

Abstract
Modest protocols of repetitive acute intermittent hypoxia (rAIH) enhance motor function in patients with chronic incomplete spinal injury. Since chronic intermittent hypoxia (CIH) elicits neuroinflammation, there is potential for rAIH to have similar effects. Thus, we tested the hypothesis that rAIH has minimal impact on microglial inflammatory gene expression, but up-regulates key neurotrophic factor expression in a CNS region-specific manner. Using real time PCR, we evaluated mRNA levels of inflammatory and neurotrophic factors in immunomagnetically-isolated microglia from rat frontal cortex, brainstem and upper and lower cervical spinal cord following rAIH (ten, 5-minute episodes, thrice weekly, 4 weeks). In agreement with our hypothesis, rAIH had no significant impact on microglial inflammatory gene expression in any region studied. On the other hand, neurotrophic factor expression was altered in a gene-and region-specific pattern. These results have important implications for the safety of rAIH as a potential therapy to enhance neuroplasticity and motor function in patients with spinal injury or other neurologic disorders.
INTRODUCTION
Intermittent hypoxia (IH) can have both detrimental and beneficial effects, depending on the frequency of cycles per day, the severity of hypoxic episodes and the duration of exposure (i.e. the “dose” of IH; (Navarrete-Opazo and Mitchell, 2014; Xi and Serebrovskaya, 2012). For example, IH protocols simulating that experienced during sleep apnea (hundreds of hypoxic episodes per night for many nights; chronic intermittent hypoxia, CIH) is associated with maladaptive changes, including insomnia, cardiovascular disease, hypertension, metabolic syndrome and cognitive impairments (Brooks et al., 1997; Gozal et al., 2001; Lesske et al., 1997; Perry et al., 2008; Savransky et al., 2007). Cognitive impairments arise from neuronal apoptosis in brain regions necessary for learning and memory, including the hippocampus and frontal cortex (Gozal et al., 2001; Row et al., 2003). CIH-induced hippocampal apoptosis involves neuroinflammation and oxidative stress (Li et al., 2003; Li et al., 2004; Shan et al., 2007). In contrast, exposure to 3, 5-minute bouts of hypoxia (acute intermittent hypoxia; AIH), elicits neuroplasticity that persists for hours post-hypoxia (Bach and Mitchell, 1996; Terada and Mitchell, 2011). Regimens of repetitive acute intermittent hypoxia (rAIH) consisting of either daily (dAIH) or 3 times weekly (3×W AIH) exposure to 10, 5-minute hypoxic episodes also promote neuroplasticity (Lovett-Barr et al., 2012; MacFarlane, 2010; Satriotomo et al., 2012; Vinit, 2010; Wilkerson and Mitchell, 2009) and restore motor function in clinical disorders that compromise movements (e.g. spinal injury and ALS) such as breathing (Lovett-Barr et al., 2012; Nichols et al., 2013) or limb function (Hayes et al., 2014; Lovett-Barr et al., 2012; Trumbower et al., 2012).
rAIH is thought to promote neuroplasticity by up-regulating key proteins critical for neuroplasticity, including brain-derived neurotrophic factor (BDNF) and its receptor TrkB (Lovett-Barr et al., 2012; Satriotomo et al., 2012). Importantly, dAIH promotes neuroplasticity in the absence of hypertension or neuronal apoptosis typically associated with CIH (Lovett-Barr et al., 2012; Wilkerson and Mitchell, 2009), suggesting that it does not induce pathologies characteristic of CIH. In a cervical hemisection model of spinal cord injury, dAIH nearly completely restores the capacity to increase tidal volume during maximal chemoreceptor stimulation, and strengthens spontaneous and evoked electrical potentials in the phrenic nerve below the site of hemisection (Golder and Mitchell, 2005). Repetitive AIH also improved lower extremity strength and walking in humans with incomplete, chronic spinal cord injuries (Hayes et al., 2014; Trumbower et al., 2012), indicating that even mild paradigms of intermittent hypoxia may have clinical benefit.
Multiple CNS cell types produce inflammatory mediators, although microglia (resident CNS immune cells) are most often implicated. Microglia respond to CIH with increased inflammatory gene expression in a region- and time-dependent manner (Smith et al., 2013). However, the impact of rAIH on microglial or CNS inflammatory gene expression has not yet been investigated. Thus, a major goal of the present study was to examine regional CNS differences in inflammatory gene expression in microglia and CNS tissue homogenates (that include all CNS cell types) after AIH delivered 3 times per week, for 4 weeks (3×W AIH). After 3×W AIH, respiratory motor plasticity is enhanced in rats (MacFarlane, 2010). Since microglial neurotrophic factor production promotes neuronal survival and plasticity (Lalancette-Hebert et al., 2007; Parkhurst et al., 2013), we hypothesized that 3×W AIH would also increase microglial and tissue homogenate neurotrophic, but not inflammatory, gene expression in a region-specific manner.
We evaluated the expression of 10 genes (Table 1) in isolated microglia and regional CNS tissue homogenates. As “markers” of inflammation, we chose to analyze the expression of the pro-inflammatory cytokines interleukin (IL)-1β, IL-6 and tumor necrosis factor (TNF)α, and the enzyme cyclooxygenase (COX)-2, that inducibly produces prostaglandins. All inflammatory genes chosen are well-known to contribute to neuronal death in multiple models of CNS injury (Iadecola et al., 2001; Merrill, 1992; Munoz-Fernandez and Fresno, 1998; Spooren et al., 2011; Tzeng et al., 2005), and we have previously shown that CIH upregulates the expression of inducible nitric oxide synthase (iNOS), COX-2, IL-1β and IL-6 in microglia, depending on the CNS region evaluated (Smith et al., 2013). Also, important in the context of CIH, are the observations that inhibiting COX-2 protects neurons and reduces behavioral deficits induced by CIH exposure (Li et al., 2003). We included evaluation of toll-like receptor (TLR)4 as well, since we have previously reported that CIH upregulates its expression in microglia (Smith et al., 2013), and increased TLR4 activities are associated with neuroinflammation and neurodegeneration (Lehnardt et al., 2008; Rivest, 2009). We evaluated the expression of 4 neurotrophic factors, each of which have been shown in other models of CNS injury and pain to be upregulated specifically in microglia [e.g. insulin-like growth factor (IGF)-1 (Lalancette-Hebert et al., 2007) and BDNF (Coull et al., 2005)], and/or that are known to promote various forms of neuronal plasticity or neuroprotection [e.g. vascular endothelial growth factor (VEGF; Dale-Nagle et al., 2011; Storkebaum et al., 2004), BDNF (Numakawa, 2014; Parkhurst et al., 2013), IGF-1 (Benarroch, 2012; Deak and Sonntag, 2012), and adrenomedullin (ADM)-2 (Cheyuo et al., 2012)]. ADM-2 and VEGF are also highly hypoxia-responsive in other systems (via HIF activation) (Berra et al., 2000; Kaur et al., 2005).
TABLE 1.
| Gene | Forward (5′→3′) | Reverse (5′→3′) |
|---|---|---|
| 18s | CGG GTG CTC TTA GCT GAG TGT CCC G | CTC GGG CCT GCT TTG AAC AC |
| BDNF | CCC ATC ACA ATC TCA CGG TAT TC | TGC GGA GGG TCT CCT ATG AA |
| COX-2 | TGT TCC AAC CCA TGT CAA AA | CGT AGA ATC CAG TCC GGG TA |
| IGF-1 | CAG TTC GTG TGT GGA CCA AG | TCT TGG GCA TGT CAG TGT G |
| IFN-γ | CAT GGA TGC TAT GGA AGG AAA GA | GGC TCT CAA GTA TTT TCG TGT TAC C |
| IL-lβ | CTG CAG ATG CAA TGG AAA GA | TTG CTT CCA AGG CAG ACT TT |
| IL-4 | GCT TCC AGG GTG CTT CGC AAA T | ACG GTG CAG CTT CTC AGT GAG TT |
| IL-6 | GTG GCT AAG GAC CAA GAC CA | GGT TTG CCG AGT AGA CCT CA |
| IL-10 | AAG TGA TGC CCC AGG CAG AGA A | AAA TCG ATG ACA GCG TCG CAG C |
| IL-17 | GGG AAG TTG GAC CAC CAC CT | TTC TCC ACC CGG AAA GTG AA |
| iNOS | AGG GAG TGT TGT TCC AGG TG | TCT GCA GGA TGT CTT GAA CG |
| TLR4 | AGG CAG CAG GTG GAA TTG TAT C | TCG AGG CTT TTC CAT CCA ATA G |
| TNFα | TGC CAC TTC ATA CCA GGA GA | CCG GAC TCC GTG ATG TCT A |
| VEGF | TTG AGA CCC TGG TGG ACA TCT | CAC ACA GGA CGG CTT GAA GA |
As predicted, 3×W AIH did not increase inflammatory gene expression in microglia from frontal cortex, brainstem, upper cervical spinal cord (C1–C3) or lower cervical spinal cord (C4–C6). However, there were region- and gene-specific effects of 3×W AIH on certain inflammatory genes in brainstem and frontal cortex homogenates, although there were no detectable effects on inflammatory gene expression in cervical spinal homogenates. Contrary to our hypothesis, 3×W AIH did not increase microglial expression of the growth/trophic factors examined here, although there were gene-specific increases in brainstem tissue homogenates. Collectively, our results suggest that 3×W AIH does not promote microglial inflammation, nor does it augment their expression of growth/trophic factors that are critical for neuroplasticity. Other cell types do increase neurotrophic factor expression in some CNS regions, although this appears to only occur in CNS regions in which increased inflammatory gene expression was observed in tissue homogenates.
METHODS
Drugs/Materials
Anti-PE magnetic beads and MACS MS columns were purchased from Miltenyi Biotech (Germany). PE-conjugated mouse anti-rat CD11b was purchased from BD Biosciences (San Jose, CA). Papain was purchased from Sigma-Aldrich (St. Louis, MO) and DNaseI was purchased from Worthington Biochemical Corporation (Lakewood, NJ). Hank’s Buffered Salt Solution (HBSS) was purchased from Cellgro (Herndon, VA). TRI-Reagent was purchased from Sigma Aldrich (St. Louis, MO). Glycoblue reagent was purchased from Ambion (Austin, TX). MMLV reverse transcriptase was purchased from Invitrogen (Carlsbad, CA). Oligo dT, random primers, and RNAse inhibitor were purchased from Promega (Madison, WI). Primers were designed using Primer 3 software and were purchased from Integrated DNA Technologies (Coralville, IA). Power SYBR green was purchased from Applied Biosystems (Foster City, CA).
Animals
Adult male Sprague Dawley rats (3–4 months old) were housed in an AAALAC-accredited facility in 12hr:12hr light-dark conditions and were provided with food and water ad libitum. All surgical and experimental procedures were approved by the Animal Care and Use Committee at the University of Wisconsin. All procedures conformed to policies enumerated in the NIH Guide for the Care and Use of Laboratory Animals.
Rats were divided into two groups (n=8 each). The experimental group (3×W AIH) received ten, 5-minute episodes of 10.5% O2, alternating with 5-minute intervals of room air (21% O2), three days/week for 4 weeks, as described previously (Satriotomo et al., 2012). The control group received room air for 95 minutes, three times weekly for four weeks (normoxia; Nx). Both groups were placed in custom-made Plexiglas cylinders measuring 4.5 × 12 in, where oxygen and carbon dioxide levels were continuously monitored and regulated by a computer controlled using a specific script written in LabVIEW Software (National Instruments, New Delhi, India). Appropriate admixtures of O2 and N2 were piped into the chambers at a speed of 4 liters per minute. Control animals were exposed to room air only. Twenty-four hours following the last exposure, rats were euthanized by an overdose of isoflurane. Spleen samples were collected, and rats were perfused with ice-cold intraaortic phosphate-buffered saline to remove circulating myeloid cells before harvesting the following central nervous system regions: frontal cortex, brainstem, upper cervical spinal cord (C1–C3) and lower cervical spinal cord (C4–C6).
Microglial and CNS tissue homogenate sample preparation
Microglia were isolated from each CNS region using immunomagnetic separation, as previously described (Crain et al., 2009) with the exception that the tissue was dissociated with papain (0.7 mg/mL) and DNAse I (2 mg/mL) in HBSS for 24 minutes at 37C. Myelin was removed with high-speed centrifugation at 850g in 27% Percoll solution, and remaining cells were resuspended in buffer (PBS, BSA, EDTA). This suspension was incubated with PE-conjugated anti-CD11b+ antibodies, followed by incubation with an anti-PE antibody conjugated to a magnetic bead. Samples were loaded onto MACS MS columns, washed in a magnetic field, and CD11b+ cells were eluted. Prior to the anti-CD11b+ antibody incubation step, a sample was taken for CNS homogenate tissue analysis. As previously shown, this method of immunomagnetic isolation results in >97% of CD11b+/CD45low cells (Nikodemova and Watters, 2012). We subsequently refer to these CD11b+ cells as “microglia.”
RNA isolation and quantitative RT-PCR
Total RNA from CD11b+ cells, spleen and CNS region tissue homogenates was isolated using TriReagent and Glycoblue reagent according to the manufacturers’ instructions. Complementary DNA (cDNA) was synthesized from total RNA using MMLV reverse transcriptase and Oligo dT, random primers, and RNAse inhibitor as we have previously described (Crain et al., 2009). Based on RNA abundance, 150 ng, 300 ng and 1 μg of total RNA from isolated microglia, CNS tissue homogenates and spleen, respectively, were used in the cDNA synthesis reactions. qPCR was performed using PowerSYBR green PCR master mix on an ABI 7500 Fast system. Standard curves (Rutledge and Cote, 2003) were used to determine the relative expression of each gene in microglial and homogenate tissue samples using the average of duplicate, interpolated CT values, which were normalized to the expression of 18s ribosomal RNA. The delta-delta CT method was employed to determine relative expression of splenic genes relative to 18s ribosomal RNA (Livak and Schmittgen, 2001). The primer sequences used for qRT-PCR are shown in Table 1. Primers were designed using Primer 3 software and were purchased from Integrated DNA Technologies (Coralville, IA). Primers were designed to span introns wherever possible, and primer efficiency was assessed by use of standard curves, as we have previously reported (Crain et al., 2009). Gene transcripts were considered undetectable and not included in statistical analyses if their CT values fell outside of the linear range of the standard curve for that primer set; for all genes assessed here, this value was ≥34 cycles. If 50% or more of the animals in a treatment group had CT values that were undetectable or that fell outside the linear range of the primer set, that gene was considered below assay detection limits and data were excluded.
Statistical Analyses
For CNS tissues, analyses were performed on normalized, interpolated CT values from standard curves for each gene, as previously described (Smith et al., 2013). Data from spleen were similarly analyzed using fold change of each gene in question relative to ribosomal RNA. CT values were identified and discarded as outliers if they fell outside of 1.96 z-scores from the mean, as evaluated by the Grubb’s test. The number of outliers per sample varied from zero to three and is indicated in the Results. Student t-tests were conducted using Sigma Plot 11.0 software (San Jose CA). Data sets failing normality were logarithmically transformed prior to analysis with the Whitney-Mann Rank Sum Test. Statistical significance was indicated by a p-value <0.05.
RESULTS
3×W AIH does not elicit cortical microglial inflammation, but induces gene-specific effects in CNS homogenates
Cortical microglial expression of inflammatory and neurotrophic genes was unchanged by 3×W AIH (Fig. 1A), although there was a non-significant trend towards decreased Cox-2 gene expression (~50%; p=0.169). The only significant change detected in cortical homogenates following 3×W AIH was increased Cox-2 expression (>70%; p=0.042; Fig. 1B), suggesting that the relevant Cox-2 was in cells other than microglia.
Figure 1. 3×W AIH does not promote inflammatory gene expression in rat cortical microglia, but it has gene-specific effects in cortical homogenates.
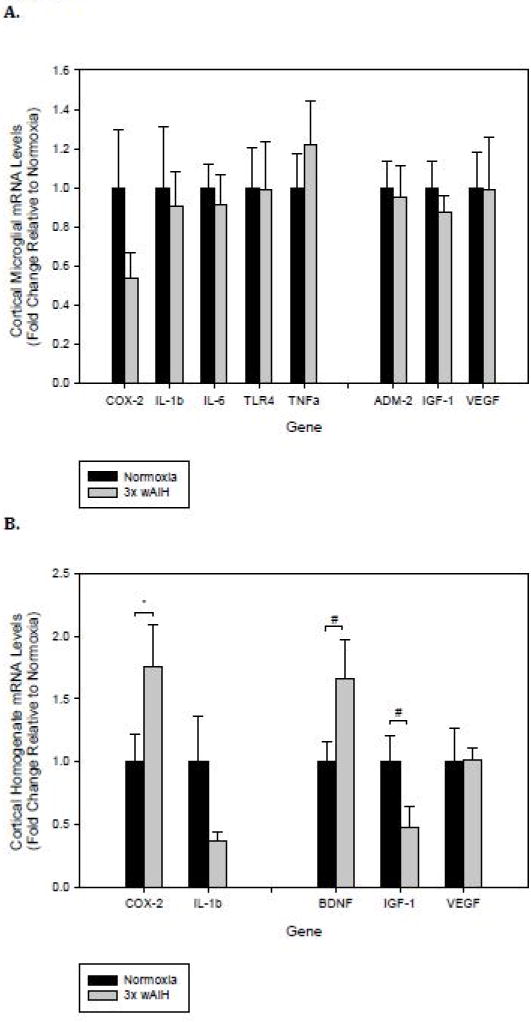
Gene expression was assessed by qRT-PCR in immunomagnetically-isolated microglia (A), and tissue homogenates from the frontal cortex (B) following 4 weeks of 3×W AIH. Treatment did not alter the expression of any gene evaluated in microglia, but there were gene-specific effects in tissue homogenates. Data shown are means ± 1 SEM of n=5–8. *p<0.05 and #0.05<p<0.1 versus normoxia.
3×W AIH differentially affects gene expression in brainstem microglia versus homogenates
3×W AIH did not change inflammatory or neurotrophic factor gene expression in brainstem microglia (Fig. 2A), although there was a non-significant trend towards decreased Bdnf mRNA levels (~50%; p=0.084). In contrast, there were selective increases in pro-inflammatory gene expression in brainstem homogenates (Fig. 2B). Specifically, Tlr4 gene expression increased >2.5 fold (p=0.017). With regard to neurotrophic factor expression, Vegf mRNA levels increased >50% in brainstem homogenates (p= 0.018).
Figure 2. 3×W AIH does not promote inflammatory gene expression in rat brainstem microglia, but it increases expression of Tlr4 and Vegf in brainstem homogenates.
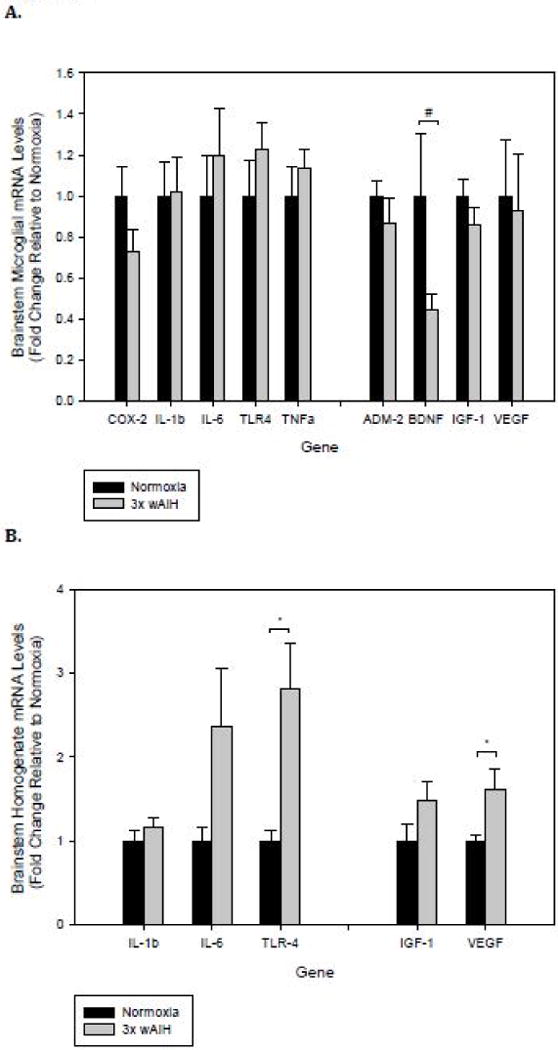
Gene expression was assessed by qRT-PCR in immunomagnetically-isolated microglia (A), and tissue homogenates from the brainstem (B) following 4 weeks of 3×W AIH. Treatment did not alter the expression of any inflammatory gene evaluated in microglia, though there was a non-significant reduction in Bdnf mRNA levels. 3×W AIH increased Tlr4 and Vegf gene expression in tissue homogenates. Data shown are means ± 1 SEM of n=5–8. *p<0.05 and #0.05<p<0.1 versus normoxia.
3×W AIH gene-specifically decreases expression of microglial neurotrophic factors in the upper cervical spinal cord
Inflammatory gene expression in microglia from the upper cervical spinal cord was unchanged by 3×W AIH (Fig. 3A). However, in this region, trophic genes demonstrated a varied response. Specifically, microglial Adm-2 (p=0.015) and Vegf (p=0.021) expression decreased by nearly 50% (Fig. 3B).
Figure 3. C1–C3 spinal microglial inflammatory gene expression was unaltered following 3×W AIH exposure, but Adm-2 and Vegf mRNA levels were down-regulated.
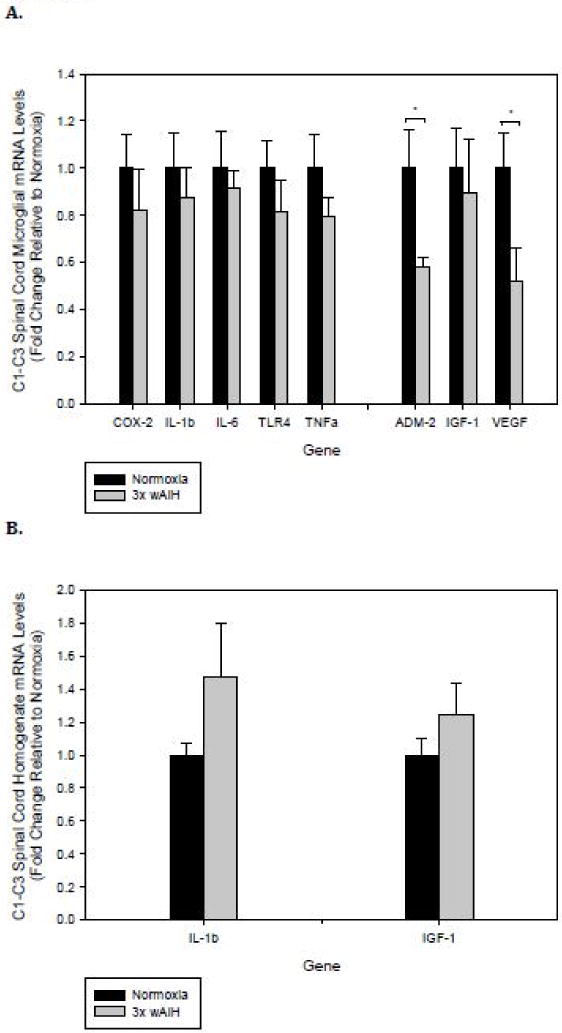
Gene expression was assessed by qRT-PCR in immunomagnetically-isolated microglia (A), and tissue homogenates from C1–C3 cervical spinal segments (B) following 4 weeks of 3×W AIH. Treatment did not alter the expression of any inflammatory gene evaluated in microglia, but it reduced Adm-2 and Vegf mRNA levels. 3×W AIH had no effects on the expression of any gene evaluated in C1–C3 tissue homogenates. Data shown are means ± 1 SEM of n=6–8. *p<0.05 versus normoxia.
3×W AIH has no detectable effects on microglial or homogenate inflammatory or neurotrophic factor gene expression in the lower cervical spinal cord
Neither pro-inflammatory nor trophic factor gene expression was altered by 3×W AIH in C4–C6 microglia (Fig. 4A) or homogenates (Fig. 4B).
Figure 4. 3×W AIH had no effect on C4–C6 spinal microglial or tissue homogenate gene expression.
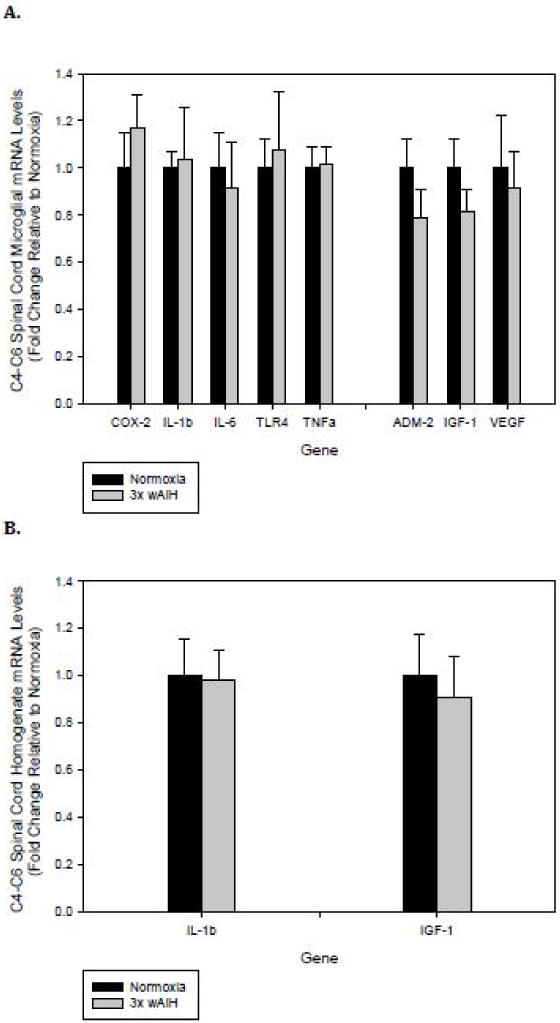
Gene expression was assessed by qRT-PCR in immunomagnetically-isolated microglia (A), and tissue homogenates from C4–C6 cervical spinal segments (B) following 4 weeks of 3×W AIH. Treatment did not alter the expression of any inflammatory or trophic factor gene evaluated in either microglia or C4–C6 tissue homogenates. Data shown are means ± 1 SEM of n=5–8.
3×W AIH does not promote observable inflammation in the spleen
We evaluated the expression of 6 key pro-inflammatory and 2 anti-inflammatory genes in the spleens of rats exposed to 3×W AIH. We were unable to detect mRNA levels of the cytokine interleukin-4 following 3×W AIH (data not shown), and minimal change was observed in IL-10 mRNA, providing no evidence for anti-inflammatory (M2) macrophage activation. Although basal Cox-2, Ifn-g, Il-1β, Il-6, Il-17, and iNos expression could be detected, no significant changes were observed after 3×W AIH (Fig. 5), suggesting that 3×W AIH does not promote systemic inflammation.
Figure 5. 3×W AIH had no effect on the expression of any gene evaluated in the spleen.
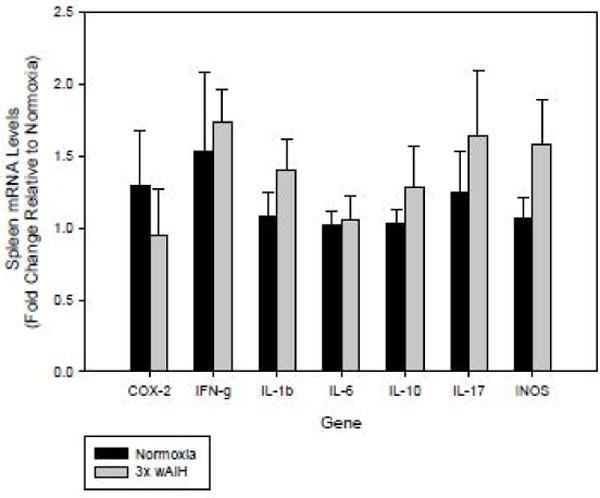
Gene expression was assessed by qRT-PCR in the spleen following 4 weeks of 3×W AIH. Treatment did not alter the expression of any pro- or anti-inflammatory gene evaluated. Data shown are means ± 1 SEM of n=7–8.
DISCUSSION
Modest protocols of repetitive acute intermittent hypoxia (rAIH) have therapeutic potential to treat clinical disorders that impair movement, inducing spinal motor plasticity, without the attendant pathologies of neuronal loss, reactive gliosis, or systemic hypertension (Dale et al., 2014; Navarrete-Opazo and Mitchell, 2014; Satriotomo et al., 2012). Microglial inflammatory activities likely contribute in part to neuroinflammation and neuronal loss resulting from “high dose” pathologic intermittent hypoxia [i.e. CIH; (Yang et al., 2013; Zhang et al., 2012)]. However, the effects of rAIH on neuroinflammation, and microglial inflammatory gene expression have not been previously examined. Here we show that a protocol of rAIH known to elicit functional benefits in rodent injury models [i.e. 3×W AIH; (MacFarlane, 2010)] does not increase microglial inflammatory gene expression in any CNS region examined here, although the expression of two pro-inflammatory genes increased in homogenates, suggesting a cellular source other than microglia (e.g. astrocytes or neurons). Likewise, we did not detect evidence of peripheral (splenic) inflammation based on the expression of 8 classical pro- and anti-inflammatory genes examined after 3×W AIH.
Given the beneficial effects of rAIH paradigms in rodent injury models and patients (Golder and Mitchell, 2005; Hayes et al., 2014; Trumbower et al., 2012), and known neuroprotective effects of microglial neurotrophic factor production (Lalancette-Hebert et al., 2007; Parkhurst et al., 2013), we also expected rAIH-induced upregulation of microglial expression of key growth factors important for neuronal plasticity and/or survival. However, we did not observe increases in any microglial neurotrophic/growth factor expression examined, although there were region- and gene-specific increases in tissue homogenates. Together, these results suggest that, unlike CIH (Smith et al., 2013), 4 weeks of 3×W AIH does not promote CNS neuroinflammation, nor are there changes in the expression of any of the 10 inflammatory or neurotrophic genes evaluated here in microglia. Microglial and tissue homogenate expression of iNOS was also analyzed, and showed no statistically significant differences upon 3×W AIH exposure, regardless of the CNS region studied (data not shown). Thus, these results contribute to accumulating evidence that rAIH is a safe and effective therapeutic approach to treat motor impairment (Dale et al., 2014; Navarrete-Opazo and Mitchell, 2014).
Because of its importance to rAIH-induced respiratory motor plasticity and metaplasticity (Dale et al., 2014), we were especially interested in modulation of gene expression in the region of the phrenic motor nucleus (i.e. C4–C6). Similar rAIH protocols increase expression of proteins critical for phrenic motor plasticity (Satriotomo et al., 2012; Wilkerson and Mitchell, 2009), leading to our hypothesis that rAIH would increase mRNA levels of these same genes. However, no significant changes in microglial or homogenate gene expression of either neurotrophic factors or inflammatory markers were observed in this region. Growth/trophic factor upregulation within phrenic motor neurons (Satriotomo et al., 2012) is difficult to detect in tissue homogenates due to the extensive dilution of ~600 phrenic motor neurons in the large volume of the cervical ventral horn. From another perspective, the absence of cervical spinal inflammation is meaningful because systemic inflammation induces cervical spinal inflammatory gene expression and undermines AIH-induced spinal, respiratory motor plasticity (Huxtable et al., 2013). Therefore, it is encouraging that this rAIH protocol, which may be therapeutic in spinal injury patients, does not cause inflammation in the region of the phrenic motor nucleus that may undermine its therapeutic efficacy.
Significant increases in the expression of two pro-inflammatory genes were detected in tissue homogenates: Cox-2 in the cortex and Tlr4 in the brainstem. Since corresponding changes in microglial expression of these genes were not observed, other cell types are most likely responsible for these changes in tissue homogenate mRNA levels (e.g. astrocytes or neurons). The functional significance of these observations is not yet known. Since COX-2 plays critical roles in synaptic plasticity (Gruol, 2014; Yang and Chen, 2008), and TLR4 signaling can promote neuroprotection (Chen et al., 2012; Marsh et al., 2009; Vartanian et al., 2011), increased expression of these genes, which are typically thought of as pro-inflammatory, may play roles other than neuroinflammatory/pathologic in this situation.
3×W AIH effects on microglial trophic factor gene expression varied by CNS region. Although we had hypothesized that this rAIH protocol would up-regulate microglial expression of genes important for neuroprotection and neuroplasticity, we found no detectable increases in the microglial expression of any of the gene studied here. However, there were limited region-specific decreases in microglial expression of certain genes. For example, C1–C3 cervical microglia demonstrated a reduced expression of Adm-2 and Vegf - genes whose products exert neuroprotective effects (Cheyuo et al., 2012; Rosenstein et al., 2010) and are directly upregulated by the hypoxia-sensitive transcription factor HIF-1 (hypoxia-inducible factor 1) (Shibuya, 2013; Zudaire et al., 2006). In contrast, in brainstem homogenates, Vegf mRNA levels increased. Thus, it is important to understand cell- and region-specific responses to 3×W AIH since microglial and CNS homogenate responses sometimes differ depending on the CNS region and the specific gene in question.
Leconte and colleagues (Leconte et al., 2012) demonstrated that a similar intermittent hypoxia protocol induced statistically significant increases in the expression of Adm-2 and Vegf genes in mouse whole brain homogenates after 41 days of AIH exposure. However, there are multiple, important variables that differ between that study and ours, aside from species. First, the expression of genes in all cell types from whole brain will differ from results obtained in microglia and tissue homogenates from individual CNS regions as in our study. Further, their hypoxic exposures consisted of 8% O2 for 1 hour per day, 3 days per week for 6 weeks; our exposures consisted of 10, 5 min episodes of 10.5% O2, 3 days per week for 4 weeks. It is well-established that the CNS responds differently to stimuli delivered in massed versus spaced patterns (Baker and Mitchell, 2000; Glanzman et al., 1990).
Brain-derived neurotrophic factor (BDNF) is important during development for neuronal survival and differentiation (Xie and Yung, 2012), motor learning (Parkhurst et al., 2013) including AIH-induced phrenic long-term facilitation (Baker-Herman et al., 2004), and hippocampal synaptic plasticity (Pang et al., 2004). Because AIH-induced phrenic long-term facilitation relies on increased BDNF synthesis (Baker-Herman et al., 2004; Wilkerson and Mitchell, 2009), and because rAIH elicits respiratory metaplasticity (Lovett-Barr et al., 2012; MacFarlane et al., 2008; MacFarlane, 2010; Vinit et al., 2009; Vinit, 2010; Wilkerson and Mitchell, 2009), we had hypothesized that BDNF expression in at least some CNS regions, would be up-regulated by 3×W AIH. In agreement, increased Bdnf expression has been reported in the pons, but not medulla, after 3 brief bouts of hypoxia in mice (Vermehren-Schmaedick et al., 2012). However, of the four CNS regions evaluated after 3×W AIH in our study, we detected BDNF mRNA only in cortical tissue homogenates and in brainstem microglia. Yet, despite trends, there were no significant changes after exposure to 3×W AIH. Interpretation of this finding must be made with caution however, since we do not consider three important factors in this study: 1) increased BDNF protein expression may occur via increased translation from pre-existing mRNA versus new transcriptional upregulation; 2) persistent changes in protein and mRNA levels within limited populations of cells (e.g. motor neurons; Satriotomo et al., 2012; Lovett-Barr et al., 2012) may be obscured by dilution in a large volume constituting the tissue homogenate; or 3) increased BDNF mRNA levels may be transient, and are no longer evident after 4 weeks of 3×W AIH.
In this regard, it is important to acknowledge the limitations of the present study. We utilized an immunomagnetic cell isolation approach to isolate microglia from different regions of the CNS. This method does not allow distinction of heterogeneous microglial inflammatory responses from cell populations that are low in frequency. Thus, it is feasible that there are small populations of microglia that respond differently to 3×W AIH that are too few in number to influence the overall means of the inflammatory gene expression measured here. However, the functional significance of such small populations is also unclear at this time, as are the mechanisms underlying their heterogeneic responses. In addition, inflammatory gene and growth factor mRNAs, tend to have very short half-lives due to the presence of adenine uridine rich elements within their 3′ untranslated regions (Caput et al., 1986; Hamilton et al., 2012; Shaw and Kamen, 1986), so the timing of gene analysis performed here (1 day after the last AIH exposure for 4 weeks) may be too late to detect changes in inflammatory gene mRNA levels.
To assess the possibility of systemic inflammation induced by rAIH, we evaluated splenic expression of 7 prototypical (M1) pro-inflammatory cytokines – cyclooxygenase-2, interferon gamma, inducible nitric oxide synthase, interleukin-1β, interleukin-4, interleukin-6, and interleukin-17, as well as the canonical anti-inflammatory (M2) cytokine interleukin-10. We found no evidence for pro- or anti-inflammatory gene expression in the spleen after 3×W AIH, although there were insignificant increases in mRNA levels of some inflammatory genes. We conclude that 4 weeks of 3×W AIH does not initiate or maintain inflammation in the spleen, a key peripheral immune organ rich in B and T lymphocytes.
Modest intermittent hypoxia protocols are being investigated as a means to treat neuromotor disorders (Dale et al., 2014; Navarrete-Opazo et al., 2015). However, it is important to analyze the ability of such intermittent hypoxia therapy to cause pathologies such as untoward neural and systemic inflammation. The results presented here directly evaluate inflammatory gene expression in microglia and tissue homogenates from various CNS regions after a therapeutically applicable protocol of repetitive acute intermittent hypoxia. Although we found little evidence for appreciable inflammation after 4 weeks of rAIH, it would be useful to evaluate other time points in future investigations since it remains possible that immediate or long term rAIH could elicit pathologic inflammatory responses.
Highlights.
-
■
The objective is proposing an evolutionary model which accepts knowledge as intensely active and dynamic and evolving in maturity from the early stages of research.
-
■
But this is possible only if the research team adopt a clear, clean and joint process of disciplinary integration and transdisciplinary integration of the produced and discovered knowledge.
-
■
In this way, the results of research will have a greater influence on society and they also will be adopted by society.
Acknowledgments
We would like to thank Dr. Stephanie Smith, Mr. Jonathan Ouellette, Dr. Maria Nikodemova, and Mr. Scott Ray for their technical support on this project. We would also like to thank Mr. Bradley Wathen for his technical support and maintenance of the intermittent hypoxia exposure system. This work was supported by NIH R01 HL111598 (GSM and JJW).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Bach KB, Mitchell GS. Hypoxia-induced long-term facilitation of respiratory activity is serotonin dependent. Respiration physiology. 1996;104:251–260. doi: 10.1016/0034-5687(96)00017-5. [DOI] [PubMed] [Google Scholar]
- Baker TL, Mitchell GS. Episodic but not continuous hypoxia elicits long-term facilitation of phrenic motor output in rats. The Journal of physiology. 2000;529(Pt 1):215–219. doi: 10.1111/j.1469-7793.2000.00215.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baker-Herman TL, Fuller DD, Bavis RW, Zabka AG, Golder FJ, Doperalski NJ, Johnson RA, Watters JJ, Mitchell GS. BDNF is necessary and sufficient for spinal respiratory plasticity following intermittent hypoxia. Nature neuroscience. 2004;7:48–55. doi: 10.1038/nn1166. [DOI] [PubMed] [Google Scholar]
- Benarroch EE. Insulin-like growth factors in the brain and their potential clinical implications. Neurology. 2012;79:2148–2153. doi: 10.1212/WNL.0b013e3182752eef. [DOI] [PubMed] [Google Scholar]
- Berra E, Pages G, Pouyssegur J. MAP kinases and hypoxia in the control of VEGF expression. Cancer metastasis reviews. 2000;19:139–145. doi: 10.1023/a:1026506011458. [DOI] [PubMed] [Google Scholar]
- Brooks D, Horner RL, Kozar LF, Render-Teixeira CL, Phillipson EA. Obstructive sleep apnea as a cause of systemic hypertension. Evidence from a canine model. The Journal of clinical investigation. 1997;99:106–109. doi: 10.1172/JCI119120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caput D, Beutler B, Hartog K, Thayer R, Brown-Shimer S, Cerami A. Identification of a common nucleotide sequence in the 3′-untranslated region of mRNA molecules specifying inflammatory mediators. Proceedings of the National Academy of Sciences of the United States of America. 1986;83:1670–1674. doi: 10.1073/pnas.83.6.1670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Z, Jalabi W, Shpargel KB, Farabaugh KT, Dutta R, Yin X, Kidd GJ, Bergmann CC, Stohlman SA, Trapp BD. Lipopolysaccharide-induced microglial activation and neuroprotection against experimental brain injury is independent of hematogenous TLR4. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2012;32:11706–11715. doi: 10.1523/JNEUROSCI.0730-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheyuo C, Yang WL, Wang P. The critical role of adrenomedullin and its binding protein, AMBP-1, in neuroprotection. Biological chemistry. 2012;393:429–439. doi: 10.1515/hsz-2012-0103. [DOI] [PubMed] [Google Scholar]
- Coull JA, Beggs S, Boudreau D, Boivin D, Tsuda M, Inoue K, Gravel C, Salter MW, De Koninck Y. BDNF from microglia causes the shift in neuronal anion gradient underlying neuropathic pain. Nature. 2005;438:1017–1021. doi: 10.1038/nature04223. [DOI] [PubMed] [Google Scholar]
- Crain JM, Nikodemova M, Watters JJ. Expression of P2 nucleotide receptors varies with age and sex in murine brain microglia. Journal of neuroinflammation. 2009;6:24. doi: 10.1186/1742-2094-6-24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dale EA, Ben Mabrouk F, Mitchell GS. Unexpected benefits of intermittent hypoxia: enhanced respiratory and nonrespiratory motor function. Physiology (Bethesda, Md) 2014;29:39–48. doi: 10.1152/physiol.00012.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dale-Nagle EA, Satriotomo I, Mitchell GS. Spinal vascular endothelial growth factor induces phrenic motor facilitation via extracellular signal-regulated kinase and Akt signaling. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2011;31:7682–7690. doi: 10.1523/JNEUROSCI.0239-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deak F, Sonntag WE. Aging, synaptic dysfunction, and insulin-like growth factor (IGF)-1. The journals of gerontology. Series A, Biological sciences and medical sciences. 2012;67:611–625. doi: 10.1093/gerona/gls118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glanzman DL, Kandel ER, Schacher S. Target-dependent structural changes accompanying long-term synaptic facilitation in Aplysia neurons. Science (New York, NY) 1990;249:799–802. doi: 10.1126/science.2389145. [DOI] [PubMed] [Google Scholar]
- Golder FJ, Mitchell GS. Spinal synaptic enhancement with acute intermittent hypoxia improves respiratory function after chronic cervical spinal cord injury. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2005;25:2925–2932. doi: 10.1523/JNEUROSCI.0148-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gozal E, Row BW, Schurr A, Gozal D. Developmental differences in cortical and hippocampal vulnerability to intermittent hypoxia in the rat. Neuroscience letters. 2001;305:197–201. doi: 10.1016/s0304-3940(01)01853-5. [DOI] [PubMed] [Google Scholar]
- Gruol DL. IL-6 Regulation of Synaptic Function in the CNS. Neuropharmacology. 2014:1–13. doi: 10.1016/j.neuropharm.2014.10.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamilton T, Li X, Novotny M, Pavicic PG, Jr, Datta S, Zhao C, Hartupee J, Sun D. Cell type- and stimulus-specific mechanisms for post-transcriptional control of neutrophil chemokine gene expression. Journal of leukocyte biology. 2012;91:377–383. doi: 10.1189/jlb.0811404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayes HB, Jayaraman A, Herrmann M, Mitchell GS, Rymer WZ, Trumbower RD. Daily intermittent hypoxia enhances walking after chronic spinal cord injury: a randomized trial. Neurology. 2014;82:104–113. doi: 10.1212/01.WNL.0000437416.34298.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huxtable AG, Smith SM, Vinit S, Watters JJ, Mitchell GS. Systemic LPS induces spinal inflammatory gene expression and impairs phrenic long-term facilitation following acute intermittent hypoxia. Journal of applied physiology (Bethesda, Md: 1985) 2013;114:879–887. doi: 10.1152/japplphysiol.01347.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iadecola C, Niwa K, Nogawa S, Zhao X, Nagayama M, Araki E, Morham S, Ross ME. Reduced susceptibility to ischemic brain injury and N-methyl-D-aspartate-mediated neurotoxicity in cyclooxygenase-2-deficient mice. Proceedings of the National Academy of Sciences of the United States of America. 2001;98:1294–1299. doi: 10.1073/pnas.98.3.1294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaur B, Khwaja FW, Severson EA, Matheny SL, Brat DJ, Van Meir EG. Hypoxia and the hypoxia-inducible-factor pathway in glioma growth and angiogenesis. Neuro-oncology. 2005;7:134–153. doi: 10.1215/S1152851704001115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lalancette-Hebert M, Gowing G, Simard A, Weng YC, Kriz J. Selective ablation of proliferating microglial cells exacerbates ischemic injury in the brain. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2007;27:2596–2605. doi: 10.1523/JNEUROSCI.5360-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leconte C, Leger M, Boulouard M, Tixier E, Freret T, Bernaudin M, Schumann-Bard P. Repeated mild hypoxic exposures decrease anxietylike behavior in the adult mouse together with an increased brain adrenomedullin gene expression. Behavioural brain research. 2012;230:78–84. doi: 10.1016/j.bbr.2012.01.054. [DOI] [PubMed] [Google Scholar]
- Lehnardt S, Schott E, Trimbuch T, Laubisch D, Krueger C, Wulczyn G, Nitsch R, Weber JR. A vicious cycle involving release of heat shock protein 60 from injured cells and activation of toll-like receptor 4 mediates neurodegeneration in the CNS. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2008;28:2320–2331. doi: 10.1523/JNEUROSCI.4760-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lesske J, Fletcher EC, Bao G, Unger T. Hypertension caused by chronic intermittent hypoxia–influence of chemoreceptors and sympathetic nervous system. Journal of hypertension. 1997;15:1593–1603. doi: 10.1097/00004872-199715120-00060. [DOI] [PubMed] [Google Scholar]
- Li RC, Row BW, Gozal E, Kheirandish L, Fan Q, Brittian KR, Guo SZ, Sachleben LR, Jr, Gozal D. Cyclooxygenase 2 and intermittent hypoxia-induced spatial deficits in the rat. American journal of respiratory and critical care medicine. 2003;168:469–475. doi: 10.1164/rccm.200211-1264OC. [DOI] [PubMed] [Google Scholar]
- Li RC, Row BW, Kheirandish L, Brittian KR, Gozal E, Guo SZ, Sachleben LR, Jr, Gozal D. Nitric oxide synthase and intermittent hypoxia-induced spatial learning deficits in the rat. Neurobiology of disease. 2004;17:44–53. doi: 10.1016/j.nbd.2004.05.006. [DOI] [PubMed] [Google Scholar]
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods (San Diego, Calif) 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- Lovett-Barr MR, Satriotomo I, Muir GD, Wilkerson JE, Hoffman MS, Vinit S, Mitchell GS. Repetitive intermittent hypoxia induces respiratory and somatic motor recovery after chronic cervical spinal injury. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2012;32:3591–3600. doi: 10.1523/JNEUROSCI.2908-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacFarlane PM, Wilkerson JE, Lovett-Barr MR, Mitchell GS. Reactive oxygen species and respiratory plasticity following intermittent hypoxia. Respiratory physiology & neurobiology. 2008;164:263–271. doi: 10.1016/j.resp.2008.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacFarlane PMVS, Roopra A, Mitchell GS. Enhanced Phrenic Long-Term Facilitation Following Repetitive Acute Intermittent Hypoxia: Role of Glygolytic Flux. Federation of American Societies for Experimental Biology 2010 [Google Scholar]
- Marsh B, Stevens SL, Packard AE, Gopalan B, Hunter B, Leung PY, Harrington CA, Stenzel-Poore MP. Systemic lipopolysaccharide protects the brain from ischemic injury by reprogramming the response of the brain to stroke: a critical role for IRF3. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2009;29:9839–9849. doi: 10.1523/JNEUROSCI.2496-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merrill JE. Tumor necrosis factor alpha, interleukin 1 and related cytokines in brain development: normal and pathological. Developmental neuroscience. 1992;14:1–10. doi: 10.1159/000111642. [DOI] [PubMed] [Google Scholar]
- Munoz-Fernandez MA, Fresno M. The role of tumour necrosis factor, interleukin 6, interferon-gamma and inducible nitric oxide synthase in the development and pathology of the nervous system. Progress in neurobiology. 1998;56:307–340. doi: 10.1016/s0301-0082(98)00045-8. [DOI] [PubMed] [Google Scholar]
- Navarrete-Opazo A, Mitchell GS. Therapeutic potential of intermittent hypoxia: a matter of dose. American journal of physiology. Regulatory, integrative and comparative physiology. 2014;307:R1181–R1197. doi: 10.1152/ajpregu.00208.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Navarrete-Opazo A, Vinit S, Dougherty BJ, Mitchell GS. Daily acute intermittent hypoxia elicits functional recovery of diaphragm and inspiratory intercostal muscle activity after acute cervical spinal injury. Experimental neurology. 2015;266C:1–10. doi: 10.1016/j.expneurol.2015.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nichols NL, Gowing G, Satriotomo I, Nashold LJ, Dale EA, Suzuki M, Avalos P, Mulcrone PL, McHugh J, Svendsen CN, Mitchell GS. Intermittent hypoxia and stem cell implants preserve breathing capacity in a rodent model of amyotrophic lateral sclerosis. American journal of respiratory and critical care medicine. 2013;187:535–542. doi: 10.1164/rccm.201206-1072OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nikodemova M, Watters JJ. Efficient isolation of live microglia with preserved phenotypes from adult mouse brain. Journal of neuroinflammation. 2012;9:147. doi: 10.1186/1742-2094-9-147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Numakawa T. Possible protective action of neurotrophic factors and natural compounds against common neurodegenerative diseases. Neural regeneration research. 2014;9:1506–1508. doi: 10.4103/1673-5374.139474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pang PT, Teng HK, Zaitsev E, Woo NT, Sakata K, Zhen S, Teng KK, Yung WH, Hempstead BL, Lu B. Cleavage of proBDNF by tPA/plasmin is essential for long-term hippocampal plasticity. Science (New York, NY) 2004;306:487–491. doi: 10.1126/science.1100135. [DOI] [PubMed] [Google Scholar]
- Parkhurst CN, Yang G, Ninan I, Savas JN, Yates JR, 3rd, Lafaille JJ, Hempstead BL, Littman DR, Gan WB. Microglia promote learning-dependent synapse formation through brain-derived neurotrophic factor. Cell. 2013;155:1596–1609. doi: 10.1016/j.cell.2013.11.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perry JC, D’Almeida V, Lima MM, Godoi FR, Vital MA, Oliveira MG, Tufik S. Intermittent hypoxia and sleep restriction: motor, cognitive and neurochemical alterations in rats. Behavioural brain research. 2008;189:373–380. doi: 10.1016/j.bbr.2008.01.014. [DOI] [PubMed] [Google Scholar]
- Rivest S. Regulation of innate immune responses in the brain. Nature reviews. Immunology. 2009;9:429–439. doi: 10.1038/nri2565. [DOI] [PubMed] [Google Scholar]
- Rosenstein JM, Krum JM, Ruhrberg C. VEGF in the nervous system. Organogenesis. 2010;6:107–114. doi: 10.4161/org.6.2.11687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Row BW, Liu R, Xu W, Kheirandish L, Gozal D. Intermittent hypoxia is associated with oxidative stress and spatial learning deficits in the rat. American journal of respiratory and critical care medicine. 2003;167:1548–1553. doi: 10.1164/rccm.200209-1050OC. [DOI] [PubMed] [Google Scholar]
- Rutledge RG, Cote C. Mathematics of quantitative kinetic PCR and the application of standard curves. Nucleic acids research. 2003;31:e93. doi: 10.1093/nar/gng093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Satriotomo I, Dale EA, Dahlberg JM, Mitchell GS. Repetitive acute intermittent hypoxia increases expression of proteins associated with plasticity in the phrenic motor nucleus. Experimental neurology. 2012;237:103–115. doi: 10.1016/j.expneurol.2012.05.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savransky V, Nanayakkara A, Li J, Bevans S, Smith PL, Rodriguez A, Polotsky VY. Chronic intermittent hypoxia induces atherosclerosis. American journal of respiratory and critical care medicine. 2007;175:1290–1297. doi: 10.1164/rccm.200612-1771OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shan X, Chi L, Ke Y, Luo C, Qian S, Gozal D, Liu R. Manganese superoxide dismutase protects mouse cortical neurons from chronic intermittent hypoxia-mediated oxidative damage. Neurobiology of disease. 2007;28:206–215. doi: 10.1016/j.nbd.2007.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaw G, Kamen R. A conserved AU sequence from the 3′ untranslated region of GM-CSF mRNA mediates selective mRNA degradation. Cell. 1986;46:659–667. doi: 10.1016/0092-8674(86)90341-7. [DOI] [PubMed] [Google Scholar]
- Shibuya M. Vascular endothelial growth factor and its receptor system: physiological functions in angiogenesis and pathological roles in various diseases. Journal of biochemistry. 2013;153:13–19. doi: 10.1093/jb/mvs136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith SM, Friedle SA, Watters JJ. Chronic intermittent hypoxia exerts CNS region-specific effects on rat microglial inflammatory and TLR4 gene expression. PloS one. 2013;8:e81584. doi: 10.1371/journal.pone.0081584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spooren A, Kolmus K, Laureys G, Clinckers R, De Keyser J, Haegeman G, Gerlo S. Interleukin-6, a mental cytokine. Brain research reviews. 2011;67:157–183. doi: 10.1016/j.brainresrev.2011.01.002. [DOI] [PubMed] [Google Scholar]
- Storkebaum E, Lambrechts D, Carmeliet P. VEGF: once regarded as a specific angiogenic factor, now implicated in neuroprotection. BioEssays : news and reviews in molecular, cellular and developmental biology. 2004;26:943–954. doi: 10.1002/bies.20092. [DOI] [PubMed] [Google Scholar]
- Terada J, Mitchell GS. Diaphragm long-term facilitation following acute intermittent hypoxia during wakefulness and sleep. Journal of applied physiology (Bethesda, Md: 1985) 2011;110:1299–1310. doi: 10.1152/japplphysiol.00055.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trumbower RD, Jayaraman A, Mitchell GS, Rymer WZ. Exposure to acute intermittent hypoxia augments somatic motor function in humans with incomplete spinal cord injury. Neurorehabilitation and neural repair. 2012;26:163–172. doi: 10.1177/1545968311412055. [DOI] [PubMed] [Google Scholar]
- Tzeng SF, Hsiao HY, Mak OT. Prostaglandins and cyclooxygenases in glial cells during brain inflammation. Current drug targets. Inflammation and allergy. 2005;4:335–340. doi: 10.2174/1568010054022051. [DOI] [PubMed] [Google Scholar]
- Vartanian KB, Stevens SL, Marsh BJ, Williams-Karnesky R, Lessov NS, Stenzel-Poore MP. LPS preconditioning redirects TLR signaling following stroke: TRIF-IRF3 plays a seminal role in mediating tolerance to ischemic injury. Journal of neuroinflammation. 2011;8:140. doi: 10.1186/1742-2094-8-140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vermehren-Schmaedick A, Jenkins VK, Knopp SJ, Balkowiec A, Bissonnette JM. Acute intermittent hypoxia-induced expression of brain-derived neurotrophic factor is disrupted in the brainstem of methyl-CpG-binding protein 2 null mice. Neuroscience. 2012;206:1–6. doi: 10.1016/j.neuroscience.2012.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vinit S, Lovett-Barr MR, Mitchell GS. Intermittent hypoxia induces functional recovery following cervical spinal injury. Respiratory physiology & neurobiology. 2009;169:210–217. doi: 10.1016/j.resp.2009.07.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vinit SMPM, Satriotomo I, Mitchell GS. Enhanced Phrenic Long-Term Facilitation (pLTF) Following Repetitive Acute Intermittent Hypoxia. Federation of American Societies for Experimental Biology 2010 [Google Scholar]
- Wilkerson JE, Mitchell GS. Daily intermittent hypoxia augments spinal BDNF levels, ERK phosphorylation and respiratory long-term facilitation. Experimental neurology. 2009;217:116–123. doi: 10.1016/j.expneurol.2009.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xi L, Serebrovskaya TV. Intermittent Hypoxia and Human Diseases. 1. Springer-Verlag; London: 2012. [Google Scholar]
- Xie H, Yung WH. Chronic intermittent hypoxia-induced deficits in synaptic plasticity and neurocognitive functions: a role for brain-derived neurotrophic factor. Acta pharmacologica Sinica. 2012;33:5–10. doi: 10.1038/aps.2011.184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang H, Chen C. Cyclooxygenase-2 in synaptic signaling. Current pharmaceutical design. 2008;14:1443–1451. doi: 10.2174/138161208784480144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Q, Wang Y, Feng J, Cao J, Chen B. Intermittent hypoxia from obstructive sleep apnea may cause neuronal impairment and dysfunction in central nervous system: the potential roles played by microglia. Neuropsychiatric disease and treatment. 2013;9:1077–1086. doi: 10.2147/NDT.S49868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang SX, Wang Y, Gozal D. Pathological consequences of intermittent hypoxia in the central nervous system. Comprehensive Physiology. 2012;2:1767–1777. doi: 10.1002/cphy.c100060. [DOI] [PubMed] [Google Scholar]
- Zudaire E, Portal-Nunez S, Cuttitta F. The central role of adrenomedullin in host defense. Journal of leukocyte biology. 2006;80:237–244. doi: 10.1189/jlb.0206123. [DOI] [PubMed] [Google Scholar]