

Abstract
Honey bee virus prevalence data are an essential prerequisite for managing epidemic events in a population. A survey study was carried out for seven viruses in colonies representing a healthy Danish honey bee population. In addition, colonies from apiaries with high level Varroa infestation or high level of winter mortality were also surveyed. Results from RT-qPCR showed a considerable difference of virus levels between healthy and sick colonies. In the group of healthy colonies, no virus was detected in 36% of cases, while at least one virus was found in each of the sick colonies. Virus titers varied among the samples, and multiple virus infections were common in both groups with a high prevalence of Sacbrood virus (SBV), Black queen cell virus (BQCV) and Deformed wing virus (DWV). Based on the distribution of virus titers, we established four categories of infection: samples free of virus (C = 0), samples with low virus titer (estimated number of virus copies 0 < C < 103), samples with medium virus titer (103 ≤ C < 107) and samples with high virus titer (C ≥ 107). This allowed us to statistically compare virus levels in healthy and sick colonies. Using categories to communicate virus diagnosis results to beekeepers may help them to reach an informed decision on management strategies to prevent further spread of viruses among colonies.
Introduction
Honey bee colonies provide valuable pollinators that enable secure crop productivity [1]. The decline of pollinators in many countries has received considerable public and scientific attention in the past decade [2]. In particular, honey bees have been experiencing considerable colony losses worldwide [3], with yearly estimates of about 30% in the U.S. since 2006 [4–6]. Losses have also been reported from Europe, although Europe appears to experience generally lower levels of losses at a less constant rate [7–9]. Multiple agents, environmental and biological, have been highlighted in correlation to colony losses [3, 7, 10–12]. However, there exists as yet no comprehensive conclusion about the cause or the most probable combination of causes [6, 13, 14].
The ectoparasitic mite, Varroa destructor, originating from one of the Asian honey bee species (Apis cerana) is an invasive species on the European honey bee (Apis mellifera) [15] and considered one major cause responsible for colony losses [4, 11, 12, 16]. The mites puncture the cuticle of honey bee workers and pupae to suck hemolymph, which may lead to immunosuppression of the parasitized host [11, 17, 18], and can also transfer virus particles [11, 19]. The detrimental impact of an infestation with high numbers of Varroa mites together with closely associated viruses (DWV, Acute bee paralysis virus (ABPV), Kashmir bee virus (KBV), and Israeli acute paralysis virus (IAPV)) on individual bees as well as colony survival has been established in previous studies [17, 19–22]. In addition to Varroa mites, the microsporidian Nosema ceranae, another possible agent involved in collapse of honey bee colonies, also created concern in some countries [12, 23, 24]. Infections with the related species Nosema apis have been linked with infections of BQCV [25].
Numerous viruses of honey bees are known and occur in different geographical regions [26, 27]. Most surveys focus on DWV, and the ABPV complex (including KBV and IAPV [28]), since they are closely associated with and transmitted by the Varroa mite. Furthermore, three viruses for which Varroa seems to play no significant role in transmission, namely Chronic bee paralysis virus (CBPV), SBV, and BQCV are frequently surveyed, too [29–31].
Based on results of infection studies, these seven viruses can be characterized as either acute or persistent. In particular, the three closely related viruses from the family Dicistroviridae (ABPV, KBV and IAPV) and the yet unclassified virus (CBPV) can cause acute infection of adult bees with a high rate of viral replication leading to high mortality of workers within a short time span [28, 32]. These viruses are found infrequently; in most prevalence studies they have been detected at low titers from bees without obvious symptoms that were sampled from healthy looking colonies. In contrast, DWV and SBV from the family Iflaviridae and BQCV from the Dicistroviridae family are known to be near omnipresent and persistent viruses, often characterized by an absence of clear disease symptoms [26]. However, they occasionally turn problematic in association with specific biotic and abiotic stress factors [13, 25, 33, 34]. Results from honey bee virus studies reveal that only a minor fraction of the honey bee populations are free of persistent viruses throughout the year [29–31]. These observations support the theory that coevolution between viruses and their hosts may lead towards less malign virus strains [35, 36]. This allows both virus and host to sustain a host-parasite relationship, for mutual persistence. It has been shown for IAPV that some colonies apparently are able to cope with the virus [37, 38]. In consequence, selection and breeding from colonies resistant to pests and pathogens can lead to a sustainable solution to combat honey bee disease [39, 40], since any chemical treatment comes with the risk of inducing resistance in the pathogens and a possible contamination of hive products.
For example, efforts to breed honey bee strains that are more tolerant towards Varroa mites are being conducted in several countries [41–43]. In Denmark, a selection program to reduce the impact of Nosemosis has been carried out over two decades which resulted in a strain of Nosema-tolerant bees [44, 45]. The ability to reliably quantify the mites and Nosema spores enables beekeepers and scientists to monitor the prevalence and quantity of parasites easily and select the best colonies for the next generations.
For virus diagnosis, quantitative RT-PCR is an advanced and more sensitive technology than qualitative gel-based technique [46]. The ability of RT-qPCR to generate accurate quantitative data has had a positive impact on honey bee viral diagnosis and on our understanding of the problems associated with viral infection [30, 47, 48]. In contrast to the qualitative techniques, quantification allows us to categorize the findings according to the level of viral load.
Colony losses in Denmark reached up to 32% in the winter of 2007 to 2008 and were largely attributed to honey bee viruses, Varroa mites and interactions between Varroa and viruses [13, 31, 49]. It was found that both high viral titers and the proportion of sick workers within colonies are directly correlated with the number of Varroa mites [13]. However, the diagnostic data of this study were obtained from symptomatic colonies and are thus likely to over-represent the prevalence of viral infections in the population. The present study aims to survey seven viruses in the healthy Danish honey bee population during the spring of 2012, to generate baseline data of virus levels in healthy and sick colonies after a long winter. The application of highly sensitive quantitative RT-PCR enables detection of virus titers much below those observed in earlier studies based on qualitative techniques and, thus, contributes to improving prevalence data [31, 49]. In addition, quantitative results may easily be categorized and thus become accessible to statistical evaluation. For beekeepers and breeders, categorized results are more easily understandable and consequently can be translated into management and selection decisions.
Material and Methods
Honey bee samples
The survey was designed to determine viral presence from apparently healthy honey bee colonies all over Denmark (Fig 1). In total, 241 samples (further referred to as healthy colonies) from 98 apiaries were received. The samples were sent by skilled Danish beekeepers, bee health inspectors, and breeders of queens, all trained in field diagnosis. Each beekeeper was asked to send a standard queen cage with up to 20 live bees per colony, exclusively from colonies that had low levels of Varroa mites and were determined free of symptomatic diseases (European Foulbrood, American Foulbrood and Chalkbrood) based on thorough visual inspection. We pooled each sample of 20 bees, which is considered sufficient for a reliable quantification of virus levels in a colony [50]. For comparison, 28 additional samples (further referred to as sick colonies) were collected from the colonies at Flakkebjerg (n = 12) (Department of Agroecology, Aarhus University) that suffered from a heavy infestation with mites and from an apiary (n = 16) with high colony mortality in the previous winter. All samples were collected alive and sent via mail during the spring of 2012 (from mid-April to mid-June). Upon arrival bees were frozen and stored at -80°C until RNA extraction.
Fig 1. Geographic distribution of honey bee samples in Denmark.

Molecular approaches
Bees of each sample were placed in a 15 mL plastic bottle together with 5–10 steel bearing balls. Using a technique adapted from plant virology, the samples were freeze dried, homogenized in a genogrinder and thereafter, RNA was extracted according to the manufacturer’s manual (for details see [13]). Following RNA extraction, a two-step real-time RT-PCR assay was used to detect and quantify seven honey bee viruses, BQCV, CBPV, SBV, DWV, ABPV, KBV, and IAPV. The three closely related viruses of the ABPV complex (ABPV, KBV and IAPV) were detected in a single assay (AKI) [51]. The housekeeping gene, β-Actin, was used as an internal control, where the presence and quantification of this reference gene ensured that the entire procedure from extraction to quantification was done without degradation of RNA [13].
Quantitative PCR amplifications were carried out on a vii7 apparatus (Applied Biosystems) in duplicate for each sample using SYBR Green DNA binding dye. Final volumes of 12μL with a primer concentration of 0.4μM were loaded on optical 384 well PCR plates. Primers [30, 47, 51–53] used in this study are listed in Table 1.
Table 1. Primers used to establish the standard curve and qRT-PCR.
| Source | Primers name | Primer sequence | Product size(bp) | Reference |
|---|---|---|---|---|
| AKI | F-AKI | 5’-CTTTCATGATGTGGAAACTCC | 100bp | [51] |
| R-AKI | 5’-AAACTGAATAATACTGTGCGTA | |||
| DWV | F-DWV | 5’-GGATGTTATCTCCTGCGTGGAA | 69bp | [30] |
| R-DWV | 5’-CTTCATTAACTGTGTCGTTGATAATTG | |||
| BQCV | BQCV-qF | 5’-AGTGGCGGAGATGTATGC | 294bp | [52] |
| BQCV-qB | 5’-GGAGGTGAAGTGGCTATATC | |||
| SBV | F-SBV | 5’- ACCAACCGATTCCTCAGTAG | 258bp | [53] |
| R-SBV | 5’- TCTTCGTCCACTCTCATCAC | |||
| CBPV | F-CBPV | 5’-CGCAAGTACGCCTTGATAAAGAAC | 101bp | [47] |
| R-CBPV | 5’-ACTACTAGAAACTCGTCGCTTCG | |||
| β.Actin | F-β-Actin | 5’-TGCCAACACTGTCCTTTCTGGAGGT | 96bp | [13] |
| R-β-Actin | 5’- TTCATGGTGGATGGTGCTAGGGCAG |
Calibration curve and data analysis
For each virus genome, a standard curve was calculated by plotting the serial dilutions of known amounts of the amplification product against the corresponding Ct values as described previously [13]. Slope and intercept of each curve were calculated with a correlation coefficient of 0.99. The amplification curve for β-Actin was used to confirm that the integrity of the RNA was preserved during the entire procedure, from the preparation of samples and RNA extraction to RT-qPCR (data not shown). Virus loads in each sample were quantified using the absolute quantification method described before [13]. Fisher’s exact test was applied to estimate the variation between healthy and sick colonies. Analysis of the data and visualization were performed using the softwares; Highcharts and R [54].
Results
Observed frequencies of the viruses in the population
A total of 241 apparently healthy and 28 sick colonies were screened for SBV, BQCV, DWV, CBPV and ABPV complex viruses. No virus at all was detected in 36% (n = 86) of the healthy colonies, while all of the sick colonies had at least one virus. The frequencies of the viruses in the two groups of colonies are shown in Fig 2.
Fig 2. Percentage of different viruses in healthy colonies (A) and sick colonies (B).
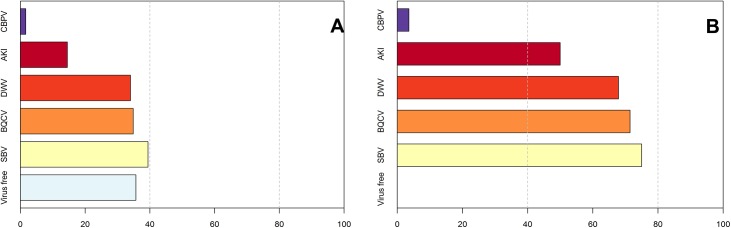
The most prevalent viruses in the healthy colonies are SBV, BQCV and DWV, with frequencies of 39% (n = 95), 35% (n = 84) and 34% (n = 82), respectively. These three viruses are also highly prevalent and frequent (75%, 71% and 68%) in the sick colonies. In contrast, viruses of the ABPV complex are present in only 14% of the healthy colonies, but 50% of the sick colonies. CBPV was only observed in four healthy colonies and one sick colony.
Twenty-seven percent of the healthy colonies contained at least one virus, mainly SBV and DWV. Multiple virus infections are also not uncommon in healthy colonies. We observed high rates of duplicate and triplicate infections (18% and 15%, respectively), the majority of them with DWV, SBV and BQCV (Fig 3). As expected, multiple virus infections are very common in sick colonies, with more than half of the samples (57.2%) simultaneously infected by three viruses. Compared to the healthy colonies, the rate of double and quadruple infections in the sick colonies was also much higher (Fig 3).
Fig 3. Percentage of single and multiple infections in healthy colonies (A) and sick colonies (B).

Viral titers in the populations
For each investigated virus the samples of healthy and sick colonies were categorized according to the quantified virus titer. Beyond the two obvious categories, bees free of virus (copy number, C = 0) and symptomatic bees (C ≥ 107), we decided to subdivide the remaining samples into two groups: samples with low virus titer (0 < C < 103) and samples with medium virus titer (103 ≤ C < 107). For both DWV and SBV we found samples falling in all four categories, whereas high BQCV and AKI titers were never observed. Finally, CBPV was observed only in the two categories no virus or medium virus level. The distribution of samples within these four categories is displayed in Fig 4 for all viruses under investigation, demonstrating significant differences between healthy and sick colonies for the virus titers of SBV, DWV, BQCV and AKI (P < 0.001), but not for CBPV (P = 0.3842).
Fig 4. Percentage of viral titer categories for each virus in two groups of healthy and sick colonies.

Discussion
We report data on the prevalence of viruses in samples of healthy Danish honey bee colonies collected in the spring time. Similar to earlier studies from other countries [29, 30], no virus was found in 36% (n = 86) of the healthy colonies. In spite of us requesting bees to be collected from healthy colonies, a few samples contained at least one bee with a high virus titer. Instead of interpreting this as a beekeeper’s mistake, rather this result is expected since the virus is probably present at high levels in just a few bees in most colonies, however in samples of 20 bees it mostly goes undetected.
Sacbrood virus is found to be the most prevalent virus in Danish honey bees for the period of investigation. Almost 40% of the healthy colonies carry SBV, with 18% of the population infected by a medium or high titer of the virus. This virus was even more frequently detected in our samples of sick colonies (75%) which is in accordance with a previous study from Denmark (81% [31]). Sacbrood is known as a disease affecting the brood of the honey bee, but it has also been reported from adult honey bees lacking any obvious sign of disease [14, 30, 34]. The occurrence of Sacbrood in the spring and summer may be related to colony growth with a high ratio of brood to nurse bees [55], or the quality of available pollen [29]. Even though an association between SBV and Varroa mites was reported in different studies [29, 56], the effect is most likely resulting from stress at colony level, since the disease occurs in larvae before these are attractive to Varroa mites. In Denmark it has frequently been reported that colonies suffer from severe outbreaks of SBV, not commonly found elsewhere in Europe [57].
