982 | 0.884 | 0.871 |
| TC (mg/dL) b | 5.26±1.11 | 5.17±1.02 | 5.24±1.11 | 0.438 | 0.232 | 0.954 |
| TG (mg/dL) b | 1.59±1.09 | 1.51±0.98 | 1.36±0.43 | 0.193 | 0.266 | 0.032 |
| FPG (mg/dL) b | 5.53±0.82 | 5.72±1.74 | 5.66±1.07 | 0.057 | 0.211 | 0.637 |
| ALT (IU/L) b | 30.22±33.28 | 34.75±90.03 | 28.73±22.89 | 0.425 | 0.312 | 0.648 |
| AST (IU/L) b | 24.36±18.29 | 25.35±37.67 | 22.86±8.46 | 0.693 | 0.707 | 0.545 |
Continuous variables are expressed as mean ± standard deviation (SD); n: number of individuals
P-values a: obtained by one-way ANOVA
b: obtained by Kruskal–Wallis test or Mann-Whitney U-test
c additive model: GG vsGA. GG vs AA, and GA vs. AA
d:a dominant model: GG vs AA+GA
e: a recessive model (GA+GG vs.AA)
The effects of rs2919872 genotype on TG levels were further assessed using multiple linear regression models with adjustment for age, sex, income, marital status, education, smoking, tea drinking, BMI and other clinical features. TG values were defined as the dependent variable and were log-transformed before statistical analysis. In stepwise regression analysis, rs2919872G>A (recessive model) (β = -0.0515, P < 0.05), BMI (β = 0.134, P < 0.001), TC (β = 0.437, P < 0.001), HDL-C (β = -0.243, P < 0.001), LDL-C (β = -0.209, P < 0.001), SBP (β = 0.123, P < 0.001), and ALT (β = 0.063, P < 0.05), in this order, were independently associated with serum logTG concentrations (Table 3), explaining a total of 29.9% of its variability. The results were similar when age, sex, fasting glucose, income, marital status, education, smoking, alcohol drinking and tea drinking were added to the model.
Table 3. Variables independently associated with plasma TG levels by multivariate lineal regression analysis.
| Variables | Standardized coefficient(β) | P | Adjusted R2 |
|---|---|---|---|
| Model 1 a | |||
| rs2919872G>A (recessive model) | -0.061 | 0.037 | 0.003 |
| Model 2 b | 0.299 | ||
| rs2919872G>A (recessive model) | -0.051 | 0.042 | |
| BMI | 0.134 | 0.000 | |
| TC | 0.437 | 0.000 | |
| HDL-C | -0.243 | 0.000 | |
| UA | 0.201 | 0.000 | |
| LDL-C | 0.209 | 0.000 | |
| SABP | 0.123 | 0.000 | |
| ALT | 0.063 | 0.011 | |
| Model 3 c | 0.303 | ||
| rs2919872G>A (recessive model) | -0.049 | 0.046 | |
| Others investigated characteristics |
a: Only the variable rs2919872G>A was entered into this model
b: Variables were selected by a forward stepwise selection procedure, in which variables were sequentially entered into this model.
c: All variables were entered into this model in a single step.
Functional Relevance of rs2919872
Variations in regulatory sequences might influence gene expression following binding of transcriptional activators or inhibitors that instruct their regulatory control. Therefore, we examined the effect of the G>A polymorphism at rs2919872 on FABP1 promoter activity. To address this issue, the full length human FABP1 promoter (from −2125 to +51, relative to the transcription start site), with either the G or A at position rs2919872, was cloned into the pGL3-basic luciferase reporter plasmid. These two constructs, i.e., pGL3-rs2919872G and pGL3-rs2919872A, were then transfected separately into HepG2 and Huh7 hepatoma cells. As shown in Fig 2, cells transfected with pGL3-rs2919872G displayed much higher luciferase activity than that of cells transfected with pGL3-rs2919872A, suggesting that the rs2919872 A allele dramatically reduced FABP1 promoter activity.
Fig 2. FABP1 promoter activities in A. HepG2 cells and B. Huh7 cells.
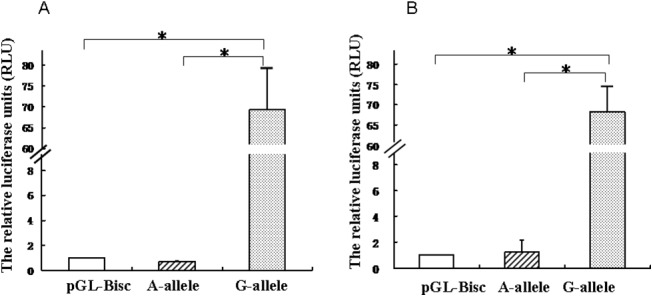
Cells were co-transfected with 10 ng of the Renilla luciferase expression vector pRL-SV40 and 0.2 μg each of the pGL3-rs2919872G and pGL3-rs2919872A plasmids; the promoterless pGL3-Basic vector served as the negative control. Intracellular luciferase activity was measured 48 h after transfection. The relative luciferase units (RLU) were determined by comparison with the promoterless pGL3-Basic plasmid, which was assigned an arbitrary value of 1. Each transfection was performed in duplicate and the data are expressed as the mean ± SD of three separate experiments. (*P < 0.05).
Correlation of Serum FABP1 Concentration with the rs2919872 G>A Polymorphism in FABP1 Promoter
Serum FABP1 levels correlate closely with those in the liver, which is main site of FABP1 expression [15]. To investigate the association between the rs2919872 genotype and serum FABP1 levels in our study population, FABP1 levels were evaluated by ELISA in serum samples from 95 individuals with different rs2919872A or rs2919872G alleles. There were no significant differences in the distributions of age, sex, marital status, education or income among the different rs2919872 genotypes (all P > 0.05). As shown in Fig 3, subjects with the rs2919872 AA genotype had significantly lower serum FABP1 concentrations (geometric mean ± geometric standard deviation) than those carrying the GG genotypes (GG genotype: 13.67 ± 2.60 ng/mL, n = 53 versus AA genotype: 5.13 ± 4.38 ng/mL, n = 10, P < 0.01). Multiple linear regression analysis to predict the variables independently associated with serum FABP1 showed that, after adjustment for all the covariables in Table 4, rs2919872 A allele was negatively (β = -0.331, P = 0.003) associated with serum logFABP1 concentrations, while serum TG levels were positively (β = 0.487, P = 0.014) associated with serum logFABP1 concentrations(Table 4). These results suggested that rs2919872 is associated with TG levels by modulating FABP1 expression.
Fig 3. Serum FABP1 levels in healthy individuals with different FABP1 rs2919872 genotypes.

Serum FABP1 levels (geometric mean ± geometric standard deviation) were measured among the subjects with different genotypes of the FABP1 rs2919872G>A (GG genotype: 13.67 ± 2.60 ng/mL (n = 53); GA genotype: 9.44 ± 2.29 ng/mL (n = 32); AA genotype: 5.13 ± 4.38 ng/mL (n = 10); *P < 0.01)
Table 4. Variables independently associated with plasma FABP1 levels by multivariate lineal regression analysis.
| Variables | Standardized coefficient(β) | P | Adjusted R2 |
|---|---|---|---|
| 0.250 | |||
| rs2919872 G>A (recessive model) | -0.320 | 0.003 | |
| TG | 0.487 | 0.014 | |
| ALT | 0.045 | 0.837 | |
| AST | -0.018 | 0.926 | |
| SABP | 0.140 | 0.511 | |
| DABP | -0.136 | 0.512 | |
| HDL-C | 0.059 | 0.761 | |
| LDL-C | 0.680 | 0.151 | |
| TC | 0.066 | 0.209 | |
| BMI | -0.046 | 0.746 | |
| Age | -0.118 | 0.349 | |
| Sex | 0.230 | 0.051 |
All variables were entered into this model in a single step.
Discussion
In this study, we investigated the common polymorphisms located in the promoter region of the FABP1 gene that may affect lipid metabolism by influencing FABP1 gene transcription. To the best of our knowledge, this is the first study to examine the association of FABP1 promoter polymorphisms with lipid levels. In this study, decreased levels of serum FABP1 were observed in subjects with the rs2919872 A allele, indicating that a G>A transition at rs2919872, which is located in the promoter region of the FABP1 gene is responsible for a major effect on serum TG levels in a Chinese Han population. Furthermore, in vitro experiments indicated that the FABP1 rs2919872 A allele decreased the transcriptional activity of the FABP1 gene, probably due to the disturbance of a transcription factor binding site, suggesting the functional relevance of this site.
FABP1 is a key regulator of hepatic lipid metabolism and is required for optimal activity of transacylase enzymes in the murine TG synthesis pathway. Overexpression of murine FABP1 is associated with increased LCFA uptake and hepatic TG levels [25,26,27], while FABP1 −/− mice exhibit decreased hepatic triglyceride content with altered FA uptake kinetics [12], with protection against diet-induced obesity and hepatic steatosis [14,28]. In humans, amino acid variations in FABP1 could be functionally relevant. The functional mutation rs2241883 (p.Thr94Ala) in the FABP1 gene has been extensively studied, and the Ala94/Ala polymorphism shows a significant association with increased serum LDL-C and TG levels[15], as well as decreased response to lipid-lowering therapy with fenofibrate (a cholesterol synthesis inhibitor) and glycogenolysis [16,29,30]. To date, little is known about the potential influence of nucleic acid variants in the FABP1 gene promoter on lipid metabolism. In the present study, we analyzed four polymorphisms that appeared to be relevant in the regulation of FABP1 expression. We found that only the rs2919872 A allele contributed significantly to reduced serum TG levels in a Chinese population. Furthermore, regression analyses also showed a similar independent inverse association of the FABP1 rs2919872 G>A variant with TG levels and that, together with the BMI, the concentrations of TC, HDL-C, LDL-C, SBP, and ALT, explained approximately 30% of TG variability. The increased minor allele frequency in populations with lower TG levels suggests that the A allele of the rs2919872 of the FABP1 gene may influence FABP1 expression and protect against hypertriglyceridemia. This hypothesis is further supported by physiological data as well as by genetic findings. Functional assays using the luciferase gene as a reporter that were performed as part of this study showed that the A allele almost abolished the promoter activity of FABP1 gene compared with effect of the G allele. In addition, we also found that healthy individuals carrying with the A allele had a considerably lower serum FABP1 concentration than the G allele carriers. Furthermore, regression analyses showed that rs2919872 G>A was negatively associated with serum FABP1 concentrations, while serum TG levels were positively associated with serum FABP1 concentrations after controlling for potential confounding factors. It was noteworthy that eQTL data analysis showed that only one SNP (rs2241883) was reported to be associated with FABP1 mRNA levels, however, this SNP was not in high LD with rs2919872.The observed phenotype shows obvious conformity with that of FABP1 knockout mice [12,14,28], indicating that the rs2919872 G>A replacement in FABP1 is likely to be a ‘loss-of-function’ variant analogous to the FABP1 null mouse. It is noteworthy that our findings are also consistent with those obtained for the rs2241883 (p.Thr94Ala) polymorphism. The FABP1 T94A substitution in this variant was shown to be associated with upregulation of total FABP1, which in turn may stimulate enzymes in the TG synthesis pathway and lead to the elevated plasma TGs in human subjects [15,31].
The association of downregulated FABP1 with protection against high TG may be explained in two ways. First, because FABP1 is involved in the budding of pre-chylomicron transport vesicles from the endoplasmic reticulum of the enterocyte [32], downregulation of FABP1 would be expected to result in subtle differences in intestinal FA trafficking and delayed appearance of dietary TG in the serum [12,14]. Second, FABP1 is abundantly expressed in both hepatocytes and enterocytes and binds to multiple ligands, including saturated FA and cholesterol[12]; therefore downregulation of FABP1 would result in impaired FA uptake, decreased production of very low density lipoproteins, and reduced hepatic TG accumulation.
The mechanisms involved in the downregulation of FABP1 by the G-to-A substitution at rs2919872 in the promoter region remain to be elucidated. However, it can be speculated that the rs2919872 G>A mutation either generates a binding site for a negative regulator or abolishes the existing binding site for a positive regulator. However, besides the MatInspector and CHIP Mapper, searches of the TESS (http://www.cbil.upenn.edu/cgi-bin/tess/tess) and TFSEARCH (http://www.cbrc.jp/research/db/TFSEARCH.html) databases did not reveal any potential binding sites changes for transcription factors due to the presence of rs2919872 A(data not shown). Therefore, further DNA pull-down and yeast one hybrid screening studies are required to identify the possible function of rs2919872 G>A.
Several limitations in our study need to be discussed. Firstly, in the present study, although the rs2919872 variant affected transcription efficiency of the FABP1’s promoter and the levels of FABP1 protein, FABP1 mRNA levels for each SNP allele or genotype were not measured in our population. However, this limitation is unavoidable, because for ethical reasons it is hard for us to obtain liver tissue even the biopsy samples from individuals taking a regular medical health check-up. Secondly, the SNPs were chosen to maximize SNP tagging for genetic variation rather than for the functionality of the FABP1 gene in the present study. Although the luciferase data indicated that only rs2919872 SNP was functional, it did not exclude the possibility that other SNPs in high LD may be responsible for the observed associations, the exact mechanisms linking rs2919872 A allele to lower serum TG levels in humans needs further studies. Thirdly, the weak P value of association in the initial population (P = 0.032) was not corrected for multiple testing. Multiple testing correction is necessary to exclude false-positive loci, however, it may simultaneously hid many true-positive loci which were biologically associated with clinical traits[20,21]. Therefore, these reports suggested that reducing the number of tests may be advantageous for finding true-positive loci [21,22]. Finally, our study firstly reported the relationship between SNPs (rs2919872) in FABP1 gene and serum TG levels in Chinese Han subjects. It is unclear whether the present findings can be generalized to other ethnicities. Thus, only specific testing of putative causal variants directly in extensive cohorts and population-based samples will help validate the role of FABP1 rs2919872 in lipid metabolism.
In summary, the results of this study demonstrate that rs2919872 G>A, a common polymorphism in the promoter region of FABP1, may be of functional importance in regulating the expression of the FABP1 gene and might influence the serum TG concentrations in a Chinese Han population. Considering the association of increased serum TG levels with increased risk of metabolic syndrome and CVD [2,33,34], this variant still represents an interesting potential target for future lipid-lowering therapies to reduce the risk of CVD.
Acknowledgments
We thank all study participants for their cooperation and also acknowledge the efforts of our recruiting and technical staff.
Abbreviations
- ALT
alanine aminotransferase
- AST
aspartate aminotransferase
- BMI
body mass index
- CVD
cardiovascular disease
- DABP
diastolic arterial blood pressure
- FABP1
fatty acid-binding protein 1
- FPG
fasting plasma glucose
- HWE
Hardy–Weinberg Equilibrium
- HDL-C
high density lipoprotein cholesterol
- LD
linkage disequilibrium
- LDL-C
low density lipoprotein cholesterol
- L-FABP
Liver fatty acid-binding protein
- NAFLD
nonalcoholic fatty liver disease
- SABP
systolic arterial blood pressure
- SNP
single-nucleotide polymorphisms
- TC
total cholesterol
- TG
triglyceride
Data Availability
All relevant data are within the paper.
Funding Statement
This work was supported by the National Natural Science Foundation of China,(http://www.nsfc.gov.cn/), Grant Number:No. 81473047 to X-EP, the National Natural Science Foundation of China,(http://www.nsfc.gov.cn/), Grant Number:No.81271822 to XL, the Natural Science Foundation of Fujian province (http://xmgl.fjkjt.gov.cn/), Grant Number: No. 2015J01303 to X-EP, the Training Program Foundation for Middle-aged and Young Talents from Sanitation System of Fujian province (http://www.fjhfpc.gov.cn/), Grant Number:No. 2014-ZQN-ZD-23 to X-EP, the New Century Excellent Talents in Fujian Province University, Grant Number:No. JA11106 and No. JA14127 to X-EP and the Foundation of Fujian Educational Committee (http://www.fjedu.gov.cn/), Grant Number:No. JK2013021 to Y-LW. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Degoma EM, Rader DJ (2011) Novel HDL-directed pharmacotherapeutic strategies. Nat Rev Cardiol 8: 266–277. 10.1038/nrcardio.2010.200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Nordestgaard BG, Varbo A (2014) Triglycerides and cardiovascular disease. Lancet 384: 626–635. 10.1016/S0140-6736(14)61177-6 [DOI] [PubMed] [Google Scholar]
- 3. Eeg-Olofsson K, Gudbjornsdottir S, Eliasson B, Zethelius B, Cederholm J (2014) The triglycerides-to-HDL-cholesterol ratio and cardiovascular disease risk in obese patients with type 2 diabetes: an observational study from the Swedish National Diabetes Register (NDR). Diabetes Res Clin Pract 106: 136–144. 10.1016/j.diabres.2014.07.010 [DOI] [PubMed] [Google Scholar]
- 4. Miller M, Stone NJ, Ballantyne C, Bittner V, Criqui MH, et al. (2011) Triglycerides and cardiovascular disease: a scientific statement from the American Heart Association. Circulation 123: 2292–2333. 10.1161/CIR.0b013e3182160726 [DOI] [PubMed] [Google Scholar]
- 5. Zhang Y, Smith EM, Baye TM, Eckert JV, Abraham LJ, et al. (2010) Serotonin (5-HT) receptor 5A sequence variants affect human plasma triglyceride levels. Physiol Genomics 42: 168–176. 10.1152/physiolgenomics.00038.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Kathiresan S, Willer CJ, Peloso GM, Demissie S, Musunuru K, et al. (2009) Common variants at 30 loci contribute to polygenic dyslipidemia. Nat Genet 41: 56–65. 10.1038/ng.291 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Willer CJ, Sanna S, Jackson AU, Scuteri A, Bonnycastle LL, et al. (2008) Newly identified loci that influence lipid concentrations and risk of coronary artery disease. Nat Genet 40: 161–169. 10.1038/ng.76 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Bass NM, Manning JA (1986) Tissue expression of three structurally different fatty acid binding proteins from rat heart muscle, liver, and intestine. Biochem Biophys Res Commun 137: 929–935. [DOI] [PubMed] [Google Scholar]
- 9. Pelsers MM, Namiot Z, Kisielewski W, Namiot A, Januszkiewicz M, et al. (2003) Intestinal-type and liver-type fatty acid-binding protein in the intestine. Tissue distribution and clinical utility. Clin Biochem 36: 529–535. [DOI] [PubMed] [Google Scholar]
- 10. Petrescu AD, McIntosh AL, Storey SM, Huang H, Martin GG, et al. (2013) High glucose potentiates L-FABP mediated fibrate induction of PPARalpha in mouse hepatocytes. Biochim Biophys Acta 1831: 1412–1425. 10.1016/j.bbalip.2013.05.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Veerkamp JH, van Moerkerk HT (1993) Fatty acid-binding protein and its relation to fatty acid oxidation. Mol Cell Biochem 123: 101–106. [DOI] [PubMed] [Google Scholar]
- 12. Newberry EP, Xie Y, Kennedy S, Han X, Buhman KK, et al. (2003) Decreased hepatic triglyceride accumulation and altered fatty acid uptake in mice with deletion of the liver fatty acid-binding protein gene. J Biol Chem 278: 51664–51672. [DOI] [PubMed] [Google Scholar]
- 13. Chen A, Tang Y, Davis V, Hsu FF, Kennedy SM, et al. (2013) Liver fatty acid binding protein (L-Fabp) modulates murine stellate cell activation and diet-induced nonalcoholic fatty liver disease. Hepatology 57: 2202–2212. 10.1002/hep.26318 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Newberry EP, Xie Y, Kennedy SM, Luo J, Davidson NO (2006) Protection against Western diet-induced obesity and hepatic steatosis in liver fatty acid-binding protein knockout mice. Hepatology 44: 1191–1205. [DOI] [PubMed] [Google Scholar]
- 15. Fisher E, Weikert C, Klapper M, Lindner I, Mohlig M, et al. (2007) L-FABP T94A is associated with fasting triglycerides and LDL-cholesterol in women. Mol Genet Metab 91: 278–284. [DOI] [PubMed] [Google Scholar]
- 16. Brouillette C, Bosse Y, Perusse L, Gaudet D, Vohl MC (2004) Effect of liver fatty acid binding protein (FABP) T94A missense mutation on plasma lipoprotein responsiveness to treatment with fenofibrate. J Hum Genet 49: 424–432. [DOI] [PubMed] [Google Scholar]
- 17. Peng XE, Wu YL, Lu QQ, Hu ZJ, Lin X (2012) Two genetic variants in FABP1 and susceptibility to non-alcohol fatty liver disease in a Chinese population. Gene 500: 54–58. 10.1016/j.gene.2012.03.050 [DOI] [PubMed] [Google Scholar]
- 18. Riva A (2012) The MAPPER2 Database: a multi-genome catalog of putative transcription factor binding sites. Nucleic Acids Res 40: D155–161. 10.1093/nar/gkr1080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Wu YL, Peng XE, Wang D, Chen WN, Lin X (2012) Human liver fatty acid binding protein (hFABP1) gene is regulated by liver-enriched transcription factors HNF3beta and C/EBPalpha. Biochimie 94: 384–392. 10.1016/j.biochi.2011.08.006 [DOI] [PubMed] [Google Scholar]
- 20. Fransen K, Visschedijk MC, van Sommeren S, Fu JY, Franke L, et al. (2010) Analysis of SNPs with an effect on gene expression identifies UBE2L3 and BCL3 as potential new risk genes for Crohn's disease. Hum Mol Genet 19: 3482–3488. 10.1093/hmg/ddq264 [DOI] [PubMed] [Google Scholar]
- 21. Hong KW, Jin HS, Lim JE, Cho YS, Go MJ, et al. (2010) Non-synonymous single-nucleotide polymorphisms associated with blood pressure and hypertension. J Hum Hypertens 24: 763–774. 10.1038/jhh.2010.9 [DOI] [PubMed] [Google Scholar]
- 22. Hong KW, Lim JE, Oh B (2011) A regulatory SNP in AKAP13 is associated with blood pressure in Koreans. J Hum Genet 56: 205–210. 10.1038/jhg.2010.167 [DOI] [PubMed] [Google Scholar]
- 23. Guzman C, Benet M, Pisonero-Vaquero S, Moya M, Garcia-Mediavilla MV, et al. (2013) The human liver fatty acid binding protein (FABP1) gene is activated by FOXA1 and PPARalpha; and repressed by C/EBPalpha: Implications in FABP1 down-regulation in nonalcoholic fatty liver disease. Biochim Biophys Acta 1831: 803–818. 10.1016/j.bbalip.2012.12.014 [DOI] [PubMed] [Google Scholar]
- 24. Brahe LK, Angquist L, Larsen LH, Vimaleswaran KS, Hager J, et al. (2013) Influence of SNPs in nutrient-sensitive candidate genes and gene-diet interactions on blood lipids: the DiOGenes study. Br J Nutr 110: 790–796. 10.1017/S0007114512006058 [DOI] [PubMed] [Google Scholar]
- 25. Murphy EJ, Prows DR, Jefferson JR, Schroeder F (1996) Liver fatty acid-binding protein expression in transfected fibroblasts stimulates fatty acid uptake and metabolism. Biochim Biophys Acta 1301: 191–198. [DOI] [PubMed] [Google Scholar]
- 26. Murphy EJ (1998) L-FABP and I-FABP expression increase NBD-stearate uptake and cytoplasmic diffusion in L cells. Am J Physiol 275: G244–249. [DOI] [PubMed] [Google Scholar]
- 27. Wolfrum C, Buhlmann C, Rolf B, Borchers T, Spener F (1999) Variation of liver-type fatty acid binding protein content in the human hepatoma cell line HepG2 by peroxisome proliferators and antisense RNA affects the rate of fatty acid uptake. Biochim Biophys Acta 1437: 194–201. [DOI] [PubMed] [Google Scholar]
- 28. Newberry EP, Kennedy SM, Xie Y, Luo J, Davidson NO (2009) Diet-induced alterations in intestinal and extrahepatic lipid metabolism in liver fatty acid binding protein knockout mice. Mol Cell Biochem 326: 79–86. 10.1007/s11010-008-0002-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Weickert MO, Loeffelholz CV, Roden M, Chandramouli V, Brehm A, et al. (2007) A Thr94Ala mutation in human liver fatty acid-binding protein contributes to reduced hepatic glycogenolysis and blunted elevation of plasma glucose levels in lipid-exposed subjects. Am J Physiol Endocrinol Metab 293: E1078–1084. [DOI] [PubMed] [Google Scholar]
- 30. Baier LJ, Sacchettini JC, Knowler WC, Eads J, Paolisso G, et al. (1995) An amino acid substitution in the human intestinal fatty acid binding protein is associated with increased fatty acid binding, increased fat oxidation, and insulin resistance. J Clin Invest 95: 1281–1287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. McIntosh AL, Huang H, Storey SM, Landrock KK, Landrock D, et al. (2014) Human FABP1 T94A variant impacts fatty acid metabolism and PPAR-alpha activation in cultured human female hepatocytes. Am J Physiol Gastrointest Liver Physiol 307: G164–176. 10.1152/ajpgi.00369.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Neeli I, Siddiqi SA, Siddiqi S, Mahan J, Lagakos WS, et al. (2007) Liver fatty acid-binding protein initiates budding of pre-chylomicron transport vesicles from intestinal endoplasmic reticulum. J Biol Chem 282: 17974–17984. [DOI] [PubMed] [Google Scholar]
- 33. Boullart AC, de Graaf J, Stalenhoef AF (2012) Serum triglycerides and risk of cardiovascular disease. Biochim Biophys Acta 1821: 867–875. 10.1016/j.bbalip.2011.10.002 [DOI] [PubMed] [Google Scholar]
- 34.Chen BD, Yang YN, Ma YT, Pan S, He CH, et al. (2015) Waist-to-Height Ratio and Triglycerides/High-Density Lipoprotein Cholesterol Were the Optimal Predictors of Metabolic Syndrome in Uighur Men and Women in Xinjiang, China. Metab Syndr Relat Disord. [DOI] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper.