

Abstract
Chronic social subordination is a well-known precipitant of numerous psychiatric and physiological health concerns. In this study, we examine the effects of chronic social stress in the visible burrow system (VBS) on the expression of glutamic acid decarboxylase (GAD) 67 and brain-derived neurotropic factor (BDNF) mRNA in forebrain stress circuitry. Male rats in the VBS system form a dominance hierarchy, whereby subordinate males exhibit neuroendocrine and physiological profiles characteristic of chronic exposure to stress. We found that social subordination decreases GAD67 mRNA in the peri-paraventricular nucleus region of the hypothalamus and the interfascicular nucleus of the bed nucleus of the stria terminalis (BNST), and increases in GAD67 mRNA in the hippocampus, medial prefrontal cortex, and dorsal medial hypothalamus. Expression of BDNF mRNA increased in the dorsal region of the BNST, but remained unchanged in all other regions examined. Results from this study indicate that social subordination is associated with several region-specific alterations in GAD67 mRNA expression in central stress circuits, whereas changes in the expression of BDNF mRNA are limited to the BNST.
Keywords: BDNF, Visible Burrow System, BNST, mPFC, hippocampus
1. Introduction
Chronic exposure to stress is linked to numerous psychiatric and physiological health concerns. Epidemiological studies consistently associate exposure to stressors to the development of depression and metabolic disorders (Kessler et al., 2005, Sapolsky, 2005, Bergmann et al., 2014). Subjection to social subordination status is a potent form of chronic stress. In rats, the effect of chronic stress imposed by social subordination can be studied in the well-characterized visible burrow system (VBS), characterized originally by the Blanchards (Blanchard et al., 1985, Blanchard and Blanchard, 1989, Blanchard et al., 1995, Albeck et al., 1997). In this model, four male and two female rats are housed together, an arrangement that causes formation of a dominance hierarchy among the males. Within one to two days, one male emerges as a dominant. Subordinate members are typically characterized by weight loss, bite wounds to the back and tail, and spend the most amount of time in the non-open areas of the VBS (Blanchard et al., 1995, Tamashiro et al., 2007a). This dominance hierarchy, once formed, is very stable and presents a potent form of chronic stress to subordinate members.
The physiological profile of dominant and subordinate members in the VBS model also varies. Subordinate members in the VBS are characterized by hypertrophy of the adrenal glands, lower testosterone levels, elevated basal and stress-induced corticosterone levels, atrophy of the thymus, and marked and persistent loss of both fat mass and lean body mass (Blanchard et al., 1993, Blanchard et al., 1995, Tamashiro et al., 2004, Tamashiro et al., 2005, Choi et al., 2006, Tamashiro et al., 2007a). Dominant rats, however, have a very limited weight loss in comparison to non-VBS controls, which is largely due to loss of body fat (Blanchard et al., 1993, Hardy et al., 2002, Choi et al., 2006).
Stress integration involves a variety of limbic structures, including the ventromedial prefrontal cortex (prelimbic and infralimbic cortices), hippocampus and the medial and central amygaloid nuclei (Ulrich-Lai and Herman, 2009). These limbic structures project to a number of hypothalamic structures that appear to relay output to stress effector systems such as the paraventricular nucleus of the hypothalamus (PVN). Prior studies from our group and others have underscored the importance of one of these relay nuclei, the bed nucleus of the stria terminalis (BNST), in control of acute and chronic stress responses (Choi et al., 2006, Choi et al., 2008a, Choi et al., 2008b, Radley et al., 2009). Moreover, chronic stress increases dendritic branching in the BNST and enhances brain derived neurotrophic factor (BDNF) expression in the dorsal BNST, suggesting that stress induces neuroplastic responses in this region that may impact output. Importantly, VBS exposure increases corticotropin releasing hormone (CRH) expression in the dorsal BNST of subordinates, while decreasing expression of glutamic acid decarboxylase (GAD) 67 in the intrafascicular BNST, suggesting gain of function in putative stress-excitatory nucleus (Choi et al., 2006).
The current study was designed to determine the consequences of social subordination on BDNF as well as GAD67 expression in the BNST as well as other brain regions responsible for brain stress integration. Our data indicate a marked increase in BDNF mRNA expression in the dorsal BNST of subordinates, accompanied by an increase in GAD67 mRNA levels in the prefrontal cortices and hippocampus. In combination, the data support the importance of the BNST in control of chronic stress reactivity in multiple experimental contexts, and suggests that drive of regions such as the BNST may be related to increased inhibition in upstream limbic structures.
2. Materials and methods
2.1 Subjects
Adult male Long-Evans rats (n=32) and intact adult female Long-Evans rats (n=12) (Harlan, Indianapolis, IN, USA; 100–120 days of age upon arrival) were obtained and allowed to habituate in individual housing in shoebox cages for three weeks before exposure to VBS. Water and food were provided ad libitum in a temperature/humidity-controlled room on a 12/12-h light/dark cycle. Rats were fed standard rat chow (Harlan Teklad, Indianapolis, IN, USA) and provided tap water for the duration of the experiment. All procedures were carried out in accordance with the guidelines of the National Institutes of Health (NIH) and were approved by the Institutional Animal Care and Use Committee of the University of Cincinnati.
2.2 The visible burrow system (VBS)
Detailed descriptions of the VBS have been published previously (Blanchard et al., 1995, Tamashiro et al., 2004, Tamashiro et al., 2007a). Briefly, the VBS was constructed from black Plexiglas material with one large open area and two smaller chambers. Clear Plexiglas-covered tunnels connected the three chambers, and the tops of the two smaller chambers were made of clear Plexiglas to allow for visual monitoring. The larger, open field area was illuminated by a mounted 15 W light bulb on a 12/12-h light cycle with lights on at 6:00am. The two other small chambers were kept in constant darkness. Food and water were provided ad libitum in each of the three chambers. For behavioral reference, each VBS colony was videotaped starting at 6:00pm (at lights out) for 6 h on days 0, 1, 2, 4, 6, 8, 10 and 12.
Six VBS colonies were created for this study, with each colony consisting of four males and two females. One day prior to entry into the VBS, male rats were weight-matched. Pictures of the dorsal hair patterns were taken for identification purposes. Control male rats were pair-housed with a female in a standard plastic shoebox cage in a separate room. Each control male rat was weight-matched with each colony and remained in its housing for the duration of the study (14 days). Experimental animals remained in the VBS apparatus for a total of 14 days.
2.3 Body weight and wound counts
On days 0, 1, 3, 5, 7, 9, 11 and 12, male rats were removed from their VBS colony and placed in their original home cage with ad libitum access to water and food. The rats were then weighed and wound counts recorded. Wounds to the face and head were accounted for separately from wounds to the flank, tail and underside regions. Similarly, controls were removed from their female partner, weighed and handled for the same duration that experimental VBS animals spent in their home cages. The time the experimental animals spent out of the VBS apparatus was limited to no more than 1.5 h.
2.4 Assessment of social status
Dominance status was determined by three criteria: (1) time in open area of the VBS, (2) body weight and composition and (3) wound counts. Typically, a social hierarchy is established within one day of entering into the VBS. Each day at 12:00pm, the VBS colonies were checked, and animals spending the most amount of time in the open area of the VBS were recorded. The second criterion was assessed as a percentage of weight gain or loss from original starting weight and by the percent of lean/fat mass gain or loss. Whole body composition was assessed on day 0 and 13 of the experiment. Animals were placed into a clear Plexiglas tube, which was then inserted into an EchoMRI whole body composition analyzer system (Echo Medical Systems, Houston, TX). This system provides a count of fat mass, lean mass, and water content (Smeltzer et al., 2012). The third criterion was assessed by counting the total number of wounds to each part of the body. Typically, dominant animals are characterized as having the least amount of weight loss, fewest number of bite wounds, and the most amount of time spent in the open area of the VBS. Subordinates, on the other hand, have substantially more bite wounds to the back and tail regions, lose significantly more weight, and spend virtually no time in the open area region of the VBS. In the present study, each of the six VBS colonies established one clear dominant and three subordinate members.
2.5 Acute restraint stress test and sacrifice
On day 14 of the study, experimental rats were removed from their VBS apparatus and immediately placed into clear, ventilated Plexiglas restraint tubes (length 17 cm and inner diameter of 7 cm) within their original home cages. For the pair-housed controls, females were removed from the control males, whereupon the males were placed in restraint tubes within their home cage. A small blood sample (50 μl) from each rat was collected into 1.5-ml microcentrifuge tubes containing 4 μl of 100 mM EDTA by creating a nick at the tip of the tail (Vahl et al., 2005). A second blood sample was collected at 60 min by removing the blood clot at the end of the tail. At this time, the animals were removed from the restrainers and allowed to move freely within their home cage. At 120 min, a third blood sample was collected. Females were then returned to their pair-housed controls, and all other rats returned to their respective VBS apparatus. All samples were immediately placed on ice after collection and centrifuged at 0°C. The plasma was then removed and stored at −20°C until assayed by radioimmunoassay (RIA). Initial baseline blood collection occurred at 9:00am. Approximately 24 h after the 120 min time-point bleed (12:00pm), all animals were euthanized by rapid decapitation. Brains were rapidly removed and flash-frozen on dry ice before being stored at −80°C. Plasma corticosterone (CORT) levels were determined using an RIA kit (CORT DA; MP Biomedicals, Solon, OH). The RIA kit had an inter-assay coefficient of variation (CV) of ~7.2% and an intra-assay CV of ~10.3%. Samples were run in duplicates within the same assay.
2.6 In situ hybridization
Frozen brains were sectioned at 12-μm thickness in the coronal plane using a Leica 3050 S cryostat and thaw-mounted onto Fisher Superfrost Plus slides (Fisher, Hampton, NJ). Slides were stored at −20°C until processing for the detection of BDNF and GAD67 mRNAs by using in situ hybridization, as detailed previously (Seroogy and Herman, 1997, Hemmerle et al., 2012). Briefly, the sections were fixed in a 4% paraformaldehyde/0.1 M phosphate-buffered saline (PBS) solution for 10 min, rinsed twice in 0.1 M PBS, once in 0.1 M PBS with 0.2% glycine for 5 min and then twice in 0.1 M PBS for 5 min each. Sections were then acetylated in 0.1M triethanolamine (pH 8.0) with 0.25% acetic anhydride for 10 min. Next, sections were then washed in 0.2X saline-sodium citrate (SSC) (2 × 5 min) before being dehydrated through a graded series of ethanol washes and delipidated with chloroform.
Antisense pan-BDNF and GAD67 cRNA probes were labeled with 35S-UTP (PerkinElmer, Boston, MA) and diluted in hybridization buffer to yield a count of 1.0×106 cpm/50 μl buffer. The remaining hybridization cocktail consisted of 50% deionized formamide; 335 mM NaCl; 10% dextran sulfate; 0.3 mg/ml ssDNA; 1 mM EDTA; 1X Denhardt’s solution; 0.15 mg/ml tRNA; 40 mM DTT; 20 mM Tris-HCl. Fifty μl of hybridization solution were pipetted onto each slide followed by coverslipping. All slides were incubated at 60°C overnight in hybridization chambers containing moist blotting paper soaked in 50% formamide. Following 18 h incubation, coverslips were removed, and the slides were washed in 4XSSC for 30 min, rinsed in RNase A (50 μg/ml for 30 min at 37°C), and then washed in 2X SSC (2 × 20 min), 0.5XSSC (2 × 20 min), and 0.1XSSC (30 min). Slides were then quickly dipped in dH20, 95% ethanol, and air-dried.
2.7 Image analysis
Hybridized slides were exposed to BioMax MR film (Kodak, Rochester, NY) for 16 days for BDNF and 7 days for GAD67. Images of the brain sections were obtained via a digital video camera. Semi-quantitative image analyses were performed by densitometry using Scion Image software (Scion, Frederick, MD). Regions of interests were identified using the rat stereotaxic brain atlas of Paxinos and Watson (Paxinos and Watson, 1998). Gray level intensities were determined in anatomically defined regions showing hybridization signal using BDNF or GAD67 cRNA probes. Background gray level signal was taken from a comparably sized region showing no hybridization signal, and was subtracted from each gray level measure to yield a corrected gray level.
2.8 Data analysis
Data are presented as mean ± standard error of the mean (S.E.M). Body weights are expressed as percent change from original body weight. Because adrenal weight correlates to the overall weight of the animal, adrenal weights were corrected for by taking the weight of the organ divided by the final body weight. Organ weight, body mass, and corrected GAD67 and BDNF mRNA hybridization signal were analyzed using a one-way factorial ANOVA design. Differences in means of CORT measurements were analyzed via two-way repeated measures ANOVA. Post hoc comparisons of significant interactions were made using Fisher’s least significant difference (LSD) analysis.
3. Results
3.1 Body composition and adrenal weights
The physiological profiles of subordinate rats (SUB) following 14d in the VBS are consistent with chronic stress exposure. Control rats (CON) and dominant rats (DOM) gained or maintained their weight while SUB lost weight as a percentage of original body weight (Fig. 1A). There were main effects of social status (F2,162=25.006, P<0.05), time (F2,126=9.442, P<0.05) and status x time interaction (F2,162=10.230, P<0.05). SUB rats had lower body weight on days 4–12 compared to CON and DOM (P<0.05). Body composition was determined via nuclear magnetic resonance at the start of the VBS and on day 13. Main effects of fat mass loss and social status were seen with SUB and DOM losing significantly more fat mass compared to CON (F2,28=23.995, P<0.05) (Fig. 1B). Similarly, a main effect of social status and lean mass loss was seen with SUB losing significantly more of their original lean body mass compared to CON (F2,28=3.552, P<0.05) (Fig. 1C). There were significant differences in adjusted adrenal weights among social status (F2,28=4.996, P<0.05) with SUB having greater adrenal weights compared to CON.
Fig. 1.

The physiological profile of SUB is consistent with exposure to chronic stress. The VBS had a significant effect on body mass (A), fat mass (B), lean mass (C) and adrenal weight (D). Body mass, fat mass and lean mass measurements were made as a percent change from day 1 VBS. Different symbols indicate significant differences between groups. * p<0.05 SUB vs. CON. # p<0.05 SUB vs. DOM. $ p<0.05 DOM vs. CON. Data are presented as mean±S.E.M. CON=control rats, SUB=subordinate rats, DOM=dominant rats. CON n=8, SUB n=18, DOM n=6.
3.2 Restraint stress test
On day 14, rats were placed into restraint chambers and plasma CORT levels were measured (Fig. 2). There was a significant effect of time on plasma CORT (F2,54=120.11, P<0.001), with all groups displaying increased plasma CORT after 1 h of novel restraint challenge. There was no significant time x social status interactions.
Fig. 2.

Plasma CORT levels were elevated after 60 min of restraint stress. 0-min =before restraint; 60-min =1 h after restraint; 120 =1 h after removal from restraint. No significant differences were noted between groups at each time point. Data are presented as mean±S.E.M.
3.3 GAD67 mRNA expression
GAD67 mRNA was robustly expressed in regions regulating mood, anxiety, and the stress response, including the dorsal and ventral peri-PVN, interfascicular (BNSTif) and principle (BNSTpr) nucleus of the bed nucleus of the stria terminalis, DMH, the CA1, CA3 and DG regions of the hippocampal formation, the prelimbic (PL) and infralimbic (IL) of the mPFC, and the suprachiasmatic nucleus (SCN) (Figs. 3, 4, 5, Table 1). Significant interactions of social status and GAD67 mRNA expression were noted in the dorsal peri-PVN (F2,24=12.674, P<0.05) (Fig. 4A), ventral peri-PVN (F2,23=9.633, P<0.05) (Fig. 4B), the BNSTif (F2, P<0.05) (Fig. 4C), DMH (F2,26=6.355, P<0.05) (Fig. 4D), the CA1 (F2,25=17.079, P<0.05), CA3 (F2,25=17.459, P<0.05) and DG (F2,24=20.768, P<0.05) regions of the hippocampus (Fig. 4E,F,G) and the PL/IL (F2,23=7.281, P<0.05) (Fig. 4H). Post hoc analysis revealed a significant increase in GAD67 mRNA expression in the PL/IL, ventral peri-PVN, CA1 and CA3 regions in the SUB group (P<0.05). A slight but significant increase in GAD67 mRNA hybridization was also observed in the DMH of the SUB group. In contrast, GAD67 mRNA expression was found to be significantly decreased in in the dorsal peri-PVN and BNSTif in DOM. No differences in GAD67 mRNA expression were observed among social status groups in the SCN and the BNSTpr (Table 1).
Fig. 3.
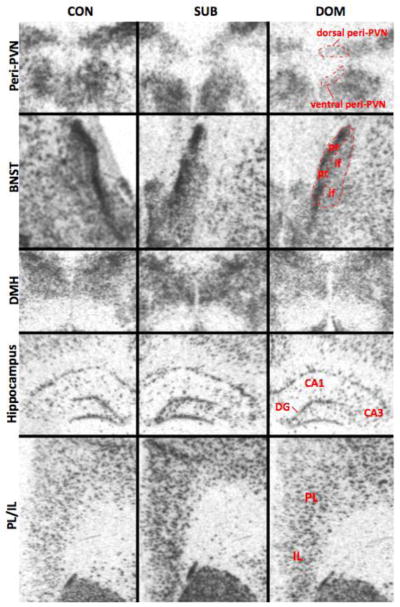
Representative film autoradiograms showing examples of chronic social subordination-induced modulation of GAD67 mRNA expression in several forebrain regions. Top row: Note the apparent decrease in hybridization signal in the dorsal and ventral peri-PVN of DOM (third column) compared to both SUB (middle column) and CON (first column). In addition, GAD67 expression appears reduced in the ventral peri-PVN of SUB vs. CON. Second row: In the bed nucleus of the stria terminalis (BNST), hybridization signal for GAD67 mRNA was apparently reduced in the interfascicular nucleus (BNSTif) of DOM vs. both CON and SUB. Third row: Labeling for GAD67 mRNA was increased in the dorsal medial hypothalamus (DMH) of SUB compared to DOM. Fourth row: In the hippocampal formation, GAD67 mRNA expression appears elevated in the dentate gyrus granule cell layer (DG) and hippocampal CA1 and CA3 regions of SUB compared to both CON and DOM. In addition, GAD67 mRNA hybridization appears decreased in the CA3 region of DOM vs. CON. Bottom row: GAD67 mRNA hybridization is elevated in the prelimbic (PL) and infralimbic (IL) regions of the medial prefrontal cortex of SUB compared to both CON and DOM. Semi-quantitative densitometric analyses of these data are shown in Figure 4. CON=control rats, SUB=subordinate rats, DOM=dominant rats.
Fig. 4.

Densitometric measurements of GAD67 mRNA hybridization in the peri-hypothalamic periventricular nucleus (peri-PVN), the bed nucleus of the stria terminalis (BNST), the dorsal medial hypothalamus (DMH), the hippocampal formation and the medial prefrontal cortex (mPFC). Expression of GAD67 mRNA was decreased in the dorsal peri-PVN (A), ventral peri-PVN (B) and in the interfascicular nucleus of the BNST (BNSTif) (C) in subordinate rats (SUB) and dominant rats (DOM). GAD67 mRNA was up-regulated in the DMH (D), the CA1 and CA3 regions of the hippocampus (E, F), in the granule cell layer of the dentate gyrus (DG) (G), and the prelimbic (PL) and infralimbic (IL) regions of the mPFC (H) in SUB. Hybridization for GAD67 mRNA was also decreased in the CA3 region of DOM compared to both SUB and control rats (CON) (B * p<0.05 SUB vs. CON. # p<0.05 SUB vs. DOM. $ p<0.05 DOM vs. CON. Data are presented as mean±S.E.M.
Fig. 5.

Densitometric measurements of BDNF mRNA hybridization signal in the dorsal bed nucleus of the stria terminalis (dBNST) (A). Subordinate rats (SUB) exhibited increased BDNF mRNA in the dBNST compared to dominant rats (DOM) and control rats (CON). * p<0.05 SUB vs. CON. # p<0.05 SUB vs. DOM. Data are presented as mean±S.E.M. Figure 5B is representative film autoradiograms showing BDNF mRNA expression in the dorsal bed nucleus of the stria terminalis (dBNST) of the social status groups (B). Note the increased hybridization signal present in the dBNST of SUB compared to both CON and DOM.
Table 1.
Densitometric measurements of GAD67 mRNA hybridization signal in the suprachiasmatic nucleus (SCN) and the principal (BNSTpr) nuclei of the bed nucleus of the stria terminalis. No group differences in BDNF mRNA expression were found in the examined regions. Data are presented as mean±S.E.M. of corrected grey levels.
| Brain Region | CON | SUB | DOM |
|---|---|---|---|
| BNSTpr | 117±2 | 118±2 | 113±6 |
| SCN | 115±6 | 112±2 | 109±6 |
3.4 BDNF mRNA expression
BDNF mRNA was robustly expressed in the dorsal BNST, the PVN and the CA1 and CA3 regions and DG of the hippocampal formation (Figs. 5, Table 2). BDNF mRNA expression in the PL/IL, PVN, CA1, CA3 and DG was not significantly different among social status groups (Table 2). However, LSD post hoc analysis indicated a significant increase in BDNF mRNA hybridization in the dorsal BNST in the SUB group (F2,21=4.446, P<0.05) (Fig. 5).
Table 2.
Densitometric measurements of BDNF mRNA hybridization signal in the hippocampal formation, the paraventricular nucleus of the hypothalamus (PVN) and the prelimbic (PL) and infralimbic (IL) regions of the mPFC. No group differences in BDNF mRNA expression were found in examined regions. Data are presented as mean±S.E.M. of corrected grey levels.
| Brain Region | CON | SUB | DOM |
|---|---|---|---|
| PL/IL | 45±4 | 41±4 | 46±3 |
| PVN | 34±3 | 40±2 | 42±3 |
| CA1 | 40±5 | 37±2 | 37±3 |
| CA3 | 43±3 | 41±2 | 41±2 |
| DG | 80±3 | 78±2 | 78±4 |
4. Discussion
Results from this study suggest that prolonged exposure to social subordination is associated with physiological signs of stress and region-specific alterations in BDNF and GAD67 mRNA expression in central stress circuitry. Of note, BDNF was up-regulated in the dorsal region of the BNST, a CRH-rich region thought to be involved in activation of physiological stress responses. GAD67 mRNA expression was increased within the hippocampus and mPFC (regions known to regulate HPA axis inhibition and emotional behavior), and in the DMH, which projects directly to the PVN and regulates activation of ACTH release (DiMicco et al., 2002, Fontes et al., 2011). Collectively, these data support a role for limbic GABAergic interneurons and dorsolateral BNST projection neurons in physiological and behavioral changes associated with social subordination.
As noted in previous studies, the VBS model produced SUB with physiological profiles associated with exposure to chronic stress (Blanchard et al., 1993, Blanchard et al., 1995, Albeck et al., 1997, Choi et al., 2006). DOM were clearly identified by time spent in the open area section of the VBS, little or no weight loss, and smaller adrenal sizes compared to SUB. DOM also received fewer bite wounds than SUB. On the other hand, SUB lost significantly more weight, including lean and fat mass, as compared to CON. DOM also lost a significantly greater amount of fat mass (but not lean mass) than CON. This may reflect the fact that DOM were more physically active (primarily sexual and aggressive behaviors) and had greater physical space to move around relative to SUB. Additionally, all three chambers of each VBS colony had ad libitum access to food and water for the duration of the study. Thus, it is unlikely that SUB would enter into the open area section of the VBS for need of nutrients. Weight loss in this model is, therefore, most likely attributed to exposure to chronic physical and psychological stress.
Along with body weight loss, increased adrenal weight suggests that ongoing/episodic social stress is sufficient to have a cumulative impact on secretion of ACTH. However, at the time of stress testing (day 14) there was no effect of social status on either basal CORT or CORT release during restraint stress. Exposure to VBS is known to increase baseline CORT; however, increases are not seen in all experiments, including those within our group (Nguyen et al., 2007). Thus, it is possible that animals are able to habituate to chronic stress over the two-week period so as to normalize basal CORT activity. Indeed, analysis of wounding reveals that most of the agonistic interactions occurred within the first few days of colony establishment. Thus, although the impact of social stress on somatic and brain endpoints is clear, these do not necessarily translate into frank long-term glucocorticoid hypersecretion.
Additionally, previous VBS studies have delineated between responsive and non-responsive SUB based on CORT secretion to restraint stress (Blanchard et al., 1993, Blanchard et al., 2001). As previously defined, non-responsive SUB have a stress-induced increase of plasma CORT of less than 10 μg/dl compared to the average basal CORT level of all SUB (Albeck et al., 1997). These non-responsive rats also exhibit other signs of HPA hypoactivity, such as reduced CRH mRNA expression in the PVN and central amygdala (Albeck et al., 1997). No SUB in this study, however, clearly met this criterion as a non-responder. One difference between this current investigation and previous VBS studies is that food and water were provided ad libitum in each of the three chambers. Earlier VBS studies provided food and water only in the open field area of the VBS (Blanchard et al., 1993, Blanchard et al., 1995, McKittrick et al., 1995). This design creates an approach-avoidance scenario, whereby SUB must risk antagonistic interactions with a dominant rat to obtain nutrients. Previous VBS studies also reported greater weight loss, more wounds and higher mortality rates amongst SUB (Blanchard et al., 1985, Blanchard et al., 1993, Blanchard et al., 1995). This more intense social stress experience may underlay the greater prevalence of non-responders reported in these studies. In this investigation and other recent VBS studies, however, SUB had greater access to food and water allowing them to spend more time in the smaller tunnels and inner chambers of the VBS, reducing the risk of an attack from a dominant rat (Choi et al., 2006, Tamashiro et al., 2007b, Davis et al., 2009). Despite this alteration, SUB still exhibit a number of physiological signs associated with chronic stress exposure, such as reduced body weight and enlarged adrenals.
Behavioral and endocrine phenotypes associated with SUB or DOM can each have adaptive advantages. For example, SUB show increase caution in risk assessment behaviors (i.e. an animal pokes its head out of the tunnel to investigate the open area chamber of the VBS) after cat exposure in the VBS compared to DOM (Blanchard and Blanchard, 1989, Blanchard et al., 1993). This reduction in risk assessment by SUB is adaptive in some contexts, as it reduces opportunities for exposure to a predator or an aggressive conspecific. Conversely, rats that exhibit decreased anxiety-like behaviors on the elevated plus maze and augmented responses to food reward are more likely to assume dominant status when later placed in the VBS (Davis et al., 2009). This increased risk-taking and reward seeking behaviors by DOM may confer advantages in reproductive opportunities and greater control over limited resources in the environment. Overall, risk-avoidant and risk-taking behaviors likely reflect important tradeoffs, as each has specific adaptive advantages. How stressed-induced alterations in GAD67 and BDNF expression in examined brain regions influence risk-taking and reward-based decision making, however, remains poorly understood. One interesting observation, nonetheless, is that DOM showed marked decreases in expression in the BNSTif, which is in contrast with the tendency for enhanced GAD67 expression in SUB. The BNSTif plays a prominent role in circuitry regulating aggression and submissive behaviors (Cooper and Huhman, 2005, Markham et al., 2009, Hammack et al., 2012). Thus, it is possible that lower GAD67 levels may be linked to aggressive behaviors in the VBS, either as a consequence of dominance status or as an innate behavioral trait.
DOM also manifested lower GAD67 expression in the peri-PVN zones. Decreased GAD67 expression in the peri-PVN suggests attenuated GABAergic input to the PVN, which may be responsible for enhanced CRH mRNA expression observed in DOM groups (Albeck et al., 1997). Interestingly, no changes in peri-PVN GAD67 were observed in the SUB group or in previous studies using non-social chronic stress exposure (Bowers et al., 1998). These data suggest that the role of local GABAergic neurons may differ in SUB vs. DOM, implying differential behavioral and endocrine responses to stress.
A number of studies have implicated BDNF in mood disorders and stress (Hashimoto et al., 2005, Yulug et al., 2009, Bath et al., 2013). Rodent studies indicate that hippocampal BDNF mRNA decreases following both acute and chronic stress (Smith et al., 1995, Umemoto et al., 1997, Russo-Neustadt et al., 2001, Murakami et al., 2005). However, other studies have failed to detect differences in BDNF expression following chronic stress or social defeat, suggesting that BDNF down-regulation may be situation- or stressor-specific (Lauterborn et al., 1995, Kuroda and McEwen, 1998, Hammack et al., 2009, Coppens et al., 2011). In line with the latter observation, we did not observe changes in BDNF mRNA in the hippocampus, suggesting that social subordination (or dominance) is not sufficient to modulate BDNF in this region. We were also unable to detect changes in other key stress-regulatory regions, such as the mPFC and PVN, suggesting that chronic social subordination does not alter BDNF mRNA levels in these regions.
The only region showing significant up-regulation of BDNF was the dBNST. This region has a central role in responding to acute-unexpected threats and anxiety behaviors (Kalin et al., 2005, Pego et al., 2008, Somerville et al., 2010) and is heavily connected with both the amygdala and brainstem stress regulatory regions, such as the nucleus of the solitary tract and ventrolateral medulla (Walker et al., 2003, Dong and Swanson, 2004, 2006). The dBNST also receives synaptic input from the paraventricular nucleus of the thalamus, postpiriform transition area and the insular region, and thus acts as a potential limbic stress integration center (Cullinan et al., 1993, Dong et al., 2001). Notably, the observed up-regulation of BDNF in SUB is consistent with prior studies using a chronic variable stress model (Hammack et al., 2009). An increase in dBNST BDNF is suggestive of ongoing neuroplasticity in BNST circuits, as exposure to chronic stress is known to increase dendritic remodeling in this region (Vyas et al., 2003). Whereas future studies may clarify the role of BDNF within the dBNST, results from this investigation suggest involvement in neuronal changes pursuant to chronic stress exposure.
The fact that both BDNF and GAD67 mRNA expression are altered by social stress suggests modification of BNST signaling is of relevance to behavioral and physiological sequelae of subordination. The ifBNST is a component of the rodent defense circuitry, being activated under conditions of agonist interactions or predator exposure (Canteras, 2002, Figueiredo et al., 2003). Loss of GABA output from this region may be linked to altered coping strategies in the SUB vs. DOM animals. Enhanced BDNF in the dBNST may be linked to stress-induced sprouting. This region of the BNST is known to play a role in autonomic function and behavior with enhanced function predictive of increased anxiety phenotypes (Egli and Winder, 2003, Hammack et al., 2009).
In summary, chronic stress imposed by social subordination status is associated with several region- and paradigm-specific alterations in GAD67 and BDNF mRNA expression in central stress circuitry. In combination with prior studies performed in other models, these data suggest that modulation of limbic GABAergic signaling is a common feature of chronic stress exposure. Modulation of BNST BDNF and GAD67 expression may be linked with specific features of VBS exposure, and may contribute to anxiety phenotypes seen under conditions of chronic stress.
Highlights.
We examine the effects of chronic social stress in the visible burrow system (VBS)
Chronic stress induced changes in the expression of GABAergic synthesizing enzyme
Chronic stress increased expression of BDNF mRNA in the BNST
These changes were region-specific in central stress circuitry, including the BNST
Modulation of BNST BDNF and GAD67 mRNA expression may be linked with anxiety
Acknowledgments
This work was supported by NIH grants T32NS007453, R01 NS060114 (KBS), R01 MH049698 (JPH), The Selma Schottenstein Harris Lab for Research in Parkinson’s, The Gardner Family Center for Parkinson’s Disease and Movement Disorders, The National Science Foundation Graduate Research Fellowship, and The University of Cincinnati Neuroscience Graduate Program. We would also like to thank Dr. Randall Sakai and Dr. Michael Schmeltzer for their guidance on the VBS experiments.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Albeck DS, McKittrick CR, Blanchard DC, Blanchard RJ, Nikulina J, McEwen BS, Sakai RR. Chronic social stress alters levels of corticotropin-releasing factor and arginine vasopressin mRNA in rat brain. Journal of Neuroscience. 1997;17:4895–4903. doi: 10.1523/JNEUROSCI.17-12-04895.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bath KG, Schilit A, Lee FS. Stress effects on BDNF expression: effects of age, sex, and form of stress. Neuroscience. 2013;239:149–156. doi: 10.1016/j.neuroscience.2013.01.074. [DOI] [PubMed] [Google Scholar]
- Bergmann N, Gyntelberg F, Faber J. The appraisal of chronic stress and the development of the metabolic syndrome: a systematic review of prospective cohort studies. Endocrine connections. 2014;3:R55–R80. doi: 10.1530/EC-14-0031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blanchard DC, Sakai RR, McEwen B, Weiss SM, Blanchard RJ. Subordination stress: behavioral, brain, and neuroendocrine correlates. Behavioral Brain Research. 1993;58:113–121. doi: 10.1016/0166-4328(93)90096-9. [DOI] [PubMed] [Google Scholar]
- Blanchard DC, Spencer RL, Weiss SM, Blanchard RJ, McEwen B, Sakai RR. Visible burrow system as a model of chronic social stress: behavioral and neuroendocrine correlates. Psychoneuroendocrinology. 1995;20:117–134. doi: 10.1016/0306-4530(94)e0045-b. [DOI] [PubMed] [Google Scholar]
- Blanchard RJ, Blanchard DC. Antipredator defensive behaviors in a visible burrow system. J Comp Psychol. 1989;103:70–82. doi: 10.1037/0735-7036.103.1.70. [DOI] [PubMed] [Google Scholar]
- Blanchard RJ, Blanchard DC, Flannelly KJ. Social stress, mortality and aggression in colonies and burrowing habitats. Behavioural processes. 1985;11:209–213. doi: 10.1016/0376-6357(85)90062-2. [DOI] [PubMed] [Google Scholar]
- Blanchard RJ, Yudko E, Dulloog L, Blanchard DC. Defense changes in stress nonresponsive subordinate males in a visible burrow system. Physiol Behav. 2001;72:635–642. doi: 10.1016/s0031-9384(00)00449-2. [DOI] [PubMed] [Google Scholar]
- Bowers G, Cullinan WE, Herman JP. Region-specific regulation of glutamic acid decarboxylase (GAD) mRNA expression in central stress circuits. Journal of Neuroscience. 1998;18:5938–5947. doi: 10.1523/JNEUROSCI.18-15-05938.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Canteras NS. The medial hypothalamic defensive system: hodological organization and functional implications. Pharmacology, biochemistry, and behavior. 2002;71:481–491. doi: 10.1016/s0091-3057(01)00685-2. [DOI] [PubMed] [Google Scholar]
- Choi DC, Evanson NK, Furay AR, Ulrich-Lai YM, Ostrander MM, Herman JP. The anteroventral bed nucleus of the stria terminalis differentially regulates hypothalamic-pituitary-adrenocortical axis responses to acute and chronic stress. Endocrinology. 2008a;149:818–826. doi: 10.1210/en.2007-0883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi DC, Furay AR, Evanson NK, Ulrich-Lai YM, Nguyen MM, Ostrander MM, Herman JP. The role of the posterior medial bed nucleus of the stria terminalis in modulating hypothalamic-pituitary-adrenocortical axis responsiveness to acute and chronic stress. Psychoneuroendocrinology. 2008b;33:659–669. doi: 10.1016/j.psyneuen.2008.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi DC, Nguyen MM, Tamashiro KL, Ma LY, Sakai RR, Herman JP. Chronic social stress in the visible burrow system modulates stress-related gene expression in the bed nucleus of the stria terminalis. Physiology & behavior. 2006;89:301–310. doi: 10.1016/j.physbeh.2006.05.046. [DOI] [PubMed] [Google Scholar]
- Cooper MA, Huhman KL. Corticotropin-releasing factor type II (CRF-sub-2) receptors in the bed nucleus of the stria terminalis modulate conditioned defeat in Syrian hamsters (Mesocricetus auratus) Behavioral Neuroscience. 2005;119:1042–1051. doi: 10.1037/0735-7044.119.4.1042. [DOI] [PubMed] [Google Scholar]
- Coppens CM, Siripornmongcolchai T, Wibrand K, Alme MN, Buwalda B, de Boer SF, Koolhaas JM, Bramham CR. Social Defeat during Adolescence and Adulthood Differentially Induce BDNF-Regulated Immediate Early Genes. Frontiers in behavioral neuroscience. 2011;5:72. doi: 10.3389/fnbeh.2011.00072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cullinan WE, Herman JP, Watson SJ. Ventral subicular interaction with the hypothalamic paraventricular nucleus: evidence for a relay in the bed nucleus of the stria terminalis. Journal of Comparative Neurology. 1993;332:1–20. doi: 10.1002/cne.903320102. [DOI] [PubMed] [Google Scholar]