Deformed wing virus is the other highly prevalent virus in this study. It was detected in 34% of the healthy colonies with almost 11% of these carrying a medium or high titer which is associated with reduced life span of the adult bees [58]. This result is concurrent with previous reports from Denmark [13, 31, 49]. DWV has also been found to be highly prevalent in honey bees in other countries [12, 22, 30]. DWV can be detected in all developmental stages and castes of bees [29, 59]. It is closely associated with the Varroa mite and strongly suspected to be one of the biological agents for honeybee colony losses [12, 59], since a significant relationship between DWV and colony collapse was reported in several studies [10, 11, 13, 34, 50, 60]. Our data for the sick colonies (68% prevalence with 40% medium and high titer) may be associated with high levels of Varroa mites infestation during the preceding winter.
Black queen cell virus is another highly prevalent virus in the healthy colonies in this study. It has been reported as a common virus in adult European honey bees [30], however, in Denmark only one single case was reported [31]. We hypothesize that the absence of BQCV positive sample in the previous study may be the result of lower sensitivity of the technique they used. We observed only low and medium titer infections, in both the healthy and the sick colonies.
BQCV has been detected with high titers in collapsed colonies [10]. A recent study suggests that BQCV has limited effects on both drone and worker health [61]. There are reports regarding a close association between BQCV and N. apis [25, 29]. As yet, no synergistic interaction has been found between BQCV and N. ceranae [61].
Acute bee paralysis virus, Kashmir bee virus and Israeli bee paralysis virus are three closely related viruses that were analyzed together in a single assay [51]. Almost 14% of the healthy colonies were found positive, but only 2% of colonies carried a medium titer of virus and not a single case with high titer was detected. All three viruses are equally and highly virulent and have a close association with the Varroa mite [13, 20, 21, 28]. While our results do not distinguish between the three subtypes (ABPV, KBV and IAPV), all three are known to rapidly kill the bees. Several studies in recent years have identified the family of Acute paralysis viruses as one of the major biological agents detected in collapsed colonies [3, 13, 62]. Our results are in agreement with previous studies that report a low prevalence of these viruses in spring [13, 34], and observe high viral titers only in collapsing colonies [10, 30]. Therefore, high titers of these viruses are rarely detected in active survey studies since highly infected bees die fast and are therefore not sampled.
Chronic bee paralysis virus is the rarest detected virus, also amongst the sick colonies. CBPV is known as a disease of adult honey bees, mostly with outbreaks during high nectar flow [32]. We only detected CBPV in 1.7% of the healthy colonies with a medium titer. This result is in agreement with previous studies that also reported a low prevalence of CBPV [29, 31].
Several of the viruses we discuss above are considered serious pathogens in regard to their impact on colony survival [10, 12, 13]. However, viruses often persist in colonies as covert infections [26] with limited consequences for colony health, and of little concern to beekeepers. Nonetheless, the standard procedure in virus analysis thus far has been to report the result as “negative” or “positive” [12, 59, 63]. In fact, a positive result from a whole-body extraction of 20 workers, as in our study, could result from a covert infection. Thus, a positive qualitative result does not necessarily allow for a meaningful prognosis.
For instance, even after successful Varroa treatment, DWV seems to prevail in the colony and the infection level will build up again, when the Varroa population increases. Individual bees parasitized by Varroa mites, either in the pupal stage or as adults, exhibit extraordinary high viral titers [64]. As the Varroa population in a colony increases, more and more bees carry such high infection levels and therefore the chance of including one of those bees in a random sample will also increase. Similar patterns are expected for the Acute bee paralysis virus complex, which may result in colony loss [65].
On the other hand, a virus like CBPV, which is not vectored by Varroa, may spread readily between colonies and is considered very contagious [32, 66]. We therefore consider it helpful if colonies carrying high virus titers within an apiary can be identified and adequately taken care of, for instance, by removing them to a quarantine apiary. In our experience beekeepers and breeders listen more readily and conceive the meaning of virus analysis results more easily when these are communicated as categories.
Frequently, due to the non-normal distribution of virus titers over several magnitudes, problems arise when trying to statistically compare two sets of samples. Categorization of quantitative virus titers renders the results accessible to statistical evaluation beyond non-parametric tests and thus allows comparisons between different samplings. Such data enable monitoring the progression of a viral infection in a colony or apiary, or comparisons between different breeding lines in a selection program. Based on a sufficient number of observations, critical thresholds will emerge that can be used for management decisions [57]. Especially in ongoing selection programmes for Varroa tolerance [42, 67] information on the virus load of potential queen and drone mother colonies may be provide useful information in the evaluation of breeding stock.
Acknowledgments
The authors would like to thank numerous Danish beekeepers, bee health inspectors and breeders of queens for providing us with samples on time.
Data Availability
All relevant data are within the paper.
Funding Statement
This research was founded by a grant, Disease and pesticide resistance in plants and animals, from the Danish Ministry of Science, Innovation and Higher Education.
References
- 1. Delaplane KS, Mayer DR, Mayer DF. Crop pollination by bees: Cabi; 2000. [Google Scholar]
- 2. Potts SG, Biesmeijer JC, Kremen C, Neumann P, Schweiger O, Kunin WE. Global pollinator declines: trends, impacts and drivers. Trends Ecol Evol 2010;25(6):345–53. 10.1016/j.tree.2010.01.007 [DOI] [PubMed] [Google Scholar]
- 3. vanEngelsdorp D, Meixner MD. A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J Invertebr Pathol. 2010;103 S80–95. 10.1016/j.jip.2009.06.011 [DOI] [PubMed] [Google Scholar]
- 4. vanEngelsdorp D, Hayes J Jr, Underwood RM, Caron D, Pettis J. A survey of managed honey bee colony losses in the USA, fall 2009 to winter 2010. J Apicult Res. 2011;50(1):1–10. 10.3896/ibra.1.50.1.01 [DOI] [Google Scholar]
- 5. Spleen AM, Lengerich EJ, Rennich K, Caron D, Rose R, Pettis JS, et al. A national survey of managed honey bee 2011–12 winter colony losses in the United States: results from the Bee Informed Partnership. J Apicult Res. 2013;52(2):44–53. 10.3896/ibra.1.52.2.07 [DOI] [Google Scholar]
- 6. vanEngelsdorp D, Caron D, Hayes J, Underwood R, Henson M, Rennich K, et al. A national survey of managed honey bee 2010–11 winter colony losses in the USA: results from the Bee Informed Partnership. J Apicult Res. 2012;51(1):115–24. 10.3896/ibra.1.51.1.14 [DOI] [Google Scholar]
- 7. Neumann P, Carreck NL. Honey bee colony losses. J Apicult Res. 2010;49(1):1–6. 10.3896/IBRA.1.49.1.01 [DOI] [Google Scholar]
- 8. Potts S, Roberts S, Dean R, Marris G, Brown M, Jones R, et al. Declines of managed honey bees and beekeepers in Europe. J Apicult Res. 2010;49(1):15 10.3896/ibra.1.49.1.02 [DOI] [Google Scholar]
- 9. van der Zee R, Pisa L, Andonov S, Brodschneider R, Charrière J-D, Chlebo R, et al. Managed honey bee colony losses in Canada, China, Europe, Israel and Turkey, for the winters of 2008–9 and 2009–10. J Apicult Res. 2012;51(1):100–14. 10.3896/ibra.1.51.1.12 [DOI] [Google Scholar]
- 10. Cornman RS, Tarpy DR, Chen Y, Jeffreys L, Lopez D, Pettis JS, et al. Pathogen webs in collapsing honey bee colonies. PLoS One. 2012;7(8):e43562 10.1371/journal.pone.0043562 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Nazzi F, Brown SP, Annoscia D, Del Piccolo F, Di Prisco G, Varricchio P, et al. Synergistic parasite-pathogen interactions mediated by host immunity can drive the collapse of honeybee colonies. PLoS Pathog. 2012;8(6):e1002735 10.1371/journal.ppat.1002735 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Genersch E, von der Ohe W, Kaatz H, Schroeder A, Otten C, Büchler R, et al. The German bee monitoring project: a long term study to understand periodically high winter losses of honey bee colonies. Apidologie. 2010;41(3):332–52. [Google Scholar]
- 13. Francis RM, Nielsen SL, Kryger P. Varroa-virus interaction in collapsing honey bee colonies. PLoS One. 2013;8(3):e57540 10.1371/journal.pone.0057540 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Runckel C, Flenniken ML, Engel JC, Ruby JG, Ganem D, Andino R, et al. Temporal analysis of the honey bee microbiome reveals four novel viruses and seasonal prevalence of known viruses, Nosema, and Crithidia . PLoS One. 2011;6(6):e20656 10.1371/journal.pone.0020656 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Rosenkranz P, Aumeier P, Ziegelmann B. Biology and control of Varroa destructor . J Invertebr Pathol. 2010;103 S96–119. 10.1016/j.jip.2009.07.016 [DOI] [PubMed] [Google Scholar]
- 16. Meixner MD, Francis RM, Gajda A, Kryger P, Andonov S, Uzunov A, et al. Occurrence of parasites and pathogens in honey bee colonies used in a European genotype–environment interactions experiment. J Apicult Res. 2014;53(2):215–9. 10.3896/IBRA.1.53.2.04 [DOI] [Google Scholar]
