

Abstract
Background
Macrophages play critical roles in liver regeneration, fibrosis development and resolution. They are among the first responders to liver injury and are implicated in orchestrating the fibrogenic response via multiple mechanisms. Macrophages are also intimately associated with the activated hepatic progenitor cell (HPC) niche or ductular reaction that develops in parallel with fibrosis. Among the many macrophage-derived mediators implicated in liver disease progression, a key role for macrophage-derived Wnt proteins in driving pro-regenerative HPC activation towards a hepatocellular fate has been suggested. Wnt proteins, in general, however, have been associated with both pro- and anti-fibrogenic activities in the liver and other organs. We investigated the role of macrophage-derived Wnt proteins in fibrogenesis and HPC activation in murine models of chronic liver disease by conditionally deleting Wntless expression, which encodes a chaperone essential for Wnt protein secretion, in LysM-Cre-expressing myeloid cells (LysM-Wls mice).
Results
Fibrosis and HPC activation were exacerbated in LysM-Wls mice compared to littermate controls, in the absence of an apparent increase in myofibroblast activation or interstitial collagen mRNA expression, in both the TAA and CDE models of chronic liver disease. Increased Epcam mRNA levels paralleled the increased HPC activation and more mature ductular reactions, in LysM-Wls mice. Increased Epcam expression in LysM-Wls HPC was also observed, consistent with a more cholangiocytic phenotype. No differences in the mRNA expression levels of key pro-inflammatory and pro-fibrotic cytokines or the macrophage-derived HPC mitogen, Tweak, were observed. LysM-Wls mice exhibited increased expression of Timp1, encoding the key Mmp inhibitor Timp1 that blocks interstitial collagen degradation, and, in the TAA model, reduced expression of the anti-fibrotic matrix metalloproteinases, Mmp12 and Mmp13, suggesting a role for macrophage-derived Wnt proteins in restraining fibrogenesis during ongoing liver injury.
Conclusion
In summary, these data suggest that macrophage-derived Wnt proteins possess anti-fibrogenic potential in chronic liver disease, which may be able to be manipulated for therapeutic benefit.
Electronic supplementary material
The online version of this article (doi:10.1186/s13069-015-0036-7) contains supplementary material, which is available to authorized users.
Keywords: Liver fibrosis, Ductular reaction, Macrophages, Matrix remodelling, TAA, CDE
Background
Regardless of their aetiology, chronic liver diseases (CLDs) share a common pathological mechanism, liver-injury stimulated fibrosis, which, if progressive, can lead to cirrhosis, liver failure, and cancer. The inflammatory response to chronic injury, mediated by multiple parenchymal and non-parenchymal cells, plays critical roles in fibrogenesis. Like inflammation, fibrosis is an adaptive response to liver injury that, when appropriately regulated, facilitates tissue repair. How these protective responses become maladaptive and the interplay between fibrogenesis and repair, in the context of chronic injury and inflammation, is not well understood. Clinical observations and experimental models suggest that liver fibrosis is a dynamic, bidirectional process with the capacity for recovery even at advanced stages [1]. With no therapies available to prevent or treat liver fibrosis, better understanding of the mechanisms of fibrosis and repair and the role of inflammatory cells and mediators is crucial to developing new therapeutic approaches.
In a healthy liver, injured parenchymal cells can be regenerated through division of mature epithelial cells (hepatocytes and bile duct-forming cholangiocytes). Following severe or chronic injury, hepatocyte regeneration is impaired and an alternative regenerative pathway involving hepatic progenitor cells (HPCs) can be activated, which is associated with the development of ductular reactions (DRs) and progressive fibrosis, both in humans and mouse models of CLD [2–4]. HPCs are thought to be bipotential, capable of proliferation and differentiation into hepatocytes, to replace injured cells or along the ‘ductular’ cholangiocyte lineage. Although capable of differentiation into hepatocytes and cholangiocytes under clonogenic conditions in vitro, the extent to which HPCs directly replace epithelial cells to regenerate the liver is controversial [5–7]. The contribution of HPCs to liver regeneration in mouse models likely depends on the extent to which epithelial self-renewal is impaired. Indeed, it was recently demonstrated that HPC activation in the setting of near total hepatocyte death or senescence was necessary for survival [7]. DRs correlate with the extent of liver fibrosis and may be postulated to represent a failed attempt to regenerate the liver, but the causal relationship between DRs and fibrosis remains to be established. HPCs are intimately associated with fibrogenic activity, however, as they exist in a niche composed of myofibroblasts, macrophages and other leukocytes and extracellular matrix components. Both cellular and matrix components of the niche shape HPC activation and fate, and HPCs, in turn, influence myofibroblasts and inflammatory cells.
Macrophages play critical roles in liver regeneration, fibrosis development and resolution and are prominent features of the fibrotic scar and DR niche [8–11]. Macrophages have been shown to play a dichotomous role in the context of liver fibrosis, as they play a pro-fibrogenic role during the development of fibrosis whereas macrophage expression of matrix metalloproteinases and pro-angiogenic factors crucially contributes to the resolution of fibrosis during recovery from liver injury [10, 12] and potentially even during ongoing injury [13]. Macrophage-derived factors, including Wnt proteins, are also directly implicated in controlling HPC proliferation, differentiation and invasion into the parenchyma [14–16]. Macrophage ablation reduced DR proliferation and expansion in several models of CLD [10, 16] and, strikingly, injection of macrophages caused transient TWEAK-dependent HPC activation in healthy mice [15].
HPC activation during injury appears to recapitulate liver epithelial cell development from hepatoblasts during embryogenesis and involves reactivation of developmental cell signalling pathways, including Wnt, Notch and Hedgehog [17]. Wnt signalling drives hepatocellular differentiation during development [18], and a similar function for Wnt proteins (‘Wnts’) during chronic liver injury has been suggested [14]. In contrast, macrophage-derived Wnt7b was recently shown to drive cholangiocarcinoma growth [19], highlighting the complex role of Wnt signalling in chronic liver disease. Inflammatory cytokines (e.g. IL-6, TNF, LTβ, IFNγ) [20–23], injury-induced growth factors (e.g. TWEAK [24]) and extracellular matrix components (e.g. collagen, laminin and fibronectin [25, 26]) are also implicated in regulating HPC proliferation and fate.
Wnts comprise a family of 19 secreted glycoproteins that activate target cells via β-catenin and β-catenin-independent pathways to regulate cell proliferation, differentiation and migration during development and homeostasis, including in multiple adult stem cell niches [18]. The pleiotropic targets and biological effects of Wnt signalling in the liver are not well understood. A loss-of-function mutation in the LRP6 gene, encoding a Wnt co-receptor, was recently associated with human non-alcoholic fatty liver disease [27]. Introduction of this mutation in mice caused steatohepatitis via a β-catenin-independent pathway, which was rescued by therapeutic administration of Wnt3a [27]. β-catenin signalling is implicated in liver repair and fibrogenesis, but the extent to which this is Wnt-dependent is not clear, as β-catenin stabilisation also occurs downstream of non-Wnt growth factors, such as hepatocyte growth factor [28]. In patients and mice with acute liver failure, β-catenin activation correlated with increased liver regeneration and (in patients) precluded the need for transplantation [29]. Multiple studies in rodent models of CLD have also suggested a role for β-catenin signalling in chronic liver injury, including driving HPC proliferation and differentiation [30–34]. By contrast, Wnt/β-catenin signalling in fibroblasts was reported to be pro-fibrotic in systemic sclerosis, skin, lung, liver and kidney disease [35–38]. The cellular source and type of Wnt proteins expressed in response to injury and their target cells may determine whether tissue regeneration or fibrosis occurs. Macrophage- but not epithelial-derived Wnts contributed to liver regeneration after partial hepatectomy [39]; however, the role of macrophage-derived Wnts in chronic liver disease, in particular in the HPC niche, is unknown. In the current study, we investigated the contribution of macrophage-derived Wnts to HPC activation and fibrogenesis during chronic liver disease. To do so, we examined the phenotype of mice with myeloid-targeted deletion of Wntless (Wls), encoding the chaperone Wls required for Wnt secretion [40]. Mice with Wls-deficient macrophages exhibited enhanced ductular reactions and fibrosis in two murine models of chronic liver disease. Our data are consistent with roles for macrophage-derived Wnts in HPC maturation and matrix remodelling to restrain net collagen deposition during chronic liver disease progression.