- Davis JF, Krause EG, Melhorn SJ, Sakai RR, Benoit SC. Dominant rats are natural risk takers and display increased motivation for food reward. Neuroscience. 2009;162:23–30. doi: 10.1016/j.neuroscience.2009.04.039. [DOI] [PubMed] [Google Scholar]
- DiMicco JA, Samuels BC, Zaretskaia MV, Zaretsky DV. The dorsomedial hypothalamus and the response to stress: part renaissance, part revolution. Pharmacology Biochemisty & Behavior. 2002;71:469–480. doi: 10.1016/s0091-3057(01)00689-x. [DOI] [PubMed] [Google Scholar]
- Dong HW, Petrovich GD, Watts AG, Swanson LW. Basic organization of projections from the oval and fusiform nuclei of the bed nuclei of the stria terminalis in adult rat brain. Journal of Comparative Neurology. 2001;436:430–455. doi: 10.1002/cne.1079. [DOI] [PubMed] [Google Scholar]
- Dong HW, Swanson LW. Projections from bed nuclei of the stria terminalis, posterior division: implications for cerebral hemisphere regulation of defensive and reproductive behaviors. Journal of Comparative Neurology. 2004;471:396–433. doi: 10.1002/cne.20002. [DOI] [PubMed] [Google Scholar]
- Dong HW, Swanson LW. Projections from bed nuclei of the stria terminalis, anteromedial area: cerebral hemisphere integration of neuroendocrine, autonomic, and behavioral aspects of energy balance. Journal of Comparative Neurology. 2006;494:142–178. doi: 10.1002/cne.20788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egli RE, Winder DG. Dorsal and ventral distribution of excitable and synaptic properties of neurons of the bed nucleus of the stria terminalis. Journal of neurophysiology. 2003;90:405–414. doi: 10.1152/jn.00228.2003. [DOI] [PubMed] [Google Scholar]
- Figueiredo HF, Bodie BL, Tauchi M, Dolgas CM, Herman JP. Stress integration after acute and chronic predator stress: differential activation of central stress circuitry and sensitization of the hypothalamo-pituitary-adrenocortical axis. Endocrinology. 2003;144:5249–5258. doi: 10.1210/en.2003-0713. [DOI] [PubMed] [Google Scholar]
- Fontes MA, Xavier CH, de Menezes RC, Dimicco JA. The dorsomedial hypothalamus and the central pathways involved in the cardiovascular response to emotional stress. Neuroscience. 2011;184:64–74. doi: 10.1016/j.neuroscience.2011.03.018. [DOI] [PubMed] [Google Scholar]
- Hammack SE, Cheung J, Rhodes KM, Schutz KC, Falls WA, Braas KM, May V. Chronic stress increases pituitary adenylate cyclase-activating peptide (PACAP) and brain-derived neurotrophic factor (BDNF) mRNA expression in the bed nucleus of the stria terminalis (BNST): roles for PACAP in anxiety-like behavior. Psychoneuroendocrinology. 2009;34:833–843. doi: 10.1016/j.psyneuen.2008.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammack SE, Cooper MA, Lezak KR. Overlapping neurobiology of learned helplessness and conditioned defeat: implications for PTSD and mood disorders. Neuropharmacology. 2012;62:565–575. doi: 10.1016/j.neuropharm.2011.02.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hardy MP, Sottas CM, Ge R, McKittrick CR, Tamashiro KL, McEwen BS, Haider SG, Markham CM, Blanchard RJ, Blanchard DC, Sakai RR. Trends of reproductive hormones in male rats during psychosocial stress: role of glucocorticoid metabolism in behavioral dominance. Biology of reproduction. 2002;67:1750–1755. doi: 10.1095/biolreprod.102.006312. [DOI] [PubMed] [Google Scholar]
- Hashimoto K, Koizumi H, Nakazato M, Shimizu E, Iyo M. Role of brain-derived neurotrophic factor in eating disorders: recent findings and its pathophysiological implications. Progress in neuro-psychopharmacology & biological psychiatry. 2005;29:499–504. doi: 10.1016/j.pnpbp.2005.01.007. [DOI] [PubMed] [Google Scholar]
- Hemmerle AM, Dickerson JW, Herring NR, Schaefer TL, Vorhees CV, Williams MT, Seroogy KB. (+/−)3,4-methylenedioxymethamphetamine (“ecstasy”) treatment modulates expression of neurotrophins and their receptors in multiple regions of adult rat brain. Journal of Comparative Neurology. 2012;520:2459–2474. doi: 10.1002/cne.23048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalin NH, Shelton SE, Fox AS, Oakes TR, Davidson RJ. Brain regions associated with the expression and contextual regulation of anxiety in primates. Biological Psychiatry. 2005;58:796–804. doi: 10.1016/j.biopsych.2005.05.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kessler RC, Berglund P, Demler O, Jin R, Merikangas KR, Walters EE. Lifetime prevalence and age-of-onset distributions of DSM-IV disorders in the National Comorbidity Survey Replication. Archives of general psychiatry. 2005;62:593–602. doi: 10.1001/archpsyc.62.6.593. [DOI] [PubMed] [Google Scholar]
- Kuroda Y, McEwen BS. Effect of chronic restraint stress and tianeptine on growth factors, growth-associated protein-43 and microtubule-associated protein 2 mRNA expression in the rat hippocampus. Molecular Brain Research. 1998;59:35–39. doi: 10.1016/s0169-328x(98)00130-2. [DOI] [PubMed] [Google Scholar]
- Lauterborn J, Berschauer R, Gall C. Cell-specific modulation of basal and seizure-induced neurotrophin expression by adrenalectomy. Neuroscience. 1995;68:363–378. doi: 10.1016/0306-4522(95)00150-h. [DOI] [PubMed] [Google Scholar]
- Markham CM, Norvelle A, Huhman KL. Role of the bed nucleus of the stria terminalis in the acquisition and expression of conditioned defeat in Syrian hamsters. Behavioral Brain Research. 2009;198:69–73. doi: 10.1016/j.bbr.2008.10.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKittrick CR, Blanchard DC, Blanchard RJ, McEwen BS, Sakai RR. Serotonin receptor binding in a colony model of chronic social stress. Biological psychiatry. 1995;37:383–393. doi: 10.1016/0006-3223(94)00152-s. [DOI] [PubMed] [Google Scholar]
- Murakami S, Imbe H, Morikawa Y, Kubo C, Senba E. Chronic stress, as well as acute stress, reduces BDNF mRNA expression in the rat hippocampus but less robustly. Neuroscience research. 2005;53:129–139. doi: 10.1016/j.neures.2005.06.008. [DOI] [PubMed] [Google Scholar]
- Nguyen MM, Tamashiro KL, Melhorn SJ, Ma LY, Gardner SR, Sakai RR. Androgenic influences on behavior, body weight, and body composition in a model of chronic social stress. Endocrinology. 2007;148:6145–6156. doi: 10.1210/en.2007-0471. [DOI] [PubMed] [Google Scholar]
- Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. San Diago, CA: Academic Press; 1998. [Google Scholar]
- Pego JM, Morgado P, Pinto LG, Cerqueira JJ, Almeida OF, Sousa N. Dissociation of the morphological correlates of stress-induced anxiety and fear. European Journal of Neuroscience. 2008;27:1503–1516. doi: 10.1111/j.1460-9568.2008.06112.x. [DOI] [PubMed] [Google Scholar]
- Radley JJ, Gosselink KL, Sawchenko PE. A discrete GABAergic relay mediates medial prefrontal cortical inhibition of the neuroendocrine stress response. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2009;29:7330–7340. doi: 10.1523/JNEUROSCI.5924-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russo-Neustadt A, Ha T, Ramirez R, Kesslak JP. Physical activity-antidepressant treatment combination: impact on brain-derived neurotrophic factor and behavior in an animal model. Behavioral Brain Research. 2001;120:87–95. doi: 10.1016/s0166-4328(00)00364-8. [DOI] [PubMed] [Google Scholar]
- Sapolsky RM. The Influence of Social Hierarchy on Primate Health. Science. 2005;308:648–652. doi: 10.1126/science.1106477. [DOI] [PubMed] [Google Scholar]
- Seroogy K, Herman J. In situ hybridization approaches to the study of the nervous system. In: Turner A, Bachelard H, editors. Neurochemistry: a practical approach. Vol. 2. Oxford: Oxford University Press; 1997. pp. 121–150. [Google Scholar]
- Smeltzer M, Scott K, Melhorn S, Krause E, Sakai R. Amylin blunts hyperphagia and reduces weight and fat gain during recovery in socially stressed rats. American Journal of Physiology Regulatory Integrative Comparative Physiology. 2012;303:R676–682. doi: 10.1152/ajpregu.00090.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith MA, Makino S, Kvetnansky R, Post RM. Stress and glucocorticoids affect the expression of brain-derived neurotrophic factor and neurotrophin-3 mRNAs in the hippocampus. Journal of Neuroscience. 1995;15:1768–1777. doi: 10.1523/JNEUROSCI.15-03-01768.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Somerville LH, Whalen PJ, Kelley WM. Human bed nucleus of the stria terminalis indexes hypervigilant threat monitoring. Biological Psychiatry. 2010;68:416–424. doi: 10.1016/j.biopsych.2010.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tamashiro KL, Hegeman MA, Nguyen MM, Melhorn SJ, Ma LY, Woods SC, Sakai RR. Dynamic body weight and body composition changes in response to subordination stress. Physiology & Behavior. 2007a;91:440–448. doi: 10.1016/j.physbeh.2007.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tamashiro KL, Nguyen MM, Fujikawa T, Xu T, Yun Ma L, Woods SC, Sakai RR. Metabolic and endocrine consequences of social stress in a visible burrow system. Physiology & Behavior. 2004;80:683–693. doi: 10.1016/j.physbeh.2003.12.002. [DOI] [PubMed] [Google Scholar]
- Tamashiro KL, Nguyen MM, Ostrander MM, Gardner SR, Ma LY, Woods SC, Sakai RR. Social stress and recovery: implications for body weight and body composition. American journal of physiology Regulatory, integrative and comparative physiology. 2007b;293:R1864–1874. doi: 10.1152/ajpregu.00371.2007. [DOI] [PubMed] [Google Scholar]
- Tamashiro KL, Nguyen MM, Sakai RR. Social stress: from rodents to primates. Frontiers in neuroendocrinology. 2005;26:27–40. doi: 10.1016/j.yfrne.2005.03.001. [DOI] [PubMed] [Google Scholar]
- Ulrich-Lai YM, Herman JP. Neural regulation of endocrine and autonomic stress responses. Nature reviews Neuroscience. 2009;10:397–409. doi: 10.1038/nrn2647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umemoto S, Kawai Y, Ueyama T, Senba E. Chronic glucocorticoid administration as well as repeated stress affects the subsequent acute immobilization stress-induced expression of immediate early genes but not that of NGFI-A. Neuroscience. 1997;80:763–773. doi: 10.1016/s0306-4522(97)00050-x. [DOI] [PubMed] [Google Scholar]
- Vahl TP, Ulrich-Lai YM, Ostrander MM, Dolgas CM, Elfers EE, Seeley RJ, D’Alessio DA, Herman JP. Comparative analysis of ACTH and corticosterone sampling methods in rats. American journal of physiology Endocrinology and metabolism. 2005;289:E823–828. doi: 10.1152/ajpendo.00122.2005. [DOI] [PubMed] [Google Scholar]
- Vyas A, Bernal S, Chattarji S. Effects of chronic stress on dendritic arborization in the central and extended amygdala. Brain research. 2003;965:290–294. doi: 10.1016/s0006-8993(02)04162-8. [DOI] [PubMed] [Google Scholar]
- Walker DL, Toufexis DJ, Davis M. Role of the bed nucleus of the stria terminalis versus the amygdala in fear, stress, and anxiety. European journal of pharmacology. 2003;463:199–216. doi: 10.1016/s0014-2999(03)01282-2. [DOI] [PubMed] [Google Scholar]
- Yulug B, Ozan E, Gonul AS, Kilic E. Brain-derived neurotrophic factor, stress and depression: a minireview. Brain Research Bulletin. 2009;78:267–269. doi: 10.1016/j.brainresbull.2008.12.002. [DOI] [PubMed] [Google Scholar]