- 17. Di Prisco G, Pennacchio F, Caprio E, Boncristiani HF Jr., Evans JD, Chen Y. Varroa destructor is an effective vector of Israeli acute paralysis virus in the honeybee, Apis mellifera . J Gen Virol. 2011;92(1):151–5. 10.1099/vir.0.023853-0 [DOI] [PubMed] [Google Scholar]
- 18. Yang X, Cox-Foster DL. Impact of an ectoparasite on the immunity and pathology of an invertebrate: evidence for host immunosuppression and viral amplification. P Natl Acad Sci USA. 2005;102(21):7470–5. 10.1073/pnas.0501860102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Bowen-Walker PL, Martin SJ, Gunn A. The transmission of Deformed wing virus between honeybees (Apis mellifera L.) by the ectoparasitic mite Varroa jacobsoni Oud. J Invertebr Pathol. 1999;73(1):101–6. 10.1006/jipa.1998.4807 [DOI] [PubMed] [Google Scholar]
- 20. Chen Y, Pettis JS, Evans JD, Kramer M, Feldlaufer MF. Transmission of Kashmir bee virus by the ectoparasitic mite Varroa destructor . Apidologie. 2004;35:441–8. 10.1051/apido:2004031 [DOI] [Google Scholar]
- 21. Shen M, Yang X, Cox-Foster D, Cui L. The role of varroa mites in infections of Kashmir bee virus (KBV) and Deformed wing virus (DWV) in honey bees. Virology. 2005;342(1):141–9. 10.1016/j.virol.2005.07.012 [DOI] [PubMed] [Google Scholar]
- 22. Dainat B, Neumann P. Clinical signs of Deformed wing virus infection are predictive markers for honey bee colony losses. J Invertebr Pathol. 2013;112(3):278–80. 10.1016/j.jip.2012.12.009 [DOI] [PubMed] [Google Scholar]
- 23. Higes M, Martin-Hernandez R, Botias C, Bailon EG, Gonzalez-Porto AV, Barrios L, et al. How natural infection by Nosema ceranae causes honeybee colony collapse. Environmental microbiology. 2008;10(10):2659–69. 10.1111/j.1462-2920.2008.01687.x [DOI] [PubMed] [Google Scholar]
- 24. Higes M, Meana A, Bartolomé C, Botías C, Martín-Hernández R. Nosema ceranae (Microsporidia), a controversial 21st century honey bee pathogen. Environ Microbiol Rep. 2013;5(1):17–29. 10.1111/1758-2229.12024 [DOI] [PubMed] [Google Scholar]
- 25. Bailey L, Ball BV, Perry JN. Association of viruses with two protozoal pathogens of the honey bee. Ann App Biol. 1983;103(1):13–20. 10.1111/j.1744-7348.1983.tb02735.x [DOI] [Google Scholar]
- 26. Chen YP, Siede R. Honey bee viruses. Advances in virus research. 702007. p. 33–80. [DOI] [PubMed] [Google Scholar]
- 27. Maori E, Lavi S, Mozes-Koch R, Gantman Y, Peretz Y, Edelbaum O, et al. Isolation and characterization of Israeli acute paralysis virus, a dicistrovirus affecting honeybees in Israel: evidence for diversity due to intra- and inter-species recombination. J Gen Virol. 2007;88(12):3428–38. 10.1099/vir.0.83284-0 [DOI] [PubMed] [Google Scholar]
- 28. de Miranda JR, Cordoni G, Budge G. The Acute bee paralysis virus–Kashmir bee virus–Israeli acute paralysis virus complex. J Invertebr Pathol. 2010;103:S30–S47. 10.16/j.jip.2009.06.014 [DOI] [PubMed] [Google Scholar]
- 29. Tentcheva D, Gauthier L, Zappulla N, Dainat B, Cousserans F, Colin ME, et al. Prevalence and seasonal variations of six bee viruses in Apis mellifera L. and Varroa destructor mite populations in France. Appl Environ Microbiol. 2004;70(12):7185–91. 10.1128/AEM.70.12.7185-7191.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Gauthier L, Tentcheva D, Tournaire M, Dainat B, Cousserans F, Colin M, Edouard, et al. Viral load estimation in asymptomatic honey bee colonies using the quantitative RT-PCR technique. Apidologie. 2007;38(5):426–35. [Google Scholar]
- 31. Nielsen S, Lykke, Nicolaisen M, Kryger P. Incidence of acute bee paralysis virus, black queen cell virus, chronic bee paralysis virus, deformed wing virus, Kashmir bee virus and sacbrood virus in honey bees (Apis mellifera) in Denmark. Apidologie. 2008;39(3):310–4. [Google Scholar]
- 32. Ribiere M, Olivier V, Blanchard P. Chronic bee paralysis: a disease and a virus like no other? J Invertebr Pathol. 2010;103 S120–31. 10.1016/j.jip.2009.06.013 [DOI] [PubMed] [Google Scholar]
- 33. Di Prisco G, Zhang X, Pennacchio F, Caprio E, Li J, Evans JD, et al. Dynamics of persistent and acute Deformed wing virus infections in honey bees, Apis mellifera . Viruses. 2011;3(12):2425–41. 10.3390/v3122425 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Dainat B, Evans JD, Chen YP, Gauthier L, Neumann P. Predictive markers of honey bee colony collapse. PLoS One. 2012;7(2):e32151 10.1371/journal.pone.0032151 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Goic B, Saleh M-C. Living with the enemy: viral persistent infections from a friendly viewpoint. Curr Opin Microbiol. 2012;15(4):531–7. 10.1016/j.mib.2012.06.002 [DOI] [PubMed] [Google Scholar]
- 36. Fries I, Camazine S. Implications of horizontal and vertical pathogen transmission for honey bee epidemiology. Apidologie. 2001;32(3):199–214. [Google Scholar]
- 37. Maori E, Tanne E, Sela I. Reciprocal sequence exchange between non-retro viruses and hosts leading to the appearance of new host phenotypes. Virology. 2007;362(2):342–9. 10.1016/j.virol.2006.11.038 [DOI] [PubMed] [Google Scholar]
- 38. Chejanovsky N, Ophir R, Schwager MS, Slabezki Y, Grossman S, Cox-Foster D. Characterization of viral siRNA populations in honey bee colony collapse disorder. Virology. 2014;454–455(0):176–83. 10.1016/j.virol.2014.02.012 [DOI] [PubMed] [Google Scholar]
- 39. Kulincevic JM, Rothenbuhler WC. The effects of artificial infection with Chronic bee paralysis virus on queens from strains of honeybee resistant or susceptible to Hairless-black syndrome. J Apicult Res. 1989;28(2):79–80. [Google Scholar]
- 40. Kulincevic JM, Rothenbuhler WC. Selection for resistance and susceptibility to Hairless-black syndrome in honeybee. J Invertebr Pathol. 1975;25(3):289–95. 10.1016/0022-2011(75)90084-1 [DOI] [PubMed] [Google Scholar]
- 41. Fries I, Imdorf A, Rosenkranz P. Survival of mite infested (Varroa destructor) honey bee (Apis mellifera) colonies in a Nordic climate. Apidologie. 2006;37(5):564–70. [Google Scholar]
- 42. Büchler R, Berg S, Le Conte Y. Breeding for resistance to Varroa destructor in Europe. Apidologie. 2010;41(3):393–408. 10.1051/apido/2010011 [DOI] [Google Scholar]
- 43. Le Conte Y, Alaux C, Martin JF, Harbo JR, Harris JW, Dantec C, et al. Social immunity in honeybees (Apis mellifera): transcriptome analysis of varroa-hygienic behaviour. Insect Mol Biol. 2011;20(3):399–408. 10.1111/j.1365-2583.2011.01074.x [DOI] [PubMed] [Google Scholar]
- 44. Huang Q, Kryger P, Le Conte Y, Moritz RF. Survival and immune response of drones of a Nosemosis tolerant honey bee strain towards N. ceranae infections. J Invertebr Pathol. 2012;109(3):297–302. 10.1016/j.jip.2012.01.004 [DOI] [PubMed] [Google Scholar]
- 45. Huang Q, Lattorff HMG, Kryger P, Le Conte Y, Moritz RFA. A selective sweep in a microsporidian parasite Nosema-tolerant honeybee population, Apis mellifera . Anim Genet. 2013;45(2):267–73. 10.1111/age.12114 [DOI] [PubMed] [Google Scholar]
- 46. Bustin SA. Why the need for qPCR publication guidelines?—The case for MIQE. Methods. 2010;50(4):217–26. 10.1016/j.ymeth.2009.12.006 [DOI] [PubMed] [Google Scholar]
- 47. Blanchard P, Ribiere M, Celle O, Lallemand P, Schurr F, Olivier V, et al. Evaluation of a real-time two-step RT-PCR assay for quantitation of Chronic bee paralysis virus (CBPV) genome in experimentally-infected bee tissues and in life stages of a symptomatic colony. J Virol Methods. 2007;141(1):7–13. 10.1016/j.jviromet.2006.11.021 [DOI] [PubMed] [Google Scholar]
- 48. Chen YP, Higgins JA, Feldlaufer MF. Quantitative real-time reverse transcription-PCR analysis of Deformed wing virus infection in the honeybee (Apis mellifera L.). Appl Environ Microbiol. 2005;71(1):436–41. 10.1128/aem.71.1.436-441.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Vejsnæs F, Nielsen SL, Kryger P. Factors involved in the recent increase in colony losses in Denmark. J Apicult Res. 2010;49(1):109–10. [Google Scholar]
- 50. Highfield AC, El Nagar A, Mackinder LCM, Noël LM-LJ, Hall MJ, Martin SJ, et al. Deformed Wing Virus Implicated in Overwintering Honeybee Colony Losses. Appl Environ Microbiol. 2009;75(22):7212–20. 10.1128/aem.02227-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Francis R, Kryger P. Single assay detection of Acute bee paralysis virus, Kashmir bee virus and Israeli acute paralysis virus. J Apicult Sci. 2012;56(1):137 10.2478/v10289-012-0014-x [DOI] [Google Scholar]
- 52. Lindström M. Detection of honey bee viruses in Apis mellifera and Apis cerana Sweden: Uppsala University; 2011. [Google Scholar]
- 53. Grabensteiner E, Ritter W, Carter MJ, Davison S, Pechhacker H, Kolodziejek J, et al. Sacbrood virus of the honeybee (Apis mellifera): rapid identification and phylogenetic analysis using reverse transcription-PCR. Clin Diagn Lab Immunol. 2001;8(1):93–104. 10.1128/CDLI.8.1.93-104.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. R Development Core Team. R: A language and environment for statistical computing Vienna, Austria: R Foundation for Statistical Computing; 2010. [Google Scholar]
- 55. Bailey L. The multiplication and spread of sacbrood virus of bees. Ann App Biol. 1969;63(3):483–91. 10.1111/j.1744-7348.1969.tb02844.x [DOI] [PubMed] [Google Scholar]
- 56. Shen MQ, Cui LW, Ostiguy N, Cox-Foster D. Intricate transmission routes and interactions between picorna-like viruses (Kashmir bee virus and Sacbrood virus) with the honeybee host and the parasitic varroa mite. J Gen Virol. 2005;86:2281–9. 10.1099/vir.0.80824-0 [DOI] [PubMed] [Google Scholar]
- 57. Blanchard P, Guillot S, Antùnez K, Köglberger H, Kryger P, de Miranda JR, et al. Development and validation of a real-time two-step RT-qPCR TaqMan® assay for quantitation of Sacbrood virus (SBV) and its application to a field survey of symptomatic honey bee colonies. J Virol Methods. 2014;197:7–13. 10.1016/j.jviromet.2013.09.012 [DOI] [PubMed] [Google Scholar]
- 58. Dainat B, Evans JD, Chen YP, Gauthier L, Neumann P. Dead or Alive: Deformed Wing Virus and Varroa destructor Reduce the Life Span of Winter Honeybees. Appl Environ Microbiol. 2012;78(4):981–7. 10.1128/aem.06537-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. de Miranda JR, Genersch E. Deformed wing virus. J Invertebr Pathol. 2010;103:S48–S61. 10.1016/j.jip.2009.06.012 [DOI] [PubMed] [Google Scholar]
- 60. Baker A, Schroeder D. Occurrence and genetic analysis of picorna-like viruses infecting worker bees of Apis mellifera L. populations in Devon, South West England. J Invertebr Pathol. 2008;98(2):239–42. 10.1016/j.jip.2008.02.010 [DOI] [PubMed] [Google Scholar]
- 61. Retschnig G, Williams GR, Mehmann MM, Yañez O, de Miranda JR, Neumann P. Sex-specific differences in pathogen susceptibility in honey bees (Apis mellifera). PLoS One. 2014;9(1):e85261 10.1371/journal.pone.0085261 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Cox-Foster DL, Conlan S, Holmes EC, Palacios G, Evans JD, Moran NA, et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science. 2007;318(5848):283–7. 10.1126/science.1146498 [DOI] [PubMed] [Google Scholar]
- 63. Chen Y, Pettis JS, Feldlaufer MF. Detection of multiple viruses in queens of the honey bee Apis mellifera L. J Invertebr Pathol. 2005;90(2):118–21. 10.1016/j.jip.2005.08.005 [DOI] [PubMed] [Google Scholar]
- 64. Yue C, Genersch E. RT-PCR analysis of Deformed wing virus in honeybees (Apis mellifera) and mites (Varroa destructor). J Gen Virol. 2005;86(12):3419–24. 10.1099/vir.0.81401-0 [DOI] [PubMed] [Google Scholar]
- 65. Martin SJ. The role of Varroa and viral pathogens in the collapse of honeybee colonies: a modelling approach. J Appl Ecol. 2001;38(5):1082–93. 10.1046/j.1365-2664.2001.00662.x [DOI] [Google Scholar]
- 66. Bailey L, Gibbs AJ, Woods RD. Two viruses from adult honey bees (Apis mellifera Linnaeus). Virology. 1963;21:390–5. [DOI] [PubMed] [Google Scholar]
- 67. Locke B, Forsgren E, de Miranda J, Rueppell O. Increased Tolerance and Resistance to Virus Infections: A Possible Factor in the Survival of Varroa destructor-Resistant Honey Bee (Apis mellifera). PLoS One. 2014;9(6):e99998 10.1371/journal.pone.0099998 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper.