

Abstract
Connexins are widely distributed proteins in the body that are crucially important for heart and brain function. Six connexin subunits form a connexon or hemichannel in the plasma membrane. Interactions between two hemichannels in a head-to-head arrangement result in the formation of a gap junction channel. Gap junctions are necessary to coordinate cell function by passing electrical current flow between heart and nerve cells or by allowing exchange of chemical signals and energy substrates. Apart from its localisation at the sarcolemma of cardiomyocytes and brain cells, connexins are also found in mitochondria where they are involved in the regulation of mitochondrial matrix ion fluxes and respiration. Connexin expression is affected by age and gender as well as several pathophysiological alterations such as hypertension, hypertrophy, diabetes, hypercholesterolemia, ischemia, post-myocardial infarction remodelling or heart failure, and post-translationally connexins are modified by phosphorylation/de-phosphorylation and nitros(yl)ation which can modulate channel activity. Using knockout/knockin technology as well as pharmacological approaches, one of the connexins, namely connexin 43, has been identified to be important for cardiac and brain ischemia/reperfusion injury as well as protection from it. Therefore, the current review will focus on the importance of connexin 43 for irreversible injury of heart and brain tissue following ischemia/reperfusion and will highlight the importance of connexin 43 as an emerging therapeutic target in cardio- and neuroprotection.
1. General introduction to connexins
Connexins (Cx) are widely distributed proteins in the body that are crucially important for heart and brain function. Connexins are integral membrane proteins which span the plasma membrane four times with N- and C-terminal ends facing the cytosol (Figure 1). Six Cx monomers form a connexon or hemichannel in the plasma membrane. Interactions between the extracellular loops (EL) of two hemichannels in a head-to-head arrangement result in the formation of a gap junction channel (reviewed in Desplantez et al., 2007; Saez et al., 2003). In this process, the two closed hemichannels open, thereby forming a channel that directly connects the cytoplasm of the two adjacent cells allowing the exchange of ions and small molecular weight substances up to ~2 kDa. Gap junctions are assemblies of closely packed gap junction channels that often appear as gap junction plaques containing thousands of gap junction channels. Gap junction plaques are typically observed in the heart at the intercalated disks of adjacent cardiomyocytes (Revel & Karnovsky, 1967; Severs, 1990) where they facilitate electrical current flow that coordinates cardiomyocyte contraction to sustain its pump function (Severs et al., 2004). In the brain, gap junctions communicate electrical signals between neurons (Pereda, 2014) and pass chemical signals and metabolites (glucose, lactate) between glial cells to support the function of the neuronal, glial and vascular cell ensemble in the neurovascular unit (Giaume et al., 2010).
Figure 1. Schematic drawing of the arrangement of Cx43 in the plasma membrane (PM).

Cx43 has a tetraspan topology with two extracellular loops (EL1 and EL2), one cytoplasmic loop (CL) and intracellulary located N- and C-terminal tails (NT, CT). Prototypic Cx43 mimetic peptide sequences are indicated and localized on EL1 (Gap26), EL2 (Gap27) and CL (Gap19). The CT contains a large range of phosphorylation sites that are the target of the various kinases indicated. Illustration generated with the Protter tool (Omasits et al. 2014).
Hemichannels are precursors of gap junctions but they also exist as free, non-junctional channels in the plasma membrane. They are normally closed but may open in response to various triggers including cell depolarization, decreased extracellular calcium ion (Ca2+) concentration, increased intracellular Ca2+ concentration and alterations in the phosphorylation or redox status (D'Hondt et al., 2014; Giaume et al., 2013; Orellana et al., 2013; Saez & Leybaert, 2014). In the brain, glial plasma membrane hemichannels may function as gliotransmitter release pores (Cheung et al., 2014; Montero & Orellana, 2015; Orellana & Stehberg, 2014). Most of the evidence currently available, however, implicates plasma membrane hemichannels as pathological rather than physiological entities, contributing to cell swelling and cell death. In heart and brain cells, excessive hemichannel opening allows the entry of sodium (Na+) and Ca2+ and the escape of potassium (K+), adenosine triphosphate (ATP) and other small metabolites, leading to osmotic shifts, energy depletion, Ca2+ overload and cell death promotion (John et al., 1999; Kondo et al., 2000; Bargiotas Monyer, & Schwaninger, 2009; Contreras, et al., 2004; Davidson et al., 2013; Davidson et al., 2014; Decrock et al., 2009; Li et al., 2001; Orellana et al., 2014; Wang et al., 2013).
Mutations of the Cx protein may lead to congenital diseases; the most frequent one is hearing loss linking to Cx26, Cx30 and other Cxs (Kelly et al., 2014). In the heart atrial fibrillation may result from mutations in Cx40 (Molica et al., 2014). Cxs also play prominent roles in acquired diseases. Under pathological conditions, gap junctions may expand cell injury/cell death to surrounding healthy cells causing bystander (Lin et al., 1998) or “spreading of injury” (Garcia-Dorado et al., 2004) effects. Conversely, gap junctions may also act in a protective manner by supplying essential nutrients and metabolites, thereby preventing injury or death of healthy neighboring cells (Decrock et al., 2009). In cardiac infarction, ischemia triggers a decrease in gap junctional communication as a result of acidosis, increased intracellular Ca2+ concentration ([Ca2+]i) and altered phosphorylation and nitrosylation levels (Fig. 2). This along with alterations in other ion channels, leads to slowed electrical impulse conduction and increased risk for arrhythmias (Cascio et al., 2005; Dhein, 2006; Dhein et al., 2014). In brain ischemia, astrocytic gap junctional coupling decreases, and like in heart, closure is incomplete (Cotrina et al., 1998; Eugenin et al., 2012). Decreased coupling hampers the spatial buffering of potassium and glutamate, and limits sharing of energy substrates from the astrocytic syncytium to neurons (Contreras et al., 2004; Rossi et al., 2007). Unfortunately, the remaining coupling may be sufficient to propagate bystander injury and cell death between astrocytes and between gap junction-coupled neurons (Belousov & Fontes, 2013).
Figure 2. Connexin channels in cardiac ischemia.
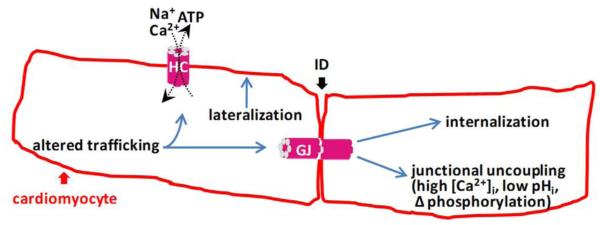
Connexins form gap junctions (GJ) that connect cardiomyocytes with each other at the site of cell-cell junction located intercalated disks (ID). They also form free unapposed hemichannels (HC) in the plasma membrane not incorporated in GJs. Ischemic conditions lead to junctional uncoupling and GJ closure as a result of [Ca2+]i elevation (Dekker et al., 1996; Peracchia, 2004; Xu et al., 2012), acidosis (Ek-Vitorin et al., 1996; Ek et al., 1994), altered phosphorylation status (Ek-Vitorin et al., 2006; Pahujaa et al., 2007) and other ischemia-related factors (Sanchez et al., 2011). In addition to this, connexins are remodeled as a result of processes that involve lateralization of connexin protein (Chkourko, et al., 2012; Kieken et al., 2009), altered trafficking (Remo et al., 2011; Smyth et al., 2010) and internalization (Duffy et al., 2004; Smyth et al., 2014; Sorgen et al., 2004). Most evidence comes from Cx43 which is a major connexin in ventricular cardiomyocytes that is also present in atria (in addition to Cx40). Cx43 hemichannels are normally closed but open in response to ischemia mimicking conditions (Kondo et al., 2000; Contreras et al., 2002), lowered redox status (Retamal et al., 2007; Saez et al., 2010), lowering of extracellular [Ca2+] (Li et al., 1996; Torres et al., 2012), moderate elevation (≤500 nM) of [Ca2+]i (De Vuyst et al., 2009; Ponsaerts et al., 2010; Wang et al., 2012a) and mechanical stress (Batra et al., 2014). They open with alkaninization, close with acidosis (Schalper et al., 2010) and close with above 500 nM [Ca2+]i elevation (Wang et al., 2012a). It is currently not clear how connexin remodeling impacts hemichannel function. GJ closure acts in a protective manner by limiting cell death spread to neighboring cardiomyocytes but may also lead to increased propensity for postischemic arrhythmogenesis because of heterogeneities in conduction velocity and consequent conduction delays. Hemichannel opening may lead to excessive entry of Na+ and Ca2+ and the loss of essential metabolites (ATP and others) from the cells (Saez et al., 2010).
In physiology as well as in pathology, Cx functions are not always linked to their roles as channels (reviewed in Giepmans, 2004; Vinken et al., 2012)). Many Cxs can be anchored to scaffolding proteins via CT interaction. For example, the CT of connexin 43 (Cx43) can interact with the PDZ-domain of zona occludens protein-1 (ZO-1), linking it to the cytoskeleton. CT-ZO-1 interaction may compete with the binding of transcription factors to ZO-1 (for example, ZO-1-associated nucleic acid binding proteins (ZONAB), a transcription repressor) and in this way influence gene expression (Balda & Matter, 2000). Interaction of the CT with β-catenin may influence Wnt signaling while interactions between CT and c-Src (Herrero-Gonzalez et al., 2010) inhibits c-Src-related oncogenic activity and key steps of cell cycle regulation (Zhang et al., 2003). Finally, the Cx43 CT can potentially localize to the nucleus and thereby inhibit cell growth (Dang et al., 2003; Vinken et al., 2012).
The most abundant Cx in brain and heart is Cx43. In the brain, Cx43 is most prominently expressed in astrocytes but is also present in microglial cells (Giaume & Theis, 2010). Astrocytes additionally express Cx30 and microglial cells express Cx32 and Cx36. Oligodendrocytes express Cx32, Cx47 and Cx29, and neurons mainly Cx36. Brain capillary endothelial cells that form the blood-brain barrier mainly express Cx37 and Cx40, with some low Cx43 signal that is increased with inflammation (Cronin et al., 2008; Danesh-Meyer et al., 2012). Neurons do not express Cx43 but they do express Cx36 and Cx45.
In the heart, Cx43 is most prominently expressed in cardiomyocytes of the ventricles, with some signals obtained also in the atria and endothelial cells (Severs et al., 2008). The atria mainly contain Cx40, which is also expressed together with Cx37 in coronary endothelial cells. Apart from Cx43, the special conduction system in the ventricles contains Cx45. In the atrioventricular and sinoatrial node of mice, Cx30.2 is expressed while its orthologous protein Cx31.9 is not detectable in the human cardiac conduction system (Kreuzberg et al., 2008).
Using knockout/knockin and pharmacological approaches Cx43 has been shown to play important roles during cardiac and brain ischemia/reperfusion injury as well as protection from it. Therefore, the current review will summarize the existing evidence for the role of Cx43 in irreversible injury of heart and brain tissue following ischemia/reperfusion highlights Cx43 as an emerging drug target for cardio- and neuroprotection.
2. Regulation of Cx43 in heart and CNS
2.1. Posttranslational modification
With an increasing number of new phosphorylation specific antibodies and constructs, research related to Cx43 regulation has gone past a simple correlation of unspecified phosphorylation-related migration shifts on Western blots to an approach whereby Cx43 phosphorylation can be mechanistically linked to changes in Cx43-interacting protein binding, gap junctional communication, hemichannel function, kinase activity and the underlying signalling pathways affecting cell biological function (Chen et al. 2013; Cooper & Lampe, 2002; Dunn et al., 2012; Dyce et al., 2012; Ek-Vitorin et al., 2006; Johnstone et al., 2009; Johnstone et al., 2012; Lampe et al., 1998b; Lampe et al., 2006; Marquez-Rosado et al., 2012; Solan & Lampe, 2008; Solan et al., 2007; TenBroek et al., 2001).
Cx43, like Cx31, 32, 37, 40, 43, 45, 46, and 50 is a phosphoprotein with clear evidence for phosphorylation at more than 12 serine (S) and tyrosine (Y) sites in the CT region of the protein via at least 6 kinases (Axelsen et al., 2006; Dunn et al., 2012; Ek-Vitorin et al., 2006; Johnstone et al., 2009; Johnstone et al., 2012; Lampe, 1994; Lampe et al., 1998a; Lampe et al., 2006; Marquez-Rosado et al., 2012; Richards et al., 2004; Saez et al., 1998; Solan & Lampe, 2008; Solan et al., 2007; Stagg & Fletcher, 1990; TenBroek et al., 2001). Also like most other Cx, Cx43 has a short half-life reported to be ~2 hours in cell lines and cardiac tissue (Beardslee et al., 1998; Crow et al., 1990; Darrow et al., 1995; Hertlein et al., 1998; Laird et al., 1991; Lampe, 1994; Musil et al., 1990; Musil & Goodenough, 1991), and phosphorylation status has been linked to differences in Cx43 localization and half-life (Axelsen et al., 2006; Dunn & Lampe, 2014; Lampe, 1994; Lampe et al., 1998a; Marquez-Rosado et al., 2012; Richards et al., 2004; Solan et al., 2007). Truncated Cx43 that lacks the CT portion (Cx43K258 stop) can form gap junctions but their channels have different permeability/electrophysiological properties (Dunham et al., 1992; Fishman et al., 1991; Moreno et al., 2002). Furthermore, truncated Cx43 has a prolonged half-life and a knockin mouse expressing truncated Cx43 (Cx43K258stop) died shortly after birth (Maass et al., 2004).
Cx43 phosphorylation at S364 (TenBroek et al., 2001) and S365 (Solan et al., 2007) increases in response to stimuli that enhance gap junction assembly, and phosphorylation at S325/328/330 via casein kinase 1 (CK1) regulates assembly into gap junction channels (Cooper & Lampe, 2002; Lampe et al., 2006).
Cx43 localization and phosphorylation are dramatically modulated during ischemia and injury (Axelsen et al., 2006) most likely depending on the cellular ATP content (Turner et al., 2004).
In isolated rat (Matsushita et al., 2006) and rabbit (Tansey et al., 2006) hearts, prolonged ischemia/hypoxia (>15 minutes) induces sarcolemmal redistribution of Cx43 with a reduction of gap junctional Cx43 [and N-cadherin (Tansey et al., 2006)] and increased expression at the lateral surface of cardiomyocytes (Beardslee et al., 2000). These changes are concomitant with changes in Cx43 phosphorylation: ischemia results in a 8-fold loss of pS365 (Solan et al., 2007) and S325/328/330 (Lampe et al., 2006), and a 5-fold and 3.5-fold increase at S368 (Ek-Vitorin et al., 2006) and S373, respectively (Dunn & Lampe, 2014). The dephosphorylation of S365 in response to ischemia occurs rapidly (5 min) (Sosinsky et al., 2007) followed by increases in S368 phosphorylation, consistent with a “gatekeeper” concept where S365 phosphorylation prevents S368 phosphorylation leading to an inverse relationship in vivo (Solan et al., 2007). Thus, the role of Cx43 phosphorylation in regulating cardiac injury in response to hypoxia appears to be multifold. Proteasomal inhibition increases and stabilizes phosphorylated Cx43 present in gap junctions via increased Akt activity (Dunn et al., 2012), and Akt phosphorylation of Cx43 at S373 (pS373) eliminates ZO-1 interaction and causes a dramatic increase in gap junction size that can be mimicked by expression of Cx43 with a S373D mutation (Dunn & Lampe, 2014). Cx43 is sequentially phosphorylated (Figure 1) by Akt (Dunn & Lampe, 2014), c-Src (Solan & Lampe, 2008), mitogen activated protein kinases (MAPK) (Johnstone et al., 2012) and protein kinase (PKC) (Lampe, 1994; Richards et al., 2004; Solan et al., 2003) in response to growth factors, hypoxia and other stimuli which induce acute gap junction turnover. Gap junction size is negatively regulated by Cx43 interaction with ZO-1 in experiments where the endogenous ZO-1:Cx43 interaction was outcompeted with a Cx43 CT peptide mimetic resulting in a dramatic increase in gap junction size (Hunter et al., 2005). During ischemia, S373 phosphorylation is coincident with an increase in gap junction size and precedes internalization (Dunn & Lampe, 2014). Internalized Cx43 in isolated mice hearts following 30 min global ischemia shows increased phosphorylation at Ser373 and Ser368 which enables the binding of the 14-3-3 protein leading to ubiquitination of Cx43 (Smyth et al., 2014), a process which can be attenuated by protein kinase C inhibition (Liao et al., 2013).
A gap junction can be internalized in its entirety via formation of a double membrane structure termed an annular junction (Archard & Denys, 1979; Fong et al., 2012; Johnson et al., 2013; Jordan et al., 2001; Laird, 2006; Leithe et al., 2006; Nickel et al., 2013; Piehl et al., 2007; Severs et al., 1989) or via loss of extracellular Cx interactions followed by endocytosis of the remaining connexon from a single membrane. During internalization, Cx43 can be phosphorylated by c-Src and inhibition of c-Src activity via protein phosphatase 2 blocks growth factor-induced gap junction turnover (Gilleron et al., 2008; Spinella et al., 2003). Glycyrrhetinic acid-related gap junction inhibitors remodel gap junctions into a looser packing arrangement (Goldberg et al., 1996) in a process that involves c-Src binding (Chung et al., 2007) and leads to disruption of Cx43-ZO-1 interaction (Gilleron et al., 2008). C-Src can directly interact with ZO-1 and compete for binding to the CT region of Cx43 (Gilleron et al., 2008; Kieken et al., 2009; Sorgen et al., 2004). Clearly, c-Src plays a role in gap junction turnover, but whether c-Src phosphorylation of Cx43 plays a direct role is not known. C-Src phosphorylation of the N-methyl-d-aspartate (NMDA) receptor, GluN3A (Chowdhury et al., 2013), triggers its endocytosis. Whether c-Src phosphorylation can direct the endocytic route of internalization through annular junction formation or by “unzippering” gap junctions via loss of extracellular interactions followed by endocytosis is unknown. The formation of double membrane endocytic vesicles (i.e., annular junctions) appears to be fairly specific to gap junctions though there are a few reports of “trans-endocytosis” occurring in dendritic cells (Spacek & Harris, 2004) and in response to receptor ligand complex formation during neural (Marston et al., 2003) and Drosophila development (Klueg et al., 1998). Cx43-containing annular junctions have been well-documented (Archard & Denys, 1979; Fong et al., 2012; Johnson et al., 2013; Jordan et al., 2001; Laird, 2006; Leithe et al., 2006; Nickel et al., 2013; Piehl et al., 2007; Severs et al., 1989) but the role Cx43 phosphorylation plays is still unclear.
Knockin of Cx43 phosphoserine-site mutants (S to A, resulting in a non-phosphorylation mutant version of Cx43 at that site) into mice are of particular interest to determine the functional necessity for Cx43 phosphorylation in vivo. Three knockin mice lines with Cx43 mutations at sites phosphorylated by MAPK, CK1 and PKC have been generated, all knocked-in under the endogenous Cx43 promoter (referred to as Cx43PKC, Cx43CK1 and Cx43MAPK knockin mouse lines) (Huang et al., 2011). All 3 knockin mice lines show different changes in Cx43 phosphorylation, different migration in SDS-PAGE, and varying gap junction stability (e.g., two distinctly derived lines of the Cx43PKC KI mouse show 60% reduced Cx43 levels in heart). The Cx43CK1 mice where the S325/328/330 serines are converted to glutamates were resistant to pathological remodeling and to the induction of ventricular arrhythmias while the line with conversion to alanine were prone to it (Remo et al., 2011). The Cx43MAPK mice show dramatically reduced vascular smooth muscle cell proliferation and neointima formation during arteriole injury (Johnstone et al., 2012).
These results prove that Cx43 phosphorylation plays a key role in modulation of the response to injury in heart and brain in vivo and that these findings might translate into putative therapeutic applications in the future.
2.2. Expression of Cx43
Estrogen (Yu et al., 1994) and triiodothyronine (Stock and Sies, 2000) directly bind to the promoter region of the Cx43 gene thereby increasing its expression. Other transcription factors involved in the regulation of Cx43 expression in cardiomyocytes are TBX18 (family member of T-box transcription factor suppressing Cx43, Kapoor et al., 2011), MSX1/2 (Msh homebox suppressing Cx43, (Boogerd et al., 2008)), AP1 and CREB (cAMP responsive element binding protein stimulating Cx43, (Salameh et al., 2009)) or NKx2.5 (suppressing Cx43, (Kasahara et al., 2003)). AP1 and CREB are translocated to the nucleus secondary to the activation or PKA or MAPK, such as p38 or extracellular regulated kinase (Erk) 1/2 (Salameh et al., 2009).
Cyclic stretch increases N-cadherin expression at the longitudinal sites of cardiomyocytes, a process requiring rho/rac (Matsuda et al., 2006). Loss of N-cadherin reduces Cx43 expression at gap junctions (Li et al., 2008;Li et al., 2005;Palatinus et al., 2011;Zhu et al., 2010). As cyclic stretch increases N-cadherin in cardiomyocytes, it also increases total Cx43 expression and polarization of Cx43 to the longitudinal sites (Salameh et al., 2012). The increases in Cx43 expression and localisation to gap junctions in stretched cardiomyocytes involve angiotensin II, Akt, Erk1/2 and glycogen synthase kinase (GSK) 3ß (Salameh et al., 2012;Salameh et al., 2010a;Salameh et al., 2010b;Shyu et al., 2001). Angiotensin II also increases Cx43 expression (Dodge et al., 1998;Polontchouk et al., 2002) and N-cadherin expression in cardiomyocytes (Adam et al., 2010).
In addition to angiotensin II, other stimuli can also increase Cx43 expression in cardiomyocytes including norepinephrine (Salameh et al., 2006;Salameh et al., 2008), endothelin 1 (Polontchouk et al., 2002), low concentration of aldosterone (Suzuki et al., 2009) and tumor necrosis factor alpha (Salameh et al., 2004). However, long-term treatment with stimuli inducing cardiomyocyte hypertrophy might also activate signaling cascades which suppress Cx43 expression; indeed, norepinephrine caused by activation of c-Jun N-terminal kinase (JNK)1/2 attenuates Cx43 expression over time, an effect that also involves the sodium-proton and sodium-calcium exchanger (Stanbouly et al., 2008). Over-expression of active JNK decreases Cx43 expression at gap junctions (Petrich et al., 2002) and may activate Cx43 degradation through ubiquitination and proteosomal degradation (Mollerup et al., 2011). Similar to norepinephrine, endothelin-1 increases cardiomyocyte size more than Cx43 expression, leading to a relative reduction in sarcolemmal Cx43 expression and cardiomyocyte coupling (McSpadden et al., 2009). Cx43 in cardiomyocytes is not only involved in cellular coupling via gap junctions but through formation of hemichannels that play a role in basal calcium signaling (Li et al., 2012). While signaling in cardiomyocytes is important, the interaction of cardiomyocytes with other cell types may modify these responses. Prolonged stimulation of cardiomyocytes with isoprenaline increases Cx43 expression through activation of Erk1/2 (Salameh et al., 2013;Salameh et al., 2009); however, in co-cultures of cardiomyocytes and fibroblasts, paracrine factors released by fibroblasts suppress the isoprenaline-induced increase in Cx43 expression in cardiomyocytes (Pedrotty et al., 2009;Salameh et al., 2013). Such suppression can partially be reversed by blocking angiotensin II or PKC (Pedrotty et al., 2009), and in cardiac tissue from mice, inhibition of the renin angiotensin system increases Cx43 expression (Iravanian et al., 2011).
2.3. Ischemia/hypoxia-induced alterations of Cx43 expression
Hypoxia of cardiomyocytes changes the Cx43 expression and phosphorylation time-dependently. During short periods of hypoxia (up to 15 minutes) Cx43 content remains unchanged (Matsumura et al., 2006;Zeevi-Levin et al., 2005). With prolonged hypoxia (several hours), downregulation of Cx43 at gap junctions occurs (Danon et al., 2010), Cx43 is internalized (Sato et al., 2009) and subsequently the total cellular Cx43 content decreases (Zeevi-Levin et al., 2005).
Metabolic inhibition activates c-Src kinase, an effect inhibited by 17ß-estradiol (Chung et al., 2009). Furthermore, 17ß-estradiol treatment preserves Cx43 phosphorylation at PKC epitopes during metabolic inhibition (Chung et al., 2004) thereby contributing to maintained localization of Cx43 to gap junctions.
In cardiomyocytes, the muscarinic M3 receptor is associated with Cx43 (Yue et al., 2006) and the hypoxia-induced decrease in total Cx43 expression is attenuated by acetylcholine (Zhang et al., 2006). Similarly, proteasome inhibitors attenuate the hypoxia-induced decline in Cx43 expression (Laing and Beyer, 1995;Zhang et al., 2006).
In summary, stimulation of Cx43 expression in Cx43 deficient pathologies may be a therapeutic option to reverse loss of Cx43.
3. Pharmacological modulation of Cx43-formed channels
The available arsenal of pharmacological agents modulating (mostly inhibiting) gap junctions and hemichannels is extensive and contains chemically diverse compounds. A large group is formed by small molecule inhibitor substances; most of these have targets other than Cx and are therefore non-selective. This group (reviewed in (Bodendiek & Raman, 2010)) includes glycyrrhetinic acid and its derivative carbenoxolone (steroid-like structures from Glycyrrhiza also exhibit anti-inflammatory and mineralocorticoid effects), long-chain alcohols like heptanol and octanol (having general biophysical effects at the level of the plasma membrane), halothane (an inhalational general anesthetic), fatty acids like arachidonic acid (a poly-unsaturated fatty acid present in plasma membrane phospholipids and involved in phospholipase C (PLC) and inflammatory signaling) and oleic acid (a mono-unsaturated fatty acid with various biological effects, abundantly present in olive oil), fatty acid amides like oleamide and anandamide (targeting cannabinoid receptors), fenamates like flufenamic acid, niflumic acid and meclofenamic acid (non-steroid anti-inflammatory molecules), and quinine, its stereo-isomer quinidine and its derivative mefloquine (antimalarial drugs). Several other compounds that inhibit gap junctions are best known for their other actions, like 2-aminoethoxydiphenyl borate (2-APB, a blocker of inositol trisphosphate (IP3) receptors and store-operated Ca2+ entry (Bootman et al., 2002)), polyamines like spermine and spermidine (acting on NMDA channels and several other targets), 5-Nitro-2-(3-phenyl-propylamino)benzoic acid (NPPB, a chloride channel blocker), disodium 4,4'-diisothiocyanatostilbene-2,2'-disulfonate (DIDS, an anion transport blocker), and certain triphenylmethanes, triphenylethanes, triarylmethanes and cyclodextrins. Importantly, most of these substances have been characterized for their gap junction-inhibiting effects but they invariably also inhibit hemichannels. The mode of action of arachidonic acid is different because it inhibits gap junctions but promotes hemichannel opening (Contreras et al., 2002; De Vuyst et al., 2007; De Vuyst et al., 2009). Furthermore, quinine has been reported to activate hemichannels as well (based on ATP release measurements (Stout et al., 2002)). Many of the above listed compounds additionally block channels composed of pannexins, including carbenoxolone, flufenamic acid (Bruzzone et al., 2005), NPPB and DIDS. Of note, low concentrations of carbenoxolone (5 μM) preferentially inhibit pannexin channels while higher concentrations (50 μM) are needed to also inhibit connexin channels (Schalper et al., 2008).
Because of the lack of specificity of small molecule inhibitors of Cx channels, substantial interest has been directed to peptides as agents with better selectivity potential. AAP10 peptide is one of the first peptides reported to have connexin modulating properties, and it increased gap junctional coupling. Later, peptides identical to certain Cx domains called Cx mimetic peptides, were introduced as agents with better specificity towards Cx channels.
AAP10 is a 6 amino acid peptide that was found to promote gap junctional coupling (Grover & Dhein, 1998, 2001; reviewed in Dhein et al., 2010; De Vuyst et al., 2011). It binds with nanomolar affinity to a yet unindentified membrane G-protein coupled receptor (typical active AAP10 concentrations are 10-50 nM), thereby resulting in the activation of certain protein kinases. The junctional coupling-promotive effect has been demonstrated to be mediated by activation of PKCα. There is some selectivity in the effect on gap junctions, with Cx43 and Cx45 junctional coupling being promoted and Cx40 not affected. Rotigaptide (ZP123) is a more stable version of AAP10 composed of D-amino acids concatenated in reverse order, resulting in higher stability after administration to animals. These peptides have been shown to be effective against ischemia-induced ventricular fibrillation and possibly also atrial fibrillation (see Dhein et al, 2010). Gap-134 is a small molecule version designed based on insights obtained from the pharmacophore of AAP10. This substance has been tested in phase I clinical trials on healthy volunteers and was demonstrated to be safe (NCT00783341, 2009). Conclusive information on phase II clinical trials is lacking. Of note, AAP10 promotes hemichannel opening as judged from ATP release studies performed in cardiomyocytes exposed to ischemia-mimicking conditions (Clarke et al., 2009). Uncontrolled opening of Cx43 hemichannels can mediate Na+ or Ca2+ entry and loss of intracellular K+, so any potential side effects of these compounds will have to be carefully followed.
Cx mimetic peptides were introduced over 20 years ago with the aim of finding sequences that had specific physiological effects (mainly channel block) on Cx channels. The first Cx mimetic peptides described were identical to sequences on extracellular loop (EL) 1 or 2. Peptide B/1 (VCYDHFFPISHVR) on EL1 and peptide5 (SRPTEKTVFTV) on EL2 of Cx32 were most effective in delaying synchrony in spontaneously beating chick cardiomyocytes in a myoball assay (Warner et al., 1995). These two peptides contain the VCYD and SHVR sequences on EL1, and the SRPTEK sequence on EL2 that are conserved between Cx32, Cx42, Cx43 and Cx45. For Cx43, the corresponding sequences are VCYDKSFPISHVR, called Gap26, and SRPTEKTIFII, called Gap27 (Figure 1). Because these sequences are very well conserved between different Cx, it comes to no surprise that Cx43-based mimetic peptides also inhibit gap junctions composed of other Cx. For example, the Cx43 sequence of Gap27 (sometimes denoted as 43Gap27) also inhibits gap junctions composed of Cx37 (Chaytor et al., 2001). However, it has not been systematically investigated whether Cx other than Cx37 and Cx43 are also targeted by 43Gap27. The Gap26/Gap27 sequences are not found in other proteins, which may be key for their specificity for the Cx family rather than non-Cx targets. However, 20 years after the discovery that Gap-peptides can block gap junctions, it still remains unknown which domains on the extracellular loops they interact with so the actual mechanism of how Gap26/27 peptides inhibit gap junctions is still elusive. They do not disrupt existing gap junctions but rather act by preventing the formation of new gap junctions (Evans & Boitano, 2001). The peptides thus probably interact with the ELs on free hemichannels and thereby hinder/prevent the docking and assembly of two hemichannels into new gap junction channels. Gap26/27 also inhibit conductance of hemichannels (Braet, Vandamme, Martin, Evans, & Leybaert, 2003). In fact, inhibition of hemichannels occurs faster, within minutes, than inhibition of gap junctions which takes something in the range of hours, at least in cell culture assays (Decrock et al., 2009). In terms of diffusion, minutes is very long suggesting that the interaction site is poorly accessible (Wang et al., 2012). Single-channel patch clamp studies of Cx43 hemichannel block indicate a half-maximal inhibitory concentration of 81 μM for Gap26 and 161 μM for Gap27. However, inhibition obtained with Gap27 is stronger as compared to Gap26. Single-channel analysis further demonstrated that Gap26/27 do not cause hemichannel pore block (provided their concentration is below 1 mM) and do not influence the unitary hemichannel conductance of ~220 pS (Wang et al., 2012). Instead, Gap26/27 appear to act on channel gating as they reduce the open probability and increase the voltage threshold for electrical activation. Hemichannels can also be activated by chemical signals, for example by an increase in the intracellular Ca2+ concentration. It is not entirely clear how Gap26/27 inhibit chemically-triggered hemichannel opening but a likely scenario is that they interact with a binding site outside the permeation pathway of the closed hemichannel and thereby lock the channel in a stabilized closed state (Wang et al., 2013).
3.1. Blocking hemichannels without blocking gap junctions
As mentioned earlier, most gap junction blockers, including carbenoxolone/glycyrrhetinic acid-derived molecules also block hemichannels and are thus not hemichannel-specific (Takeuchi et al., 2011). Lanthanum ions (La3+) block hemichannels and do not inhibit gap junctions (Anselmi et al., 2008) but this trivalent ion also blocks Ca2+ channels (Mlinar & Enyeart, 1993; Young et al., 2002). Gadolinium (Gd3+) is another trivalent ion hemichannel blocker (Stout et al., 2002). These trivalent ions can be used for in vitro work on cells with low expression of Ca2+ channels but they are not appropriate tools to block hemichannels in a complex in vivo setting. Gap26/27 peptides are also useful, provided cell exposure is short enough (tens of minutes) to prevent effects on gap junctions. Peptide5 (VDCFLSRPTEKT (Figure 1) [see Table 1], not to be confused with peptide5 from the Warner et al. (1995a) study, overlaps with Gap27 (SRPTEKT) but lacks the last 4 C-terminal amino acids and has 5 extra amino acids N-terminally. This peptide was identified by testing various slightly overlapping mimetic sequences of the Cx43 EL2 for their potential to inhibit the swelling of rat pup spinal cord segments kept in organotypic culture; peptide5 more than halved swelling after 1 or 4 days in culture. Propidium iodide hemichannel dye uptake assays with a low extracellular Ca2+ concentration showed hemichannel block at 5 μM while calcein dye spread studies indicated gap junction block at 500 μM (O'Carroll et al., 2008). Interestingly, peptide5 also reduced Cx43 expression and had beneficial effects on glial fibrillary acidic protein (GFAP) expression in astrocytes and neuronal nuclei (NeuN) and SMI-32 (marker of nonphosphorylated neurofilaments) expression. The distinct concentration-dependence of hemichannel and gap junction block indicates there may be two different, yet unknown interaction sites for peptide5. It should be noted that, although peptide5 mimics a sequence on EL2 of Cx43, this does not mean that this peptide is necessarily selective for this particular Cx – further testing of its selectivity is therefore indicated.
Table 1.
Connexin mimetic peptides
| Peptide | Cx43 sequence (human) |
|---|---|
| Gap26 | VCYDKSFPISHVR |
| Gap27 | SRPTEKTIFII |
| Peptide5 | VDCFLSRPTEKT |
| L2 | DGVNVDMHLKQIEIKKFKYGIEEHGK |
| Gap19 | KQIEIKKFK |
Recent work has identified two new peptides, called L2 and Gap19, (Figure 1) which specifically block Cx43 hemichannels without inhibiting gap junctions. In contrast to Gap26/27 and peptide5, these two peptides mimic sequences located at the intracellular side of the Cx43 protein which is known to vary widely between different connexins. The L2 sequence is located on the cytoplasmic loop (CL) of Cx43 and the Gap19 sequence is a 9 amino acid stretch within the L2 domain (Wang et al., 2013) (Table 1)(L2 sequence: DGANVDMHLKQIEIKKFKYGIEEHGK, Gap19 sequence underlined). The L2 region is involved in CL interactions with the CT and the short Gap19 sequence is a crucial domain for this interaction. CT-CL interaction results in closure of gap junctions, according to a putative ball-and-chain model (reviewed in (Delmar et al., 2004)). Exogenous addition of L2-derived RXP Cx43-binding peptides prevents CT-CL interaction and thereby prevents gap junction closure induced by acidification or octanol (Verma et al., 2009), i.e., L2 peptide acts to stabilize the open state of gap junctions. Surprisingly, the L2 peptide, in contrast to its effects on gap junctions, blocks Cx43-based hemichannels (Ponsaerts et al., 2010). Gap19 displays similar effects: inhibition of Cx43 hemichannels while not affecting gap junctions upon short exposure (30 min) but slightly stimulating them upon longer exposures (24-48 hrs) (Wang et al., 2013). In depth mechanistic investigations based on single-channel patch clamp experiments, surface plasmon resonance (SPR) and amino acid substitution experiments showed that CT-CL interactions are necessary for hemichannel opening (Abudara et al., 2014; D'Hondt et al., 2013; Iyyathurai et al., 2013; Ponsaerts et al., 2012; Ponsaerts et al., 2010; Ponsaerts et al., 2012; Wang et al., 2013). CT-CL interactions by themselves do not trigger hemichannel opening per se but bring the hemichannels into an available-to-open state that allows their opening by other stimuli of electrical or chemical nature (Saez & Leybaert, 2014; Wang et al., 2013). Moreover, Gap19 specifically acts on Cx43 hemichannels while not influencing Cx40 hemichannels or pannexin 1 channels (Wang et al., 2013). Thus, Gap19 and L2 peptides are interesting tools to investigate contributions of Cx43 hemichannels in brain and heart diseases, in particular ischemia and/or inflammatory conditions. L2 is not membrane permeant but Gap19 has some intrinsic plasma membrane permeability related to its high lysine content (4 out of 9 amino acids are lysines). Both Gap19 and L2 peptides can be linked to the TAT membrane translocation motif to improve/facilitate cell entry and access to the cytoplasmic Cx43 CT target. Determination of the half-maximal hemichannel inhibition concentration for the TAT-linked versions of these peptides gave values of ~10 μM for TAT-L2 and ~7 μM for TAT-Gap19, which are in good agreement to the Kd values for both peptides to interact with the CT determined from SPR experiments. Other novel interesting molecules, for example the linear peptide RRNYRRNY, the cyclic peptide CyRP-71 and the peptidomimetic ZP2519 target the Cx43 CT and act to prevent Cx43-based gap junction closure (De Vuyst et al., 2011; Verma et al., 2009; Verma et al., 2010). These substances are of potential translational value because they prevent gap junction closure and thereby preserve the physiological roles of gap junctions while they are expected (but still not proven) to inhibit hemichannel opening (by preventing CT-CL interaction).
Another approach for blocking hemichannels is the use of antibodies directed against the extracellular portions of the Cx proteins. Antibodies against the ELs have been generated for Cx26 (Clair et al., 2008) and Cx43 (Baklaushev et al., 2009; Riquelme et al., 2013; Hofer & Dermietzel, 1998; Siller-Jackson et al., 2008). Antibodies raised against the EL2 of Cx43 (Figure 1) have higher affinity than those against EL1, hence EL2 antibodies have been most frequently used. EL2 antibodies have been raised against the sequence T186 – K206 (polyclonal Ab, (Hofer & Dermietzel, 1998)), Y185 – K206 (polyclonal Ab, (Siller-Jackson et al., 2008)) or Q173-I208 (monoclonal Ab, (Baklaushev et al., 2009)) which include part of the Gap27 sequence (SRPTEKTI for Q173-I208). These antibodies block hemichannels as determined in assays of low extracellular Ca2+-triggered Lucifer Yellow dye uptake in astrocytes (Hofer & Dermietzel, 1998) and fluid shear stress-induced prostaglandin E2 release in osteocytes (Siller-Jackson et al., 2008). Because they are bulky, it is conceivable that hemichannel block is caused by pore obstruction. Additionally, EL-antibodies invariably also inhibit gap junctions: they interact with the ELs of free hemichannels and thereby prevent the docking and formation of gap junctions (Riquelme et al., 2013). Like for Gap26/27 peptides, gap junction block occurs with some latency which likely depends on the turn-over rate of the Cx protein (antibody-bound hemichannels cannot form new gap junction channels resulting in slow disappearance of junctional coupling). Although EL-antibodies have significantly higher affinities than EL-peptides, they may suffer from the same specificity limitations because of the conserved nature of the EL sequences (for example: Cx32 EL2 has a homology score of ~70% with Cx43 EL2, (Hofer & Dermietzel, 1998)). EL2 antibodies do however not block pannexin channels (Siller-Jackson et al., 2008) that have strongly different sequences compared to Cx.
4. Ischemia/reperfusion injury and neuroprotection in the central nervous system
4.1. Irreversible brain injury
Numerous studies of the effect of gap junction blockers and knockout of specific Cx have been performed on models of brain ischemia and central nervous system trauma. Most of the gap junction blockers used have off-target effects (see 5. Pharmacological modulation of Cx43-formed channels) and also affect hemichannels, making mechanistic interpretation of the reported results difficult.
Early studies using halothane reported a reduction in infarct size in a mid-cerebral artery occlusion (MCAO)/reperfusion model in rats and cats (Saito et al., 1997; Warner et al., 1995). Intraperitoneal injection of octanol 30 min before MCAO (without reperfusion) in rats reduced infarct size measured 24 hrs later by 50% (Rawanduzy et al., 1997). A similar octanol treatment schedule gave a 30-50% reduction of cell death in the hippocampus following transient (10 min) bilateral clamping of the common carotid arteries in rats (Rami et al., 2001). In a model of transient (12.5 min) intrauterine global hypoxia applied to at term pregnant rats, carbenoxolone administration reduced pup mortality by 40% 24 hrs later and by 55% 21 days later. Caspase-3 activation in the brain was reduced by 57% 24hrs after hypoxia (de Pina-Benabou et al., 2005). Using a model of transient (18 min) bilateral global ischemia (4 vessel occlusion), carbenoxolone and 18α-glycyrrhetinic administered 30-40 min before ischemia directly into the hippocampus reduced cell death by 27% in the CA1 region 5 days after ischemia (Perez Velazquez et al., 2006). Interestingly, treatment was also effective when substances were applied 30-40 min after ischemia induction but not when applied at 90 min post-induction, indicating that the first 1.5 hrs of ischemia/reperfusion are critical for protection by carbenoxolone/18α-glycyrrhetinic treatment.
While the previous studies with Cx channel blockers reported improved outcomes, subsequent studies in knockout animals showed the opposite result. Permanent MCAO gave a 83% larger infarct size and 120% larger rate of apoptosis (TUNEL) in the penumbra zone (zone around the ischemic core) when applied to global heterozygous Cx43 knockout mice as compared to wildtype mice (Nakase et al., 2003; Siushansian et al., 2001). Interestingly, astrogliosis was reduced in heterozygous Cx43 knockout mice, which might result from diminished astrocytic intercellular Ca2+ wave activity (De Bock et al., 2014). Follow-up work by the same group in astrocyte-specific Cx43 knockout mice confirmed these results: larger infarct size and rate of apoptosis in the penumbra (Nakase et al., 2004). An up-regulation of astrocytic Cx30 (the second most prevalent Cx in astrocytes) in heterozygous Cx43 knockout mice was reported (Nakase et al., 2003) but no data were included on the effect of Cx30/Cx43 double knockout animals on infarct size and astrogliosis. The impact of CT-truncation of Cx43 (Cx43 K258stop; Cx43ΔCT/+) was studied in the MCAO model. Infarct volume was twice as large in Cx43ΔCT/+ and Cx43ΔCT/− as compared to wildtype mice (Kozoriz et al., 2010). Astrogliosis was reduced by 30% (Cx43ΔCT/+) to 50% (Cx43ΔCT/−) and microglial infiltration of the peri-infarct zone was almost twice as large in Cx43ΔCT/+ and Cx43ΔCT/− compared to wildtype mice. Interpretation of these results is more difficult, as the CT-truncation used affects channel as well as non-channel functions of Cx43. The effect of Cx43ΔCT/+ in astrocyte cultures demonstrated slightly reduced gap junctional coupling in Cx43ΔCT/+ and a more pronounced reduction in coupling (halving) in Cx43ΔCT/−. A subconductance state of gap junctional gating appeared to be lost. Hemichannels were also tested and propidium iodide dye uptake triggered by a low extracellular Ca2+ concentration suggested a doubling of hemichannel function in Cx43ΔCT/+ and halved in Cx43ΔCT/− relative to wildtype mice. Intercellular Ca2+ waves, which are propagated by both gap junction and hemichannel related mechanisms (Leybaert & Sanderson, 2012) were normal in Cx43ΔCT/+ but reduced in Cx43ΔCT/−. Overall, the infarct size was increased to a comparable degree in both Cx43ΔCT/+ and Cx43ΔCT/− while the effect on channel functions was rather diverse. Although fine-detailed interpretation of these results has been proposed in terms of channel functions of Cx43 (Kozoriz et al., 2010), overall it looks like the increased brain infarct size associated with CT-truncation has more to do with an influence on non-channel functions of Cx43.
Remarkably, no reports are available investigating the use of Gap26/27 as potential protective agents against ischemic injury in the brain, while these peptides display clear cardioprotective potential (Hawat et al., 2010; Hawat et al., 2012). In part, this is related to the fact that these peptides cannot pass the blood-brain barrier (BBB) although they might pass through once the BBB has lost its integrity (molecular weight of Gap26 is 1551 and for Gap27 1305). Importantly, Cx in BBB endothelial cells are an accessible and promising target for neuroprotection purposes in their own right. For example, intravenously administered Gap27 protected the BBB against bradykinin-induced leakage by inhibiting oscillations in the endothelial cytoplasmic Ca2+ concentration (De Bock et al., 2011). Ca2+ entry via Cx hemichannels (Cx43 but probably also Cx37) contributed to endothelial Ca2+ oscillations and inhibiting this Ca2+ entry pathway with Gap27 preserved BBB function (De Bock et al., 2011; De Bock et al., 2012; De Bock et al., 2013). Peptide5, which has some sequence overlap with Gap27 (see Table 1) and blocks hemichannels at 5 μM concentration has been demonstrated to be a potential neuroprotectant in various in vivo models of ischemia in brain as well as in spinal cord injury. In most cases, application was done directly to the neuronal tissue to circumvent problems of limited BBB passage of this peptide. In a model of transient 30 min ischemia by bilateral clamping of the carotid arteries in fetal sheep, peptide5 when infused intraventrically 90 min after ischemia for 1 hr or 25 hrs, improved oligodendrocyte survival and prevented brain weight loss associated with ischemia (Davidson et al., 2012). Neuronal cell loss was partly but not significantly reduced. Making use of the same model, the authors compared intraventricular peptide5 infusion starting 1 hr before and applied during ischemia or starting 90 min after ischemia and infused for the next 25 hrs. Only the latter post-ischemia treatment protected significantly against neuronal cell loss in the cortex and oligodendrocyte loss in various white matter regions (Davidson et al., 2013). Neuronal cell loss in the most vulnerable hippocampal CA1 and CA3 regions was however not prevented by any of those treatments. Overall, this work demonstrates that peptide5 action is most prominent after ischemia, presumably by preventing hemichannel opening and thereby limiting injury spread in gray and white matter during reperfusion. Intraperitoneal administration of peptide5 at the start of reperfusion (100 μM estimated systemic concentration) limited Evans blue dye leakage from blood vessels 4 hrs and 24 hrs later and prevented vascular endothelial cell death after 6 hrs of reperfusion (Danesh-Meyer et al., 2012). GFAP astrocyte immunoreactivity was (non-significantly) reduced and Cx43 expression was doubled in astrocytes and endothelial cells starting from 4 hrs.
4.2. Spinal cord injury
In vivo data are also available from spinal cord injury models. The pathophysiology of spinal cord injury involves the mechanotrauma proper, associated with direct cell injury, as well as ischemia related to vascular thrombosis or tissue swelling-induced vessel compression, leading to a secondary/delayed phase of cell death. In a rat spinal cord compression model, Cx43 downregulation by local application of antisense oligodeoxynucleotides (Cx43-asODN) immediately after injury improved the locomotion score from 24 hrs after injury up to 4 weeks later (Cronin et al., 2008). It also decreased cord tissue swelling and disruption, gave less astrocyte activation, less extravasation of bovine serum albumin and less infiltration of neutrophils. In another spinal cord transection model, the observations of less astrocyte activation and neutrophil recruitment upon Cx43-asODN treatment were confirmed, and it was further demonstrated that treatment decreased OX42+ microglial cell recruitment to the lesion site. Animals with astrocyte specific Cx43 knockout combined with global knockout of Cx30 (Cx30/Cx43 double knockout) displayed a consistently improved locomotor score starting from post-injury day 3 and lasting for at least 56 days (Huang et al., 2012). Electrophysiological recovery estimated from compound action potential amplitude was also better in the Cx30/Cx43 double knockout mice. The lesion volume was more than halved at 8 weeks post-injury in the double knockout mice and the myelin mass was 3.5 times larger in the double knockout compared to wild-type mice. These are striking observations as astrocyte-specific Cx43 knockout results in a larger infarct size in brain ischemia as discussed above ((Nakase et al., 2004) – though no data are available on Cx30/Cx43 double knockout mice for brain ischemia). Counts of reactive astrocytes (GFAP positive) and microglial cells were also decreased (Huang et al., 2012). Interestingly, post-injury ATP release measured in the tissue was decreased in the double knockout and the authors suggested a link between suppressed ATP release and protection against neuronal and oligodendrocyte cell death as well as decreased astrocyte and microglial activation. Hemichannel blockade with peptide5 had similar effects as decreasing Cx43 expression. In a spinal cord weight drop rat model, local application of peptide5 for 24 hrs (applied via an intrathecal catheter and an osmotic pump) improved locomotor recovery, prevented the Cx43 increase at 4 hrs and 8 hrs post-injury and restored its phosphorylation at S368 (O'Carroll et al., 2013). This treatment also decreased astrocyte activation (GFAP expression) adjacent to the lesion site and cell counts of activated microglia up to several millimeters away from the lesion site in gray and white matter. Neuron counts were also increased after treatment up to several mm away from the lesion (almost double counts of SMI32+ cells). Tumor necrosis factor-α and interleukin 1-β expression at the injury site was diminished by peptide5 treatment, suggesting a suppressive effect on inflammation and microglial activation via inhibition of ATP release or other mechanisms.
4.3. Conditioning
Ischemic preconditioning has been best characterized in the heart (see 7. Ischemia/reperfusion injury and protection in the heart) but also exerts protective effects in the brain (Gidday, 2006). Similar to the heart, the signaling cascade involves mitochondrial ATP-dependent potassium channels and K+ fluxes (Wu et al., 2011). However, in contrast to the heart where preconditioning is linked to mitochondrial Cx43, brain preconditioning has been linked to the opening of plasma membrane hemichannels, with subsequent ATP release and formation of extracellular adenosine (Lin et al., 2008). A transient MCAO (45 min) followed by reperfusion for 24 hrs had a reduced infarct volume when occlusion was preceded by ischemic preconditioning in wildtype mice (infarct volume more than halved) while there was no protection in Cx30/Cx43 double knockout animals (global knockout for Cx30, astrocyte specific knockout for Cx43).
In summary, although results using hemichannel inhibitors and genetic deletion of the hemichanel gene provided conflicting results, selective Cx43 hemichannel inhibition might be a promising target in neuroprotection.
5. Myocardial ischemia/reperfusion injury and cardioprotection
5.1. Irreversible injury
Cx43 forms hemichannels, which are predominantly closed in healthy myocardium (Goodenough et al, 2004; Krysko et al., 2005). However, they can open in response to electrical and chemical triggers, most notably during ischemia and inflammatory conditions (Saez et al., 2005; Saez & Leybaert, 2014; Wang et al., 2012a; Wang et al., 2013a; Wang et al., 2013b). In cardiomyocytes, metabolic inhibition activated a non-selective current through Cx43 hemichannels (Kondo et al., 2000). Single channel analysis of this current demonstrated a reversal potential of 0 mV and a unitary conductance of ~200 pS, which are biophysical properties compatible with Cx43 hemichannel activity (Wang et al., 2012a; Wang et al., 2013b). The 0 mV reversal potential suggests non-selectivity in terms of ion permeation. As a result, prolonged opening of Cx43 hemichannels during ischemia may lead to loss of ionic gradients, excessive Ca2+ entry, cell swelling and cellular damage. Cx43 hemichannels are also a well-defined release pathway for several metabolites such as ATP (Kang et al., 2008) and glutamate (Ye et al., 2003) to the extracellular space affecting homeostasis of neighboring cells (Tsukimoto et al., 2005).
Cardiomyocyte dye uptake following 1 hr simulated ischemia was blocked by Gap26 (Shintani-Ishida et al., 2007), and cardiomyocyte swelling caused by simulated ischemia/reoxygenation was blocked by Gap19 (Wang et al., 2013b). As a consequence, cardiomyocyte survival after simulated ischemia/reoxygenation was increased by Gap19 (Wang et al., 2013b) or Gap26 (Hawat et al., 2010). Also in mice hearts in vivo, infarct size after ischemia/reperfusion was attenuated by blockade of Cx43 hemichannels using Gap19 (Wang et al., 2013b). Gap26/Gap27 reduced myocardial infarct size after ischemia/reperfusion in rats (Hawat et al., 2010;Hawat et al., 2012;Johansen et al., 2011). The effect of Gap26/27 on infarct size appears to be larger than the one obtained with Gap19, which might be due to the fact that Gap26/27 is less selective towards gap junctions and hemichannels and also inhibits channels composed of Cx other than Cx43 (see 5. Pharmacological modulation of Cx43-formed channels).
Replacement of Cx43 by Cx32 in mice reduced infarct size following ischemia/reperfusion (Rodriguez-Sinovas et al., 2010) and also partial (50%) depletion of myocardial Cx43 attenuated infarct size in one (Kanno et al., 2003) but not in other studies (Heinzel et al., 2005;Schwanke et al., 2002). Thus, sustained ischemia/reperfusion opening of Cx43-formed hemichannels combined with residual gap junctional communication contributes to irreversible myocardial injury (the latter phenomenon has been termed “spreading of injury”, for a review, see (Garcia-Dorado et al., 2004)). Interestingly, in cultured cardiomyocytes, the rate of apoptosis increased rather than decreased when gap junctional communication was reduced (Yasui et al., 2000).
Many pharmacological interventions which reduce infarct size after ischemia/reperfusion such as blockade of p38 MAPK (Surinkaew et al., 2013), high density lipoprotein/sphingosine-1 phosphate (Morel et al., 2012), PKC epsilon activating peptide (Lancaster et al., 2011), FGF2/diazoxide (Srisakuldee et al., 2009) or estradiol (Lee et al., 2004) preserved Cx43 phosphorylation during ischemia/reperfusion, although causality between altered Cx43 phosphorylation and reduced infarct size was proven only in one study (Srisakuldee et al., 2009).
5.2. Conditioning
In cardiomyocytes, preconditioning by simulated ischemia/reoxygenation reduced Lucifer Yellow uptake prior to the sustained ischemic period but maintained Lucifer Yellow uptake following the prolonged lethal simulated ischemia (Sundset et al., 2007). Maintained Lucifer Yellow uptake followed by prolonged simulated ischemia was associated with preserved Cx43 phosphorylation. Cardiomyocytes isolated from heterozygous Cx43 knockout mice – in contrast to cardiomyocytes from wildtype mice - could not be conditioned by simulated ischemia/reoxygenation (Li et al., 2004).
Similar to the data in isolated cardiomyocytes, infarct size reduction by ischemic (Sanchez et al., 2013;Schwanke et al., 2002) or pharmacological (Heinzel et al., 2005) preconditioning was absent in Cx43 knockout mice or in mice in which Cx43 was replaced by Cx32 (Rodriguez-Sinovas et al., 2010). Also blockade of Cx43-formed channels using heptanol during the preconditioning cycles of ischemia/reperfusion blocked the infarct size reduction in mice hearts (Li et al., 2002). In many studies, ischemic or pharmacological preconditioning increased the extent of phosphorylated Cx43 during sustained ischemia (Hatanaka et al., 2004; Hund et al., 2007; Miura et al., 2010; Miura et al., 2007; Naitoh et al., 2006;Naitoh et al., 2009; Totzeck et al., 2008;Jain et al., 2003; Lee & Chou, 2003; Schulz et al., 2003) and subsequently reduced gap junction permeability (Lee&Chou, 2003; Miura et al., 2010; Miura et al., 2007; Naitoh et al., 2006; Naitoh et al., 2009) and increased lateralization of Cx43 (Vetterlein et al., 2006). Preserved phosphorylation was caused by activated PKC epsilon (Jain et al., 2003; Lee and Chou, 2003; Miura et al., 2010; Naitoh et al., 2009), and knockout of PKC abolished both the reduction in infarct size and the preservation of Cx43 phosphorylation by ischemic preconditioning in mice hearts (Hund et al., 2007). Apart from PKC, MAPK also contributed to Cx43 phosphorylation maintenance during sustained ischemia by ischemic or pharmacological preconditioning (Naitoh et al., 2006; Schulz et al., 2003). In some studies, however, the rate of Cx43 during sustained ischemia remained unaffected by ischemic preconditioning (Muhlfeld et al., 2010) and rate of gap junction uncoupling was decreased (Jain et al., 2003).
Importantly, although Cx43 deficient mice cannot be preconditioned by ischemic preconditioning, the RISK and SAFE pathways, which have been often associated with cardioprotection, became activated in these mice to a similar extent as in wildtype and conferred a protective effect (Sanchez et al., 2013). Indeed, the lack of cardioprotection by preconditioning in Cx43 deficient mice appears to be related to mitochondrial Cx43 and the insufficient mitochondrial production of reactive oxygen species (Heinzel et al., 2005).
Similarly to ischemic preconditioning, ischemic or pharmacological postconditioning reduced infarct size and preserved myocardial Cx43 expression and phosphorylation during sustained ischemia in rat hearts (Wu et al., 2012; Zhao et al., 2013). In contrast, to ischemic preconditioning, however, infarct size was reduced by ischemic postconditioning in heterozygous Cx43 knockout mice (Heusch et al., 2006) (for review, see (Schulz et al., 2007)).
Remote ischemic preconditioning (i.e. the preconditioning ischemia/reperfusion occurs in an organ distant from the heart) in rats preserved Cx43 protein expression and phosphorylation potentially contributing to the protection of the rat heart in vivo (Brandenburger et al., 2014).
In summary, blockade of Cx43 prior to ischemia and/or reperfusion reduces irreversible injury following ischemia/reperfusion. However, data obtained from ischemic conditioning studies suggest that Cx43 is essential also for cardioprotection. Thus, timing of blockade of Cx43 appears to be of utmost importance in order to maximize the cardioprotective effect.
6. Mitochondrial Cx 43
Apart from being present at the sarcolemma, Cx43 has been identified in mitochondria of cardiomyocytes (Boengler et al., 2005), astrocytes (Kozoriz et al., 2010a), endothelial cells (Kiec-Wilk et al., 2012; Mohammad & Kowluru, 2011;Trudeau et al., 2012), stem cells (Lu et al., 2012), hepatocytes (Vinken et al., 2013) and pancreatic cancer cells (Sun et al., 2012).
In cardiomyocytes, Cx43 is mainly located in the inner mitochondrial membrane of subsarcolemmal mitochondria (Figure 2; Boengler et al., 2009;Rodriguez-Sinovas et al., 2006), although one study reported Cx43 being present also in the outer mitochondrial membrane (Goubaeva et al., 2007). Mitochondrial Cx43 expression in the heart decreased with age (Boengler et al., 2007). A decline in mitochondrial Cx43 content was also seen after activation of the NMDA receptor 1 (NMDAR-1) in cardiomyocytes; NMDAR-1 activation increased translocation of matrix metalloproteinase 9 to mitochondria thereby inducing mitochondrial Cx43 degradation and mitophagy (Tyagi et al., 2010). [A similar effect was also seen in retinal endothelial cells (Mohammad & Kowluru, 2011; Trudeau et al., 2012)]. In contrast, ischemia/reperfusion (Boengler et al., 2005;Penna et al., 2009) or diazoxide (Yang et al., 2011) increased cardiac mitochondrial Cx43 content.
Mitochondrial Cx43 is involved in loss of cardiomyocyte division after birth; the signaling cascade involves reactive oxygen species formation and p38 MAPK activation (Matsuyama & Kawahara, 2011). Knockdown of mitochondrial Cx43 or inhibition of p38 MAPK restored the proliferation of neonatal rat cardiomyocytes (Matsuyama and Kawahara, 2009).
Mitochondrial connexin 43 impacts on respiratory function (Boengler et al., 2012;Ruiz-Meana et al., 2014), and the ischemia-associated reduction in mitochondrial respiration is associated with increased oxidized Cx43 cysteines. The latter effects are attenuated by ischemic preconditioning (Ruiz-Meana et al., 2014). Apart from its effect on respiration, mitochondrial Cx43 modulates the matrix potassium flux in heart (Boengler et al., 2013;Miro-Casas et al., 2009) and brain (Kozoriz et al., 2010a) mitochondria. S-nitrosation of mitochondrial Cx43 increases mitochondrial permeability, especially for potassium, and leads to increased reactive oxygen species formation. The increased amount of S-nitrosation mitochondrial Cx43 by ischemic preconditioning or nitrite administration may link nitric oxide and Cx43 in the signal transduction cascade of cardioprotective interventions (Soetkamp et al., 2014).
Regulation of the matrix calcium content also depends on mitochondrial Cx43; the calcium retention capacity of subsarcolemmal mitochondria was reduced by blocking Cx43-formed channels with Gap27 (Srisakuldee et al., 2014). In mitochondria derived from rat brain, carbenoxolone reduced the calcium retention capacity (Azarashvili et al., 2011). The modification of the calcium retention capacity is always accompanied by alterations in the phosphorylation status of mitochondrial Cx43 with the S262 epitope being of particular importance (Azarashvili et al., 2011;Srisakuldee et al., 2014).
Abolishing the mitochondrial import of Cx43 by blocking its interaction with heat shock protein 90 completely blocked the infarct size reduction by diazoxide in isolated rat hearts (Miro-Casas et al., 2009). In this study, sarcolemmal Cx43 remained unaffacted.
These data indicate that the presence of mitochondrial Cx43 is of utmost importance for cardioprotective pathway(s) being functional.
Table 2.
Importance of connexin 43 is ischemia/reperfusion injury and protection from it.
| Brain | Spinal cord | Heart | |||
|---|---|---|---|---|---|
| Ischemia | Conditioning | injury | Ischemia | Conditioning | |
| Cx43+/− | ↑ | ↓ or = | Protection lost |
||
| Cx43−/− | ↑ | Protection lost |
Function improved (Cx43/30 double KO) |
↓ (Cx32 KI) |
Protection lost (Cx32 KI) |
| Cx43K258 | ↑ | ||||
| Cx43 antisense | Function improved |
||||
| Pharmacological Blockade |
↓ | Protection lost |
|||
| Peptide5 | ↓ | ↓ Inflammation decreased |
|||
| Gap19 | ↓ | ||||
| Gap26/27 | ↓ | ||||
Legend to Table: Arrows illustrate effect on irreversible injury (↑ = increased; ↓ = decreased) KO: knockout; KI: knockin. For details see text.
7. Cx43 function in the presence of major cardiovascular risk factors
It has been well established that organoprotective cellular mechanisms are inhibited by the presence of major cardiovascular risk factors (see for reviews: Ferdinandy et al., 2007, 2014). Indeed, aging, hypertension, and metabolic diseases have been shown to interfere with Cx43 signalling.
7.1. Age and gender
In aged mice and rat hearts, total and mitochondrial Cx43 expression are decreased (Boengler et al., 2007; Fannin et al., 2014; Watanabe et al., 2004; Zaman et al., 2014). Also lateralization of Cx43 is enhanced in aged cardiomyocytes (Dhein and Hammerath, 2001). The increased heterogenety of Cx43 distribution correlates with age-associated alterations in heart rhythm and increased atrial fibrillation in patients (Nattel et al. 2007; Fannin et al., 2014).There are differences regarding gender as Cx43 expression is higher in female compared to male hearts (Knezl et al., 2008; Tribulova et al., 2005). Starvation in mice reduced the expression of Cx43 at the sarcolemma of cardiomyocytes (McLachlan et al., 2009).
7.2. Hypertension and cardiac hypertrophy
Most studies related to hypertension are done in spontaneously hypertensive rats. The results obtained differ in terms of Cx43 expression with Cx43 being up-regulated and phosphorylation of Cx43 being increased (Bacova et al., 2010; Bacova et al., 2012; Benova et al., 2013; Fialova et al., 2008; Mitasikova et al., 2008; Radosinska et al., 2011; Radosinska et al., 2013; Zhao et al., 2008) in some studies and Cx43 expression being decreased (Bacharova et al., 2008; Zhang et al., 2014) in others. Part of the observed differences might relate to the degree of left ventricular hypertrophy associated with hypertension, since in human hearts mild hypertrophy increased while extensive hypertrophy reduced left ventricular Cx43 expression (Kostin et al., 2004). Mild left ventricular hypertrophy due to pressure or volume overload was associated with changes in Cx43 sarcolemmal distribution in human hearts (Vetter et al., 2010). Similarly in mice with aortic banding, total Cx43 expression remained unaltered but Cx43 became redistributed (Boulaksil et al., 2010) or total Cx43 expression decreased (Qu et al., 2009; Yasuno et al., 2013) and Cx43 was dephosphorylated (Yasuno et al., 2013). Indeed, many interventions which reduce hypertension and/or hypertrophy (angiotensin II (AT) receptor 1-blockade(Zhao et al., 2008) or knockout (Yasuno et al., 2013), aldosterone antagonists (Yasuno et al., 2013), renin inhibition (Zhang et al., 2014) but also compounds such as melatonin (Benova et al., 2013), atorvastatin (Bacova et al., 2010; Chen et al., 2007), omega-3 free fatty acids (Bacova et al., 2012) or red palm oil (Mitasikova et al., 2008)) attenuate the observed changes in Cx43 expression and/or phosphorylation. Interestingly, blockade of AT- receptor 1 attenuated microRNA-1 expression (Curcio et al., 2013) which subsequently affected Cx43 expression: an increase in microRNA-1 secondary to p38MAPK activation in cardiomyocytes (Zhang et al., 2010b) or overexpression of microRNA-1 in mice depressed Cx43 expression (Zhang et al., 2010b) but a decrease in microRNA-1 increased Cx43 expression in female hearts (Stauffer et al., 2011).
7.3. Diabetes and hypercholesterolemia
Cx43 expression under diabetic conditions also shows variability from being increased (Anna et al., 2014; Howarth et al., 2008; Joshi et al., 2014), unaltered (Nygren et al., 2007) or even decreased (Lin et al., 2006a;Lin et al., 2006b; Sheu et al., 2007). In contrast, data on Cx43 phosphorylation and distribution are more homogenous: almost all studies show an increase in Cx43 phosphorylation (Anna et al., 2014; Howarth et al., 2008; Lin et al., 2006a; Lin et al., 2006b) [at PKC phosphorylation sites (Lin et al., 2006a; Lin et al., 2006b)] and a redistribution/lateralization of Cx43 in cardiomyocytes (Anna et al., 2014; Howarth et al., 2008; Joshi et al., 2014; Lin et al., 2006a; Lin et al., 2006b; Nygren et al., 2007), the latter being associated with an increase also in Cx43 nitrosation (Joshi et al., 2014).
In rabbits with hypercholesterolemia, JNK2 was activated which subsequently led to a reduction in Cx43 expression and redistribution of Cx43 away from gap junctions (Lin et al., 2005). Similarly in rats fed a high fat-diet, myocardial Cx43 expression was reduced and Cx43 was redistributed away from gap junctions and mitochondria in cardiomyocytes (Gorbe et al., 2011).
7.4. Post-myocardial infarction
There is a substantial amount of literature demonstrating a reduction and disorganization of myocardial Cx43 in the infarcted area as well as in the border zone. The decrease in myocardial Cx43 expression post-myocardial infarction in mice (Jackson et al., 2008; Lindsey et al., 2006) was related to increased NO availability since Cx43 expression was preserved in nitric oxide synthase 2 knockout mice despite a similar extent of infarction (Jackson et al., 2008). In cardiomyocytes, a nitric oxide donor dose-dependently decreased Cx43 expression over time (Jackson et al., 2008). Part of the degradation of Cx43 appears to be through matrix metalloproteinase 7 since the reduction in Cx43 expression post-myocardial infarction was not detected in matrix metalloproteinase 7 knockout mice (Lindsey et al., 2006). Also an increase in micro RNA-1 in the infarcted heart (Yang et al., 2007) might contribute to the decline in myocardial Cx43 expression (see above). Interestingly, the amount of fibrosis post-myocardial infarction is reduced in heterozygous Cx43 knockout mice through decreases in phosphoSMAD despite increased TGFß concentrations (Zhang et al., 2010a), again pointing to non-channel effects of Cx43.
7.5. Heart Failure
In failing animal hearts, total Cx43 expression (Ai & Pogwizd, 2005; Danielson et al., 2013; Givvimani et al., 2014; Wang et al., 2012b) and Cx43 expression at gap junctions is reduced (Hesketh et al., 2010; Petrich et al., 2004; Wang et al., 2012b; Zhong et al., 2007) independent of the cause of heart failure. Similarly, in tissue samples from failing human hearts (dilated or ischemic cardiomyopathy), Cx43 expression is reduced and Cx43 is redistributed away from the gap junctions (Dupont et al., 2001; Kostin et al., 2003). In some heart failure studies, Cx43 became dephosphorylated and dye transfer was reduced (Ai et al., 2011; Ai& Pogwizd, 2005) while in one study the phosphorylation of the S255 residue of Cx43 increased (Sato et al., 2008). Overexpression of Cx43 in failing hearts through adenovirus application restored cardiomyocyte coupling (Ai et al., 2010). The decrease in Cx43 expression is related to increased mitophagy and matrix metalloproteinase activation and can be blocked by inhibiting mitochondrial division (Givvimani et al., 2014), but also an increase in micro RNA (19a/b) contributes to loss of Cx43 (Danielson et al., 2013). In viral myocarditis, increases in micro RNA-1 repressed Cx43 expression in mice (Xu et al., 2012).
In conclusion, cardiovascular risk factors often are associated with a Cx43 deficient state especially in mitochondria thereby interfering with cardioprotection.
10. Conclusion and perspectives
Cx43 has multiple cellular locations and functions and changes in its expression and/or its channel and non-channel activities contribute to several cardiovascular pathologies (brain and heart irreversible injury, arrhythmias). More research is needed to understand the pathophysiological alterations in Cx43 and to develop potential selective pharmacological approaches to reverse these effects. Development of mimetic peptides (opening/closing Cx43-formed channels) which may differentiate between the different channel subtypes at the sarcolemma (hemichannels, gap junctions) or mitochondria or interfere with the non-channel function of Cx43 might be a promising approach to help treating cardiovascular pathologies in the future.
Figure 3. Cx43 in mitochondria (Cx43).

Cx43 is imported into the inner mitochondrial membrane of mitochondria in a heat shock protein (HSP) 90– dependent pathway. Its import into subsarcolemmal mitochindria in cardiomyocytes is increased by preconditioning interventions. Cx43 is S-nitrosylated and influences mitochondrial matrix potassium (K+) and calcium (Ca2+) influx and reactive oxygen species (ROS) formation. Miotchondrial Cx43 is important for cardioprotection through modification of ROS formation. Apart from its influence on ion fluxes, Cx43 affects mitochondrial respiratory complex function. Some studies indicate that Cx43 might also be localized at the outer mitochondrial membrane where it is important for cytochrome C (Cyt C) release. Finally, matrix metalloproteinases (MMP) affect the stability of mitochondrial Cx43. For details see text.
8. Acknowledgements
The National Research Fund of Hungary [Grants ANN 107803, K 109737) (to PF);
The European Foundation for the Study of Diabetes (to PF and RS)
The US National Institutes of Health [Grant GM55632 to PL]
The Hungarian Scientific Research Fund (OTKA PD 106001 (to GA), the Fund for Scientific Research Flanders, Belgium (Grant N° G.0298.11, G.0571.12, G.0A54.13 and G.0320.15N to LL) and the Interuniversity Attraction Poles Program (Belgian Science Policy, Project P7/10 to LL).
P.F. is a Szentágothai fellow (National Excellence Program of Hungary, TAMOP 4.2.4.A/2-11-1-2012-0001)
GA is a Bolyai János fellow of the Hungarian Academy of Sciences.
9. List of abbreviations
- 2-APB
2-aminoethoxydiphenyl borate
- asODN
antisense oligodeoxynucleotides
- ATP
adenosine trisphosphate
- BBB
blood-brain barrier
- Ca2+
calcium
- C-Src
tyrosine kinase cellular und sarcoma
- CK
casein kinase
- CL
cytoplasmic loop
- CREB
cyclic adenosine monophosphate responsive element binding protein
- CK1
casein kinase 1
- CT
carboxy-terminal tail
- Cx
connexin
- Cx43
connexin 43
- DIDS
disodium 4,4'-diisothiocyanatostilbene-2,2'-disulfonate
- EL
extracellular loop
- Gd3+
Gadolinium
- GFAB
Glial fibrillary acidic protein
- GSK3ß
glycogen synthase kinase 3ß
- Hr(s)
hour(s)
- IP3
inositol triphosphate
- JNK
c-Jun N-terminal kinase
- K+
potassium
- La3+
Lanthanum ions
- MAPK
mitogen activated protein kinase
- MCAO
mid cerebral artery occlusion
- Min
minutes
- MSX
Msh homebox
- Na+
sodium
- NeuN
neuronal nuclei
- NMDA
N-Methyl-d-aspartate
- NPPB
5-Nitro-2-(3-phenyl-propylamino)benzoic acid
- PLC
phospholipase C
- PKA
protein kinase A
- PKC
protein kinase C
- S
serine
- SPR
surface plasmon resonance
- TBX18
family member of T-box transcription factors
- ZO-1
zona occuldens proteins 1
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
12. Conflict of Interest Statement:
The authors declare there are not conflicts of interest for this paper.
14. List of References
- Abudara V, Bechberger J, Freitas-Andrade M, De Bock, M., Wang N, Bultynck G, et al. The connexin43 mimetic peptide Gap19 inhibits hemichannels without altering gap junctional communication in astrocytes. Front Cell Neurosci. 2014;8:306. doi: 10.3389/fncel.2014.00306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adam O, Lavall D, Theobald K, Hohl M, Grube M, Ameling S, et al. Rac1-induced connective tissue growth factor regulates connexin 43 and N-cadherin expression in atrial fibrillation. J Am Coll Cardiol. 2010;55:469–480. doi: 10.1016/j.jacc.2009.08.064. [DOI] [PubMed] [Google Scholar]
- Ai X, Jiang A, Ke Y, Solaro RJ, Pogwizd SM. Enhanced activation of p21-activated kinase 1 in heart failure contributes to dephosphorylation of connexin 43. Cardiovasc Res. 2011;92:106–114. doi: 10.1093/cvr/cvr163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ai X, Pogwizd SM. Connexin 43 downregulation and dephosphorylation in nonischemic heart failure is associated with enhanced colocalized protein phosphatase type 2A. Circ Res. 2005;96:54–63. doi: 10.1161/01.RES.0000152325.07495.5a. [DOI] [PubMed] [Google Scholar]
- Ai X, Zhao W, Pogwizd SM. Connexin43 knockdown or overexpression modulates cell coupling in control and failing rabbit left ventricular myocytes. Cardiovasc Res. 2010;85:751–762. doi: 10.1093/cvr/cvp353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anna Z, Angela S, Barbara B, Jana R, Tamara B, Csilla V, et al. Heart-protective effect of n-3 PUFA demonstrated in a rat model of diabetic cardiomyopathy. Mol Cell Biochem. 2014;389:219–227. doi: 10.1007/s11010-013-1943-9. [DOI] [PubMed] [Google Scholar]
- Anselmi F, Hernandez VH, Crispino G, Seydel A, Ortolano S, Roper SD, et al. ATP release through connexin hemichannels and gap junction transfer of second messengers propagate Ca2+ signals across the inner ear. Proc Natl Acad Sci U S A. 2008;105:18770–18775. doi: 10.1073/pnas.0800793105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Archard HO, Denys FR. Development of annular gap junctions in guinea pig epithelia. J Oral Pathol. 1979;8:187–197. doi: 10.1111/j.1600-0714.1979.tb01885.x. [DOI] [PubMed] [Google Scholar]
- Axelsen LN, Stahlhut M, Mohammed S, Larsen BD, Nielsen MS, Holstein-Rathlou NH, et al. Identification of ischemia-regulated phosphorylation sites in connexin43: A possible target for the antiarrhythmic peptide analogue rotigaptide (ZP123) J Mol Cell Cardiol. 2006;40:790–798. doi: 10.1016/j.yjmcc.2006.03.005. [DOI] [PubMed] [Google Scholar]
- Azarashvili T, Baburina Y, Grachev D, Krestinina O, Evtodienko Y, Stricker R, et al. Calcium-induced permeability transition in rat brain mitochondria is promoted by carbenoxolone through targeting connexin43. Am J Physiol Cell Physiol. 2011;300:C707–C720. doi: 10.1152/ajpcell.00061.2010. [DOI] [PubMed] [Google Scholar]
- Bacharova L, Plandorova J, Klimas J, Krenek P, Kyselovic J. Discrepancy between increased left ventricular mass and "normal" QRS voltage is associated with decreased connexin 43 expression in early stage of left ventricular hypertrophy in spontaneously hypertensive rats. J Electrocardiol. 2008;41:730–734. doi: 10.1016/j.jelectrocard.2008.01.013. [DOI] [PubMed] [Google Scholar]
- Bacova B, Radosinska J, Knezl V, Kolenova L, Weismann P, Navarova J, et al. Omega-3 fatty acids and atorvastatin suppress ventricular fibrillation inducibility in hypertriglyceridemic rat hearts: implication of intracellular coupling protein, connexin-43. J Physiol Pharmacol. 2010;61:717–723. [PubMed] [Google Scholar]
- Bacova B, Radosinska J, Viczenczova C, Knezl V, Dosenko V, Benova T, et al. Up-regulation of myocardial connexin-43 in spontaneously hypertensive rats fed red palm oil is most likely implicated in its anti-arrhythmic effects. Can J Physiol Pharmacol. 2012;90:1235–1245. doi: 10.1139/y2012-103. [DOI] [PubMed] [Google Scholar]
- Baklaushev VP, Gurina OI, Yusubalieva GM, Grinenko NF, Cytrin EB, Victorov IV, et al. Immunofluorescent analysis of connexin-43 using monoclonal antibodies to its extracellular domain. Bull Exp Biol Med. 2009;148:725–730. doi: 10.1007/s10517-010-0802-x. [DOI] [PubMed] [Google Scholar]
- Balda MS, Matter K. The tight junction protein ZO-1 and an interacting transcription factor regulate ErbB-2 expression. EMBO J. 2000;19:2024–2033. doi: 10.1093/emboj/19.9.2024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bargiotas P, Monyer H, Schwaninger M. Hemichannels in cerebral ischemia. Curr Mol Med. 2009;9:186–194. doi: 10.2174/156652409787581646. [DOI] [PubMed] [Google Scholar]
- Batra N, Riquelme MA, Burra S, Kar R, Gu S, Jiang JX. Direct regulation of osteocytic connexin 43 hemichannels through AKT kinase activated by mechanical stimulation. J Biol Chem. 2014;289:10582–10591. doi: 10.1074/jbc.M114.550608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beardslee M, Laing J, Beyer E, Saffitz J. Rapid turnover of connexin43 in the adult rat heart. Circ Res. 1998;83:629–635. doi: 10.1161/01.res.83.6.629. [DOI] [PubMed] [Google Scholar]
- Beardslee MA, Lerner DL, Tadros PN, Laing JG, Beyer EC, Yamada KA, et al. Dephosphorylation and intracellular redistribution of ventricular connexin43 during electrical uncoupling induced by ischemia. Circ Res. 2000;87:656–662. doi: 10.1161/01.res.87.8.656. [DOI] [PubMed] [Google Scholar]
- Belousov AB, Fontes JD. Neuronal gap junctions: making and breaking connections during development and injury. Trends Neurosci. 2013;36:227–236. doi: 10.1016/j.tins.2012.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benova T, Viczenczova C, Radosinska J, Bacova B, Knezl V, Dosenko V, et al. Melatonin attenuates hypertension-related proarrhythmic myocardial maladaptation of connexin-43 and propensity of the heart to lethal arrhythmias. Can J Physiol Pharmacol. 2013;91:633–639. doi: 10.1139/cjpp-2012-0393. [DOI] [PubMed] [Google Scholar]
- Bodendiek SB, Raman G. Connexin modulators and their potential targets under the magnifying glass. Curr Med Chem. 2010;17:4191–4230. doi: 10.2174/092986710793348563. [DOI] [PubMed] [Google Scholar]
- Boengler K, Dodoni G, Rodriguez-Sinovas A, Cabestrero A, Ruiz-Meana M, Gres P, et al. Connexin 43 in cardiomyocyte mitochondria and its increase by ischemic preconditioning. Cardiovasc Res. 2005;67:234–244. doi: 10.1016/j.cardiores.2005.04.014. [DOI] [PubMed] [Google Scholar]
- Boengler K, Konietzka I, Buechert A, Heinen Y, Garcia-Dorado D, Heusch G, et al. Loss of ischemic preconditioning's cardioprotection in aged mouse hearts is associated with reduced gap junctional and mitochondrial levels of connexin 43. Am J Physiol Heart Circ Physiol. 2007;292:H1764–H1769. doi: 10.1152/ajpheart.01071.2006. [DOI] [PubMed] [Google Scholar]
- Boengler K, Ruiz-Meana M, Gent S, Ungefug E, Soetkamp D, Miro-Casas E, et al. Mitochondrial connexin 43 impacts on respiratory complex I activity and mitochondrial oxygen consumption. J Cell Mol Med. 2012;16:1649–1655. doi: 10.1111/j.1582-4934.2011.01516.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boengler K, Stahlhofen S, van de Sand A., Gres P, Ruiz-Meana M, Garcia-Dorado D, et al. Presence of connexin 43 in subsarcolemmal, but not in interfibrillar cardiomyocyte mitochondria. Basic Res Cardiol. 2009;104:141–147. doi: 10.1007/s00395-009-0007-5. [DOI] [PubMed] [Google Scholar]
- Boengler K, Ungefug E, Heusch G, Leybaert L, Schulz R. Connexin 43 impacts on mitochondrial potassium uptake. Front Pharmacol. 2013;4:73. doi: 10.3389/fphar.2013.00073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boogerd KJ, Wong LY, Christoffels VM, Klarenbeek M, Ruijter JM, Moorman AF, et al. Msx1 and Msx2 are functional interacting partners of T-box factors in the regulation of Connexin43. Cardiovasc Res. 2008;78:485–493. doi: 10.1093/cvr/cvn049. [DOI] [PubMed] [Google Scholar]
- Bootman MD, Collins TJ, Mackenzie L, Roderick HL, Berridge MJ, Peppiatt CM. 2-aminoethoxydiphenyl borate (2-APB) is a reliable blocker of store-operated Ca2+ entry but an inconsistent inhibitor of InsP3-induced Ca2+ release. FASEB J. 2002;16:1145–1150. doi: 10.1096/fj.02-0037rev. [DOI] [PubMed] [Google Scholar]
- Boulaksil M, Winckels SK, Engelen MA, Stein M, van Veen T. A., Jansen JA, et al. Heterogeneous Connexin43 distribution in heart failure is associated with dispersed conduction and enhanced susceptibility to ventricular arrhythmias. Eur J Heart Fail. 2010;12:913–921. doi: 10.1093/eurjhf/hfq092. [DOI] [PubMed] [Google Scholar]
- Braet K, Vandamme W, Martin PE, Evans WH, Leybaert L. Photoliberating inositol-1,4,5-trisphosphate triggers ATP release that is blocked by the connexin mimetic peptide gap 26. Cell Calcium. 2003;33:37–48. doi: 10.1016/s0143-4160(02)00180-x. [DOI] [PubMed] [Google Scholar]
- Brandenburger T, Huhn R, Galas A, Pannen BH, Keitel V, Barthel F, et al. Remote ischemic preconditioning preserves Connexin 43 phosphorylation in the rat heart in vivo. J Transl Med. 2014;2712:228. doi: 10.1186/s12967-014-0228-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruzzone R, Barbe MT, Jakob NJ, Monyer H. Pharmacological properties of homomeric and heteromeric pannexin hemichannels expressed in Xenopus oocytes. J Neurochem. 2005;92:1033–1043. doi: 10.1111/j.1471-4159.2004.02947.x. [DOI] [PubMed] [Google Scholar]
- Cagan RL, Kramer H, Hart AC, Zipursky SL. The bride of sevenless and sevenless interaction: internalization of a transmembrane ligand. Cell. 1992;69:393–399. doi: 10.1016/0092-8674(92)90442-f. [DOI] [PubMed] [Google Scholar]
- Cascio WE, Yang H, Muller-Borer BJ, Johnson TA. Ischemia-induced arrhythmia: the role of connexins, gap junctions, and attendant changes in impulse propagation. J Electrocardiol. 2005;38:55–59. doi: 10.1016/j.jelectrocard.2005.06.019. [DOI] [PubMed] [Google Scholar]
- Chaytor AT, Martin PE, Edwards DH, Griffith TM. Gap junctional communication underpins EDHF-type relaxations evoked by ACh in the rat hepatic artery. Am J Physiol Heart Circ Physiol. 2001;280:H2441–2450. doi: 10.1152/ajpheart.2001.280.6.H2441. [DOI] [PubMed] [Google Scholar]
- Chen HJ, Yao L, Chen TG, Yu M, Wang LH, Chen JZ. Atorvastatin prevents connexin43 remodeling in hypertrophied left ventricular myocardium of spontaneously hypertensive rats. Chin Med J (Engl ) 2007;120:1902–1907. [PubMed] [Google Scholar]
- Chen SC, Kennedy BK, Lampe PD. Phosphorylation of connexin43 on S279/282 may contribute to laminopathy-associated conduction defects. Exp Cell Res. 2013;319:888–896. doi: 10.1016/j.yexcr.2012.12.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheung G, Chever O, Rouach N. Connexons and pannexons: newcomers in neurophysiology. Front Cell Neurosci. 2014;8:348. doi: 10.3389/fncel.2014.00348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chkourko HS, Guerrero-Serna G, Lin X, Darwish N, Pohlmann JR, Cook KE, et al. Remodeling of mechanical junctions and of microtubule-associated proteins accompany cardiac connexin43 lateralization. Heart Rhythm. 2012;9:1133–1140. doi: 10.1016/j.hrthm.2012.03.003. e1136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chowdhury D, Marco S, Brooks IM, Zandueta A, Rao Y, Haucke V, et al. Tyrosine phosphorylation regulates the endocytosis and surface expression of GluN3A-containing NMDA receptors. J Neurosci. 2013;33:4151–4164. doi: 10.1523/JNEUROSCI.2721-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung TH, Wang SM, Chang YC, Chen YL, Wu JC. 18beta-glycyrrhetinic acid promotes src interaction with connexin43 in rat cardiomyocytes. J Cell Biochem. 2007;100:653–664. doi: 10.1002/jcb.21018. [DOI] [PubMed] [Google Scholar]
- Chung TH, Wang SM, Liang JY, Yang SH, Wu JC. The interaction of estrogen receptor alpha and caveolin-3 regulates connexin43 phosphorylation in metabolic inhibition-treated rat cardiomyocytes. Int J Biochem Cell Biol. 2009;41:2323–2333. doi: 10.1016/j.biocel.2009.06.001. [DOI] [PubMed] [Google Scholar]
- Chung TH, Wang SM, Wu JC. 17beta-estradiol reduces the effect of metabolic inhibition on gap junction intercellular communication in rat cardiomyocytes via the estrogen receptor. J Mol Cell Cardiol. 2004;37:1013–1022. doi: 10.1016/j.yjmcc.2004.08.003. [DOI] [PubMed] [Google Scholar]
- Clair C, Combettes L, Pierre F, Sansonetti P, Tran Van Nhieu, G. Extracellular-loop peptide antibodies reveal a predominant hemichannel organization of connexins in polarized intestinal cells. Exp Cell Res. 2008;314:1250–1265. doi: 10.1016/j.yexcr.2007.12.021. [DOI] [PubMed] [Google Scholar]
- Clark RAF. Cutaneous tissue repair: basic biological considerations. J Am Acad Derm. 1985;13:701–725. doi: 10.1016/s0190-9622(85)70213-7. [DOI] [PubMed] [Google Scholar]
- Contreras JE, Sanchez HA, Eugenin EA, Speidel D, Theis M, Willecke K, et al. Metabolic inhibition induces opening of unapposed connexin 43 gap junction hemichannels and reduces gap junctional communication in cortical astrocytes in culture. Proc Natl Acad Sci U S A. 2002;99:495–500. doi: 10.1073/pnas.012589799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Contreras JE, Sanchez HA, Veliz LP, Bukauskas FF, Bennett MV, Saez JC. Role of connexin-based gap junction channels and hemichannels in ischemia-induced cell death in nervous tissue. Brain Res Brain Res Rev. 2004;47:290–303. doi: 10.1016/j.brainresrev.2004.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper CD, Lampe PD. Casein kinase 1 regulates connexin43 gap junction assembly. J Biol Chem. 2002;277:44962–44968. doi: 10.1074/jbc.M209427200. [DOI] [PubMed] [Google Scholar]
- Cotrina ML, Kang J, Lin JH, Bueno E, Hansen TW, He L, et al. Astrocytic gap junctions remain open during ischemic conditions. J Neurosci. 1998;18:2520–2537. doi: 10.1523/JNEUROSCI.18-07-02520.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cronin M, Anderson PN, Cook JE, Green CR, Becker DL. Blocking connexin43 expression reduces inflammation and improves functional recovery after spinal cord injury. Mol Cell Neurosci. 2008;39:152–160. doi: 10.1016/j.mcn.2008.06.005. [DOI] [PubMed] [Google Scholar]
- Crow DS, Beyer EC, Paul DL, Kobe SS, Lau AF. Phosphorylation of connexin43 gap junction protein in uninfected and Rous sarcoma virus-transformed mammalian fibroblasts. Mol Cell Biol. 1990;10:1754–1763. doi: 10.1128/mcb.10.4.1754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Curcio A, Torella D, Iaconetti C, Pasceri E, Sabatino J, Sorrentino S, et al. MicroRNA-1 downregulation increases connexin 43 displacement and induces ventricular tachyarrhythmias in rodent hypertrophic hearts. PLoS One. 2013.;8:e70158. doi: 10.1371/journal.pone.0070158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D'Hondt C, Iyyathurai J, Himpens B, Leybaert L, Bultynck G. Cx43-hemichannel function and regulation in physiology and pathophysiology: insights from the bovine corneal endothelial cell system and beyond. Front Physiol. 2014;5:348. doi: 10.3389/fphys.2014.00348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D'Hondt C, Iyyathurai J, Wang N, Gourdie RG, Himpens B, Leybaert L, et al. Negatively charged residues (Asp378 and Asp379) in the last ten amino acids of the C-terminal tail of Cx43 hemichannels are essential for loop/tail interactions. Biochem Biophys Res Commun. 2013;432:707–712. doi: 10.1016/j.bbrc.2013.01.066. [DOI] [PubMed] [Google Scholar]
- Danesh-Meyer HV, Kerr NM, Zhang J, Eady EK, O'Carroll SJ, Nicholson LF, et al. Connexin43 mimetic peptide reduces vascular leak and retinal ganglion cell death following retinal ischaemia. Brain. 2012;135:506–520. doi: 10.1093/brain/awr338. [DOI] [PubMed] [Google Scholar]
- Danielson LS, Park DS, Rotllan N, Chamorro-Jorganes A, Guijarro MV, Fernandez-Hernando C, et al. Cardiovascular dysregulation of miR-17-92 causes a lethal hypertrophic cardiomyopathy and arrhythmogenesis. FASEB J. 2013;27:1460–1467. doi: 10.1096/fj.12-221994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dang X, Doble BW, Kardami E. The carboxy-tail of connexin-43 localizes to the nucleus and inhibits cell growth. Mol Cell Biochem. 2003;242:35–38. [PubMed] [Google Scholar]
- Danon A, Zeevi-Levin N, Pinkovich DY, Michaeli T, Berkovich A, Flugelman M, et al. Hypoxia causes connexin 43 internalization in neonatal rat ventricular myocytes. Gen Physiol Biophys. 2010;29:222–233. doi: 10.4149/gpb_2010_03_222. [DOI] [PubMed] [Google Scholar]
- Darrow BJ, Laing JG, Lampe PD, Saffitz JE, Beyer EC. Expression of multiple connexins in cultured neonatal rat ventricular myocytes. Circ. Res. 1995;76:381–387. doi: 10.1161/01.res.76.3.381. [DOI] [PubMed] [Google Scholar]
- Davidson JO, Green CR, Bennet L, Gunn AJ. Battle of the hemichannels - Connexins and Pannexins in ischemic brain injury. Int J Dev Neurosci. 2014 doi: 10.1016/j.ijdevneu.2014.12.007. Dec 26. pii: S0736-5748(14)00209-3. doi: 10.1016/j.ijdevneu.2014.12.007. [DOI] [PubMed] [Google Scholar]
- Davidson JO, Green CR, Bennet L, Nicholson LF, Danesh-Meyer H, O'Carroll SJ, et al. A key role for connexin hemichannels in spreading ischemic brain injury. Curr Drug Targets. 2013;14:36–46. doi: 10.2174/138945013804806479. [DOI] [PubMed] [Google Scholar]
- Davidson JO, Green CR, Nicholson LF, Bennet L, Gunn AJ. Connexin hemichannel blockade is neuroprotective after, but not during, global cerebral ischemia in near-term fetal sheep. Exp Neurol. 2013;248:301–308. doi: 10.1016/j.expneurol.2013.06.026. [DOI] [PubMed] [Google Scholar]
- Davidson JO, Green CR, Nicholson LF, O'Carroll SJ, Fraser M, Bennet L, et al. Connexin hemichannel blockade improves outcomes in a model of fetal ischemia. Ann Neurol. 2012;71:121–132. doi: 10.1002/ana.22654. [DOI] [PubMed] [Google Scholar]
- De Bock, M., Culot M, Wang N, Bol M, Decrock E, De Vuyst E, et al. Connexin channels provide a target to manipulate brain endothelial calcium dynamics and blood-brain barrier permeability. J Cereb Blood Flow Metab. 2011;31:1942–1957. doi: 10.1038/jcbfm.2011.86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Bock, M., Decrock E, Wang N, Bol M, Vinken M, Bultynck G, et al. The dual face of connexin-based astroglial Ca(2+) communication: a key player in brain physiology and a prime target in pathology. Biochim Biophys Acta. 2014;1843:2211–2232. doi: 10.1016/j.bbamcr.2014.04.016. [DOI] [PubMed] [Google Scholar]
- De Bock M, Wang N, Bol M, Decrock E, Ponsaerts R, Bultynck G, et al. Connexin 43 hemichannels contribute to cytoplasmic Ca2+ oscillations by providing a bimodal Ca2+-dependent Ca2+ entry pathway. J Biol Chem. 2012;287:12250–12266. doi: 10.1074/jbc.M111.299610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Bock M, Wang N, Decrock E, Bol M, Gadicherla AK, Culot M, et al. Endothelial calcium dynamics, connexin channels and blood-brain barrier function. Prog Neurobiol. 2013;108:1–20. doi: 10.1016/j.pneurobio.2013.06.001. [DOI] [PubMed] [Google Scholar]
- de Pina-Benabou MH, Szostak V, Kyrozis A, Rempe D, Uziel D, Urban-Maldonado M, et al. Blockade of gap junctions in vivo provides neuroprotection after perinatal global ischemia. Stroke. 2005;36:2232–2237. doi: 10.1161/01.STR.0000182239.75969.d8. [DOI] [PubMed] [Google Scholar]
- De Vuyst E, Boengler K, Antoons G, Sipido KR, Schulz R, Leybaert L. Pharmacological modulation of connexin-formed channels in cardiac pathophysiology. Br J Pharmacol. 2011;163:469–483. doi: 10.1111/j.1476-5381.2011.01244.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Vuyst E, Decrock E, De Bock M, Yamasaki H, Naus CC, Evans WH, et al. Connexin hemichannels and gap junction channels are differentially influenced by lipopolysaccharide and basic fibroblast growth factor. Mol Biol Cell. 2007;18:34–46. doi: 10.1091/mbc.E06-03-0182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Vuyst E, Wang N, Decrock E, De Bock M, Vinken M, Van Moorhem M, et al. Ca(2+) regulation of connexin 43 hemichannels in C6 glioma and glial cells. Cell Calcium. 2009;46:176–187. doi: 10.1016/j.ceca.2009.07.002. [DOI] [PubMed] [Google Scholar]
- Decrock E, De Vuyst E, Vinken M, Van Moorhem M, Vranckx K, Wang N, et al. Connexin 43 hemichannels contribute to the propagation of apoptotic cell death in a rat C6 glioma cell model. Cell Death Differ. 2009;16:151–163. doi: 10.1038/cdd.2008.138. [DOI] [PubMed] [Google Scholar]
- Decrock E, Vinken M, De Vuyst E, Krysko DV, D'Herde K, Vanhaecke T, et al. Connexin-related signaling in cell death: to live or let die? Cell Death Differ. 2009;16:524–536. doi: 10.1038/cdd.2008.196. [DOI] [PubMed] [Google Scholar]
- Dekker LR, Fiolet JW, VanBavel E, Coronel R, Opthof T, Spaan JA, et al. Intracellular Ca2+, intercellular electrical coupling, and mechanical activity in ischemic rabbit papillary muscle. Effects of preconditioning and metabolic blockade. Circ Res. 1996;79:237–246. doi: 10.1161/01.res.79.2.237. [DOI] [PubMed] [Google Scholar]
- Delmar M, Coombs W, Sorgen P, Duffy HS, Taffet SM. Structural bases for the chemical regulation of connexin43 channels. Cardiovasc Res. 2004;62:268–275. doi: 10.1016/j.cardiores.2003.12.030. [DOI] [PubMed] [Google Scholar]
- Desplantez T, Dupont E, Severs NJ, Weingart R. Gap junction channels and cardiac impulse propagation. J Membr Biol. 2007;218:13–28. doi: 10.1007/s00232-007-9046-8. [DOI] [PubMed] [Google Scholar]
- Dhein S, Hammerath SB. Aspects of the intercellular communication in aged hearts: effects of the gap junction uncoupler palmitoleic acid. Naunyn Schmiedebergs Arch Pharmacol. 2001;364:397–408. doi: 10.1007/s002100100462. [DOI] [PubMed] [Google Scholar]
- Dhein S. Cardiac ischemia and uncoupling: gap junctions in ischemia and infarction. Adv Cardiol. 2006;42:198–212. doi: 10.1159/000092570. [DOI] [PubMed] [Google Scholar]
- Dhein S, Seidel T, Salameh A, Jozwiak J, Hagen A, Kostelka M, Hindricks G, Mohr FW. Remodeling of cardiac passive electrical properties and susceptibility to ventricular and atrial arrhythmias. Front Physiol. 2014;5:424. doi: 10.3389/fphys.2014.00424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duffy HS, Ashton AW, O'Donnell P, Coombs W, Taffet SM, Delmar M, et al. Regulation of connexin43 protein complexes by intracellular acidification. Circ Res. 2004;94:215–222. doi: 10.1161/01.RES.0000113924.06926.11. [DOI] [PubMed] [Google Scholar]
- Dodge SM, Beardslee MA, Darrow BJ, Green KG, Beyer EC, Saffitz JE. Effects of angiotensin II on expression of the gap junction channel protein connexin43 in neonatal rat ventricular myocytes. J Am Coll Cardiol. 1998;32:800–807. doi: 10.1016/s0735-1097(98)00317-9. [DOI] [PubMed] [Google Scholar]
- Dunham B, Liu S, Taffet S, Trabka-Janik E, Delmar M, Petryshyn R, et al. Immunolocalization and expression of functional and nonfunctional cell-to-cell channels from wild-type and mutant rat heart connexin43 cDNA. Circ Res. 1992;70:1233–1243. doi: 10.1161/01.res.70.6.1233. [DOI] [PubMed] [Google Scholar]
- Dunn CA, Lampe PD. Injury-triggered Akt phosphorylation of Cx43: a ZO-1-driven molecular switch that regulates gap junction size. J Cell Sci. 2014;127:455–464. doi: 10.1242/jcs.142497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunn CA, Su V, Lau AF, Lampe PD. Activation of Akt, not connexin 43 protein ubiquitination, regulates gap junction stability. J Biol Chem. 2012;287:2600–2607. doi: 10.1074/jbc.M111.276261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dupont E, Matsushita T, Kaba RA, Vozzi C, Coppen SR, Khan N, et al. Altered connexin expression in human congestive heart failure. J Mol Cell Cardiol. 2001;33:359–371. doi: 10.1006/jmcc.2000.1308. [DOI] [PubMed] [Google Scholar]
- Dyce PW, Norris RP, Lampe PD, Kidder GM. Phosphorylation of serine residues in the C-terminal cytoplasmic tail of connexin43 regulates proliferation of ovarian granulosa cells. J Membr Biol. 2012;245:291–301. doi: 10.1007/s00232-012-9450-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ek-Vitorin JF, Calero G, Morley GE, Coombs W, Taffet SM, Delmar M. PH regulation of connexin43: molecular analysis of the gating particle. Biophys J. 1996;71:1273–1284. doi: 10.1016/S0006-3495(96)79328-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ek-Vitorin JF, King TJ, Heyman NS, Lampe PD, Burt JM. Selectivity of connexin 43 channels is regulated through protein kinase C-dependent phosphorylation. Circ Res. 2006;98:1498–1505. doi: 10.1161/01.RES.0000227572.45891.2c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ek JF, Delmar M, Perzova R, Taffet SM. Role of histidine 95 on pH gating of the cardiac gap junction protein connexin43. Circ Res. 1994;74:1058–1064. doi: 10.1161/01.res.74.6.1058. [DOI] [PubMed] [Google Scholar]
- Eugenin EA, Basilio D, Saez JC, Orellana JA, Raine CS, Bukauskas F, et al. The role of gap junction channels during physiologic and pathologic conditions of the human central nervous system. J Neuroimmune Pharmacol. 2012;7:499–518. doi: 10.1007/s11481-012-9352-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans WH, Boitano S. Connexin mimetic peptides: specific inhibitors of gap-junctional intercellular communication. Biochem Soc Trans. 2001;29:606–612. doi: 10.1042/bst0290606. [DOI] [PubMed] [Google Scholar]
- Fannin J, Rice KM, Thulluri S, Dornon L, Arvapalli RK, Wehner P, et al. Age-associated alterations of cardiac structure and function in the female F344xBN rat heart. Age (Dordr ) 2014;36:9684. doi: 10.1007/s11357-014-9684-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fialova M, Dlugosova K, Okruhlicova L, Kristek F, Manoach M, Tribulova N. Adaptation of the heart to hypertension is associated with maladaptive gap junction connexin-43 remodeling. Physiol Res. 2008;57:7–11. doi: 10.33549/physiolres.931101. [DOI] [PubMed] [Google Scholar]
- Fishman GI, Moreno AP, Spray DC, Leinwand LA. Functional analysis of human cardiac gap junction channel mutants. Proc Natl Acad Sci. 1991;88:3525–3529. doi: 10.1073/pnas.88.9.3525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fong JT, Kells RM, Gumpert AM, Marzillier JY, Davidson MW, Falk MM. Internalized gap junctions are degraded by autophagy. Autophagy. 2012;8:794–811. doi: 10.4161/auto.19390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia-Dorado D, Rodriguez-Sinovas A, Ruiz-Meana M. Gap junction-mediated spread of cell injury and death during myocardial ischemia-reperfusion. Cardiovasc Res. 2004;61:386–401. doi: 10.1016/j.cardiores.2003.11.039. [DOI] [PubMed] [Google Scholar]
- Giaume C, Koulakoff A, Roux L, Holcman D, Rouach N. Astroglial networks: a step further in neuroglial and gliovascular interactions. Nat Rev Neurosci. 2010;11:87–99. doi: 10.1038/nrn2757. [DOI] [PubMed] [Google Scholar]
- Giaume C, Leybaert L, Naus CC, Saez JC. Connexin and pannexin hemichannels in brain glial cells: properties, pharmacology, and roles. Front Pharmacol. 2013;4:88. doi: 10.3389/fphar.2013.00088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giaume C, Theis M. Pharmacological and genetic approaches to study connexin-mediated channels in glial cells of the central nervous system. Brain Res Rev. 2010;63:160–176. doi: 10.1016/j.brainresrev.2009.11.005. [DOI] [PubMed] [Google Scholar]
- Gidday JM. Cerebral preconditioning and ischaemic tolerance. Nat Rev Neurosci. 2006;7:437–448. doi: 10.1038/nrn1927. [DOI] [PubMed] [Google Scholar]
- Giepmans BN. Gap junctions and connexin-interacting proteins. Cardiovasc Res. 2004;62:233–245. doi: 10.1016/j.cardiores.2003.12.009. [DOI] [PubMed] [Google Scholar]
- Gilleron J, Fiorini C, Carette D, Avondet C, Falk MM, Segretain D, et al. Molecular reorganization of Cx43, Zo-1 and Src complexes during the endocytosis of gap junction plaques in response to a non-genomic carcinogen. J Cell Sci. 2008;121:4069–4078. doi: 10.1242/jcs.033373. [DOI] [PubMed] [Google Scholar]
- Givvimani S, Pushpakumar S, Veeranki S, Tyagi SC. Dysregulation of Mfn2 and Drp-1 proteins in heart failure. Can J Physiol Pharmacol. 2014;92:583–591. doi: 10.1139/cjpp-2014-0060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg GS, Moreno AP, Bechberger JF, Hearn SS, Shivers RR, MacPhee DJ, et al. Evidence that disruption of connexon particle arrangements in gap junction plaques is associated with inhibition of gap junctional communication by a glycyrrhetinic acid derivative. Exp Cell Res. 1996;222:48–53. doi: 10.1006/excr.1996.0006. [DOI] [PubMed] [Google Scholar]
- Goodenough DA, Paul DL. Beyond the gap: functions of unpaired connexon channels. Nat Rev Mol Cell Biol. 2003;4:285–294. doi: 10.1038/nrm1072. [DOI] [PubMed] [Google Scholar]
- Gorbe A, Varga ZV, Kupai K, Bencsik P, Kocsis GF, Csont T, et al. Cholesterol diet leads to attenuation of ischemic preconditioning-induced cardiac protection: the role of connexin 43. Am J Physiol Heart Circ Physiol. 2011;300:H1907–H1913. doi: 10.1152/ajpheart.01242.2010. [DOI] [PubMed] [Google Scholar]
- Goubaeva F, Mikami M, Giardina S, Ding B, Abe J, Yang J. Cardiac mitochondrial connexin 43 regulates apoptosis. Biochem Biophys Res Commun. 2007;352:97–103. doi: 10.1016/j.bbrc.2006.10.177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hatanaka K, Kawata H, Toyofuku T, Yoshida K. Down-regulation of connexin43 in early myocardial ischemia and protective effect by ischemic preconditioning in rat hearts in vivo. Jpn Heart J. 2004;45:1007–1019. doi: 10.1536/jhj.45.1007. [DOI] [PubMed] [Google Scholar]
- Hawat G, Benderdour M, Rousseau G, Baroudi G. Connexin 43 mimetic peptide Gap26 confers protection to intact heart against myocardial ischemia injury. Pflugers Arch. 2010;460:583–592. doi: 10.1007/s00424-010-0849-6. [DOI] [PubMed] [Google Scholar]
- Hawat G, Helie P, Baroudi G. Single intravenous low-dose injections of connexin 43 mimetic peptides protect ischemic heart in vivo against myocardial infarction. J Mol Cell Cardiol. 2012;53:559–566. doi: 10.1016/j.yjmcc.2012.07.008. [DOI] [PubMed] [Google Scholar]
- Heinzel FR, Luo Y, Li X, Boengler K, Buechert A, Garcia-Dorado D, et al. Impairment of diazoxide-induced formation of reactive oxygen species and loss of cardioprotection in connexin 43 deficient mice. Circ Res. 2005;97:583–586. doi: 10.1161/01.RES.0000181171.65293.65. [DOI] [PubMed] [Google Scholar]
- Herrero-Gonzalez S, Gangoso E, Giaume C, Naus CC, Medina JM, et al. Connexin43 inhibits the oncogenic activity of c-Src in C6 glioma cells. Oncogene. 2010;29:5712–5723. doi: 10.1038/onc.2010.299. [DOI] [PubMed] [Google Scholar]
- Hertlein B, Butterweck A, Haubrich S, Willecke K, Traub O. Phosphorylated carboxy terminal serine residues stabilize the mouse gap junction protein connexin45 against degradation. J. Membrane Biol. 1998;162:247–257. doi: 10.1007/s002329900362. [DOI] [PubMed] [Google Scholar]
- Hesketh GG, Shah MH, Halperin VL, Cooke CA, Akar FG, Yen TE, et al. Ultrastructure and regulation of lateralized connexin43 in the failing heart. Circ Res. 2010;106:1153–1163. doi: 10.1161/CIRCRESAHA.108.182147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heusch G, Buchert A, Feldhaus S, Schulz R. No loss of cardioprotection by postconditioning in connexin 43-deficient mice. Basic Res Cardiol. 2006;101:354–356. doi: 10.1007/s00395-006-0589-0. [DOI] [PubMed] [Google Scholar]
- Hofer A, Dermietzel R. Visualization and functional blocking of gap junction hemichannels (connexons) with antibodies against external loop domains in astrocytes. Glia. 1998;24:141–154. doi: 10.1002/(sici)1098-1136(199809)24:1<141::aid-glia13>3.0.co;2-r. [DOI] [PubMed] [Google Scholar]
- Howarth FC, Chandler NJ, Kharche S, Tellez JO, Greener ID, Yamanushi TT, et al. Effects of streptozotocin-induced diabetes on connexin43 mRNA and protein expression in ventricular muscle. Mol Cell Biochem. 2008;319:105–114. doi: 10.1007/s11010-008-9883-5. [DOI] [PubMed] [Google Scholar]
- Huang C, Han X, Li X, Lam E, Peng W, Lou N, et al. Critical role of connexin 43 in secondary expansion of traumatic spinal cord injury. J Neurosci. 2012;32:3333–3338. doi: 10.1523/JNEUROSCI.1216-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang GY, Xie LJ, Linask KL, Zhang C, Zhao XQ, Yang Y, et al. Evaluating the role of connexin43 in congenital heart disease: Screening for mutations in patients with outflow tract anomalies and the analysis of knock-in mouse models. J Cardiovasc Dis Res. 2011;2:206–212. doi: 10.4103/0975-3583.89804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hund TJ, Lerner DL, Yamada KA, Schuessler RB, Saffitz JE. Protein kinase Cepsilon mediates salutary effects on electrical coupling induced by ischemic preconditioning. Heart Rhythm. 2007;4:1183–1193. doi: 10.1016/j.hrthm.2007.05.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunter AW, Barker RJ, Zhu C, Gourdie RG. Zonula occludens-1 alters connexin43 gap junction size and organization by influencing channel accretion. Mol Biol Cell. 2005;16:5686–5698. doi: 10.1091/mbc.E05-08-0737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Incardona JP, Lee JH, Robertson CP, Enga K, Kapur RP, et al. Receptor-mediated endocytosis of soluble and membrane-tethered Sonic hedgehog by Patched-1. Proc Natl Acad Sci U S A. 2000;97:12044–12049. doi: 10.1073/pnas.220251997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iravanian S, Sovari AA, Lardin HA, Liu H, Xiao HD, Dolmatova E, et al. Inhibition of renin-angiotensin system (RAS) reduces ventricular tachycardia risk by altering connexin43. J Mol Med (Berl) 2011;89:677–687. doi: 10.1007/s00109-011-0761-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iyyathurai J, D'Hondt C, Wang N, De Bock M, Himpens B, Retamal MA, et al. Peptides and peptide-derived molecules targeting the intracellular domains of Cx43: gap junctions versus hemichannels. Neuropharmacology. 2013;75:491–505. doi: 10.1016/j.neuropharm.2013.04.050. [DOI] [PubMed] [Google Scholar]
- Jackson PE, Feng QP, Jones DL. Nitric oxide depresses connexin 43 after myocardial infarction in mice. Acta Physiol (Oxf) 2008;19:23–33. doi: 10.1111/j.1748-1716.2008.01858.x. [DOI] [PubMed] [Google Scholar]
- Jain SK, Schuessler RB, Saffitz JE. Mechanisms of delayed electrical uncoupling induced by ischemic preconditioning. Circ Res. 2003;92:1138–1144. doi: 10.1161/01.RES.0000074883.66422.C5. [DOI] [PubMed] [Google Scholar]
- Johansen D, Cruciani V, Sundset R, Ytrehus K, Mikalsen SO. Ischemia induces closure of gap junctional channels and opening of hemichannels in heart-derived cells and tissue. Cell Physiol Biochem. 2011;28:103–114. doi: 10.1159/000331719. [DOI] [PubMed] [Google Scholar]
- Johnson KE, Mitra S, Katoch P, Kelsey LS, Johnson KR, Mehta PP. Phosphorylation on Ser-279 and Ser-282 of connexin43 regulates endocytosis and gap junction assembly in pancreatic cancer cells. Mol Biol Cell. 2013;24:715–733. doi: 10.1091/mbc.E12-07-0537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnstone SR, Kroncke BM, Straub AC, Best AK, Dunn CA, Mitchell LA, et al. MAPK phosphorylation of connexin 43 promotes binding of cyclin e and smooth muscle cell proliferation. Circ Res. 2012;111:201–211. doi: 10.1161/CIRCRESAHA.112.272302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnstone SR, Ross J, Rizzo MJ, Straub AC, Lampe PD, Leitinger N, et al. Oxidized phospholipid species promote in vivo differential cx43 phosphorylation and vascular smooth muscle cell proliferation. Am J Pathol. 2009;175:916–924. doi: 10.2353/ajpath.2009.090160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jordan K, Chodock R, Hand AR, Laird DW. The origin of annular junctions: a mechanism of gap junction internalization. J Cell Sci. 2001;114:763–773. doi: 10.1242/jcs.114.4.763. [DOI] [PubMed] [Google Scholar]
- Joshi MS, Mihm MJ, Cook AC, Schanbacher BL, Bauer JA. Alterations in connexin 43 during diabetic cardiomyopathy: Competition of tyrosine nitration versus phosphorylation. J Diabetes. 2015;7:250–259. doi: 10.1111/1753-0407.12164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang J, Kang N, Lovatt D, Torres A, Zhao Z, Lin J, Nedergaard M. Connexin 43 hemichannels are permeable to ATP. J Neurosci. 2008;28:4702–4711. doi: 10.1523/JNEUROSCI.5048-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanno S, Kovacs A, Yamada KA, Saffitz JE. Connexin43 as a determinant of myocardial infarct size following coronary occlusion in mice. J Am Coll Cardiol. 2003;41:681–686. doi: 10.1016/s0735-1097(02)02893-0. [DOI] [PubMed] [Google Scholar]
- Kapoor N, Galang G, Marban E, Cho HC. Transcriptional suppression of connexin43 by TBX18 undermines cell-cell electrical coupling in postnatal cardiomyocytes. J Biol Chem. 2011;286:14073–14079. doi: 10.1074/jbc.M110.185298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kasahara H, Ueyama T, Wakimoto H, Liu MK, Maguire CT, Converso KL, et al. Nkx2.5 homeoprotein regulates expression of gap junction protein connexin 43 and sarcomere organization in postnatal cardiomyocytes. J Mol Cell Cardiol. 2003;35:243–256. doi: 10.1016/s0022-2828(03)00002-6. [DOI] [PubMed] [Google Scholar]
- Kelly JJ, Simek J, Laird DW. Mechanisms linking connexin mutations to human diseases. Cell Tissue Res. 2014 doi: 10.1007/s00441-014-2024-4. Nov 16. [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- Kiec-Wilk B, Czech U, Janczarska K, Knapp A, Goralska J, Cialowicz U, et al. Connexin 43 and metabolic effect of fatty acids in stressed endothelial cells. Genes Nutr. 2012.;7:257–263. doi: 10.1007/s12263-011-0247-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kieken F, Mutsaers N, Dolmatova E, Virgil K, Wit AL, Kellezi A, et al. Structural and molecular mechanisms of gap junction remodeling in epicardial border zone myocytes following myocardial infarction. Circ Res. 2009;104:1103–1112. doi: 10.1161/CIRCRESAHA.108.190454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- King TJ, Lampe PD. Temporal regulation of connexin phosphorylation in embryonic and adult tissues. Biochim Biophys Acta. 2005;1719:24–35. doi: 10.1016/j.bbamem.2005.07.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirca M, Kleinbongard P, Soetkamp D, Heger J, Csonka C, Ferdinandy P, et al. Interaction between Connexin 43 and nitric oxide synthase in mice heart mitochondria. J Cell Mol Med. 2015;19:815–825. doi: 10.1111/jcmm.12499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klueg KM, Parody TR, Muskavitch MA. Complex proteolytic processing acts on Delta, a transmembrane ligand for Notch, during Drosophila development. Mol Biol Cell. 1998;9:1709–1723. doi: 10.1091/mbc.9.7.1709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knezl V, Bacova B, Kolenova L, Mitasikova M, Weismann P, Drimal J, et al. Distinct lethal arrhythmias susceptibility is associated with sex-related difference in myocardial connexin-43 expression. Neuro Endocrinol Lett. 2008;29:798–801. [PubMed] [Google Scholar]
- Kondo RP, Wang SY, John SA, Weiss JN, Goldhaber JI. Metabolic inhibition activates a non-selective current through connexin hemichannels in isolated ventricular myocytes. J Mol Cell Cardiol. 2000;32:1859–1872. doi: 10.1006/jmcc.2000.1220. [DOI] [PubMed] [Google Scholar]
- Kostin S, Dammer S, Hein S, Klovekorn WP, Bauer EP, Schaper J. Connexin 43 expression and distribution in compensated and decompensated cardiac hypertrophy in patients with aortic stenosis. Cardiovasc Res. 2004;62:426–436. doi: 10.1016/j.cardiores.2003.12.010. [DOI] [PubMed] [Google Scholar]
- Kostin S, Rieger M, Dammer S, Hein S, Richter M, Klovekorn WP, et al. Gap junction remodeling and altered connexin43 expression in the failing human heart. Mol Cell Biochem. 2003;242:135–144. [PubMed] [Google Scholar]
- Kozoriz MG, Church J, Ozog MA, Naus CC, Krebs C. Temporary sequestration of potassium by mitochondria in astrocytes. J Biol Chem. 2010a;285:31107–31119. doi: 10.1074/jbc.M109.082073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kozoriz MG, Bechberger JF, Bechberger GR, Suen MW, Moreno AP, Maass K, et al. The connexin43 C-terminal region mediates neuroprotection during stroke. J Neuropathol Exp Neurol. 2010b;69:196–206. doi: 10.1097/NEN.0b013e3181cd44df. [DOI] [PubMed] [Google Scholar]
- Krysko DV, Leybaert L, Vandenabeele P, D’Herde K. Gap junctions and the propagation of cell survival and cell death signals. Apoptosis. 2005;10:459–469. doi: 10.1007/s10495-005-1875-2. [DOI] [PubMed] [Google Scholar]
- Laing JG, Beyer EC. The gap junction protein connexin43 is degraded via the ubiquitin proteasome pathway. J Biol Chem. 1995;270:26399–26403. doi: 10.1074/jbc.270.44.26399. [DOI] [PubMed] [Google Scholar]
- Laird DW. Life cycle of connexins in health and disease. Biochem J. 2006;394:527–543. doi: 10.1042/BJ20051922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laird DW, Puranam KL, Revel JP. Turnover and phosphorylation dynamics of connexin43 gap junction protein in cultured cardiac myocytes. Biochem J. 1991;273:67–72. doi: 10.1042/bj2730067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lampe PD. Analyzing phorbol ester effects on gap junction communication: A dramatic inhibition of assembly. J Cell Biol. 1994;127:1895–1905. doi: 10.1083/jcb.127.6.1895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lampe PD, Cooper CD, King TJ, Burt JM. Analysis of Connexin43 phosphorylated at S325, S328 and S330 in normoxic and ischemic heart. J Cell Sci. 2006;119:3435–3442. doi: 10.1242/jcs.03089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lampe PD, Kurata WE, Warn-Cramer B, Lau AF. Formation of a distinct connexin43 phosphoisoform in mitotic cells is dependent upon p34cdc2 kinase. J Cell Sci. 1998a;111:833–841. doi: 10.1242/jcs.111.6.833. [DOI] [PubMed] [Google Scholar]
- Lampe PD, Kurata WE, Warn-Cramer BJ, Lau AF. Mitotic cells contain a distinct connexin43 phosphoisoform. In: Werner R, editor. Gap Junctions. IOS Press; Amsterdam: 1998b. pp. 249–253. [DOI] [PubMed] [Google Scholar]
- Lampe PD, Nguyen BP, Gil S, Usui M, Olerud J, Takada Y, et al. Cellular interaction of integrin 3 1 with lamin 5 promotes gap junctional communication. J Cell Biol. 1998;143:1735–1747. doi: 10.1083/jcb.143.6.1735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lancaster TS, Jefferson SJ, Korzick DH. Local delivery of a PKCepsilon-activating peptide limits ischemia reperfusion injury in the aged female rat heart. Am J Physiol Regul Integr Comp Physiol. 2011;301:R1242–R1249. doi: 10.1152/ajpregu.00851.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee TM, Chou TF. Troglitazone administration limits infarct size by reduced phosphorylation of canine myocardial connexin43 proteins. Am J Physiol Heart Circ Physiol. 2003;285:H1650–H1659. doi: 10.1152/ajpheart.00407.2002. [DOI] [PubMed] [Google Scholar]
- Lee TM, Lin MS, Chou TF, Tsai CH, Chang NC. Adjunctive 17beta-estradiol administration reduces infarct size by altered expression of canine myocardial connexin43 protein. Cardiovasc Res. 2004;63:109–117. doi: 10.1016/j.cardiores.2004.03.009. [DOI] [PubMed] [Google Scholar]
- Leithe E, Brech A, Rivedal E. Endocytic processing of connexin43 gap junctions: a morphological study. Biochem J. 2006;393:59–67. doi: 10.1042/BJ20050674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leybaert L, Sanderson MJ. Intercellular Ca(2+) waves: mechanisms and function. Physiol Rev. 2012;92:1359–1392. doi: 10.1152/physrev.00029.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li C, Meng Q, Yu X, Jing X, Xu P, Luo D. Regulatory effect of connexin 43 on basal Ca2+ signaling in rat ventricular myocytes. PLoS One. 2012;7:e36165. doi: 10.1371/journal.pone.0036165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li F, Sugishita K, Su Z, Ueda I, Barry WH. Activation of connexin-43 hemichannels can elevate [Ca(2+)]i and [Na(+)]i in rabbit ventricular myocytes during metabolic inhibition. J Mol Cell Cardiol. 2001;33:2145–2155. doi: 10.1006/jmcc.2001.1477. [DOI] [PubMed] [Google Scholar]
- Li H, Liu TF, Lazrak A, Peracchia C, Goldberg GS, Lampe PD, et al. Properties and regulation of gap junctional hemichannels in the plasma membranes of cultured cells. J Cell Biol. 1996;134:1019–1030. doi: 10.1083/jcb.134.4.1019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li G, Whittaker P, Yao M, Kloner RA, Przyklenk K. The gap junction uncoupler heptanol abrogates infarct size reduction with preconditioning in mouse hearts. Cardiovasc Pathol. 2002;11:158–165. doi: 10.1016/s1054-8807(02)00102-3. [DOI] [PubMed] [Google Scholar]
- Li J, Levin MD, Xiong Y, Petrenko N, Patel VV, Radice GL. N-cadherin haploinsufficiency affects cardiac gap junctions and arrhythmic susceptibility. J Mol Cell Cardiol. 2008;44:597–606. doi: 10.1016/j.yjmcc.2007.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J, Patel VV, Kostetskii I, Xiong Y, Chu AF, Jacobson JT, Yu C, et al. Cardiac-specific loss of N-cadherin leads to alteration in connexins with conduction slowing and arrhythmogenesis. Circ Res. 2005;97:474–481. doi: 10.1161/01.RES.0000181132.11393.18. [DOI] [PubMed] [Google Scholar]
- Li X, Heinzel FR, Boengler K, Schulz R, Heusch G. Role of connexin 43 in ischemic preconditioning does not involve intercellular communication through gap junctions. J Mol Cell Cardiol. 2004;36:161–163. doi: 10.1016/j.yjmcc.2003.10.019. [DOI] [PubMed] [Google Scholar]
- Liao CK, Cheng HH, Wang SD, Yeih DF, Wang SM. PKCvarepsilon mediates serine phosphorylation of connexin43 induced by lysophosphatidylcholine in neonatal rat cardiomyocytes. Toxicology. 2013;314:11–21. doi: 10.1016/j.tox.2013.08.001. [DOI] [PubMed] [Google Scholar]
- Lin JH, Lou N, Kang N, Takano T, Hu F, Han X, Xu Q, et al. A central role of connexin 43 in hypoxic preconditioning. J Neurosci. 2008;28:681–695. doi: 10.1523/JNEUROSCI.3827-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin JH, Weigel H, Cotrina ML, Liu S, Bueno E, Hansen AJ, et al. Gap-junction-mediated propagation and amplification of cell injury. Nat Neurosci. 1998;1:494–500. doi: 10.1038/2210. [DOI] [PubMed] [Google Scholar]
- Lin H, Ogawa K, Imanaga I, Tribulova N. Alterations of connexin 43 in the diabetic rat heart. Adv Cardiol. 2006a;42:243–254. doi: 10.1159/000092573. [DOI] [PubMed] [Google Scholar]
- Lin H, Ogawa K, Imanaga I, Tribulova N. Remodeling of connexin 43 in the diabetic rat heart. Mol Cell Biochem. 2006b;290:69–78. doi: 10.1007/s11010-006-9166-y. [DOI] [PubMed] [Google Scholar]
- Lin JH, Lou N, Kang N, Takano T, Hu F, Han X, et al. A central role of connexin 43 in hypoxic preconditioning. J Neurosci. 2008;28:681–695. doi: 10.1523/JNEUROSCI.3827-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin LC, Wu CC, Yeh HI, Lu LS, Liu YB, Lin SF, et al. Downregulated myocardial connexin 43 and suppressed contractility in rabbits subjected to a cholesterol-enriched diet. Lab Invest. 2005;85:1224–1237. doi: 10.1038/labinvest.3700324. [DOI] [PubMed] [Google Scholar]
- Lindsey ML, Escobar GP, Mukherjee R, Goshorn DK, Sheats NJ, Bruce JA, et al. Matrix metalloproteinase-7 affects connexin-43 levels, electrical conduction, and survival after myocardial infarction. Circulation. 2006;113:2919–2928. doi: 10.1161/CIRCULATIONAHA.106.612960. [DOI] [PubMed] [Google Scholar]
- Lu G, Jiang S, Ashraf M, Haider KH. Subcellular preconditioning of stem cells: mito-Cx43 gene targeting is cytoprotective via shift of mitochondrial Bak and Bcl-xL balance. Regen Med. 2012;7:323–334. doi: 10.2217/rme.12.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maass K, Ghanem A, Kim JS, Saathoff M, Urschel S, Kirfel G, et al. Defective epidermal barrier in neonatal mice lacking the C-terminal region of connexin43. Mol Biol Cell. 2004;15:4597–4608. doi: 10.1091/mbc.E04-04-0324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marquez-Rosado L, Singh D, Rincon-Arano H, Solan JL, Lampe PD. CASK (LIN2) interacts with Cx43 in wounded skin and their coexpression affects cell migration. J Cell Sci. 2012;125:695–702. doi: 10.1242/jcs.084400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marquez-Rosado L, Solan JL, Dunn CA, Norris RP, Lampe PD. Connexin43 phosphorylation in brain, cardiac, endothelial and epithelial tissues. Biochim Biophys Acta. 2012;1818:1985–1992. doi: 10.1016/j.bbamem.2011.07.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marston DJ, Dickinson S, Nobes CD. Rac-dependent trans-endocytosis of ephrinBs regulates Eph-ephrin contact repulsion. Nat Cell Biol. 2003;5:879–888. doi: 10.1038/ncb1044. [DOI] [PubMed] [Google Scholar]
- Matsuda T, Fujio Y, Nariai T, Ito T, Yamane M, Takatani T, et al. N-cadherin signals through Rac1 determine the localization of connexin 43 in cardiac myocytes. J Mol Cell Cardiol. 2006;40:495–502. doi: 10.1016/j.yjmcc.2005.12.010. [DOI] [PubMed] [Google Scholar]
- Matsuyama D, Kawahara K. Proliferation of neonatal cardiomyocytes by connexin43 knockdown via synergistic inactivation of p38 MAPK and increased expression of FGF1. Basic Res Cardiol. 2009;104:631–642. doi: 10.1007/s00395-009-0029-z. [DOI] [PubMed] [Google Scholar]
- Matsuyama D, Kawahara K. Oxidative stress-induced formation of a positive-feedback loop for the sustained activation of p38 MAPK leading to the loss of cell division in cardiomyocytes soon after birth. Basic Res Cardiol. 2011;106:815–828. doi: 10.1007/s00395-011-0178-8. [DOI] [PubMed] [Google Scholar]
- McLachlan CS, Almsherqi ZA, Mossop P, Suzuki J, Leong ST, Deng Y. Down regulation of immuno-detectable cardiac connexin-43 in BALB/c mice following acute fasting. Int J Cardiol. 2009;136:99–102. doi: 10.1016/j.ijcard.2008.03.098. [DOI] [PubMed] [Google Scholar]
- McSpadden LC, Kirkton RD, Bursac N. Electrotonic loading of anisotropic cardiac monolayers by unexcitable cells depends on connexin type and expression level. Am J Physiol Cell Physiol. 2009;297:C339–C351. doi: 10.1152/ajpcell.00024.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miro-Casas E, Ruiz-Meana M, Agullo E, Stahlhofen S, Rodriguez-Sinovas A, Cabestrero A, et al. Connexin43 in cardiomyocyte mitochondria contributes to mitochondrial potassium uptake. Cardiovasc Res. 2009;83:747–756. doi: 10.1093/cvr/cvp157. [DOI] [PubMed] [Google Scholar]
- Mitasikova M, Smidova S, Macsaliova A, Knezl V, Dlugosova K, Okruhlicova L, et al. Aged male and female spontaneously hypertensive rats benefit from n-3 polyunsaturated fatty acids supplementation. Physiol Res. 2008;57(Suppl 2):S39–S48. doi: 10.33549/physiolres.931550. [DOI] [PubMed] [Google Scholar]
- Miura T, Miki T, Yano T. Role of the gap junction in ischemic preconditioning in the heart. Am J Physiol Heart Circ Physiol. 2010;298:H1115–H1125. doi: 10.1152/ajpheart.00879.2009. [DOI] [PubMed] [Google Scholar]
- Miura T, Yano T, Naitoh K, Nishihara M, Miki T, Tanno M, et al. Delta-opioid receptor activation before ischemia reduces gap junction permeability in ischemic myocardium by PKC-epsilon-mediated phosphorylation of connexin 43. Am J Physiol Heart Circ Physiol. 2007;293:H1425–H1431. doi: 10.1152/ajpheart.01115.2006. [DOI] [PubMed] [Google Scholar]
- Mlinar B, Enyeart JJ. Block of current through T-type calcium channels by trivalent metal cations and nickel in neural rat and human cells. J Physiol. 1993;469:639–652. doi: 10.1113/jphysiol.1993.sp019835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molica F, Meens MJ, Morel S, Kwak BR. Mutations in cardiovascular connexin genes. Biol Cell. 2014;106:269–293. doi: 10.1111/boc.201400038. [DOI] [PubMed] [Google Scholar]
- Montero TD, Orellana JA. Hemichannels: new pathways for gliotransmitter release. Neuroscience. 2015;286:45–59. doi: 10.1016/j.neuroscience.2014.11.048. [DOI] [PubMed] [Google Scholar]
- Mohammad G, Kowluru RA. Novel role of mitochondrial matrix metalloproteinase-2 in the development of diabetic retinopathy. Invest Ophthalmol Vis Sci. 2011;52:3832–3841. doi: 10.1167/iovs.10-6368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mollerup S, Hofgaard JP, Braunstein TH, Kjenseth A, Leithe E, Rivedal E, et al. Norepinephrine inhibits intercellular coupling in rat cardiomyocytes by ubiquitination of connexin43 gap junctions. Cell Commun Adhes. 2011;18:57–65. doi: 10.3109/15419061.2011.611920. [DOI] [PubMed] [Google Scholar]
- Morel S, Frias MA, Rosker C, James RW, Rohr S, Kwak BR. The natural cardioprotective particle HDL modulates connexin43 gap junction channels. Cardiovasc Res. 2012;93:41–49. doi: 10.1093/cvr/cvr257. [DOI] [PubMed] [Google Scholar]
- Moreno AP, Chanson M, Elenes S, Anumonwo J, Scerri I, Gu H, et al. Role of the carboxyl terminal of connexin43 in transjunctional fast voltage gating. Circ Res. 2002;90:450–457. doi: 10.1161/hh0402.105667. [DOI] [PubMed] [Google Scholar]
- Muhlfeld C, Cetegen C, Freese S, Volkmann R, Hellige G, Vetterlein F. Phosphorylation of extrajunctional Cx43 in ischemic-preconditioned rat hearts. J Surg Res. 2010;162:e1–e8. doi: 10.1016/j.jss.2010.02.024. [DOI] [PubMed] [Google Scholar]
- Musil LS, Beyer EC, Goodenough DA. Expression of the gap junction protein connexin43 in embryonic chick lens: Molecular cloning, ultrastructural localization, and post-translational phosphorylation. J Membr Biol. 1990;116:163–175. doi: 10.1007/BF01868674. [DOI] [PubMed] [Google Scholar]
- Musil LS, Goodenough DA. Biochemical analysis of connexin43 intracellular transport, phosphorylation and assembly into gap junctional plaques. J Cell Biol. 1991;115:1357–1374. doi: 10.1083/jcb.115.5.1357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naitoh K, Ichikawa Y, Miura T, Nakamura Y, Miki T, Ikeda Y, et al. MitoKATP channel activation suppresses gap junction permeability in the ischemic myocardium by an ERK-dependent mechanism. Cardiovasc Res. 2006;70:374–383. doi: 10.1016/j.cardiores.2006.01.023. [DOI] [PubMed] [Google Scholar]
- Naitoh K, Yano T, Miura T, Itoh T, Miki T, Tanno M, et al. Roles of Cx43-associated protein kinases in suppression of gap junction-mediated chemical coupling by ischemic preconditioning. Am J Physiol Heart Circ Physiol. 2009;296:H396–H403. doi: 10.1152/ajpheart.00448.2008. [DOI] [PubMed] [Google Scholar]
- Nakase T, Fushiki S, Naus CC. Astrocytic gap junctions composed of connexin 43 reduce apoptotic neuronal damage in cerebral ischemia. Stroke. 2003;34:1987–1993. doi: 10.1161/01.STR.0000079814.72027.34. [DOI] [PubMed] [Google Scholar]
- Nakase T, Sohl G, Theis M, Willecke K, Naus CC. Increased apoptosis and inflammation after focal brain ischemia in mice lacking connexin43 in astrocytes. Am J Pathol. 2004;164:2067–2075. doi: 10.1016/S0002-9440(10)63765-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nickel B, Boller M, Schneider K, Shakespeare T, Gay V, Murray SA. Visualizing the effect of dynamin inhibition on annular gap vesicle formation and fission. J Cell Sci. 2013;126:2607–2616. doi: 10.1242/jcs.116269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nygren A, Olson ML, Chen KY, Emmett T, Kargacin G, Shimoni Y. Propagation of the cardiac impulse in the diabetic rat heart: reduced conduction reserve. J Physiol. 2007;580:543–560. doi: 10.1113/jphysiol.2006.123729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Carroll SJ, Alkadhi M, Nicholson LF, Green CR. Connexin 43 mimetic peptides reduce swelling, astrogliosis, and neuronal cell death after spinal cord injury. Cell Commun Adhes. 2008;15:27–42. doi: 10.1080/15419060802014164. [DOI] [PubMed] [Google Scholar]
- O'Carroll SJ, Gorrie CA, Velamoor S, Green CR, Nicholson LF. Connexin43 mimetic peptide is neuroprotective and improves function following spinal cord injury. Neurosci Res. 2013;75:256–267. doi: 10.1016/j.neures.2013.01.004. [DOI] [PubMed] [Google Scholar]
- Omasits U, Ahrens CH, Muller S, Wollscheid B. Protter: interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics. 2014;30:884–886. doi: 10.1093/bioinformatics/btt607. [DOI] [PubMed] [Google Scholar]
- Orellana JA, Avendano BC, Montero TD. Role of connexins and pannexins in ischemic stroke. Curr Med Chem. 2014;21:2165–2182. doi: 10.2174/0929867321666131228191714. [DOI] [PubMed] [Google Scholar]
- Orellana JA, Martinez AD, Retamal MA. Gap junction channels and hemichannels in the CNS: regulation by signaling molecules. Neuropharmacology. 2013;75:567–582. doi: 10.1016/j.neuropharm.2013.02.020. [DOI] [PubMed] [Google Scholar]
- Orellana JA, Stehberg J. Hemichannels: new roles in astroglial function. Front Physiol. 2014;5:193. doi: 10.3389/fphys.2014.00193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pahujaa M, Anikin M, Goldberg GS. Phosphorylation of connexin43 induced by Src: regulation of gap junctional communication between transformed cells. Exp Cell Res. 2007;313:4083–4090. doi: 10.1016/j.yexcr.2007.09.010. [DOI] [PubMed] [Google Scholar]
- Palatinus JA, O'Quinn MP, Barker RJ, Harris BS, Jourdan J, Gourdie RG. ZO-1 determines adherens and gap junction localization at intercalated disks. Am J Physiol Heart Circ Physiol. 2011;300:H583–H594. doi: 10.1152/ajpheart.00999.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pedrotty DM, Klinger RY, Kirkton RD, Bursac N. Cardiac fibroblast paracrine factors alter impulse conduction and ion channel expression of neonatal rat cardiomyocytes. Cardiovasc Res. 2009;83:688–697. doi: 10.1093/cvr/cvp164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Penna C, Perrelli MG, Raimondo S, Tullio F, Merlino A, Moro F, et al. Postconditioning induces an anti-apoptotic effect and preserves mitochondrial integrity in isolated rat hearts. Biochim Biophys Acta. 2009;1787:794–801. doi: 10.1016/j.bbabio.2009.03.013. [DOI] [PubMed] [Google Scholar]
- Peracchia C. Chemical gating of gap junction channels; roles of calcium, pH and calmodulin. Biochim Biophys Acta. 2004;1662:61–80. doi: 10.1016/j.bbamem.2003.10.020. [DOI] [PubMed] [Google Scholar]
- Pereda AE. Electrical synapses and their functional interactions with chemical synapses. Nat Rev Neurosci. 2014;15:250–263. doi: 10.1038/nrn3708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez Velazquez JL, Kokarovtseva L, Sarbaziha R, Jeyapalan Z, Leshchenko Y. Role of gap junctional coupling in astrocytic networks in the determination of global ischaemia-induced oxidative stress and hippocampal damage. Eur J Neurosci. 2006;23:1–10. doi: 10.1111/j.1460-9568.2005.04523.x. [DOI] [PubMed] [Google Scholar]
- Petrich BG, Eloff BC, Lerner DL, Kovacs A, Saffitz JE, Rosenbaum DS, et al. Targeted activation of c-Jun N-terminal kinase in vivo induces restrictive cardiomyopathy and conduction defects. J Biol Chem. 2004;279:15330–15338. doi: 10.1074/jbc.M314142200. [DOI] [PubMed] [Google Scholar]
- Petrich BG, Gong X, Lerner DL, Wang X, Brown JH, Saffitz JE, et al. c-Jun N-terminal kinase activation mediates downregulation of connexin43 in cardiomyocytes. Circ Res. 2002;91:640–647. doi: 10.1161/01.res.0000035854.11082.01. [DOI] [PubMed] [Google Scholar]
- Piehl M, Lehmann C, Gumpert A, Denizot JP, Segretain D, Falk MM. Internalization of large double-membrane intercellular vesicles by a clathrin-dependent endocytic process. Mol Biol Cell. 2007;18:337–347. doi: 10.1091/mbc.E06-06-0487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polontchouk L, Ebelt B, Jackels M, Dhein S. Chronic effects of endothelin 1 and angiotensin II on gap junctions and intercellular communication in cardiac cells. FASEB J. 2002;16:87–89. doi: 10.1096/fj.01-0381fje. [DOI] [PubMed] [Google Scholar]
- Ponsaerts R, D'Hondt C, Hertens F, Parys JB, Leybaert L, Vereecke J, et al. RhoA GTPase switch controls Cx43-hemichannel activity through the contractile system. PLoS One. 2012;7:e42074. doi: 10.1371/journal.pone.0042074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ponsaerts R, De Vuyst E, Retamal M, D'Hondt C, Vermeire D, Wang N, et al. Intramolecular loop/tail interactions are essential for connexin 43-hemichannel activity. FASEB J. 2010;24:4378–4395. doi: 10.1096/fj.09-153007. [DOI] [PubMed] [Google Scholar]
- Ponsaerts R, Wang N, Himpens B, Leybaert L, Bultynck G. The contractile system as a negative regulator of the connexin 43 hemichannel. Biol Cell. 2012;104:367–377. doi: 10.1111/boc.201100079. [DOI] [PubMed] [Google Scholar]
- Qu J, Volpicelli FM, Garcia LI, Sandeep N, Zhang J, Marquez-Rosado L, et al. Gap junction remodeling and spironolactone-dependent reverse remodeling in the hypertrophied heart. Circ Res. 2009;104:365–371. doi: 10.1161/CIRCRESAHA.108.184044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radosinska J, Bacova B, Bernatova I, Navarova J, Zhukovska A, Shysh A, et al. Myocardial NOS activity and connexin-43 expression in untreated and omega-3 fatty acids-treated spontaneously hypertensive and hereditary hypertriglyceridemic rats. Mol Cell Biochem. 2011;347:163–173. doi: 10.1007/s11010-010-0625-0. [DOI] [PubMed] [Google Scholar]
- Radosinska J, Bacova B, Knezl V, Benova T, Zurmanova J, Soukup T, et al. Dietary omega-3 fatty acids attenuate myocardial arrhythmogenic factors and propensity of the heart to lethal arrhythmias in a rodent model of human essential hypertension. J Hypertens. 2013;31:1876–1885. doi: 10.1097/HJH.0b013e328362215d. [DOI] [PubMed] [Google Scholar]
- Rami A, Volkmann T, Winckler J. Effective reduction of neuronal death by inhibiting gap junctional intercellular communication in a rodent model of global transient cerebral ischemia. Exp Neurol. 2001;170:297–304. doi: 10.1006/exnr.2001.7712. [DOI] [PubMed] [Google Scholar]
- Rawanduzy A, Hansen A, Hansen TW, Nedergaard M. Effective reduction of infarct volume by gap junction blockade in a rodent model of stroke. J Neurosurg. 1997;87:916–920. doi: 10.3171/jns.1997.87.6.0916. [DOI] [PubMed] [Google Scholar]
- Remo BF, Qu J, Volpicelli FM, Giovannone S, Shin D, Lader J, et al. Phosphatase-resistant gap junctions inhibit pathological remodeling and prevent arrhythmias. Circ Res. 2011;108:1459–1466. doi: 10.1161/CIRCRESAHA.111.244046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Retamal MA, Schalper KA, Shoji KF, Bennett MV, Saez JC. Opening of connexin 43 hemichannels is increased by lowering intracellular redox potential. Proc Natl Acad Sci U S A. 2007;104:8322–8327. doi: 10.1073/pnas.0702456104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Revel JP, Karnovsky MJ. Hexagonal array of subunits in intercellular junctions of the mouse heart and liver. J Cell Biol. 1967;33:C7–C12. doi: 10.1083/jcb.33.3.c7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richards TS, Dunn CA, Carter WG, Usui ML, Olerud JE, et al. Protein kinase C spatially and temporally regulates gap junctional communication during human wound repair via phosphorylation of connexin43 on serine368. J Cell Biol. 2004;167:555–562. doi: 10.1083/jcb.200404142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riquelme MA, Kar R, Gu S, Jiang JX. Antibodies targeting extracellular domain of connexins for studies of hemichannels. Neuropharmacology. 2013;75:525–532. doi: 10.1016/j.neuropharm.2013.02.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossi DJ, Brady JD, Mohr C. Astrocyte metabolism and signaling during brain ischemia. Nat Neurosci. 2007;10:1377–1386. doi: 10.1038/nn2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez-Sinovas A, Boengler K, Cabestrero A, Gres P, Morente M, Ruiz-Meana M, et al. Translocation of connexin 43 to the inner mitochondrial membrane of cardiomyocytes through the heat shock protein 90-dependent TOM pathway and its importance for cardioprotection. Circ Res. 2006;99:93–101. doi: 10.1161/01.RES.0000230315.56904.de. [DOI] [PubMed] [Google Scholar]
- Rodriguez-Sinovas A, Sanchez JA, Gonzalez-Loyola A, Barba I, Morente M, Aguilar R, et al. Effects of substitution of Cx43 by Cx32 on myocardial energy metabolism, tolerance to ischaemia and preconditioning protection. J Physiol. 2010;588:1139–1151. doi: 10.1113/jphysiol.2009.186577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruiz-Meana M, Nunez E, Miro-Casas E, Martinez-Acedo P, Barba I, Rodriguez-Sinovas A, Inserte J, et al. Ischemic preconditioning protects cardiomyocyte mitochondria through mechanisms independent of cytosol. J Mol Cell Cardiol. 2014;68:79–88. doi: 10.1016/j.yjmcc.2014.01.001. [DOI] [PubMed] [Google Scholar]
- Saez JC, Berthoud VM, Branes MC, Martinez AD, Beyer EC. Plasma membrane channels formed by connexins: their regulation and functions. Physiol Rev. 2003;83:1359–1400. doi: 10.1152/physrev.00007.2003. [DOI] [PubMed] [Google Scholar]
- Saez JC, Leybaert L. Hunting for connexin hemichannels. FEBS Lett. 2014;588:1205–1211. doi: 10.1016/j.febslet.2014.03.004. [DOI] [PubMed] [Google Scholar]
- Saez JC, Martinez AD, Branes MC, Gonzalez HE. Regulation of gap junctions by protein phosphorylation. Brazilian J Med Biol Res. 1998;31:593–600. doi: 10.1590/s0100-879x1998000500001. [DOI] [PubMed] [Google Scholar]
- Saez JC, Retamal MA, Basilio D, Bukauskas FF, Bennett MV. Connexinbased gap junction hemichannels: gating mechanisms. Biochim Biophys Acta. 2005;1711:215–224. doi: 10.1016/j.bbamem.2005.01.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saez JC, Schalper KA, Retamal MA, Orellana JA, Shoji KF, Bennett MV. Cell membrane permeabilization via connexin hemichannels in living and dying cells. Exp Cell Res. 2010;316:2377–2389. doi: 10.1016/j.yexcr.2010.05.026. [DOI] [PubMed] [Google Scholar]
- Saito R, Graf R, Hubel K, Fujita T, Rosner G, Heiss WD. Reduction of infarct volume by halothane: effect on cerebral blood flow or perifocal spreading depression-like depolarizations. J Cereb Blood Flow Metab. 1997;17:857–864. doi: 10.1097/00004647-199708000-00004. [DOI] [PubMed] [Google Scholar]
- Salameh A, Apel D, Gonzalez CJ, von SS, Mohr FW, Daehnert I, Dhein S. On the different roles of AT1 and AT2 receptors in stretch-induced changes of connexin43 expression and localisation. Pflugers Arch. 2012;464:535–547. doi: 10.1007/s00424-012-1161-4. [DOI] [PubMed] [Google Scholar]
- Salameh A, Djilali H, Blanke K, Gonzalez CJ, von SS, Savtschenko A, Dhein S, et al. Cardiac fibroblasts inhibit beta-adrenoceptor-dependent connexin43 expression in neonatal rat cardiomyocytes. Naunyn Schmiedebergs Arch Pharmacol. 2013;386:421–433. doi: 10.1007/s00210-013-0843-6. [DOI] [PubMed] [Google Scholar]
- Salameh A, Frenzel C, Boldt A, Rassler B, Glawe I, Schulte J, et al. Subchronic alpha- and beta-adrenergic regulation of cardiac gap junction protein expression. FASEB J. 2006;20:365–367. doi: 10.1096/fj.05-4871fje. [DOI] [PubMed] [Google Scholar]
- Salameh A, Karl S, Djilali H, Dhein S, Janousek J, Daehnert I. Opposing and synergistic effects of cyclic mechanical stretch and alpha- or beta-adrenergic stimulation on the cardiac gap junction protein Cx43. Pharmacol Res. 2010a;62:506–513. doi: 10.1016/j.phrs.2010.08.002. [DOI] [PubMed] [Google Scholar]
- Salameh A, Krautblatter S, Baessler S, Karl S, Rojas GD, Dhein S, et al. Signal transduction and transcriptional control of cardiac connexin43 up-regulation after alpha 1-adrenoceptor stimulation. J Pharmacol Exp Ther. 2008;326:315–322. doi: 10.1124/jpet.108.136663. [DOI] [PubMed] [Google Scholar]
- Salameh A, Krautblatter S, Karl S, Blanke K, Gomez DR, Dhein S, et al. The signal transduction cascade regulating the expression of the gap junction protein connexin43 by beta-adrenoceptors. Br J Pharmacol. 2009;158:198–208. doi: 10.1111/j.1476-5381.2009.00344.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salameh A, Schneider P, Muhlberg K, Hagendorff A, Dhein S, Pfeiffer D. Chronic regulation of the expression of gap junction proteins connexin40, connexin43, and connexin45 in neonatal rat cardiomyocytes. Eur J Pharmacol. 2004;503:9–16. doi: 10.1016/j.ejphar.2004.09.024. [DOI] [PubMed] [Google Scholar]
- Salameh A, Wustmann A, Karl S, Blanke K, Apel D, Rojas-Gomez D, et al. Cyclic mechanical stretch induces cardiomyocyte orientation and polarization of the gap junction protein connexin43. Circ Res. 2010b;106:1592–1602. doi: 10.1161/CIRCRESAHA.109.214429. [DOI] [PubMed] [Google Scholar]
- Sanchez JA, Rodriguez-Sinovas A, Barba I, Miro-Casas E, Fernandez-Sanz C, Ruiz-Meana M, et al. Activation of RISK and SAFE pathways is not involved in the effects of Cx43 deficiency on tolerance to ischemia-reperfusion injury and preconditioning protection. Basic Res Cardiol. 2013;108:351. doi: 10.1007/s00395-013-0351-3. [DOI] [PubMed] [Google Scholar]
- Sánchez JA, Rodríguez-Sinovas A, Fernández-Sanz C, Ruiz-Meana M, García-Dorado D. Effects of a reduction in the number of gap junction channels or in their conductance on ischemia-reperfusion arrhythmias in isolated mouse hearts. Am J Physiol Heart Circ Physiol. 2011;301:H2442–53. doi: 10.1152/ajpheart.00540.2011. [DOI] [PubMed] [Google Scholar]
- Sato M, Jiao Q, Honda T, Kurotani R, Toyota E, Okumura S, et al. Activator of G protein signaling 8 (AGS8) is required for hypoxia-induced apoptosis of cardiomyocytes: role of G betagamma and connexin 43 (CX43) J Biol Chem. 2009;284:31431–31440. doi: 10.1074/jbc.M109.014068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato T, Ohkusa T, Honjo H, Suzuki S, Yoshida MA, Ishiguro YS, et al. Altered expression of connexin43 contributes to the arrhythmogenic substrate during the development of heart failure in cardiomyopathic hamster. Am J Physiol Heart Circ Physiol. 2008;294:H1164–H1173. doi: 10.1152/ajpheart.00960.2007. [DOI] [PubMed] [Google Scholar]
- Schalper KA, Palacios-Prado N, Orellana JA, Saez JC. Currently used methods for identification and characterization of hemichannels. Cell Commun Adhes. 2008;15:207–218. doi: 10.1080/15419060802014198. [DOI] [PubMed] [Google Scholar]
- Schalper KA, Sanchez HA, Lee SC, Altenberg GA, Nathanson MH, Saez JC. Connexin 43 hemichannels mediate the Ca2+ influx induced by extracellular alkalinization. Am J Physiol Cell Physiol. 2010;299:C1504–1515. doi: 10.1152/ajpcell.00015.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schulz R, Boengler K, Totzeck A, Luo Y, Garcia-Dorado D, Heusch G. Connexin 43 in ischemic pre- and postconditioning. Heart Fail Rev. 2007;12:261–266. doi: 10.1007/s10741-007-9032-3. [DOI] [PubMed] [Google Scholar]
- Schulz R, Gres P, Skyschally A, Duschin A, Belosjorow S, Konietzka I, et al. Ischemic preconditioning preserves connexin 43 phosphorylation during sustained ischemia in pig hearts in vivo. FASEB J. 2003;17:1355–1357. doi: 10.1096/fj.02-0975fje. [DOI] [PubMed] [Google Scholar]
- Schwanke U, Konietzka I, Duschin A, Li X, Schulz R, Heusch G. No ischemic preconditioning in heterozygous connexin43-deficient mice. Am J Physiol Heart Circ Physiol. 2002;283:H1740–H1742. doi: 10.1152/ajpheart.00442.2002. [DOI] [PubMed] [Google Scholar]
- Severs NJ. The cardiac gap junction and intercalated disc. Int J Cardiol. 1990;26:137–173. doi: 10.1016/0167-5273(90)90030-9. [DOI] [PubMed] [Google Scholar]
- Severs NJ, Bruce AF, Dupont E, Rothery S. Remodelling of gap junctions and connexin expression in diseased myocardium. Cardiovasc Res. 2008;80:9–19. doi: 10.1093/cvr/cvn133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Severs NJ, Coppen SR, Dupont E, Yeh HI, Ko YS, Matsushita T. Gap junction alterations in human cardiac disease. Cardiovasc Res. 2004;62:368–377. doi: 10.1016/j.cardiores.2003.12.007. [DOI] [PubMed] [Google Scholar]
- Severs NJ, Shovel KS, Slade AM, Powell T, Twist VW, Green CR. Fate of gap junctions in isolated adult mammalian cardiomyocytes. Circ Res. 1989;65:22–42. doi: 10.1161/01.res.65.1.22. [DOI] [PubMed] [Google Scholar]
- Sheu JJ, Chang LT, Chiang CH, Sun CK, Chang NK, Youssef AA, et al. Impact of diabetes on cardiomyocyte apoptosis and connexin43 gap junction integrity: role of pharmacological modulation. Int Heart J. 2007;48:233–245. doi: 10.1536/ihj.48.233. [DOI] [PubMed] [Google Scholar]
- Shintani-Ishida K, Uemura K, Yoshida K. Hemichannels in cardiomyocytes open transiently during ischemia and contribute to reperfusion injury following brief ischemia. Am J Physiol Heart Circ Physiol. 2007;293:H1714–H1720. doi: 10.1152/ajpheart.00022.2007. [DOI] [PubMed] [Google Scholar]
- Shyu KG, Chen CC, Wang BW, Kuan P. Angiotensin II receptor antagonist blocks the expression of connexin43 induced by cyclical mechanical stretch in cultured neonatal rat cardiac myocytes. J Mol Cell Cardiol. 2001;33:691–698. doi: 10.1006/jmcc.2000.1333. [DOI] [PubMed] [Google Scholar]
- Siller-Jackson AJ, Burra S, Gu S, Xia X, Bonewald LF, Sprague E, et al. Adaptation of connexin 43-hemichannel prostaglandin release to mechanical loading. J Biol Chem. 2008;283:26374–26382. doi: 10.1074/jbc.M803136200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siushansian R, Bechberger JF, Cechetto DF, Hachinski VC, Naus CC. Connexin43 null mutation increases infarct size after stroke. J Comp Neurol. 2001;440:387–394. doi: 10.1002/cne.1392. [DOI] [PubMed] [Google Scholar]
- Smyth JW, Zhang SS, Sanchez JM, Lamouille S, Vogan JM, Hesketh GG, et al. A 14-3-3 mode-1 binding motif initiates gap junction internalization during acute cardiac ischemia. Traffic. 2014;15:684–699. doi: 10.1111/tra.12169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soetkamp D, Nguyen TT, Menazza S, Hirschhäuser C, Hendgen-Cotta UB, Rassaf T, et al. S-nitrosation of mitochondrial connexin 43 regulates mitochondrial function. Basic Res Cardiol. 2014;109:433. doi: 10.1007/s00395-014-0433-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Solan JL, Fry MD, TenBroek EM, Lampe PD. Connexin43 phosphorylation at S368 is acute during S and G2/M and in response to protein kinase C activation. J Cell Sci. 2003;116:2203–2211. doi: 10.1242/jcs.00428. [DOI] [PubMed] [Google Scholar]
- Solan JL, Lampe PD. Connexin 43 in LA-25 cells with active v-src is phosphorylated on Y247, Y265, S262, S279/282, and S368 via multiple signaling pathways. Cell Commun Adhes. 2008;15:75–84. doi: 10.1080/15419060802014016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Solan JL, Marquez-Rosado L, Sorgen PL, Thornton PJ, Gafken PR, Lampe PD. Phosphorylation of Cx43 at S365 is a gatekeeper event that changes the structure of Cx43 and prevents downregulation by PKC. J Cell Biol. 2007;179:1301–1309. doi: 10.1083/jcb.200707060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sorgen PL, Duffy HS, Sahoo P, Coombs W, Delmar M, Spray DC. Structural changes in the carboxyl terminus of the gap junction protein connexin43 indicates signaling between binding domains for c-Src and zonula occludens-1. J Biol Chem. 2004;279:54695–54701. doi: 10.1074/jbc.M409552200. [DOI] [PubMed] [Google Scholar]
- Sosinsky GE, Solan JL, Gaietta GM, Ngan L, Lee GJ, Mackey MR, et al. The C-terminus of Connexin43 adopts different conformations in the golgi and gap junction as detected with structure specific antibodies. Biochem J. 2007;408:375–385. doi: 10.1042/BJ20070550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spacek J, Harris KM. Trans-endocytosis via spinules in adult rat hippocampus. J Neurosci. 2004;24:4233–4241. doi: 10.1523/JNEUROSCI.0287-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spinella F, Rosano L, Di Castro V, Nicotra MR, Natali PG, Bagnato A. Endothelin-1 decreases gap junctional intercellular communication by inducing phosphorylation of connexin 43 in human ovarian carcinoma cells. J Biol Chem. 2003;278:41294–41301. doi: 10.1074/jbc.M304785200. [DOI] [PubMed] [Google Scholar]
- Srisakuldee W, Jeyaraman MM, Nickel BE, Tanguy S, Jiang ZS, Kardami E. Phosphorylation of connexin-43 at serine 262 promotes a cardiac injury-resistant state. Cardiovasc Res. 2009;83:672–681. doi: 10.1093/cvr/cvp142. [DOI] [PubMed] [Google Scholar]
- Srisakuldee W, Makazan Z, Nickel BE, Zhang F, Thliveris JA, Pasumarthi KB, et al. The FGF-2-triggered protection of cardiac subsarcolemmal mitochondria from calcium overload is mitochondrial connexin 43-dependent. Cardiovasc Res. 2014;103:72–80. doi: 10.1093/cvr/cvu066. [DOI] [PubMed] [Google Scholar]
- Stagg RB, Fletcher WH. The hormone-induced regulation of contact-dependent cell-cell communication by phosphorylation. Endocr Rev. 1990;11:302–325. doi: 10.1210/edrv-11-2-302. [DOI] [PubMed] [Google Scholar]
- Stanbouly S, Kirshenbaum LA, Jones DL, Karmazyn M. Sodium hydrogen exchange 1 (NHE-1) regulates connexin 43 expression in cardiomyocytes via reverse mode sodium calcium exchange and c-Jun NH2-terminal kinase-dependent pathways. J Pharmacol Exp Ther. 2008;327:105–113. doi: 10.1124/jpet.108.140228. [DOI] [PubMed] [Google Scholar]
- Stauffer BL, Sobus RD, Sucharov CC. Sex differences in cardiomyocyte connexin43 expression. J Cardiovasc Pharmacol. 2011;58:32–39. doi: 10.1097/FJC.0b013e31821b70b4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stock A, Sies H. Thyroid hormone receptors bind to an element in the connexin43 promoter. Biol Chem. 2000;381:973–979. doi: 10.1515/BC.2000.120. [DOI] [PubMed] [Google Scholar]
- Stout CE, Costantin JL, Naus CC, Charles AC. Intercellular calcium signaling in astrocytes via ATP release through connexin hemichannels. J Biol Chem. 2002;277:10482–10488. doi: 10.1074/jbc.M109902200. [DOI] [PubMed] [Google Scholar]
- Sun Y, Zhao X, Yao Y, Qi X, Yuan Y, Hu Y. Connexin 43 interacts with Bax to regulate apoptosis of pancreatic cancer through a gap junction-independent pathway. Int J Oncol. 2012;41:941–948. doi: 10.3892/ijo.2012.1524. [DOI] [PubMed] [Google Scholar]
- Sundset R, Ytrehus K, Zhang Y, Saffitz JE, Yamada KA. Repeated simulated ischemia and protection against gap junctional uncoupling. Cell Commun Adhes. 2007;14:239–249. doi: 10.1080/15419060701821149. [DOI] [PubMed] [Google Scholar]
- Surinkaew S, Kumphune S, Chattipakorn S, Chattipakorn N. Inhibition of p38 MAPK during ischemia, but not reperfusion, effectively attenuates fatal arrhythmia in ischemia/reperfusion heart. J Cardiovasc Pharmacol. 2013;61:133–141. doi: 10.1097/FJC.0b013e318279b7b1. [DOI] [PubMed] [Google Scholar]
- Suzuki S, Ohkusa T, Sato T, Yoshida M, Yasui K, Miwa K, et al. Effects of aldosterone on Cx43 gap junction expression in neonatal rat cultured cardiomyocytes. Circ J. 2009;73:1504–1512. doi: 10.1253/circj.cj-08-1065. [DOI] [PubMed] [Google Scholar]
- Takeuchi H, Mizoguchi H, Doi Y, Jin S, Noda M, Liang J, et al. Blockade of gap junction hemichannel suppresses disease progression in mouse models of amyotrophic lateral sclerosis and Alzheimer's disease. PLoS One. 2011;6:e21108. doi: 10.1371/journal.pone.0021108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tansey EE, Kwaku KF, Hammer PE, Cowan DB, Federman M, Levitsky S, et al. Reduction and redistribution of gap and adherens junction proteins after ischemia and reperfusion. Ann Thorac Surg. 2006;82:1472–1479. doi: 10.1016/j.athoracsur.2006.04.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- TenBroek EM, Lampe PD, Solan JL, Reynhout JK, Johnson RG. Ser364 of connexin43 and the upregulation of gap junction assembly by cAMP. J Cell Biol. 2001;155:1307–1318. doi: 10.1083/jcb.200102017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torres A, Wang F, Xu Q, Fujita T, Dobrowolski R, Willecke K, Takano T, Nedergaard M. Extracellular Ca(2)(+) acts as a mediator of communication from neurons to glia. Sci Signal. 2012;5:ra8. doi: 10.1126/scisignal.2002160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Totzeck A, Boengler K, van de Sand A., Konietzka I, Gres P, Garcia-Dorado D, et al. No impact of protein phosphatases on connexin 43 phosphorylation in ischemic preconditioning. Am J Physiol Heart Circ Physiol. 2008;295:H2106–H2112. doi: 10.1152/ajpheart.00456.2008. [DOI] [PubMed] [Google Scholar]
- Tribulova N, Dupont E, Soukup T, Okruhlicova L, Severs NJ. Sex differences in connexin-43 expression in left ventricles of aging rats. Physiol Res. 2005;54:705–708. [PubMed] [Google Scholar]
- Trudeau K, Muto T, Roy S. Downregulation of mitochondrial connexin 43 by high glucose triggers mitochondrial shape change and cytochrome C release in retinal endothelial cells. Invest Ophthalmol Vis Sci. 2012;53:6675–6681. doi: 10.1167/iovs.12-9895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsukimoto M, Harada H, Ikari A, Takagi K. Involvement of chloride in apoptotic cell death induced by activation of ATPsensitive P2x7 purinoceptor. J. Biol. Chem. 2005;280:2653–2658. doi: 10.1074/jbc.M411072200. [DOI] [PubMed] [Google Scholar]
- Turner MS, Haywood GA, Andreka P, You L, Martin PE, Evans WH, et al. Reversible connexin 43 dephosphorylation during hypoxia and reoxygenation is linked to cellular ATP levels. Circ Res. 2004;95:726–733. doi: 10.1161/01.RES.0000144805.11519.1e. [DOI] [PubMed] [Google Scholar]
- Tyagi N, Vacek JC, Givvimani S, Sen U, Tyagi SC. Cardiac specific deletion of N-methyl-d-aspartate receptor 1 ameliorates mtMMP-9 mediated autophagy/mitophagy in hyperhomocysteinemia. J Recept Signal Transduct Res. 2010;30:78–87. doi: 10.3109/10799891003614808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verma V, Larsen BD, Coombs W, Lin X, Sarrou E, Taffet SM, et al. Design and characterization of the first peptidomimetic molecule that prevents acidification-induced closure of cardiac gap junctions. Heart Rhythm. 2010;7:1491–1498. doi: 10.1016/j.hrthm.2010.06.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verma V, Larsen BD, Coombs W, Lin X, Spagnol G, Sorgen PL, et al. Novel pharmacophores of connexin43 based on the "RXP" series of Cx43-binding peptides. Circ Res. 2009;105:176–184. doi: 10.1161/CIRCRESAHA.109.200576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vetter C, Zweifel M, Zuppinger C, Carrel T, Martin D, Haefliger JA, et al. Connexin 43 expression in human hypertrophied heart due to pressure and volume overload. Physiol Res. 2010;59:35–42. doi: 10.33549/physiolres.931654. [DOI] [PubMed] [Google Scholar]
- Vetterlein F, Muhlfeld C, Cetegen C, Volkmann R, Schrader C, Hellige G. Redistribution of connexin43 in regional acute ischemic myocardium: influence of ischemic preconditioning. Am J Physiol Heart Circ Physiol. 2006;291:H813–H819. doi: 10.1152/ajpheart.01177.2005. [DOI] [PubMed] [Google Scholar]
- Vinken M, Decrock E, Leybaert L, Bultynck G, Himpens B, Vanhaecke T, et al. Non-channel functions of connexins in cell growth and cell death. Biochim Biophys Acta. 2012;1818:2002–2008. doi: 10.1016/j.bbamem.2011.06.011. [DOI] [PubMed] [Google Scholar]
- Vinken M, Maes M, Cavill R, Valkenborg D, Ellis JK, Decrock E, et al. Proteomic and metabolomic responses to connexin43 silencing in primary hepatocyte cultures. Arch Toxicol. 2013;87:883–894. doi: 10.1007/s00204-012-0994-0. [DOI] [PubMed] [Google Scholar]
- Wang N, De Bock M, Antoons G, Gadicherla AK, Bol M, Decrock E, et al. Connexin mimetic peptides inhibit Cx43 hemichannel opening triggered by voltage and intracellular Ca2+ elevation. Basic Res Cardiol. 2012a;107:304. doi: 10.1007/s00395-012-0304-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang N, De Bock M, Decrock E, Bol M, Gadicherla A, Bultynck G, et al. Connexin targeting peptides as inhibitors of voltage- and intracellular Ca2+-triggered Cx43 hemichannel opening. Neuropharmacology. 2013a;75:506–516. doi: 10.1016/j.neuropharm.2013.08.021. [DOI] [PubMed] [Google Scholar]
- Wang N, De Vuyst E, Ponsaerts R, Boengler K, Palacios-Prado N, Wauman J, et al. Selective inhibition of Cx43 hemichannels by Gap19 and its impact on myocardial ischemia/reperfusion injury. Basic Res Cardiol. 2013b;108:309. doi: 10.1007/s00395-012-0309-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang XH, Zhuo XZ, Ni YJ, Gong M, Wang TZ, Lu Q, et al. Improvement of cardiac function and reversal of gap junction remodeling by Neuregulin-1beta in volume-overloaded rats with heart failure. J Geriatr Cardiol. 2012b;9:172–179. doi: 10.3724/SP.J.1263.2012.03271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warner A, Clements DK, Parikh S, Evans WH, DeHaan RL. Specific motifs in the external loops of connexin proteins can determine gap junction formation between chick heart myocytes. J Physiol. 1995;488:721–728. doi: 10.1113/jphysiol.1995.sp021003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warner DS, Ludwig PS, Pearlstein R, Brinkhous AD. Halothane reduces focal ischemic injury in the rat when brain temperature is controlled. Anesthesiology. 1995;82:1237–1245. doi: 10.1097/00000542-199505000-00019. [DOI] [PubMed] [Google Scholar]
- Watanabe M, Ichinose S, Sunamori M. Age-related changes in gap junctional protein of the rat heart. Exp Clin Cardiol. 2004;9:130–132. [PMC free article] [PubMed] [Google Scholar]
- Wu XF, Liu WT, Liu YP, Huang ZJ, Zhang YK, Song XJ. Reopening of ATP-sensitive potassium channels reduces neuropathic pain and regulates astroglial gap junctions in the rat spinal cord. Pain. 2011;152:2605–2615. doi: 10.1016/j.pain.2011.08.003. [DOI] [PubMed] [Google Scholar]
- Wu Y, Gu EW, Zhu Y, Zhang L, Liu XQ, Fang WP. Sufentanil limits the myocardial infarct size by preservation of the phosphorylated connexin 43. Int Immunopharmacol. 2012;13:341–346. doi: 10.1016/j.intimp.2012.04.009. [DOI] [PubMed] [Google Scholar]
- Xu HF, Ding YJ, Shen YW, Xue AM, Xu HM, Luo CL, et al. MicroRNA-1 represses Cx43 expression in viral myocarditis. Mol Cell Biochem. 2012;362:141–148. doi: 10.1007/s11010-011-1136-3. [DOI] [PubMed] [Google Scholar]
- Xu Q, Kopp RF, Chen Y, Yang JJ, Roe MW, Veenstra RD. Gating of connexin 43 gap junctions by a cytoplasmic loop calmodulin binding domain. Am J Physiol Cell Physiol. 2012;302:C1548–1556. doi: 10.1152/ajpcell.00319.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang B, Lin H, Xiao J, Lu Y, Luo X, Li B, et al. The muscle-specific microRNA miR-1 regulates cardiac arrhythmogenic potential by targeting GJA1 and KCNJ2. Nat Med. 2007;13:486–491. doi: 10.1038/nm1569. [DOI] [PubMed] [Google Scholar]
- Yang F, Chen WL, Zheng MZ, Yu GW, Xu HJ, Shen YL, et al. Heat shock protein 90 mediates anti-apoptotic effect of diazoxide by preventing the cleavage of Bid in hypothermic preservation rat hearts. J Heart Lung Transplant. 2011;30:928–934. doi: 10.1016/j.healun.2011.04.001. [DOI] [PubMed] [Google Scholar]
- Yasui K, Kada K, Hojo M, Lee JK, Kamiya K, Toyama J, et al. Cell-to-cell interaction prevents cell death in cultured neonatal rat ventricular myocytes. Cardiovasc Res. 2000;48:68–76. doi: 10.1016/s0008-6363(00)00145-0. [DOI] [PubMed] [Google Scholar]
- Yasuno S, Kuwahara K, Kinoshita H, Yamada C, Nakagawa Y, Usami S, et al. Angiotensin II type 1a receptor signalling directly contributes to the increased arrhythmogenicity in cardiac hypertrophy. Br J Pharmacol. 2013;170:1384–1395. doi: 10.1111/bph.12328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye ZC, Wyeth MS, Baltan-Tekkok S, Ransom BR. Functional hemichannels in astrocytes: a novel mechanism of glutamate release. J Neurosci. 2003;23:3588–3596. doi: 10.1523/JNEUROSCI.23-09-03588.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young RC, Schumann R, Zhang P. The signaling mechanisms of long distance intercellular calcium waves (far waves) in cultured human uterine myocytes. J Muscle Res Cell Motil. 2002;23:279–284. doi: 10.1023/a:1022052910585. [DOI] [PubMed] [Google Scholar]
- Yu W, Dahl G, Werner R. The connexin43 gene is responsive to oestrogen. Proc Biol Sci. 1994;255:125–132. doi: 10.1098/rspb.1994.0018. [DOI] [PubMed] [Google Scholar]
- Yue P, Zhang Y, Du Z, Xiao J, Pan Z, Wang N, et al. Ischemia impairs the association between connexin 43 and M3 subtype of acetylcholine muscarinic receptor (M3-mAChR) in ventricular myocytes. Cell Physiol Biochem. 2006;17:129–136. doi: 10.1159/000092074. [DOI] [PubMed] [Google Scholar]
- Zaman J, Patel P, Peters N. 207 Arrhythmia Inducibility in a Novel Normotensive Rodent Model of Arrhythmia is not Related to Connexin 43 Quantity and Phosphorylation States - Determining the Contribution of Hypertension and ageing on the Myocardial Substrate. Heart. 2014;100(Suppl 3):A113–A114. [Google Scholar]
- Zeevi-Levin N, Barac YD, Reisner Y, Reiter I, Yaniv G, Meiry G, et al. Gap junctional remodeling by hypoxia in cultured neonatal rat ventricular myocytes. Cardiovasc Res. 2005;66:64–73. doi: 10.1016/j.cardiores.2005.01.014. [DOI] [PubMed] [Google Scholar]
- Zhang YW, Kaneda M, Morita I. The gap junction-independent tumor-suppressing effect of connexin 43. J Biol Chem. 2003;278:44852–44856. doi: 10.1074/jbc.M305072200. [DOI] [PubMed] [Google Scholar]
- Zhang W, Zhao G, Hu X, Wang M, Li H, Ye Y, et al. Aliskiren-attenuated myocardium apoptosis via regulation of autophagy and connexin-43 in aged spontaneously hypertensive rats. J Cell Mol Med. 2014;18:1247–1256. doi: 10.1111/jcmm.12273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y, Kakinuma Y, Ando M, Katare RG, Yamasaki F, Sugiura T, et al. Acetylcholine inhibits the hypoxia-induced reduction of connexin43 protein in rat cardiomyocytes. J Pharmacol Sci. 2006;101:214–222. doi: 10.1254/jphs.fp0051023. [DOI] [PubMed] [Google Scholar]
- Zhang Y, Wang H, Kovacs A, Kanter EM, Yamada KA. Reduced expression of Cx43 attenuates ventricular remodeling after myocardial infarction via impaired TGF-beta signaling. Am J Physiol Heart Circ Physiol. 2010a;298:H477–H487. doi: 10.1152/ajpheart.00806.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y, Zhang L, Chu W, Wang B, Zhang J, Zhao M, et al. Tanshinone IIA inhibits miR-1 expression through p38 MAPK signal pathway in post-infarction rat cardiomyocytes. Cell Physiol Biochem. 2010b;26:991–998. doi: 10.1159/000324012. [DOI] [PubMed] [Google Scholar]
- Zhao LL, Chen HJ, Chen JZ, Yu M, Ni YL, Zhang WF. Losartan reduced connexin43 expression in left ventricular myocardium of spontaneously hypertensive rats. J Zhejiang Univ Sci B. 2008;9:448–454. doi: 10.1631/jzus.B0820050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao Q, Shao L, Hu X, Wu G, Du J, Xia J, Qiu H. Lipoxin a4 preconditioning and postconditioning protect myocardial ischemia/reperfusion injury in rats. Mediators Inflamm. 2013;2013:231351. doi: 10.1155/2013/231351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong JQ, Zhang W, Gao H, Li Y, Zhong M, Li D, et al. Changes in connexin 43, metalloproteinase and tissue inhibitor of metalloproteinase during tachycardia-induced cardiomyopathy in dogs. Eur J Heart Fail. 2007;9:23–29. doi: 10.1016/j.ejheart.2006.04.014. [DOI] [PubMed] [Google Scholar]
- Zhu H, Wang H, Zhang X, Hou X, Cao K, Zou J. Inhibiting N-cadherin-mediated adhesion affects gap junction communication in isolated rat hearts. Mol Cells. 2010;30:193–200. doi: 10.1007/s10059-010-0107-y. [DOI] [PubMed] [Google Scholar]