

Abstract
In Miocene times a vast wetland existed in Western Amazonia. Whereas the general development of this amazing ecosystem is well established, many questions remain open on sedimentary environments, stratigraphical correlations as well as its palaeogeographical configuration. Several outcrops located in a barely studied region around Eirunepé (SW Amazonas state, Brazil) were investigated to obtain basic sedimentological data. The observed deposits belong to the upper part of the Solimões Formation and are biostratigraphically dated to the Late Miocene. Vertically as well as laterally highly variable fine-grained clastic successions were recorded. Based on the lithofacies assemblages, these sediments represent fluvial deposits, possibly of an anastomosing river system. Sand bodies formed within active channels and dominant overbank fines are described (levees, crevasse splays/channels/deltas, abandoned channels, backswamps, floodplain paleosols). Lacustrine environments are restricted to local floodplain ponds/lakes. The mollusc and ostracod content as well as very light δ18O and δ13C values, measured on ostracod valves, refer to exclusively freshwater conditions. Based on palaeontological and geological results the existence of a long-lived lake (“Lake Pebas”) or any influx of marine waters can be excluded for that region during the Late Miocene.
Keywords: Western Amazonia, Late Miocene, “Lake Pebas”, Fluvial sediments
Highlights
► Around Eirunepé (Amazonia, Brazil) six outcrops were sedimentologically investigated. ► The sediments are of Late Miocene age and belong to the Solimões Formation. ► Lithofacies assemblages document a fluvial sedimentary environment. ► Molluscs, ostracods and stable isotopes refer to exclusively freshwater conditions. ► The existence of a long-lived lake or any influx of marine waters can be excluded.
1. Introduction
The geological and (palaeo-)biological evolution of lowland Amazonia during Neogene and Quaternary times remains fascinating since the early days of Natural Sciences (for summaries of research history see e.g., Loczy, 1963 and Wesselingh, 2008). Many, but partly highly controversial models have been introduced to explain its historical development (for recent compilations see e.g., Lundberg et al., 1998, Campbell et al., 2006, Wesselingh and Salo, 2006, Rossetti and Mann de Toledo, 2007, Haffer, 2008, Hoorn and Wesselingh, 2010, Hoorn et al., 2010, Latrubesse et al., 2010).
Generally, it is widely accepted that around the onset of the Miocene (∼23 Ma) a mega-wetland (“Pebas system”, also called “Lake Pebas”) developed in western Amazonia due to the subsiding Subandean foreland (e.g., Hoorn, 1994, Hoorn, 2006, Wesselingh et al., 2002, Wesselingh and Salo, 2006, Shephard et al., 2010). Short-lived marine incursions or even a transcontinental seaway from the Caribbean Sea through the Venezuelan/Columbian Llanos Basin southwards to the Argentinean Paraná Basin are proposed (e.g., Räsänen et al., 1995, Gingras et al., 2002, Hovikoski et al., 2007, Hovikoski et al., 2010, Uba et al., 2009) but heavily disputed (e.g., Campbell et al., 2006, Cozzuol, 2006, Westaway, 2006, Latrubesse et al., 2007, Latrubesse et al., 2010; compare also discussions in Vonhof et al., 2003, Hoorn and Vonhof, 2006, Wesselingh, 2006). In the Late Miocene this mega-wetland disintegrated due to enhanced uplift of the Northern/Central Andes. The drainage pattern of northern South America started to reverse completely to today’s easterly course and the modern “Amazon system” became established during the Early Pliocene (Hoorn, 2006, Figueiredo et al., 2009, Hoorn et al., 2010, Latrubesse et al., 2010).
Beside the vast size of Amazonia and the still fragmentary regional coverage with field surveys, there are considerable inconsistencies in palaeoenvironmental reconstructions and, in particular, in the chronology and correlation of scattered outcrops. Wesselingh (2008, p. 5) stated: “The lack of geological data has led to the emergence of many grand theories about the origin of present-day Amazon system and its highly diversity, often based on dubious interpretations of the little data available”.
The present paper aims to contribute basic sedimentological data from a barely studied region (Eirunepé, ∼2.000 km SW Manaus; Fig. 1a), which is supposed to be placed at the south-eastern margin of the “Pebas system” (Wesselingh and Ramos, 2010). We demonstrate that there is no evidence for a long-lived lake (sensu Gorthner, 1994) or any marine influx. Conversely, we document a well-structured, aggrading fluvial system of Late Miocene age, which is in agreement with the sedimentation model and chronology proposed by Latrubesse et al., 2007, Latrubesse et al., 2010.
Fig. 1.

Location of the study area around Eirunepé (SW Amazonia). (a) Delineation of the Acre and Upper Amazonas Basin (isopaches refer to the thickness of the Solimões Formation; after Latrubesse et al., 2010). (b) Isopaches of the Solimões Fm., position of exploration wells (after Maia et al., 1977) and location of the investigated outcrops (AQ, Aquidabã; BA, Barro Branco; MN, Morada Nova; PD, Pau D’Alho; RE, Remanso; TO, Torre da Lua).
2. Working area and geological background
The studied sections are located along the Juruá and Tarauacá River, NE respectively SE of the city Eirunepé (state of Amazonia, western Brazil; ∼245 km S of Benjamin Constant; Fig. 1a and b).
Delineations and subdivisions of basins in western Amazonia diverge notably and several authors attribute the region of Eirunepé to the Solimões Basin (Eirunepé Subbasin; e.g., Caputo, 1991, Roddaz et al., 2005, Ramos, 2006, Wesselingh and Salo, 2006, Wesselingh et al., 2006a, Barata and Caputo, 2007). However, based on subsurface information, obtained by extensive hydrocarbon and coal exploration campaigns during the 1970’s, this region is situated west of a basement high (Miura, 1972; “Iquitos arch”; corresponds to the “Carauari arch” of Caputo, 1991) on the eastern margin of the Acre Basin (e.g., Del’ Arco et al., 1977, Maia et al., 1977, Latrubesse et al., 2010; Fig. 1a and b).
Beside Quaternary deposits (terraces, alluvium), the scattered outcrops along river banks expose sediments of the upper parts of the Solimões Formation (Del’ Arco et al., 1977, Maia et al., 1977, Paz et al., in press). The Solimões Fm. comprises more than 1000 m thick, largely pelitic–sandy alternations with lignitic intercalations and covers a huge part of western Amazonia (Fig. 1a). Uncertainties in its definition, its stratigraphical and geographical extent as well as its depositional environments basically reflect the ongoing debate about Amazonia’s history through Neogene times. Comprehensive reviews of the Solimões Fm. provide for example: Del’ Arco et al., 1977, Hoorn, 1993, Hoorn, 1994, Latrubesse et al., 1997, Latrubesse et al., 2010 and Silva-Caminha et al. (2010).
3. Field and laboratory work
The sections were vertically logged by visual inspection of the lithofacies (including colour, grain size, bedding planes, sedimentary structures, macrofossil content). For lithofacies coding the scheme of Miall (1996) was used: capital letters = dominant grain size (G, gravel; S, sand; F, fine sand–clay) followed by a lowercase letter = sedimentary structures and/or biogenic features (c, clast-supported; m, massive/faint lamination; h, horizontal bedding/lamination; t and p, trough and planar cross bedding; r, ripple bedding; s, scour fill; l, lamination; r, rooted; C, coal; P, pedogenic overprint). Additionally the outcrops were mapped laterally as far as possible. Due to poor outcrop conditions an application of the architectural element concept (Miall, 1996) was only loosely possible.
For micropalaeontological investigations bulk samples (∼1–2 kg) were taken from all outcrops. 500 g of dried sediment (40 °C, 24 h) were washed by using diluted hydrogen superoxide for disintegration through standard sieves (H2O2 : H2O = 1 : 5; 63/125/250/500 μm). Wet sieve residuals were washed with ethanol prior to drying (40 °C, 24 h). The residuals ≥250 μm were picked out completely. An in-depth taxonomic examination of the obtained microfossils (including the sieve fractions <250 μm) will be subject of further studies.
For preliminary stable isotope analyses (δ18O, δ13C; 18 samples) two or three ostracod valves (∼40–60 μg; Cyprideis spp.) from earlier sampling campaigns were used (collected by M.I.R). For analyses a Thermo-Finnigan Kiel II automated reaction system and a Thermo-Finnigan Delta Plus isotope-ratio mass spectrometer were used (University of Graz; standard deviation = 0.1‰ relative to NBS-19; results in per mille relative to VPDB). More detailed investigations are in preparation (M. Fonseca/Belém, M. Caporaletti/Graz).
4. Results – sedimentology of the outcrops
4.1. Pau D’Alho
Location: 12.6 km NE Eirunepé (S 06°33’55.5”/W 069°46’11.7”, altitude: ∼108 m), left cutbank of the Juruá River (Fig. 2).
Fig. 2.

Lithological section of Pau D’Alho and interpretation of depositional environment (legend also used in Fig. 4, Fig. 6, Fig. 8, Fig. 9, Fig. 10).
Description: The basal ∼1.8 m (layers 1–9) of the section consists of partly ripple bedded sands and laminated silts; some cm-thick clay intercalations were observed. Above follows a 0.65 m thick alternation of clay with incorporated calcareous nodules and laminated silty sand or ripple bedded sand (10–17; Fig. 3a). The top is rich in vertebrate remains (crocodiles, turtles; 17; Fig. 3b). Massive to finely laminated, occasionally bivalves-bearing (silty) clay with coaly and sandy layers forms the next 1.14 m (18–23; Fig. 3c and d). Beds 24–25 (1.85 m thick) comprise massive, horizontally or trough cross bedded sand and finely laminated or massive (sandy–silty) clay/fine sand (Fig. 3e). Rarely claws of crabs were found. These strata are topped by 0.70 m thick, reddish–grey mottled, rooted and massive clay (paleosol) with a calcareous horizon on its top (26). Up-section, a 4.2 m (27–39) thick alternation of massive or laminated clay, silt and partly ripple bedded fine sand is developed (a slight coarsening-upward trend is anticipated). Bed 32 displays a slightly erosive base with intraclasts in the lower part (mud pebbles). Layers 31–39 are cut by a channel (40) with a multiphase fill. Large scale cross bedded sands (ε-cross stratification) with sporadic intraclasts (mud pebbles) on reactivation surfaces and trough cross bedded and ripple bedded fine–medium sand (Fig. 3f) form the channel fill.
Fig. 3.

Outcrop photographs from Pau D’Alho (encircled numbers indicate bed numbers; scale bar in cm). (a) Horizons of calcareous nodules. (b) Bone bed on top of layer 17. (c) Laminated, coaly fine sand. (d) Finely laminated silty clay. (e) Finely laminated silt–clay alternations. (f) Ripple bedded fine sand on lateral accretion surfaces.
Microfossil contents: Samples from the basal part of the section (PD2) yielded only scarce valves of Cyprideis spp. and rare fish elements. PD18 contained several and PD20 numerous specimen of Cyprideis spp. In PD23 only very rarely Cyprideis spp. is recorded, accompanied by small bivalves (sphaeriids). PD25 delivered the richest fauna with Cypria sp., Cyprideis spp., Cytheridella sp., darwinulids and fish remains.
Interpretation: The lithofacies of the basal part of the outcrop (1–17) refers to a deposition in a fluvial overbank environment. These layers are probably related to levee and/or crevasse splay sedimentation. Scarce ostracod and fish fragments may indicate the presence of short-lived ponds. The diffuse layers of calcareous nodules (17) are supposed to be largely a diagenetic feature (groundwater caliche). However, it may also hint to soil-forming processes at seasonally dry floodplain areas (Retallack, 2001). Up-section the sediments become finer and contain several (sandy–silty) coal-rich layers (18–23). These beds are assigned to the formation of a floodplain pond, which was inhabited by opportunistic ostracods, freshwater bivalves and fishes. However, crevasse splays influenced episodically that shallow lacustrine environment (sandy and organic matter-rich layers). Later (layers 24–25), the influx of crevasse splays into the pond, which is still populated by ostracod faunas and fishes increased (more prominent sandy beds, partly scour-fills). Pedogenic processes are indicated by incipient soil formation in layer 26 (mottled, roots). Floodplain pond conditions ended above (beds 27–39) due to enhanced clastic input from an approaching channel. Finally (bed 40), the overbank fines were intersected by a channel of at least 2–3 m in depth, which was filled up by large scale cross bedded sand. More or less perpendicular to the main accretion surfaces oriented ripple bedding indicates deposition by lateral accretion of a point bar.
4.2. Morada Nova
Location: 17.9 km NE Eirunepé (S 06°32’51.1”/W 069°42’39.4”, altitude: ∼107 m), left cutbank of the Juruá River (Fig. 4).
Fig. 4.

Lithological sections of Morada Nova and interpretation of depositional environment (CS = crevasse splay, FF (CH) = abandoned channel, FPL = floodplain lake, OF = overbank fines).
Description: The lower ∼5 m of the outcrop are formed by (section 1): (1) greenish–grey, mottled clay with ferran cutans and cm-sized carbonate nodules (Fig. 5a); (2) dark grey, massive or indistinctly laminated clay with cm-sized carbonate concretions (Fig. 5b), thin, pyrite-rich coaly layers, plant fragments, root traces and vertebrate remains (crocodiles, turtles); (3) greenish–yellowish, mottled clay with ferran cutans (passes gradually into bed 4); (4) violet-red–green-grey, mottled clay/paleosol (Fig. 5c); and (5) green–grey, mottled clay with vertebrate remains (reptiles) on top. A trough cross bedded sandy layer with coaly interlayers in the SE part of the site mentioned by Paz et al. (in press) was not exposed during our observation (section 4). In the NW part (section 1), above layer 5, ripple bedded fine sand with intraclasts (mud pebbles) at its base is recorded, which contains bivalves and coalified wood remains (6). It is topped by massive clay–laminated sand alternations (7) and massive sandy clay, which is rich in molluscs in its lower and upper part (8). Up-section (9), an alternation of clay and coal layers, rich in molluscs (i.e., Mycetopoda sp.), as well as a thinly bedded succession of laminated sand and thin clay interlayers (10) follow. Towards the SE (section 2, 3, 4) an up to ∼2 m thick, large scale cross bedded sandy unit is developed (Fig. 5d). Occasionally, intraclasts and vertebrate fragments are found at the base of this element or on accretion surfaces, which dip around NE. The dip of ripple foresets is approximately perpendicular oriented to these surfaces (SW). This unit is overlain by a layer of calcareous nodules (11; section 3) and laminated clay (silt)–coal alternations with bivalve and vertebrate remains (13 and 14 = equivalent to 9). Bed 12 (plastic, strongly weathered clay) is an equivalent of layer 10.
Fig. 5.

Outcrop photographs from Morada Nova (encircled numbers indicate bed numbers; scale bar = 50 mm). (a) Greenish–grey paleosol. (b) Faintly laminated clay. (c) Violet-red paleosol. (d) Large scale cross bedded sand topped by a layer of calcareous nodules (11) and a strongly weathered clay (12). (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article).
Microfossil contents: Samples MN1 and MN2 lack microfossils, in MN8a and MN8b only scattered valves of Cypria sp. and Heterocypris? sp. were found. MN9 and MN 14 yielded many fish, gastropod and bivalve (sphaeriids) remains as well as abundant ostracod valves (Cyprideis spp.).
Earlier reports on fossil material: Celestino and Ramos (2007; see also Wesselingh and Ramos, 2010) documented the following ostracod taxa: Cyprideis graciosa, Cyprideis lacrimata, Cyprideis longispina, Cyprideis machadoi, Cyprideis pebasae, Cyprideis olivencai, Cyprideis sp. 1 and 2. Further findings (plants, fishes, reptiles) are mentioned in Del’ Arco et al. (1977) and Paz et al. (in press). The later authors also reported the occurrence of Cyathecidites annulatus, Echitricolporites spinosus and Grimsdalea magnaclavata pollen.
Interpretation: The basal part forms a succession of variable coloured paleosols. Greenish to yellowish and dark grey colouration might indicate more or less water-logged conditions, whereas reddish colours may hint to rather well-drained, oxidizing environments. However, diagenetic alteration can hamper such an interpretation significantly (Retallack, 2001). Layer 2 is less affected by pedogenic processes (faint lamination, coaly layers, plant and reptile fragments) and perhaps represents floodplain lake/backswamp deposits. The paleosols (overbank fines; 1–5) are overlain by a sandy channel fill (point bar) with lag deposits at its base and repeatedly occurring intraclasts at its lateral accretion surfaces (channel depth >2 m based on the preserved height of this unit). In the NW part of Morada Nova layers 6–8 represent an abandoned channel fill, which could be the fill of a subordinate channel or the fill of a chute channel. Up-section, the influx of the active channel decreased and a floodplain lake developed (9, 13, 14), which enabled the establishment of plentiful ostracod and aquatic molluscs faunas. The pond itself or its surroundings were richly vegetated (coaly layers) and settled by semi-aquatic reptiles. Finally, enhanced input of sandy matter points to an increasing influence of the channel, probably due to crevasse splays.
4.3. Aquidabã
Location: 22.0 km NE Eirunepé (S 06°31’40.8”/W 069°39’52.0”; altitude: ∼106 m), left cutbank of the Juruá River (Fig. 6).
Fig. 6.

Lithological sections of Aquidabã and interpretation of depositional environment (FF (CH) = abandoned channel).
Description: Section 1 (W part) starts with a 1.6 m thick succession of laminated, ripple and cross bedded silty sand with coaly intercalations (layers 1–6/1). Up-section (7–14/1), a 2.5 m thick sequence of ripple bedded silty fine sand layers with commonly erosive basal boundaries (incl. clay intraclasts) is developed. Convolute bedding and m-thick concretions are frequent in its upper part (11–14/1). The sediments above become successively finer (silty fine sand–fine sandy silt–silty clay; 15–20/1) and comprise coal and mollusc-rich pelitic (16/1, 19/1) and coaly layers (18/1) as well as carbonate-cemented clay (20/1). Layer 21/1 (0.75 m thick) is composed of greenish–violet, mottled or poorly laminated clay with scattered bivalve shells. Intercalated are several layers (∼10 cm-thick), which consist of 1–2 mm large, rounded clay clasts (Fig. 7a). It is overlain by 1 m of massive clay with abundant mollusc (e.g., Sheppardiconcha septencincta; Fig. 7b) and plant remains (incl. leaves; 22/1) and a >0.5 m thick (weathered) coaly layer (23/1) with mollusc fragments (e.g., Anodontites? sp., Castalia sp., S. septencincta; Fig. 7c).
Fig. 7.

Outcrop photographs from Aquidabã (encircled numbers indicate bed numbers; scale bar in cm). (a) Layer of crumb peds (section 1). (b) Mass occurrence of Sheppardiconcha (section 1). (c) Weathered mollusc remains (section 1). (d) Approximately 4 m wide and 0.8 m deep channel fill (section 2). (e) Bioturbation due to bivalves (Mycetopoda sp., section 2). (f) Coal and coaly fine sand–finely laminated clay alternations (section 2).
Section 2 (E part; ∼120 m downstream of section 2) starts with >0.35 m thick, laminated fine sand with coaly interlayers (layer 1/2). It is cut by a ∼4 m wide and 0.8 m deep channel fill (2–5/2; fining-upward, from base to top: laminated, coal-rich fine sand with rare mollusc and plant remains; ripple bedded fine sand, rich in plant fragments; massive clay, rich in bivalves (sphaeriids), several wood remains; Fig. 7d). This channel fill is topped by a 0.5 m thick, massive–poorly laminated clay with thin fine sand interlayers (6/2) and 0.2 m of massive, partly indurated (cemented) clay (7/2). Coalified wood remains are rare; bivalves (sphaeriids) are abundant (especially upper part of 6/2). Layer 6/2 and 7/2 taper off towards the W. Up-section (1 m thickness), follow massive, silty–sandy clays (8–9/2) and ripple bedded silty fine sand with silt interlayers (10–14/2). Especially layer 8/2 is rich in molluscs (sphaeriids), 13–14/2 contain coaly fragments; clay intraclasts are observed at the base of 13/2. These beds are topped by 2.7 m of mottled silty fine sand with coaly fragments in the lower 0.3 m (15/2). The uppermost 0.1 m is bioturbated by bivalves (Mycetopoda sp.; Fig. 7e). Layer 16/2 (>1.2 m thick) comprises coal–coaly fine sand–finely laminated clay alternations (Fig. 7f) and is rich in bivalves (sphaeriids) in the lowermost 5–7 cm.
Microfossil contents: Whereas samples AQ5/1 and AQ6/1 contain only rare ostracod remains, AQ16/1 yields, beside fish fragments, some valves of Cytheridella sp. and darwinulids, accompanied by rare specimens of Cypria sp., Cyprideis spp. and Heterocypris? sp. Sample AQ19/1 is very rich in ostracods (Cytheridella sp., darwinulids, Cypria sp., ilyocypridids) and fish bones. Several (AQ22a/1) respectively rare (AQ22b/1) Cyprideis spp. valves are recorded in the samples of section 1 above, together with fish remains and gastropods (AQ22b/1). Sample AQ3/2 of section 2 delivered only rarely Cyprideis spp. and some fish remains. In the samples AQ 5-6-8-13/2 plentiful ostracod faunas (Cytheridella sp., ilyocyprinids, darwinulids, Cyprideis spp., Heterocypris? sp.) are documented along with fish remains and sphaeriid bivalve shells (AQ8/2 and AQ13/2). AQ15/2 and AQ16/2 are almost barren of ostracods but contain fish and bivalve (sphaeriids) fossils.
Earlier reports on fossil material: Celestino and Ramos (2007; see also Wesselingh and Ramos, 2010) recorded the following ostracod taxa: Alicenula (Darwinula) fragilis, Cypria aqualica, C. longispina, C. pebasae, Cyprideis sp. 3, Cytheridella purperae, Heterocypris? sp., Ilyocypris (Pelocypris) zilchi. Wesselingh et al. (2006b) re-examined mollusc material studied by Roxo (1937) from Aquidabã and described: Ampullariidae sp. 2, Anodontites cf. trapesialus, Diplodon cf. longulus, Eupera sp., Littoridina? sp., Pyrgophorus? sp. and Sheppardiconcha septemcincta. Additional findings (including vertebrates and plants) are mentioned by Del’ Arco et al. (1977, cum Lit.).
Interpretation: The measured litho-logs document a laterally highly variable depositional environment. The basal, sandy–silty layers (1–6/1 = 1/2) possibly represent crevasse splay deposits, which are followed by a series of crevasse splay and/or crevasse channel sediments (7–14/1 respectively 2–5/2). Alternatively, these layers could represent channel deposits of an avulsive river arm. In section 2 a subordinate abandoned channel fill (floodplain pond; 6–7/2) is developed. Afterwards (15–23/1 and 8–16/2) the influx of crevassing respectively the active channel successively decreased and led to the formation of a floodplain lake, which was only influenced by flash floods in more proximal settings (section 2; e.g., 12/2). Aquatic faunas (bivalves, ostracods, fishes) and semi-aquatic crocodilians inhabited that lake, which was surrounded by a densely vegetated backswamp. Pedogenic processes are evident in layers 21/1 and 15/2. These beds display a mottled appearance and contain (21/1) layers of crumb peds (clast-supported layers of rounded but not interlocked tiny mud balls), which indicate bioturbation due to invertebrate activity (Retallack, 2001). Finally, the lake became replaced by a swampy environment (23/1 and 16/2).
4.4. Remanso
Location: 27.4 km NE Eirunepé (S 06°31’22.0”/W 069°35’42.8”; altitude: ∼105 m), left cutbank of the Juruá River (Fig. 8).
Fig. 8.

Lithological section of Remanso (a), outcrop photograph and interpretation of depositional environment (b).
Description: At the base >2 m of slightly southwards inclined (<10°) rippled bedded silty fine sand (layer 1) are followed by 2.7 m thick violet-brown (2) and yellowish (3) mottled paleosols with calcareous nodules (between 2 and 3 a pedogenically overprinted clay plug is intercalated). Layers 1–3 are cut by a channel, whose fill starts (4) with >1.5 m thick, massive–poorly laminated silty clay with gastropods, rare bivalves and plant (leaves) remains. Bed 4 grades into a 0.8 m thick, brownish-grey, blocky structured silty clay (5) with a calcareous crust on its top. It is overlain by 2.4 m of violet-brown, mottled clay (6) with intercalated layers of calcareous nodules. Strata 1–3 as well as the channel fill (4–6) are topped by a 0.4 m thick alternation of laminated silty clay and laminated or rippled bedded fine–medium sand with a high amount of plant detritus (also wood remains) and abundant vertebrates (turtles; 7). The badly exposed, ∼7.2 m thick beds up-section (8) consist of cross and ripple bedded fine sand with intercalations of clayey lenses (∼2 m lateral extension, ∼20 cm thickness), which are formed by clay intraclasts (mud pebbles, ∼1 cm diameter). Repeatedly coaly layers are intercalated and rarely vertebrate (crocodiles) remains were found in the intraclast conglomerate. Layer 9 represents a 1.2 m thick alternation of horizontally or ripple bedded fine–medium sand, coaly (especially in the lower part) and clayey layers. Clay contents increases and the bedding becomes more rhythmical upwards. The topmost 1.2 m thick layer 10 consists of ripple bedded silty fine sand and silty clay alternations, which form cm-thick, fining-upward couplets. Oxidised plant (wood) remains are frequent.
Microfossil contents: Sample RE4 yielded only a few valves of Cyprideis spp. as well as some fish and gastropod remains.
Interpretation: Layer 1 could represent a crevasse splay or levee deposit, which is overlain by pedogenically altered floodplain fines (2–3) with intercalated minor channel fills. These overbank deposits are cut by a channel (?20 m wide and 5 m deep), which became filled by pelitic, afterwards pedogenically overprinted sediments. The rare (a matter of preservation?) mollusc and ostracod record indicates at least at the beginning (4) aquatic life in the abandoned channel. Above, a crevasse splay (7) points to rising proximity of the active channel. Large wood remains and vertebrate fragments were accumulated by this splay. The sandy succession up-section (8) can only be attributed to a sandy bedform sensu Miall (1996) due to insufficient outcrop conditions. Maybe it is formed by vertically aggrading dune fields. Intercalated shallow scours, filled up with mud balls indicate deposition of reworked material and variations of the flow regime. The more or less rhythmically stratified, sandy-pelitic alternations at the top of Remanso (9–10) mark a shift towards overbank environment and might represent a bar-top assemblage.
4.5. Torre da Lua
Location: 17.5 km SE Eirunepé (S 06°49’23”/W 069°47’04”, altitude: ∼117 m), left cutbank of the Tarauacá River (Fig. 9).
Fig. 9.

Lithological section of Torre da Lua and interpretation of depositional environment (CR = crevasse channel).
Description: The basal 1.7 m (layers 1–3) consist of massive or laminated pelite- and fine sand with rare bivalve remains. They are topped by 0.2 m of reddish-brown, massive clay (4). The sandy–silty layers 5–9 display a general coarsening-upward trend and contain only rarely bivalve fragments. Up-section (10), a 0.25 m thick intraclast conglomerate (mudstone pebbles) with abundant mollusc (e.g., Ampullariidae?, Diplodon? sp.) and vertebrate remains (crocodiles, turtles) follows. This stratum is overlain by thin sand and clay layers (11–17) with abundant mollusc remains (12, 14, 16, lower part of 17). Layer 14 is rich in coal fragments (wood, leaves) at its top. The 2.57 m thick succession above (18–23) starts with thin, ripple bedded fine sand beds (18–19), followed by massive silty/sandy clay with coaly fragments (20) and cm-thick alternations of fine sandy silt and silty fine sand (21; forming couplets of small-scale fining-upward cycles). Beds 22 and 23 comprise alternations of massive, fine sandy silt and ripple bedded, silty fine sand. Coaly remains were documented at the top of layer 22. The topmost bed 24 consists of ∼10 cm-thick alternations of indistinctly ripple bedded silty fine sand and massive or poorly laminated pelite. The upper part (∼1.5 m) is mottled and primary sedimentary structures are obscured due to pedogenic processes.
Microfossil contents: Samples TO1 and TO6 yielded several ostracod (Cyprideis spp., Cytheridella sp.) and some fish remains. TO10 is rich and TO12 extremely rich in ostracods (Cypria sp., Cyprideis spp., Cytheridella, darwinulids). These samples contain several fish remains and in TO12 rare characean gyrogonites were found additionally. The samples TO20 and TO 23 delivered only rare ostracod (Cypria sp., Cyprideis spp.) and fish fossils.
Earlier reports of fossil material and facies: Ramos (2006) mentioned the following ostracod taxa: C. aqualica, C. graciosa, C. lacrimata, C. longispina, C. pebasae, C. purperae, Cytheridella sp., Darwinula fragilis as well as additional vertebrate and crab findings.
Interpretation: The lowermost sandy-pelitic layers (1–4) might be deposited in a floodplain pond/lake, which was influenced by crevasse splays. The coarsening-upward trend of layers 5–9 hints to a progressive influx of a crevasse channel (layer 10). Intraclasts and disarticulated vertebrate fragments form a lag deposit. It remains unclear if the ostracods of this layer are allochthonous or if they are incorporated in that layer due to subaquatic inflow of the crevasse splay (crevasse delta) into a floodplain pond/lake. Up-section (11–24), plentiful ostracod and mollusc faunas (especially in the lower part: 11–17; compare Ramos, 2006) indicate the return to an aquatic environment and the abandonment of the crevasse channel. However, this overbank environment is frequently influenced by crevasse splays during floods. Small-scale fining-upward alternations of ripple bedded sand and pelite document individual flooding events or surges.
4.6. Barro Branco
Location: 22.1 km SSE Eirunepé (S 06°52’18.3”/W 069°47’05.1”; altitude: ∼120 m), left cutbank of the Tarauacá River (Fig. 10).
Fig. 10.
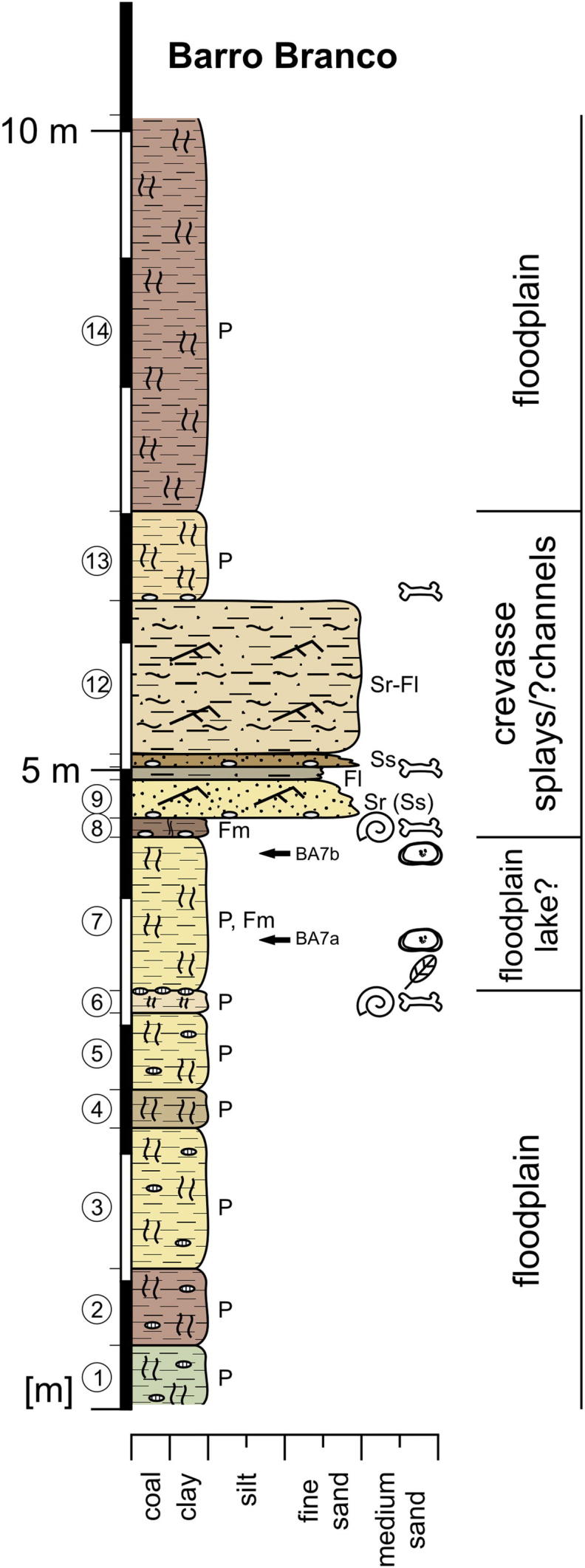
Lithological section of Barro Branco and interpretation of depositional environment.
Description: Layers 1–6 represent a ∼3.2 m thick succession of greenish (1), reddish (2), yellowish (3, 5, 6) or violet grey (4) coloured, mottled, frequently calcareous nodules-bearing paleosols. In layer 6 vertebrate remains (crocodile teeth) and gastropods were found. Above follows a 1.1 m thick, indistinctly bedded clay to fine sand (7), which contains in its lower half coaly plant fragments and becomes towards the top and laterally progressively mottled (paleosol). The 0.65 m thick sediment column up-section (8–11) is composed of massive clay (8), ripple bedded fine sand (9), laminated silty fine sand to silt (10) and fine sand (11). Each of these layers displays a slightly erosive base and an internal fining-upward trend. The beds 8, 9 and 11 each start with a basal layer of concretionary mud clasts in which (layer 8 and 11) abundant vertebrate remains were found. Gastropods (Ampullariidae?) were only observed at the base of layer 8. Layer 12 is formed by 1.2 m thick, ripple bedded fine sand with cm-thick, laminated silty clay interlayers. Sedimentary structures are obscured due to alteration. Up-section a 0.7 m thick, orange-grey, mottled paleosol with mud clasts and rare vertebrate remains at its base (13) as well as a >1.5 m thick, violet-purple coloured, mottled paleosol are developed (14).
Microfossil contents: The samples BA7a and BA7b contained several ostracod valves (mainly Cyprideis spp., some specimens of Cypria sp., Cytheridella sp. and darwinulids).
Interpretation: The basal layers (1–6) represent a succession of paleosols, which could be seriously altered floodplain deposits. Due to the occurrence of ostracods, probably bed 7 was deposited in a floodplain lake environment. Afterwards, a series of crevasse splay (maybe partly also crevasse channel) deposits follows (8–13), which is topped by pedogenically overprinted floodplain fines (14).
5. Discussion
5.1. Sedimentology
5.1.1. Depositional environment
The sedimentary record of the investigated outcrops documents vertically as well as laterally highly variable fine-grained clastic successions (predominantly clay–fine sand, along with fine–medium sand). Based on the lithofacies and, as far as possible, lateral observations, these sediments represent fluvial deposition within active channels as well as in overbank environments.
Macroforms, generated within the active channel include lateral accretion deposits (sandy point bars; Fig. 2, Fig. 4) and undifferentiated sandy bedforms (stacked dune fields?; Fig. 8), which are probably partly intersected by chute channels or are overlain by overbank fines.
However, the bulk of the sediments belongs to overbank environments. These comprise pelite–sand-alternations of levee, crevasse splay and crevasse channel deposits. Frequently, a lag of mud pebbles and vertebrate fragments is found at the base of crevasse channels/splays. In part, some splays may have entered floodplain ponds/lakes via crevasse deltas (e.g., Fig. 6, Fig. 9). Abandoned, fine-grained channel fills (mud plugs) were observed (Fig. 8). Decimetre-thick, clay–silt beds with rich ostracod and mollusc faunas indicate the presence of short-lived, shallow ponds or lakes within the floodplain (floodbasin; e.g., Fig. 7b). We found no evidence for long-lasting and deep lakes. Up to 1 m thick, lignitic beds (Fig. 6) point to partly swampy, poorly-drained conditions and a densely vegetated floodplain, which is, however, influenced by high clastic input. Intensively mottled paleosols with root casts and calcareous nodules are ubiquitous (e.g., Fig. 5a, c and 10). Differences in colour and occasional calcic horizons may reflect diverging water-logging due to e.g., local topography and/or seasonal fluctuations in discharge or climate (Kaandorp et al., 2006, Latrubesse et al., 2010). Nevertheless, diagenesis may have altered significantly primary features and more specific investigations are required (Retallack, 2001).
5.1.2. Considerations about the fluvial style
We are aware that restricted outcrop conditions and the limited areal and stratigraphical extent (∼30 m stratigraphical thickness) of the current investigation hamper conclusions about the nature of fluvial style. Reconstructions of fluvial geomorphology, based solely on lithofacies assemblages and fragmentary known fluvial elements, remain to some degree tentative (Miall, 1996, Bridge, 2003).
However, it seems obvious that we are dealing with a suspended load river system due to the high amount of overbank fines, probably within a tropical to subtropical wet–dry climate (Kaandorp et al., 2005, Latrubesse et al., 2010). We observed: (a) a large proportion of overbank deposits, (b) laterally extensive, heterogeneous, fine-grained avulsion deposits (including crevasse splays), (c) predominantly sandy channel deposits, which change laterally and vertically rapidly into overbank deposits as well as (d) lacustrine and coaly deposits. Some sandy point bars (of secondary channels?) indicate subordinate lateral sediment accretion. These features are not exclusive to anastomosing river deposits, however, they largely coincide with the “standard” model of an anastomosing river sensu Miall (1996) as well as with characteristics of such a system as reviewed by Makaske (2001; compare also Smith, 1983). The general setting (subsiding foreland basin, extensive sediment input from the Andes, possibly incompletely framed by tectonically active basement highs towards the east) as well as a wet–dry seasonal climate (seasonal floods, channel banks stabilized by vegetation) would provide the conditions for anastomosing river systems (Smith, 1983, Nanson and Knighton, 1996, Makaske, 2001, Latrubesse et al., 2010).
Facies analyses on exposures of the uppermost Solimões Fm. (Late Miocene) in the state of Acre (SW Brazil) led to similar assumptions (Latrubesse et al., 1997, Latrubesse et al., 2007, Latrubesse et al., 2010; see also Westaway, 2006). These authors proposed a megafan system, which originates in the Andes and encompass a complex mosaic of avulsive rivers and associated wetlands (including abandoned channels, floodplain lakes, floodbasin deltas, backswamps and well-drained floodplains; see also Wilkinson et al., 2006, Wilkinson et al., 2010). This system is suggested to reach to the east at least as far as 67° W (∼”Iquitos arch” sensu Del’ Arco et al., 1977) and probably also beyond (“Purus arch”; Latrubesse et al., 1997, Latrubesse et al., 2010; Fig. 1). According to that, the sections around Eirunepé were located in a more distal position, which could be the reason for a more pronounced anastomosing-anabranching pattern due to a lower gradient (compare also interpretations of the Late Miocene Tariquia Formation, southern Bolivia; Uba et al., 2005, Uba et al., 2006. However, Latrubesse et al., 2007, Latrubesse et al., 2010 compared the depositional environment of the Late Miocene Solimões Fm. in Acre with i.e., the Quaternary megafans of the Chaco plain and the Pantanal wetland (Iriondo, 1993, Horton and DeCelles, 2001, Assine, 2005, Latrubesse et al., 2005).
5.1.3. Palaeoecological remarks
The mollusc fauna, recorded from floodplain pond/lake deposits, is dominated by sphaeriids and the pachychilid Sheppardiconcha (Fig. 7b) and indicates freshwater conditions within a fluvio-lacustrine environment (Wesselingh et al., 2006b, Wesselingh and Ramos, 2010). Typical freshwater ostracods (darwinulids, ilyocyprinids, Cypria, Cytheridella) are associated with diverse species of Cyprideis. This genus is a holoeuryhaline taxon, able to cope with fluctuating salinities, aberrant water chemistries, variable temperatures and oxygenation (e.g., Meisch, 2000, Keyser, 2005). As it could adapt to freshwater conditions (e.g., Lake Tanganyika; Wouters and Martens, 2001) and co-occurs here with exclusively freshwater taxa, there is no constraint to conclude a brackish water environment (Ramos, 2006). Preliminary stable isotope data (δ18O, δ13C) measured on Cyprideis valves from Aquidabã, Morada Nova and Torre da Lua, yielded very negative values (δ18O: −5.7 to −9.7‰, δ13C: −10.3 to −12.5‰), which exclude any marine influx. Similar results are presented by Vonhof et al., 2003, Kaandorp et al., 2006, Wesselingh et al., 2006c; compare Wesselingh, 2008 and Wesselingh and Ramos, 2010).
5.2. Biostratigraphy
There is still intensive debate on definition and timing of palynological biozonation concepts (e.g., Hoorn, 1993, Hoorn, 1994, Latrubesse et al., 2007, Latrubesse et al., 2010, Jaramillo et al., 2010, Silva-Caminha et al., 2010). However, palynostratigraphy remains a cornerstone for correlation across Amazonia up to now and provides at least a rough stratigraphical framework. Subsequent ostracod and mollusc zonations of Muñoz-Torres et al., 1998, Muñoz-Torres et al., 2006 and Wesselingh et al. (2006a) are highly linked to the concept established by Hoorn (1993).
Based on the presence of some index taxa (C. annulatus, E. spinosus, G. magnaclavata) a Late Miocene age (probably Asteraceae zone sensu Lorente, 1986) is proposed by Paz et al. (in press) for the outcrops Morada Nova and Torre da Lua recently.
The vertebrate fauna from Torre da Lua (Ramos, 2006) is related to the so-called “Acre fauna”, which is one of the best documented faunas of northern South America (Cozzuol, 2006, Latrubesse et al., 2007, Negri et al., 2010). The Acre vertebrate assemblage displays significant affinities with the “Mesopotamian” faunas of Argentina and Uruguay as well as with the Urumaco assemblages (Venezuela) and is dated to the Late Miocene (Huayquerian South American Land Mammal Age; Cozzuol, 2006, Latrubesse et al., 2007, Latrubesse et al., 2010, Negri et al., 2010).
Knowledge about the aquatic “Juruá fauna” is still poor and has to be extended by later works (Wesselingh and Ramos, 2010; see Roxo, 1937, Wesselingh et al., 2006b, Wesselingh, 2008, and outcrop descriptions herein). From a stratigraphical point of view the mollusc faunas of Aquidabã and Torre da Lua are suggested by Wesselingh et al. (2006b) to be of Late Miocene origin and post-date the “Pebas fauna” (compare Celestino and Ramos, 2007, Wesselingh, 2008, Wesselingh and Ramos, 2010, which suggested a late Middle Miocene age based on the ostracod record). A Late Miocene age seems to be in agreement with the lack of typical endemic Pebasian molluscs (Wesselingh and Ramos, 2010).
The recorded Cyprideis-species from Aquidabã, Morada Nova and Torre da Lua (Ramos, 2006, Celestino and Ramos, 2007, Wesselingh and Ramos, 2010) have generally long ranges according to Muñoz-Torres et al., 1998, Muñoz-Torres et al., 2006; see also Whatley et al., 1998). The occurrence of C. lacrimata and C. pebasae might be indicative for an age not younger than the (early) Late Miocene. Nonetheless, due to some contradictions (e.g., C. lacrimata is supposed to appear first after the last appearance of C. longispina but both are co-occurring at Morada Nova and Torre da Lua) and the lack of key taxa, the current ostracod zonation needs some adjustment and is not further stressed here.
However, to date, a Late Miocene age seems most plausible for all outcrops described herein by considering the palaeontological record. Moreover, this fits well to the palaeogeographic context of this region as proposed by Latrubesse et al., 1997, Latrubesse et al., 2007, Latrubesse et al., 2010 and Cozzuol (2006).
6. Conclusions
Sedimentological observations derived from outcrops around Eirunepé (south-western part of the state of Amazonia) document various subenvironments of a fluvial system (upper part of the Solimões Formation, Late Miocene). Beside sandy channel deposits, the main part comprises overbank sediments of levees, crevasse splays/channels/deltas, abandoned channels, backswamps and floodplain paleosols. Lacustrine environments consist only of short-lived floodplain ponds/lakes. Preliminary palaeontological and geochemical results indicate exclusively freshwater conditions. Based on the lithofacies and the general geological setting, this system can be possibly related to an anastomosing river style. There is not any indication for a long-lived lake (“Lake Pebas”) or any marine influx in this region during the Late Miocene. Our data confirm the continental sedimentation model established by Latrubesse et al., 1997, Latrubesse et al., 2007, Latrubesse et al., 2010; compare also Cozzuol, 2006, Westaway, 2006).
Acknowledgements
This study was financially supported by the Austrian Science Fund project P21748-N21. We acknowledge the Conselho Nacional de Desenvolvimento Científico e Tecnológico/Ministério da Ciência e Tecnologia (CNPq/MCT; process number EXC 010389/2009-1), the Departamento Nacional de Produção Mineral (DNPM/Manaus, especially Gert Woeltje) and the Companhia de Pesquisas de Recursos Minerais (CPRM/Manaus; especially Marco Oliveira) for sampling and shipping licences as well as for access to unpublished reports. Earlier sampling campaigns (M.I.R.) were financially supported by the CNPq (process number 475475/2004-6). The authors are grateful to Marco Caporaletti (University of Graz) for the stable isotope analyses. Special thanks go to Norbert Winkler (Joanneum/Graz) and Miguel Sombra (Eirunepé) for their assistance during fieldwork. We thank two anonymous reviewers for their critical comments, which improved the manuscript.
Contributor Information
Martin Gross, Email: martin.gross@museum-joanneum.at.
Werner E. Piller, Email: werner.piller@uni-graz.at.
Maria Ines Ramos, Email: mramos@museu-goeldi.br.
Jackson Douglas da Silva Paz, Email: jackdspaz@yahoo.com.br.
References
- Assine M.L. River avulsion on the Taquari megafan, Pantanal wetland. Brazil Geomorphology. 2005;70:357–371. [Google Scholar]
- Barata C.B., Caputo M.V. Geologia do petróleo da bacia do Solimões. O “Estado da arte”. 4° Congresso Brasileiro de Pesquisa e Desenvolvimento em Petróleo e Gás. PDPETRO. 2007;4 1.1.0147–1–10. [Google Scholar]
- Bridge J.S. Blackwell Publishing; Oxford: 2003. Rivers and Floodplains: Forms, Processes, and Sedimentary Record. 491 pp. [Google Scholar]
- Campbell K.E., Jr., Frailey C.D., Romero-Pittman L. The Pan-Amazonian Ucayali Peneplain, late Neogene sedimentation in Amazonia, and the birth of the modern Amazon River system. Palaeogeography, Palaeoclimatology. Palaeoecology. 2006;239:166–219. [Google Scholar]
- Caputo M.V. Solimões megashear: intraplate tectonics in northwestern Brazil. Geology. 1991;19:246–249. [Google Scholar]
- Celestino E.A., Ramos M.I.F. Associação Brasileira de Oceanografia; Florianópolis: 2007. Sistemática de ostracodes e reconstruçã paleoambiental no Neógeno da bacia do Solimões, Formação Solimões, sudoeste do estada do Amazonas. XII Congresso Latino-Americano de Ciências do Mar. 1–3. [Google Scholar]
- Cozzuol M.A. The Acre vertebrate fauna: age, diversity, and geography. Journal of South American Earth Science. 2006;21:185–203. [Google Scholar]
- Del’ Arco J.O., Santos R.O.B., Rivetti M., Olivera Alves E.D., Fernandes C.A.C., Silva L.L. Folha SB. 19 Juruá. I–Geologia. Projeto Radambrasil. Levantamento de Recursos Naturais. 1977;15:19–88. [Google Scholar]
- Figueiredo J., Hoorn C., van der Ven P., Soares E. Late Miocene onset of the Amazon River and the Amazon deep-sea fan: evidence from the Foz do Amazonas basin. Geology. 2009;37:619–622. [Google Scholar]
- Gingras M.J., Räsänen M.E., Pemberton S.G., Romero L.P. Ichnology and sedimentology reveal depositional characteristics of bay-margin parasequences in the Miocene Amazonian foreland basin. Journal of Sedimentary Research. 2002;72:871–883. [Google Scholar]
- Gorthner A. What is an ancient lake? Archiv für Hydrobiologie. Beiheft. 1994;44:97–100. [Google Scholar]
- Haffer J. Hypotheses to explain the origin of species in Amazonia. Brazilian Journal of Biology. Supplement. 2008;68:917–947. doi: 10.1590/s1519-69842008000500003. [DOI] [PubMed] [Google Scholar]
- Hoorn C. Marine incursions and the influence of Andean tectonics on the Miocene depositional history of northwestern Amazonia: results of a palynostratigraphic study. Palaeogeography, Palaeoclimatology, Palaeoecology. 1993;105:267–309. [Google Scholar]
- Hoorn C. An environmental reconstruction of the palaeo-Amazon River system (Middle–Late Miocene, NW Amazonia). Palaeogeography, Palaeoclimatology. Palaeoecology. 1994;112:187–238. [Google Scholar]
- Hoorn C. Scientific American; 2006. The birth of the mighty Amazon. May 2006, 52–59. [DOI] [PubMed] [Google Scholar]
- Hoorn C., Vonhof H. Neogene Amazonia: Introduction to the special issue. Journal of South American Earth Sciences. 2006;21:1–4. [Google Scholar]
- Hoorn C., Wesselingh F.P. Wiley-Blackwell; Oxford: 2010. Amazonia, Landscape and Species Evolution: a Look into the Past. 464 pp. [Google Scholar]
- Hoorn C., Wesselingh F.P., ter Steege H., Bermudez M.A., Mora A., Sevink J., Sanmartín I., Sanchez-Meseguer A., Anderson C.L., Figueiredo J.P., Jaramillo C., Riff D., Negri F.R., Hooghiemstra H., Lundberg J., Stadler T., Särkinen T., Antonelli A. Amazonia through time: andean uplift, climate change, landscape evolution, and Biodiversity. Science. 2010;330:927–931. doi: 10.1126/science.1194585. [DOI] [PubMed] [Google Scholar]
- Horton B.K., DeCelles P.G. Modern and ancient fluvial megafans in the foreland basin system of the central Andes, southern Bolivia: implications for drainage network evolution in fold-thrust belts. Basin Research. 2001;13:43–63. [Google Scholar]
- Hovikoski J., Gingras M., Räsänen M., Rebata L.A., Guerrero J., Ranzi A., Melo J., Romero L., Nuñez del Prado H., Jaimes F., Lopez S. The nature of Miocene Amazonian epicontinental embayment: high-frequency shifts of the low-gradient coastline. Geological Society of America Bulletin. 2007;119:1506–1520. [Google Scholar]
- Hovikoski J., Wesselingh F.P., Räsänen M., Gingras M., Vonhof H.B. Marine Influence in Amazonia: evidence from the geological record. In: Hoorn C., Wesselingh F.P., editors. Amazonia, Landscape and Species Evolution: a Look into the Past. Wiley-Blackwell; Oxford: 2010. pp. 143–161. [Google Scholar]
- Iriondo M. Geomorphology and late Quaternary of the Chaco (South America) Geomorphology. 1993;7:289–303. [Google Scholar]
- Jaramillo C., Hoorn C., Silva S.A.F., Leite F., Herrera F., Quiroz L., Dino R., Antonioli L. The origin of the modern Amazon rainforest: implications of the palynological and palaeobotanical record. In: Hoorn C., Wesselingh F.P., editors. Amazonia, Landscape and Species Evolution: a Look into the Past. Wiley-Blackwell; Oxford: 2010. pp. 317–334. [Google Scholar]
- Kaandorp R.J.G., Vonhof H.B., Wesselingh F.P., Romero Pittman L., Kroon D., Van Hinte J.E. Seasonal Amazonian rainfall variation in the Miocene climate Optimum. Palaeogeography, Palaeoclimatology. Palaeoecology. 2005;221:1–6. [Google Scholar]
- Kaandorp R.J.G., Wesselingh F.P., Vonhof H.B. Ecological implications from geochemical records of Miocene Western Amazonian bivalves. Journal of South American Earth Sciences. 2006;21:54–74. [Google Scholar]
- Keyser D. Histological peculiarities of the noding process in Cyprideis torosa (Jones) (Crustacea, Ostracoda) Hydrobiologia. 2005;538:95–106. [Google Scholar]
- Latrubesse E.M., Bocquentin J., Santos J.C.R., Ramonell C.G. Paleoenvironmental model for the late Cenozoic of southwestern Amazonia: paleontology and geology. Acta Amazonica. 1997;27:103–118. [Google Scholar]
- Latrubesse E.M., Stevaux J.C., Sinha R. Tropical rivers. Geomorphology. 2005;70:187–206. [Google Scholar]
- Latrubesse E., Silva S.A.F., Cozzuol M., Absy M.L. Late Miocene continental sedimentation in the southwestern Amazonia and its regional significance: Biotic and geological evidence. Journal of South American Earth Science. 2007;23:61–80. [Google Scholar]
- Latrubesse E.M., Cozzuol M., Silva-Caminha S.A.F., Rigsby C.A., Absy M.L., Jaramillo C. The late Miocene paleogeography of the Amazon basin and the evolution of the Amazon River system. Earth-Science Reviews. 2010;99:99–124. [Google Scholar]
- Loczy L. Paleogeography and history of the geological development of the Amazonas basin. Jahrbuch der Geologischen Bundesanstalt Wien. 1963;106:449–502. [Google Scholar]
- Lorente M.A. Palynology and Palynofacies of the upper Tertiary in Venezuela. Dissertationes Botanicae. 1986;99:1–225. [Google Scholar]
- Lundberg J.G., Marshall L.G., Guerrero J., Horton B., Malabarba M.S.L., Wesselingh F. The stage for Neotropical fish diversification: a history of tropical south American rivers. In: Malabarba L.R., Reis R.E., Vari R.P., Lucena Z.M., Lucena CA.S., editors. Phylogeny and Classification of Neotropical Fishes. Edipucrs; Porto Alegre: 1998. pp. 13–48. [Google Scholar]
- Maia R.G.N., Godoy H.K., Yamaguti H.S., Moura P.A., Costa F.S.F., Holanda M.A., Costa J.A. Companhia de Pesquisa de Recursos Minerais-Departamento Nacional da Produçáo Mineral; Manaus: 1977. Projeto Carvão No Alto Solimões. Relatório Final. 142 pp. [Google Scholar]
- Makaske B. Anastomosing rivers: a review of their classification, origin and sedimentary products. Earth-Science Reviews. 2001;53:149–196. [Google Scholar]
- Meisch C. Freshwater Ostracoda of western and central Europe. In: Schwoerbel J., Zwick P., editors. Süßwasserfauna von Mitteleuropa. Spektrum-Verlag; Heidelberg-Berlin: 2000. p. 522. 8/3. [Google Scholar]
- Miall A.D. Springer-Verlag; Berlin-Heidelberg-New York: 1996. The Geology of Fluvial Deposits. , 582 pp. [Google Scholar]
- Miura K. Possibilidades Petrolíferas da Bacia do Acre. Anais do XXVI Congresso Brasileiro de Geologia. 1972;3:15–20. [Google Scholar]
- Muñoz-Torres F., Whatley R.C., Van Harten D. The endemic non-marine Miocene ostracod fauna of the Upper Amazon Basin. Revista Españiola de Micropaleontología. 1998;30:89–105. [Google Scholar]
- Muñoz-Torres F.A., Whatley R.C., Van Harten D. Miocene ostracod (Crustacea) biostratigraphy of the upper Amazon Basin and evolution of the genus Cyprideis. Journal of South American Earth Sciences. 2006;21:75–86. [Google Scholar]
- Nanson G.C., Knighton A.D. Anabranching rivers: their cause, character and classification. Earth Surface Processes and Landforms. 1996;21:217–239. [Google Scholar]
- Negri F.R., Bocquentin-Villanueva J., Ferigolo J., Antoine P.-O. A review of Tertiary mammal faunas and birds from western Amazonia. In: Hoorn C., Wesselingh F.P., editors. Wiley-Blackwell; Oxford: 2010. pp. 245–258. (Amazonia, Landscape and Species Evolution: a Look into the Past). [Google Scholar]
- Paz, J.D.S., Ramos, M.I.F., Moraes-Santos, H.M., Silva, S., in press. Depósitos fluviais da formação Solimões, Bacia do Solimões, Eirunepé (AM), Brasil. Latin American Journal of Sedimentology and Basin Analysis.
- Ramos M.I.F. Ostracods from the Neogene Solimões formation (Amazonas, Brazil) Journal of South American Earth Sciences. 2006;21:87–95. [Google Scholar]
- Räsänen M.E., Linna A.M., Santos J.C.R., Negri F.R. Late Miocene tidal deposits in the Amazonian foreland basin. Science. 1995;269:386–390. doi: 10.1126/science.269.5222.386. [DOI] [PubMed] [Google Scholar]
- Retallack G.J. Blackwell; Oxford: 2001. Soils of the Past. An Introduction to Paleopedology. 404 pp. [Google Scholar]
- Roddaz M., Baby P., Brusset S., Hermoza W., Darrozes J.M. Forebulge dynamics and environmental control in Western Amazonia: the case study of the Arch of Iquitos (Peru) Tectonophysics. 2005;399:87–108. [Google Scholar]
- Rossetti D.F., Mann de Toledo P. Environmental changes in Amazonia as evidenced by geological and paleontological data. Revista Brasiliera de Ornitologia. 2007;15:175–188. [Google Scholar]
- Roxo M.G. de Oliveira. Fósseis Pliocênicos do Rio luruá, Estado do Amazonas. Notas Preliminares e Estudos. Serviço Géológico e Mineralógico do Brasil. 1937;9:4–13. [Google Scholar]
- Shephard G.E., Müller R.D., Liu L., Gurnis M. Miocene drainage reversal of the Amazon River driven by plate–mantle interaction. Nature Geoscience. 2010;3:870–875. [Google Scholar]
- Silva-Caminha S.A.F., Jaramillo C.A., Absy M.L. Neogene palynology of the Solimões basin, Brazilian Amazonia. Palaeontographica B. 2010;284:13–79. [Google Scholar]
- Smith D.G. Anastomosed fluvial deposits: modern examples from Western Canada. Special Publications. International Association of Sedimentologists. 1983;6:155–168. [Google Scholar]
- Uba C.E., Heubeck C., Hulka C. Facies analysis and basin architecture of the Neogene Subandean synorogenic wedge, southern Bolivia. Sedimentary Geology. 2005;180:91–123. [Google Scholar]
- Uba C.E., Heubeck C., Hulka C. Evolution of the late Cenozoic Chaco foreland basin, southern Bolivia. Basin Research. 2006;18:145–170. [Google Scholar]
- Uba C.E., Hasler C.-A., Buatois L.A., Schmitt A.K., Plessen B. Isotopic, paleontologic, and ichnologic evidence for late Miocene pulses of marine incursions in the central Andes. Geology. 2009;37:827–830. [Google Scholar]
- Vonhof H.B., Wesselingh F.P., Kaandorp R.J.G., Davies G.R., Van Hinte J.E., Guerrero J., Räsänen M., Romero-Pittman L., Ranzi A. Paleogeography of Miocene Western Amazonia: Isotopic composition of molluscan shells constrains the influence of marine incursions. Geological Society of America Bulletin. 2003;115:983–993. [Google Scholar]
- Wesselingh F.P. Miocene long-lives lake Pebas as a stage of mollusc radiations, with implications for landscape evolution in western Amazonia. Scripta Geologica. 2006;133:1–17. [Google Scholar]
- Wesselingh F.P. Molluscan radiations and landscape evolution in Miocene Amazonia. Annales Universitates Turkuensis. Biologica-Geographica-Geologica. 2008;232:1–41. [Google Scholar]
- Wesselingh F.P., Ramos M.I.F. Amazonian aquatic invertebrate faunas (Mollusca, Ostracoda) and their development over the past 30 million years. In: Hoorn C., Wesselingh F.P., editors. Amazonia, Landscape and Species Evolution: a Look into the Past. Wiley-Blackwell; Oxford: 2010. pp. 302–316. [Google Scholar]
- Wesselingh F.P., Salo J.A. A Miocene perspective on the evolution of the Amazonian biota. Scripta Geologica. 2006;133:439–458. [Google Scholar]
- Wesselingh F.P., Räsänen M.E., Irion G., Vonhof H.B., Kaandorp R., Renema W., Romero Pittman L., Gingras M. Lake Pebas: a palaeoecological reconstruction of a Miocene, long-lived lake complex in western Amazonia. Cainozoic Research. 2002;1:35–81. [Google Scholar]
- Wesselingh F.P., Hoorn M.C., Guerrero J., Räsänen M.E., Romero Pittman L., Salo J. The stratigraphy and regional structure of Miocene deposits in western Amazonia (Peru, Columbia and Brazil), with implications for late Neogene landscape evolution. Scripta Geologica. 2006;133:291–322. [Google Scholar]
- Wesselingh F.P., Ranzi A., Räsänen M.E. Miocene freshwater Mollusca from western Brazilian Amazonia. Scripta Geologica. 2006;133:419–437. [Google Scholar]
- Wesselingh F.P., Kaandorp R.J.G., Vonhof H.B., Räsänen M.E., Renema W., Gingras M. The nature of aquatic landscapes in the Miocene of western Amazonia: an integrated palaeontological and geochemical approach. Scripta Geologica. 2006;133:363–393. [Google Scholar]
- Westaway R. Late Cenozoic sedimentary sequences in Acre state, southwestern Amazonia: fluvial or tidal? Deductions from the IGCP 449 fieldtrip. Journal of South American Earth Sciences. 2006;21:120–134. [Google Scholar]
- Whatley R.C., Muñoz-Torres F., Van Harten D. The Ostracoda of an isolated Neogene saline lake in the western Amazon Basin, Peru. Bulletin du Centres de Recherches Elf Exploration-Production. Memoires. 1998;20:231–245. [Google Scholar]
- Wilkinson M.J., Marshall L.G., Lundberg J.G. River behavior on megafans and potential influences on diversification and distribution of aquatic organisms. Journal of South American Earth Sciences. 2006;21:151–172. [Google Scholar]
- Wilkinson M.J., Marshall L.G., Lundberg J.G., Kreslavsky M.H. Megafan environments in northern South America and their impact on Amazon Neogene aquatic ecosystems. In: Hoorn C., Wesselingh F.P., editors. Amazonia, Landscape and Species Evolution: A Look into the Past. Wiley-Blackwell; Oxford: 2010. pp. 162–184. [Google Scholar]
- Wouters K., Martens K. On the Cyprideis species flock (Crustacea, Ostracoda) in Lake Tanganyika, with the description of four new species. Hydrobiologia. 2001;450:111–127. [Google Scholar]