Results
Myeloid-specific Wls knockout does not affect liver macrophage abundance or localisation
To investigate the contribution of macrophage-derived Wnt proteins to inflammation, HPC activation and fibrosis, we generated mice deficient in Wls expression in myeloid cells by crossing Wls-loxp mice [40] with LysM-Cre mice [41] for 4–10 generations. Wls knockout in macrophages was confirmed by qPCR for Wls in bone marrow-derived macrophages (Additional file 1: Figure S1A). LysM-Wls (Cre+) mice and littermate controls (Cre-) were administered thioacetamide (TAA, 300 mg/L) in drinking water for 12 weeks as a model of chronic liver injury. Reduced Wls expression in F4/80+ liver cells sorted by flow cytometry was confirmed by qPCR (Additional file 1: Figure S1B–E). By contrast, Wls expression in Epcam+ cells, sorted by flow cytometry, was comparable between untreated Cre+ and Cre− animals, confirming the specificity of targeting (Additional file 1: Figure S1F). Consistent with a recent report [39], LysM-Wls mice were viable and livers were histologically normal. Wls deficiency did not affect the number or localisation of liver macrophages in control or TAA-treated mice, as assessed by immunohistochemistry and image analysis for F4/80 and qPCR for Emr1 (encoding the F4/80 antigen) in whole liver (Fig. 1). To determine which Wnt proteins are expressed by liver macrophages, we conducted a PCR for a panel of Wnt ligands. Wnt3a, Wnt5a, Wnt7a, Wnt7b and Wnt1 mRNAs were below the limit of detection in F480+ cells from control or TAA-treated mice; however, Wnt4 and Wnt6 were robustly detected but not significantly altered by TAA treatment or genotype (Additional file 1: Figure S1G–H).
Fig. 1.
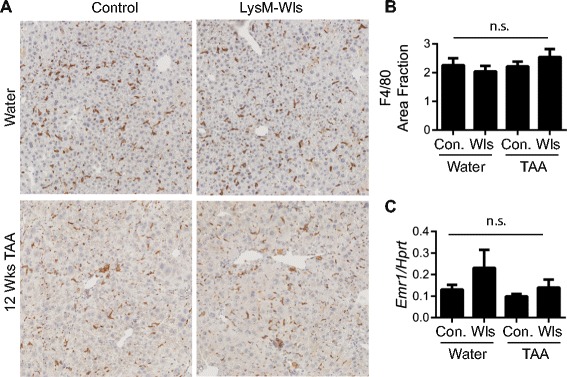
LysM-Wls depletion does not affect macrophage abundance and localisation. F4/80+ liver macrophage localisation (representative stain, 5× original magnification) (a). Quantification of F4/80+ staining (b) and whole liver Emr1 (encoding the F4/80 antigen) expression (c) in control and LysM-Wls mice treated with water or TAA for 12 weeks. Data represent mean + SEM from 5 to 12 mice from 4 independent experiments. n.s. non-significant
Reduction in Wnt/β-catenin signalling during liver injury in mice with Wls-deficient macrophages
In healthy livers, β-catenin localised to hepatocyte membranes and bile duct epithelia, whereas after 12 weeks TAA treatment, β-catenin stained hepatocyte membranes and some nuclei, as well as HPC, in both LysM-Wls mice and littermate controls (Fig. 2a). Whilst no clear difference in β-catenin expression or localisation in injured mice was discernible, expression of the Wnt/β-catenin target gene, Axin2, was elevated in livers from TAA-treated control mice but not LysM-Wls mice (Fig. 2b), consistent with a reduction in Wnt/β-catenin signalling in LysM-Wls mice during liver injury. Whole liver expression of the Wnt/β-catenin target genes Ccnd2 and Numb, which has previously been suggested to contribute to HPC hepatocellular fate determination [14], was not significantly different between genotypes, although there was a trend towards reduced Numb expression in LysM-Wls mice (Fig. 2c and data not shown).
Fig. 2.
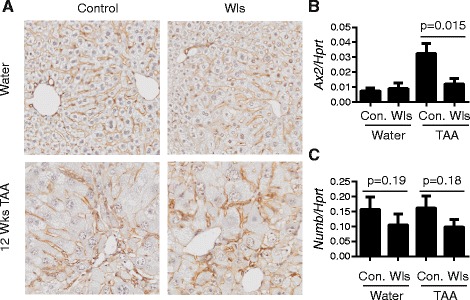
Reduced Wls expression in macrophages from LysM-Wls mice is associated with reduced β-catenin target gene expression. a β-Catenin staining (representative, 10× original magnification) and whole liver expression of b Axin2 and c Numb in control and LysM-Wls mice treated with water or TAA for 12 weeks. Data represent mean+/−SEM of 5–12 mice from 4 independent experiments
Wls depletion in macrophages exacerbates TAA-induced hepatic progenitor cell activation and fibrosis
After 12 weeks of TAA treatment, LysM-Wls mice exhibited increased HPC activation, frequently associated with mature ductular reactions, which were not observed in littermate controls (pan-keratin staining, Fig. 3a, b). This was associated with increased fibrosis (Picro-Sirius red staining, Fig. 3c–e). Female mice were used in this study because we observed that TAA-induced HPC activation and fibrosis were more severe in females than males, however Wls deficiency in macrophages similarly exacerbated the DR and fibrosis in male mice (Additional file 2: Figure S2). The increased DR and fibrosis was not explained by increased susceptibility to TAA-induced injury, as liver injury was comparable in LysM-Wls mice and littermate controls, as assessed by serum ALT and liver histology (Fig. 4a–c). The expression of the key pro-inflammatory cytokines Tnf and Il6 that are produced by macrophages and other cell types in response to liver injury and play critical roles in liver inflammation, fibrosis and HPC activation [20, 22, 42] was not significantly different between LysM-Wls and littermate controls (data not shown). Despite the increase in fibrosis in LysM-Wls mice, mRNA levels of the key interstitial collagens contributing to liver fibrosis [43], Col1a1, Col1a2 and Col3a1, were comparable between LysM-Wls and littermate controls (Fig. 5a–c). Although mRNA expression of Sma, which encodes smooth muscle actin, was comparable between genotypes (data not shown), there was a trend towards increased Sma protein expression in LysM-Wls mice (Fig. 5d, e), suggesting an increase in myofibroblast activation in these mice. Nevertheless, mRNA expression of the pro-fibrogenic cytokines Tgfb and Ctgf [44] that play key roles in myofibroblast activation did not differ between genotypes (Fig. 5f, g).
Fig. 3.
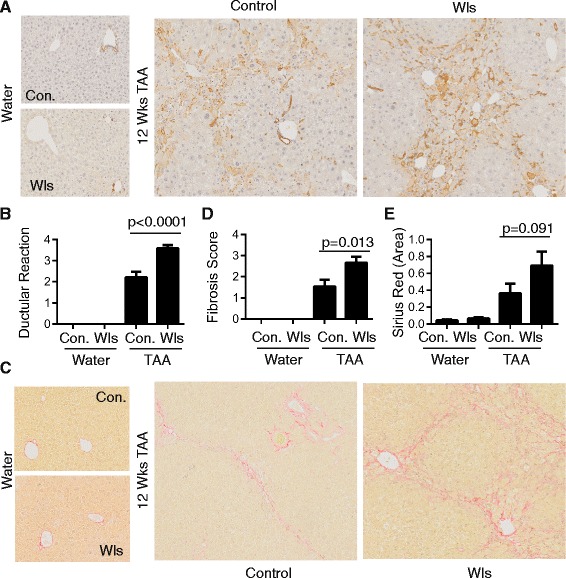
Increased ductular reaction and fibrosis in LysM-Wls mice. a Representative CKWSS staining and b ductular reaction grade in livers from 12-week water- and TAA-treated control and LysM-Wls mice. c Representative Picro-Sirius Red staining, d fibrosis score and e collagen proportionate area in livers from water- and 12-week TAA-treated control and LysM-Wls mice. Data represent mean + SEM of 8–17 mice from 4 independent experiments
Fig. 4.
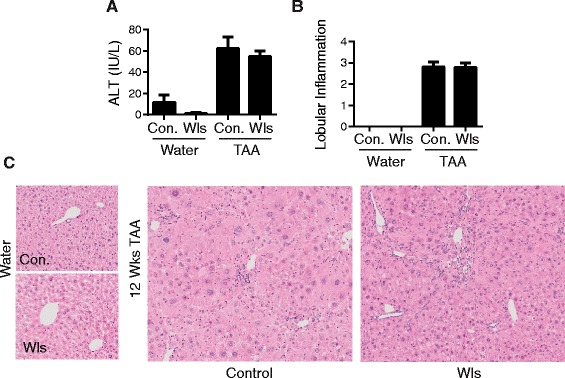
Comparable injury and inflammation in LysM-Wls and control mice. a Serum ALT levels in water- and 12-week TAA-treated control and LysM-Wls mice. Data represent mean+/−SEM of 8–10 mice from 2 independent experiments. b Lobular inflammation score and c representative H&E staining, in water- and 12-week TAA-treated control and LysM-Wls mice. Data represent mean + SEM of 8–17 mice from 4 experiments
Fig. 5.
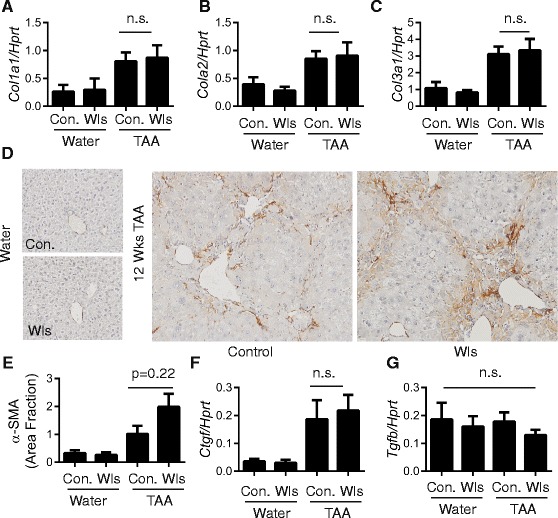
Comparable myofibroblast activation in LysM-Wls and control mice. Whole liver expression of a Col1a1, b Col1a2 and c Col3a1 in water- and 12-week TAA-treated control and LysM-Wls mice. d Representative SMA staining and e quantification of SMA staining (mean + SEM, n = 5 per group) in livers from water- and 12-week TAA-treated control and LysM-Wls mice. Whole liver expression of f Ctgf and g Tgfb in water- and 12-week TAA-treated control and LysM-Wls mice. Data represent mean + SEM of 8–17 mice from 4 independent experiments. n.s. not significant
Hepatic progenitor cells express an immature profile in mice with Wls-deficient macrophages
Paralleling the ductular reaction, Epcam (Tacstd1) liver mRNA levels were elevated in LysM-Wls mice compared to littermate controls (Fig. 6a), whereas the expression of the biliary keratin, Krt19, did not differ. Epcam is a cell surface glycoprotein expressed on tissue stem cells with regenerative capacity and is normally downregulated upon terminal differentiation [45]. To investigate the effects of macrophage-derived Wnts on HPC gene expression, we sorted Epcam+ cells from the liver non-parenchymal fraction to enrich for HPC. Expression of Epcam itself was increased in Epcam+ cells from TAA-treated LysM-Wls mice compared to littermate controls (Fig. 6b). Trop2 (Tacstd2), which encodes an Epcam-related protein reported to be specifically induced in HPC upon liver injury [46], was elevated in TAA-treated control mice and hyper-induced in LysM-Wls mice (Fig. 6c). We investigated the expression of the Notch target gene Hey1 and the Wnt/β-catenin target genes Axin2 and Numb in Epcam+ cells, as the Notch and Wnt/β-catenin pathways have previously been suggested to have opposing roles in determining HPC fate [14, 17], to provide evidence for direct effect of macrophage-derived Wnts on HPC, however these genes were only marginally detected in sorted cells (data not shown).
Fig. 6.
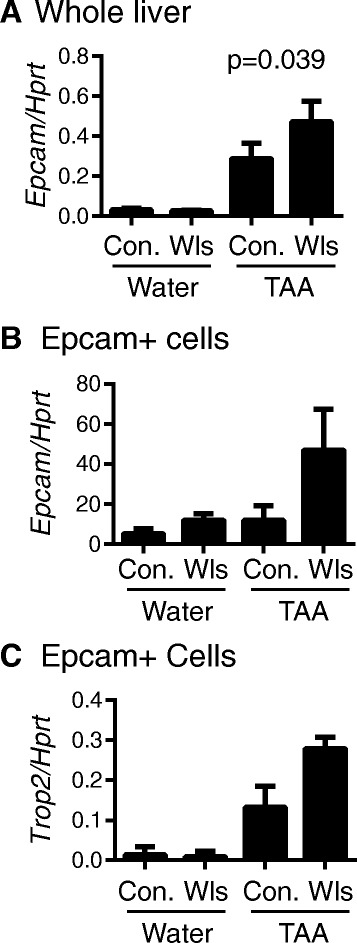
Increased Epcam expression in Epcam+ cells from LysM-Wls mice. a Whole liver expression of Epcam in water- and 12-week TAA-treated control and LysM-Wls mice (mean + SEM of 8–17 mice from 4 independent experiments). Expression of b Epcam and c Trop2 in flow cytometry-sorted Epcam+ cells (pooled from 3 to 6 mice) from water- and 12-week TAA-treated control and LysM-Wls mice (mean + range of 2 independent experiments)
Macrophage metalloproteinase expression is reduced in TAA-treated LysM-Wls mice
Macrophage production of the HPC mitogen, Tweak, has been implicated in driving HPC proliferation [15, 24]. Although TAA inducible, Tweak expression did not differ in whole liver or flow cytometry-sorted F4/80+ cells from control and LysM-Wls mice (Fig. 7a, b). As in whole liver, expression of the pro-inflammatory cytokines, Il6 and Tnf, and the pro-fibrotic cytokine, Tgfb, did not differ between LysM-Wls and control macrophages (data not shown). Since macrophages play a key role in matrix remodelling, we also examined the expression of key matrix remodelling factors implicated in liver fibrosis and repair [47]. We found myeloid deletion of Wls impaired injury-mediated upregulation of Mmp13 (encoding the key interstitial collagenase in rodents, Mmp13) and Mmp12 (encoding the elastase Mmp12) mRNAs in whole liver and purified F4/80+ cells (Fig. 7c–f). Mmp2 and Mmp8 were induced in TAA-injured livers but not differentially expressed in Wls-deficient and control mice, whereas no alteration in liver or macrophage Mmp9 expression was observed (data not shown). Furthermore, in TAA-treated LysM-Wls mice, the expression of Timp1, encoding the key Mmp inhibitor Timp1 that blocks interstitial collagen degradation [1], was higher compared to littermate controls (Fig. 7g). These data are consistent with the hypothesis that fibrolysis, as well as fibrogenesis, is dynamically regulated during TAA-mediated liver injury and suggest that macrophage-derived Wnt proteins promote matrix remodelling, via Mmp13 and Mmp12, potentially restraining fibrosis.
Fig. 7.
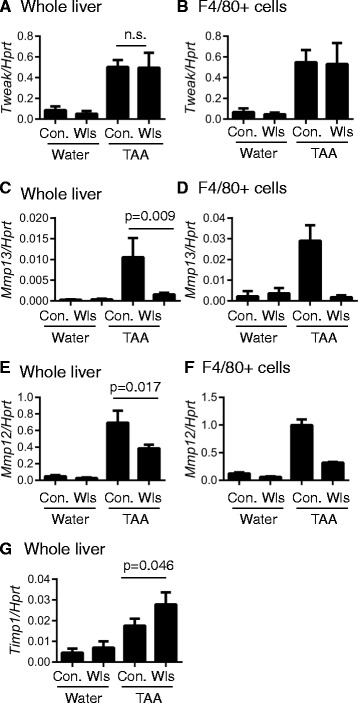
Reduced macrophage matrix metalloproteinase expression in LysM-Wls mice. Tweak expression in a whole liver and b flow cytometry-sorted macrophages. Mmp13 expression in c whole liver and d flow cytometry-sorted macrophages, Mmp12 expression in e whole liver and f flow cytometry-sorted macrophages and g whole liver expression of Timp1 in water- and 12-week TAA-treated control and LysM-Wls mice. Data represent mean + SEM of 8–17 mice (whole liver) or mean + range of 2 independent experiments (sorted macrophages pooled from 3 to 6 mice in each experiment)
Wls depletion in macrophages exacerbates CDE-induced hepatic progenitor cell activation and fibrosis
To determine whether macrophage-derived Wnts contribute to the regulation of HPC activation and fibrogenesis in multiple contexts of liver injury, we employed an alternative model of liver injury and fibrosis. The choline-deficient ethionine-supplemented (CDE) diet induces a robust DR but a different pattern of fibrosis to TAA [48]. Liver injury and fibrosis were also more severe in female mice than male mice in this model (data not shown). Similar to the TAA model, HPC activation and fibrosis were exacerbated in CDE-treated LysM-Wls mice compared to littermate controls, in the absence of alterations in whole liver Col1a1, Col1a2, Col3a1, Sma and Il6 expression between LysM-Wls mice and littermate controls (Additional file 3: Figure S3A–E and data not shown). Epcam expression was elevated in CDE-treated LysM-Wls mice, in whole liver and Epcam+ cells (Additional file 3: Figure S3F, G), suggesting similar alterations in HPC phenotype in both models. As in the TAA model, Timp1 mRNA expression was increased in CDE-treated LysM-Wls mice compared to littermate controls (Additional file 3: Figure S3I); however, Mmp13 was not detectable in the majority of CDE-treated mice and there was no difference in Mmp12, Mmp8, Mmp2 or Mmp9 expression (data not shown). Wnt4 and Wnt6 were expressed in macrophages from CDE-treated mice, regardless of genotype (data not shown).
Discussion
Macrophages contribute to the dynamic interplay between liver inflammation, fibrosis and regeneration via multiple mechanisms, at different locations and different disease stages. Inflammatory, bone marrow-derived monocytes infiltrate the liver in response to injury, where they are dynamically reprogrammed, depending on the nature and chronicity of the injury [10, 49–52]. Macrophages are among the earliest responders to liver injury, having roles in phagocytosis of apoptotic and necrotic cells and releasing soluble mediators, such as TNF and IL-6, that orchestrate inflammatory and ultimately pro-fibrogenic responses to chronic injury. Macrophages are also prominent components of the hepatic progenitor cell niche, which is closely associated with liver fibrogenesis [2, 8, 9] and repair; although the fate relationships among hepatocytes, cholangiocytes and HPC and the contribution of HPC to liver fibrosis and regeneration are controversial [5–7, 53–55]; and the factors determining their activation and differentiation are not well understood. Wnt proteins, potentially derived from macrophages, have been suggested to promote liver regeneration by driving HPC hepatocellular differentiation [14, 17, 32], which could theoretically ameliorate liver fibrosis by promoting regeneration. Wnt proteins, however, constitute a large family of proteins that have been variously attributed pro- and anti-fibrogenic properties [35–38] and which likely have multiple potential target cells and mechanisms of action during chronic liver disease. We investigated the role of macrophage-derived Wnt proteins in fibrogenesis in two models of chronic liver disease and demonstrate that ablating Wls expression in myeloid cells exacerbated HPC activation and fibrosis.
Although the role of β-catenin in liver homeostasis and injury has been extensively studied [18], the contribution and sources of specific β-catenin-dependent Wnt proteins are little studied, and β-catenin-independent Wnt signalling is even less studied [27, 32, 39]. β-catenin is essential for hepatocyte-mediated liver regeneration after partial hepatectomy, and it was recently shown that macrophage- but not epithelium-derived Wnt proteins contribute to β-catenin activation in this setting [39]. A novel, β-catenin-dependent population of self-renewing hepatocytes maintained by endothelial-derived Wnts was also recently identified, although the role of these cells in liver injury has not yet been investigated [56]. In humans with acute liver failure, nuclear β-catenin localisation in hepatocytes correlated with patient survival and reduced requirement for liver transplant [29], consistent with a pro-regenerative role for β-catenin, although Wnt proteins were not directly implicated in this study. Interestingly, β-catenin depletion from mature hepatocytes in healthy mice caused HPC expansion, and hepatocytes were gradually replaced by β-catenin-positive HPC [57]. β-catenin has also been implicated in HPC-mediated liver regeneration in murine chronic liver disease [30, 33], and hepatocyte-derived Wnt1 promoted HPC differentiation in rats, following partial hepatectomy and 2-acetylaminofluorene (AAF) treatment [32]. Our data demonstrating increased fibrosis and associated with more extensive ductular reactions and a more cholangiocytic HPC phenotype (increased Epcam and Trop2 expression) in LysM-Wls mice are consistent with a role for Wnt proteins in protecting from fibrosis by promoting HPC hepatocellular differentiation. However, we did not observe any difference in β-catenin localisation in livers from LysM-Wls and control mice. Whilst the β-catenin target gene Axin2 was reduced in whole liver in LysM-Wls mice, it was below the limit of detection in Epcam+ cells (which include HPC), so we could not ascertain whether this pathway was differentially activated in these cells in the absence of macrophage-derived Wnts. It is possible that cells with activated β-catenin signalling are not represented among the sorted population due to maturation-associated Epcam downregulation [45]. Alternatively, rather than representing a block in hepatocellular differentiation, the expanded DR observed in LysM-Wls mice could result from a pro-proliferative drive, via the activation of Hedgehog or other growth factors [58, 59], the persistence of collagen or altered matrix composition [11], or from hepatocyte metaplasia, rather than HPC activation [54].
In addition to their potential direct (or indirect) effects on HPC, macrophage-derived Wnts could influence HPC activation and fibrosis via additional target cells and mechanisms. Wnt/β-Catenin signalling, for example, has been implicated in promoting stellate cell quiescence [60]; but we did not observe any difference in SMA expression or key cytokine regulators of myofibroblast activation, in LysM-Wls and control mice. In both the TAA and CDE models, increased hepatic collagen deposition in LysM-Wls mice occurred in the absence of increased expression of the key interstitial collagens, Col1a1, Col2a1 and Col3a1. In contrast to a typical pro-fibrogenic signature, with evidence of increased myofibroblast activation and collagen production, Timp1 mRNA, encoding a key hepatic Mmp inhibitor that prevents remodelling of the fibrotic scar [61, 62], was increased in LysM-Wls mice compared to littermate controls in both the TAA and CDE models. Moreover, in the TAA model, macrophage Mmp13 and Mmp12 expression was reduced in LysM-Wls mice. These data suggest the hypothesis that macrophage-derived Wnts promote fibrolysis, which dynamically counter-balances collagen accumulation, even during active fibrogenesis. The contribution of matrix remodelling to fibrogenesis during ongoing liver injury is not well understood, but studies altering the Timp to Mmp balance have consistently confirmed the potent influence of this ratio on the development and resolution of liver fibrosis [1]. Macrophages clearly play an essential role in the resolution of established fibrosis, after cessation of liver injury [10, 51, 52]. This involves reprogramming recruited monocytes/macrophages to a pro-resolution phenotype, including increased phagocytic activity and increased Mmp12 and Mmp13 expression, in conjunction with loss of liver Timp1 mRNA and protein expression [52]. Interestingly, administration of bone marrow-derived macrophages during ongoing toxic injury was sufficient to ameliorate fibrosis, and this was associated with increased Mmp13 expression [13]. Similarly, Barnes and colleagues recently reported a reduced rate of collagen degradation, associated with reduced recruitment of macrophages to the fibrotic scar, and reduced Mmp13 activity during CCL4-induced liver injury in mice lacking the chemokine MIF [63]. Mmp and Timp functions are not, however, limited to matrix proteolysis. Timp1 inhibits myofibroblast apoptosis [64], for example, whilst Mmp13 could contribute to fibrosis resolution via activation of latent hepatocyte growth factor and multiple downstream mechanisms [65–69]. Investigating the dynamic remodelling of multiple matrix components during chronic injury will be important to determine the functional relevance of the Timp1/Mmp balance in different models at different stages of disease progression.
Conclusions
We have shown for the first time that the net effect of myeloid-derived Wnt proteins in chronic liver disease is to dampen HPC activation and fibrogenesis. Multiple Wnt proteins acting on different target cells may contribute to this phenotype, including direct effects on HPC fate, collagen-producing myofibroblasts and macrophages. Whilst Wnt induction, particularly leading to β-catenin activation, is not a viable therapeutic strategy in the setting of chronic liver disease, due to the risk of cancer development [70]; elucidation of the downstream pathways mediating Wnt proteins’ anti-fibrogenic effects may reveal novel, specific therapeutic targets. Most significantly, however, these data suggest that pro- and anti-fibrogenic macrophage functions co-exist during disease progression and support the notion that reprogramming macrophages or otherwise enhancing anti-fibrogenic pathways may be a viable therapeutic strategy to ameliorate fibrogenesis during chronic liver disease.
Methods
Transgenic mice and in vivo models
All animal experiments were performed with approval from the University of Queensland Animal Ethics Committee (MED/PAH/156/13/PAHRF/NHMRC, UQDI/571/12/NHMRC/AIDRCC). To conditionally delete Wls from macrophages, homozygous Wlsflox/flox mice [40] were crossed with LysM-Cre transgenic mice [41] for 4–10 generations. Offspring with the genotype Wlsflox/flox; LysM-Cre-positive represents Wls macrophage knockouts, whilst Wlsflox/flox; LysM-Cre-negative was used as controls. Six to 9-week-old mice were administered with 30 mg/L thioacetamide (TAA, Sigma) in drinking water for 12 weeks [71] or a modified choline-deficient ethionine-supplemented (CDE) diet for 6 weeks [48], to induce chronic liver injury and fibrosis. The modified CDE diet was optimised by Professor George Yeoh [48] (UWA, Australia) and was custom made by MP Biosciences (Santa Ana, USA). The diet consisted of 70 % choline-deficient diet (Cat#0296021410) and 30 % control (choline-sufficient) diet (Cat#0296041210). At the end of the treatment period, mice were euthanised and a blood sample taken for serum isolation. Livers were perfused with 10 ml of PBS via the portal vein in situ prior to tissue harvest to minimise blood contamination. Portions of the liver were fixed in 10 % neutral buffered formalin and embedded in paraffin or homogenised in TRI reagent and stored at −80 °C for RNA isolation. The remainder of the liver was used to isolate non-parenchymal cells. Serum ALT levels were measured using the MaxDiscovery Alanine Transaminase Color Endpoint Assay (Bioo Scientific, Austin, TX, USA).
Non-parenchymal cell isolation
Non-parenchymal cells were isolated by finely dissociating the liver tissue using surgical scissors then passing the fragments through a stainless steel strainer with the addition of 14 ml of Hank’s Based Salt Solution (HBSS) containing 1 mg/ml type 4 collagenase (Worthington Biochemical corporation, Lakewood, NJ, USA) 1 μg/ml DNase I (Sigma), 100 U/ml penicillin and 100 μg/ml streptomycin. The cell suspension was filtered through a 70-μM filter (Becton Dickinson) and incubated at 37 °C for 5 min, prior to the addition of 10 mM EDTA and a further 5-min incubation period. The lysate was washed twice in cold PBS containing 2 % FBS (FACS buffer) and resuspended in 25 ml of 33 % Percoll in PBS (isotonic) and centrifuged at 600 × g for 15 min with no brake. The supernatant was discarded and the pellet washed twice and resuspended in FACS buffer. Freshly isolated non-parenchymal cells (~5 × 106) per liver were stained with anti-F4/80-PE (clone BM8) and anti-Epcam-APC (clone G8.8) (Biolegend) antibodies for 30 min at room temperature, washed twice and resuspended in FACS buffer for sorting. PE- and APC-positive cells were sorted using an Astrios Moflo, achieving >90 % purity. Sorted cells were resuspended in TRI reagent (Sigma) for RNA purification.
RNA purification and qPCR
RNA from whole liver (5 μg) or sorted cells (entire yield) was transcribed to cDNA using oligodT priming and Superscript III (Life Technologies). Quantitative real-time PCR (qPCR) for genes of interest was performed using SYBR green (ABI) on an HT9000 or VIA-VII cycler with default cycle settings. Relative expression was determined using Hprt as housekeeping gene and the delta-Ct method and expressed as Target/Hprt. Primer sequences used in this study were the following: Hprt-F: TGCTGGATTACATCAAAGCACTG, Hprt-R: CCCCTGTTGACTGGTCATTACAA; Ccnd1-F CACAACGCACTTTCTTTCCAGA Ccnd1-R CTTGACTCCAGAAGGGCTTCAA Numb-F CAATGAGTTGCCTTCCACTATGC Numb-R ATCTGGGAACACAAGGAGCTGA Mmp12-F TACCCCAAGCTGATTTCCACAC Mmp12-R GCTCCTTGGAAGATGTAGTAGTGTCTTT Mmp13-F ACAAAGATTATCCCCGCCTCAT Mmp13-R GGCCCATTGAAAAAGTAGATATAGCC Timp1-FAAGGGCTAAATTCATGGGTTCC Timp1-R ACAGCCTTGAATCCTTTTAGCATC Mmp9-F AGGGGCGTGTCTGGAGATTC Mmp9-R TCCAGGGCACACCAGAGAAC Tweak-F CGCTCTTAGTCTGGTCCTGGTTT Tweak-R CACCAGTCTCCTCTATGGGGGTA Tgfb-F GTGGCTGAACCAAGGAGACG Tgfb-R GGCTGATCCCGTTGATTTCC F480-F CTGTCTGCTCAACCGTCAGGTA F480-R AGAAGTCTGGGAATGGGAGCTAA E Epcam-F CGGGGATTGTTGTCCTGGTT Epcam-R GCACGGCTAGGCATTAAGCTC Trop2-F AGGAGCTGGGGGAGATGAGA Trop2-R CCAACCCATCTGGTCTGAGG Krt19-F CCTAGCCAAGATCCTGAGTGAGAT Krt19-R TGGGTGTTCAGCTCCTCAATC Ctgf-F GTCAAGCTGCCTGGGAAATG Ctgf-R AATGTGTCTTCCAGTCGGTAGGC Il6-F GCTGGTGACAACCACGGCCT Il6R GGCATAACGCACTAGGTTTGCCG Tnf-F AGGGGCCACCACGCTCTTCT Tnf-R CGGGGCAGCCTTGTCCCTTG Wls-F CAAATCGTTGCCTTTCTGGTG Wls-R TTGTCACACTTGTTAGGTCCC Wnt4-F AAGAGGAGACGTGCGAGAAA Wnt4-R CACCACCTTCCCAAAGACAG Wnt6-F TCAAGACTCTTTATGGATGCGC Wnt6-R ATGGCACTTACACTCGGTG Sma-F GATCCTGACTGAGCGTGGCTAT Sma-R CGTGGCCATCTCATTTTCAAAG mMmp2-F TTGCAGGAGACAAGTTCTGGAGATA mMmp2-R CACGACGGCATCCAGGTTAT mMmp8-F GGTTACCCCAAAAGCATACCAA mMmp8-R CACTGAAGAAGAGGAAGAAGGAGTC mCol1a2-F CACCCCAGCGAAGAACTCATAC mCol1a2-R CCCCTTCTACGTTGTATTCAAACTG mCol3a1-F TGGGATCAAATGAAGGCGAAT mCol3a1-R GCTCCATTCCCCAGTGTGTTTAG.
Histology, immunohistochemistry and image analysis
H&E and Sirius red stains were performed and histological changes (inflammatory infiltrate and fibrosis) were assessed by two experienced hepatopathologists (GCM and ADC). Fibrosis stage was assessed using a modified METAVIR score as previously described [72]; briefly, stage 1: peri-portal or centrilobular fibrosis, stage 2: some septa, stage 3: many septa, stage 4: cirrhosis. Deparaffinised liver tissue sections were immunolabelled as described previously [73]. Antibodies used in this study were cytokeratin wide-spectrum screening (CKWSS, DAKO), F4/80 (Abcam), SMA (1A4, DAKO), β-catenin (Cell Signalling). HPC activation/ductular reactions were graded from CKWSS-stained sections according to the following schema: 1: CKWSS+ cells surrounding <50 % of portal vein, 2: CKWSS+ cells surrounding >50 % of portal vein; 3: CKWSS+ cells forming septa covering <50 % distance between two structures, 4: CKWSS+ cells forming septa covering >50 % distance between two structures. SMA and F4/80 image analysis were performed using ImageJ from ten captured images per section, each representing an area 310 × 250 μm in size. Sirius Red image analysis was performed using the positive pixel count algorithm in Aperio ImageScope from Aperio AT slide scans, from an area of 5–10 mm2 for each liver specimen. The algorithm was optimised to detect red pixels (hue value 0, hue width 0.1 and colour saturation threshold 0.22). All portal tracts and central veins greater than 100 μm in size were removed as they contain a large amount of collagen and therefore prevent quantification of fibrosis-associated collagen.
Statistical analysis
Group comparisons were performed using the Mann–Whitney-U test. A two-tailed p value of 0.05 was considered statistically significant. All statistical analyses were performed using Prism Version 6 (GraphPad).
Acknowledgements
We wish to acknowledge Professor George Yeoh for advice on the CDE model and helpful discussions, Ms. Tam Ngyuen for genotyping macrophage analyses, and the Translational Research Institute Flow Cytometry and Biological Resources Facilities for technical assistance. This study was funded by project grants from the Princess Alexandra Hospital Foundation and the National Health and Medical Research Council (NHMRC), Australia. EEP is supported by an NHMRC Practitioner Fellowship and KMI is supported by an Australian Liver Foundation Fellowship.
Abbreviations
- CDE
Choline-deficient ethionine-supplemented
- CLD
Chronic liver disease
- DR
Ductular reaction
- HPC
Hepatic progenitor cells
- TAA
Thioacetamide
Additional files
Wls depletion and Wnt expression in Wls-loxp × LysM-Cre mice. (A) RT-PCR quantification of Wls expression in bone marrow-derived macrophages from Wlsfl/fl and LysM-Cre+/− mice (n = 4–5, Mann–Whitney-U test). (B) Flow cytometry gating strategy for sorting F4/80+ and Epcam+ cells from liver non-parenchymal cells. Post-sort analysis of (C) F4/80+ and (D) Epcam+ cells. RT-PCR quantification of Wls expression in sorted (E) F4/80+ and (F) Epcam+ cells from control and 12-week TAA-treated mice. RT-PCR quantification of (G) Wnt4 and (H) Wnt6 expression in sorted F4/80+ cells from control and 12-week TAA-treated mice. Sorted cells were pooled from 3 to 6 mice from control and LysM-Wls mice. Data represent mean + range of 2 independent experiments. (EPS 1589 kb)
Increased ductular reaction and fibrosis in male LysM-Wls mice compared to littermate controls. (A) Ductular reaction grade, (B) fibrosis score and (C) whole liver Col1a1 expression in livers from water- and 12-week TAA-treated control and LysM-Wls male mice. Data represent mean + SEM of 4–8 mice from 3 independent experiments. (EPS 478 kb)
Increased ductular reaction and fibrosis in LysM-Wls mice in the CDE model of chronic liver disease. Ductular reaction grade (A), fibrosis score (B) and whole liver expression of Col1a1 (D), Col3a1 (E), Sma (F), Epcam (G) and Timp1 (I) in livers from LysM-Wls and control mice fed with a CDE diet for 6 weeks. Data represent mean + SEM from n = 3 mice from 1 experiment, representative of 3 independent experiments. (H) Epcam expression in flow cytometry-sorted Epcam+ cells (pooled from 3 mice) from LysM-Wls and control mice treated fed with a CDE diet for 6 weeks. Data represent mean + range of PCR duplicates, representative of 3 independent experiments. (EPS 509 kb)
Footnotes
Antje Blumenthal and Elizabeth E. Powell contributed equally to this work.
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
KMI, AB and EEP conceived and designed the study. KMI, WW, VLG, MRM, MM, KM, MJS and RL developed techniques and/or produced data. KMI, ADC, GM and VLG analysed the data. RL and MJS provided essential reagents and critical review. KMI, ADC, AB and EEP wrote the manuscript. All authors read and approved the final manuscript.
Contributor Information
Katharine M. Irvine, Phone: +61 (7) 3443 8014, Email: katharine.irvine@uq.edu.au
Andrew D. Clouston, Email: AndrewClouston@envoi.com
Victoria L. Gadd, Email: v.gadd@uq.edu.au
Gregory C. Miller, Email: gregorymiller@envoi.com
Weng-Yew Wong, Email: Jeremy.wong@centenary.org.au.
Michelle Melino, Email: Michelle.Melino@qimrberghofer.edu.au.
Muralidhara Rao Maradana, Email: m.maradana@uq.edu.au.
Kelli MacDonald, Email: Kelli.MacDonald@qimrberghofer.edu.au.
Richard A. Lang, Email: Richard.Lang@cchmc.org
Matthew J. Sweet, Email: m.sweet@imb.uq.edu.au
Antje Blumenthal, Email: a.blumenthal@uq.edu.au.
Elizabeth E. Powell, Email: e.powell@uq.edu.au
References
- 1.Iredale JP, Thompson A, Henderson NC. Extracellular matrix degradation in liver fibrosis: biochemistry and regulation. Biochim Biophys Acta. 2013;1832(7):876–83. doi: 10.1016/j.bbadis.2012.11.002. [DOI] [PubMed] [Google Scholar]
- 2.Williams MJ, Clouston AD, Forbes SJ. Ductular reactions and fibrosis: what is the link and can we break it? Gastroenterology. 2013 (Submitted).
- 3.Gouw AS, Clouston AD, Theise ND. Ductular reactions in human liver: diversity at the interface. Hepatology. 2011;54(5):1853–63. doi: 10.1002/hep.24613. [DOI] [PubMed] [Google Scholar]
- 4.Williams MJ, Clouston AD, Forbes SJ. Links between hepatic fibrosis, ductular reaction, and progenitor cell expansion. Gastroenterology. 2014;146(2):349–56. doi: 10.1053/j.gastro.2013.11.034. [DOI] [PubMed] [Google Scholar]
- 5.Espanol-Suner R, Carpentier R, Van HN, Legry V, Achouri Y, Cordi S, et al. Liver progenitor cells yield functional hepatocytes in response to chronic liver injury in mice. Gastroenterology. 2012;143(6):1564–75. doi:10.1053/j.gastro.2012.08.024. [DOI] [PubMed]
- 6.Malato Y, Naqvi S, Schurmann N, Ng R, Wang B, Zape J, et al. Fate tracing of mature hepatocytes in mouse liver homeostasis and regeneration. J Clin Invest. 2011;121(12):4850–60. doi: 10.1172/JCI59261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lu WY, Bird TG, Boulter L, Tsuchiya A, Cole AM, Hay T, et al. Hepatic progenitor cells of biliary origin with liver repopulation capacity. Nat Cell Biol. 2015;17(8):971–83. doi: 10.1038/ncb3203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gadd VL, Melino M, Roy S, Horsfall L, O’Rourke P, Williams MR, et al. Portal, but not lobular, macrophages express MMP-9: association with the ductular reaction and fibrosis in chronic HCV. Liver Int. 2013;33(4):569–79. doi: 10.1111/liv.12050. [DOI] [PubMed] [Google Scholar]
- 9.Gadd VL, Skoien R, Powell EE, Fagan KJ, Winterford C, Horsfall L, et al. The portal inflammatory infiltrate and ductular reaction in human non-alcoholic fatty liver disease. Hepatology. 2013 doi: 10.1002/hep.26937. [DOI] [PubMed] [Google Scholar]
- 10.Duffield JS, Forbes SJ, Constandinou CM, Clay S, Partolina M, Vuthoori S, et al. Selective depletion of macrophages reveals distinct, opposing roles during liver injury and repair. J Clin Invest. 2005;115(1):56–65. doi: 10.1172/JCI200522675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lorenzini S, Bird TG, Boulter L, Bellamy C, Samuel K, Aucott R, et al. Characterisation of a stereotypical cellular and extracellular adult liver progenitor cell niche in rodents and diseased human liver. Gut. 2010;59(5):645–54. doi: 10.1136/gut.2009.182345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kantari-Mimoun C, Castells M, Klose R, Meinecke AK, Lemberger UJ, Rautou PE, et al. Resolution of liver fibrosis requires myeloid cell-driven sinusoidal angiogenesis. Hepatology. 2014 doi: 10.1002/hep.27635. [DOI] [PubMed] [Google Scholar]
- 13.Thomas JA, Pope C, Wojtacha D, Robson AJ, Gordon-Walker TT, Hartland S, et al. Macrophage therapy for murine liver fibrosis recruits host effector cells improving fibrosis, regeneration, and function. Hepatology. 2011;53(6):2003–15. doi: 10.1002/hep.24315. [DOI] [PubMed] [Google Scholar]
- 14.Boulter L, Govaere O, Bird TG, Radulescu S, Ramachandran P, Pellicoro A, et al. Macrophage-derived Wnt opposes Notch signaling to specify hepatic progenitor cell fate in chronic liver disease. Nat Med. 2012;18(4):572–9. doi: 10.1038/nm.2667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bird TG, Lu WY, Boulter L, Gordon-Keylock S, Ridgway RA, Williams MJ, et al. Bone marrow injection stimulates hepatic ductular reactions in the absence of injury via macrophage-mediated TWEAK signaling. Proc Natl Acad Sci U S A. 2013;110(16):6542–7. doi: 10.1073/pnas.1302168110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Van HN, Lanthier N, Espanol SR, Abarca QJ, van Rooijen N, Leclercq I. Kupffer cells influence parenchymal invasion and phenotypic orientation, but not the proliferation, of liver progenitor cells in a murine model of liver injury. Am J Pathol. 2011;179(4):1839–50. doi: 10.1016/j.ajpath.2011.06.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Spee B, Carpino G, Schotanus BA, Katoonizadeh A, Vander BS, Gaudio E, et al. Characterisation of the liver progenitor cell niche in liver diseases: potential involvement of Wnt and Notch signalling. Gut. 2010;59(2):247–57. doi: 10.1136/gut.2009.188367. [DOI] [PubMed] [Google Scholar]
- 18.Nejak-Bowen KN, Monga SP. Beta-catenin signaling, liver regeneration and hepatocellular cancer: sorting the good from the bad. Semin Cancer Biol. 2011;21(1):44–58. doi: 10.1016/j.semcancer.2010.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Boulter L, Guest RV, Kendall TJ, Wilson DH, Wojtacha D, Robson AJ, et al. WNT signaling drives cholangiocarcinoma growth and can be pharmacologically inhibited. J Clin Invest. 2015;125(3):1269–85. doi: 10.1172/JCI76452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Yeoh GC, Ernst M, Rose-John S, Akhurst B, Payne C, Long S, et al. Opposing roles of gp130-mediated STAT-3 and ERK-1/2 signaling in liver progenitor cell migration and proliferation. Hepatology. 2007;45(2):486–94. doi: 10.1002/hep.21535. [DOI] [PubMed] [Google Scholar]
- 21.Subrata LS, Lowes KN, Olynyk JK, Yeoh GC, Quail EA, Abraham LJ. Hepatic expression of the tumor necrosis factor family member lymphotoxin-beta is regulated by interleukin (IL)-6 and IL-1beta: transcriptional control mechanisms in oval cells and hepatoma cell lines. Liver Int. 2005;25(3):633–46. doi: 10.1111/j.1478-3231.2005.01080.x. [DOI] [PubMed] [Google Scholar]
- 22.Knight B, Yeoh GC, Husk KL, Ly T, Abraham LJ, Yu C, et al. Impaired preneoplastic changes and liver tumor formation in tumor necrosis factor receptor type 1 knockout mice. J Exp Med. 2000;192(12):1809–18. doi: 10.1084/jem.192.12.1809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Knight B, Lim R, Yeoh GC, Olynyk JK. Interferon-gamma exacerbates liver damage, the hepatic progenitor cell response and fibrosis in a mouse model of chronic liver injury. J Hepatol. 2007;47(6):826–33. doi: 10.1016/j.jhep.2007.06.022. [DOI] [PubMed] [Google Scholar]
- 24.Tirnitz-Parker JE, Viebahn CS, Jakubowski A, Klopcic BR, Olynyk JK, Yeoh GC, et al. Tumor necrosis factor-like weak inducer of apoptosis is a mitogen for liver progenitor cells. Hepatology. 2010;52(1):291–302. doi: 10.1002/hep.23663. [DOI] [PubMed] [Google Scholar]
- 25.Paku S, Schnur J, Nagy P, Thorgeirsson SS. Origin and structural evolution of the early proliferating oval cells in rat liver. Am J Pathol. 2001;158(4):1313–23. doi: 10.1016/S0002-9440(10)64082-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kallis YN, Robson AJ, Fallowfield JA, Thomas HC, Alison MR, Wright NA, et al. Remodelling of extracellular matrix is a requirement for the hepatic progenitor cell response. Gut. 2011;60(4):525–33. doi: 10.1136/gut.2010.224436. [DOI] [PubMed] [Google Scholar]
- 27.Wang S, Song K, Srivastava R, Dong C, Go GW, Li N, et al. Nonalcoholic fatty liver disease induced by noncanonical Wnt and its rescue by Wnt3a. FASEB J. 2015;29(8):3436–45. doi: 10.1096/fj.15-271171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Monga SP. Role and regulation of beta-catenin signaling during physiological liver growth. Gene Expr. 2014;16(2):51–62. doi: 10.3727/105221614X13919976902138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Apte U, Singh S, Zeng G, Cieply B, Virji MA, Wu T, et al. Beta-catenin activation promotes liver regeneration after acetaminophen-induced injury. Am J Pathol. 2009;175(3):1056–65. doi: 10.2353/ajpath.2009.080976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Apte U, Thompson MD, Cui S, Liu B, Cieply B, Monga SP. Wnt/beta-catenin signaling mediates oval cell response in rodents. Hepatology. 2008;47(1):288–95. doi: 10.1002/hep.21973. [DOI] [PubMed] [Google Scholar]
- 31.Hu M, Kurobe M, Jeong YJ, Fuerer C, Ghole S, Nusse R, et al. Wnt/beta-catenin signaling in murine hepatic transit amplifying progenitor cells. Gastroenterology. 2007;133(5):1579–91. doi: 10.1053/j.gastro.2007.08.036. [DOI] [PubMed] [Google Scholar]
- 32.Williams JM, Oh SH, Jorgensen M, Steiger N, Darwiche H, Shupe T, et al. The role of the Wnt family of secreted proteins in rat oval ‘stem’ cell-based liver regeneration: Wnt1 drives differentiation. Am J Pathol. 2010;176(6):2732–42. doi: 10.2353/ajpath.2010.080486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Itoh T, Kamiya Y, Okabe M, Tanaka M, Miyajima A. Inducible expression of Wnt genes during adult hepatic stem/progenitor cell response. FEBS Lett. 2009;583(4):777–81. doi: 10.1016/j.febslet.2009.01.022. [DOI] [PubMed] [Google Scholar]
- 34.Thompson MD, Awuah P, Singh S, Monga SP. Disparate cellular basis of improved liver repair in beta-catenin-overexpressing mice after long-term exposure to 3,5-diethoxycarbonyl-1,4-dihydrocollidine. Am J Pathol. 2010;177(4):1812–22. doi: 10.2353/ajpath.2010.100173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Akhmetshina A, Palumbo K, Dees C, Bergmann C, Venalis P, Zerr P, et al. Activation of canonical Wnt signalling is required for TGF-beta-mediated fibrosis. Nat Commun. 2012;3:735. doi: 10.1038/ncomms1734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Henderson WR, Jr, Chi EY, Ye X, Nguyen C, Tien YT, Zhou B, et al. Inhibition of Wnt/beta-catenin/CREB binding protein (CBP) signaling reverses pulmonary fibrosis. Proc Natl Acad Sci U S A. 2010;107(32):14309–14. doi: 10.1073/pnas.1001520107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Beyer C, Schramm A, Akhmetshina A, Dees C, Kireva T, Gelse K, et al. beta-catenin is a central mediator of pro-fibrotic Wnt signaling in systemic sclerosis. Ann Rheum Dis. 2012;71(5):761–7. doi: 10.1136/annrheumdis-2011-200568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hao S, He W, Li Y, Ding H, Hou Y, Nie J, et al. Targeted inhibition of beta-catenin/CBP signaling ameliorates renal interstitial fibrosis. J Am Soc Nephrol. 2011;22(9):1642–53. doi: 10.1681/ASN.2010101079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yang J, Mowry LE, Nejak-Bowen KN, Okabe H, Diegel CR, Lang RA, et al. Beta-catenin signaling in murine liver zonation and regeneration: a Wnt-Wnt situation! Hepatology. 2014;60(3):964–76. doi: 10.1002/hep.27082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Carpenter AC, Rao S, Wells JM, Campbell K, Lang RA. Generation of mice with a conditional null allele for Wntless. Genesis. 2010;48(9):554–8. doi: 10.1002/dvg.20651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Clausen BE, Burkhardt C, Reith W, Renkawitz R, Forster I. Conditional gene targeting in macrophages and granulocytes using LysMcre mice. Transgenic Res. 1999;8(4):265–77. doi: 10.1023/A:1008942828960. [DOI] [PubMed] [Google Scholar]
- 42.Pradere JP, Kluwe J, De Minicis S, Jiao JJ, Gwak GY, Dapito DH, et al. Hepatic macrophages but not dendritic cells contribute to liver fibrosis by promoting the survival of activated hepatic stellate cells in mice. Hepatology. 2013;58(4):1461–73. doi: 10.1002/hep.26429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Karsdal MA, Manon-Jensen T, Genovese F, Kristensen JH, Nielsen MJ, Sand JM, et al. Novel insights into the function and dynamics of extracellular matrix in liver fibrosis. Am J Physiol Gastrointest Liver Physiol. 2015;308(10):G807–30. doi: 10.1152/ajpgi.00447.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wang Q, Usinger W, Nichols B, Gray J, Xu L, Seeley TW, et al. Cooperative interaction of CTGF and TGF-beta in animal models of fibrotic disease. Fibrogenesis Tissue Repair. 2011;4(1):4. doi: 10.1186/1755-1536-4-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Dolle L, Theise ND, Schmelzer E, Boulter L, Gires O, van Grunsven LA. EpCAM and the biology of hepatic stem/progenitor cells. Am J Physiol Gastrointest Liver Physiol. 2015;308(4):G233–G50. doi: 10.1152/ajpgi.00069.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Okabe M, Tsukahara Y, Tanaka M, Suzuki K, Saito S, Kamiya Y, et al. Potential hepatic stem cells reside in EpCAM+ cells of normal and injured mouse liver. Development. 2009;136(11):1951–60. doi: 10.1242/dev.031369. [DOI] [PubMed] [Google Scholar]
- 47.Wynn TA, Barron L. Macrophages: master regulators of inflammation and fibrosis. Semin Liver Dis. 2010;30(3):245–57. doi: 10.1055/s-0030-1255354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Akhurst B, Croager EJ, Farley-Roche CA, Ong JK, Dumble ML, Knight B, et al. A modified choline-deficient, ethionine-supplemented diet protocol effectively induces oval cells in mouse liver. Hepatology. 2001;34(3):519–22. doi: 10.1053/jhep.2001.26751. [DOI] [PubMed] [Google Scholar]
- 49.Karlmark KR, Zimmermann HW, Roderburg C, Gassler N, Wasmuth HE, Luedde T, et al. The fractalkine receptor CXCR1 protects against liver fibrosis by controlling differentiation and survival of infiltrating hepatic monocytes. Hepatology. 2010;52(5):1769–82. doi: 10.1002/hep.23894. [DOI] [PubMed] [Google Scholar]
- 50.Dal-Secco D, Wang J, Zeng Z, Kolaczkowska E, Wong CH, Petri B, et al. A dynamic spectrum of monocytes arising from the in situ reprogramming of CCR2+ monocytes at a site of sterile injury. J Exp Med. 2015;212(4):447–56. doi: 10.1084/jem.20141539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ramachandran P, Iredale JP. Macrophages: central regulators of hepatic fibrogenesis and fibrosis resolution. J Hepatol. 2012 doi: 10.1016/j.jhep.2011.10.026. [DOI] [PubMed] [Google Scholar]
- 52.Ramachandran P, Pellicoro A, Vernon MA, Boulter L, Aucott RL, Ali A, et al. Differential Ly-6C expression identifies the recruited macrophage phenotype, which orchestrates the regression of murine liver fibrosis. Proc Natl Acad Sci U S A. 2012;109(46):E3186–95. doi: 10.1073/pnas.1119964109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Huch M, Dorrell C, Boj SF, van EJH, Li VS, van dWM, et al. In vitro expansion of single Lgr5+ liver stem cells induced by Wnt-driven regeneration. Nature. 2013;494(7436):247–50. doi: 10.1038/nature11826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Yimlamai D, Christodoulou C, Galli GG, Yanger K, Pepe-Mooney B, Gurung B, et al. Hippo pathway activity influences liver cell fate. Cell. 2014;157(6):1324–38. doi: 10.1016/j.cell.2014.03.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Michalopoulos GK, Barua L, Bowen WC. Transdifferentiation of rat hepatocytes into biliary cells after bile duct ligation and toxic biliary injury. Hepatology. 2005;41(3):535–44. doi: 10.1002/hep.20600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Wang B, Zhao L, Fish M, Logan CY, Nusse R. Self-renewing diploid Axin2(+) cells fuel homeostatic renewal of the liver. Nature. 2015;524(7564):180–5. doi: 10.1038/nature14863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Wang EY, Yeh SH, Tsai TF, Huang HP, Jeng YM, Lin WH, et al. Depletion of beta-catenin from mature hepatocytes of mice promotes expansion of hepatic progenitor cells and tumor development. Proc Natl Acad Sci U S A. 2011;108(45):18384–9. doi: 10.1073/pnas.1116386108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Winkler T, Mahoney EJ, Sinner D, Wylie CC, Dahia CL. Wnt signaling activates Shh signaling in early postnatal intervertebral discs, and re-activates Shh signaling in old discs in the mouse. PLoS One. 2014;9(6) doi: 10.1371/journal.pone.0098444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Michelotti GA, Xie G, Swiderska M, Choi SS, Karaca G, Kruger L, et al. Smoothened is a master regulator of adult liver repair. J Clin Invest. 2013;123(6):2380–94. doi: 10.1172/JCI66904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kordes C, Sawitza I, Haussinger D. Canonical Wnt signaling maintains the quiescent stage of hepatic stellate cells. Biochem Biophys Res Commun. 2008;367(1):116–23. doi: 10.1016/j.bbrc.2007.12.085. [DOI] [PubMed] [Google Scholar]
- 61.Yoshiji H, Kuriyama S, Yoshii J, Ikenaka Y, Noguchi R, Nakatani T, et al. Tissue inhibitor of metalloproteinases-1 attenuates spontaneous liver fibrosis resolution in the transgenic mouse. Hepatology. 2002;36(4 Pt 1):850–60. doi: 10.1053/jhep.2002.35625. [DOI] [PubMed] [Google Scholar]
- 62.Parsons CJ, Bradford BU, Pan CQ, Cheung E, Schauer M, Knorr A, et al. Antifibrotic effects of a tissue inhibitor of metalloproteinase-1 antibody on established liver fibrosis in rats. Hepatology. 2004;40(5):1106–15. doi: 10.1002/hep.20425. [DOI] [PubMed] [Google Scholar]
- 63.Barnes MA, McMullen MR, Roychowdhury S, Madhun NZ, Niese K, Olman MA, et al. Macrophage migration inhibitory factor is required for recruitment of scar-associated macrophages during liver fibrosis. J Leukoc Biol. 2015;97(1):161–9. doi: 10.1189/jlb.3A0614-280R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Pellicoro A, Ramachandran P, Iredale JP, Fallowfield JA. Liver fibrosis and repair: immune regulation of wound healing in a solid organ. Nat Rev Immunol. 2014;14(3):181–94. doi: 10.1038/nri3623. [DOI] [PubMed] [Google Scholar]
- 65.Endo H, Niioka M, Sugioka Y, Itoh J, Kameyama K, Okazaki I, et al. Matrix metalloproteinase-13 promotes recovery from experimental liver cirrhosis in rats. Pathobiology. 2011;78(5):239–52. doi: 10.1159/000328841. [DOI] [PubMed] [Google Scholar]
- 66.Kim WH, Matsumoto K, Bessho K, Nakamura T. Growth inhibition and apoptosis in liver myofibroblasts promoted by hepatocyte growth factor leads to resolution from liver cirrhosis. Am J Pathol. 2005;166(4):1017–28. doi: 10.1016/S0002-9440(10)62323-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Marquardt JU, Seo D, Gomez-Quiroz LE, Uchida K, Gillen MC, Kitade M, et al. Loss of c-Met accelerates development of liver fibrosis in response to CCl(4) exposure through deregulation of multiple molecular pathways. Biochim Biophys Acta. 2012;1822(6):942–51. doi: 10.1016/j.bbadis.2012.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Suzumura K, Hirano T, Son G, Iimuro Y, Mizukami H, Ozawa K, et al. Adeno-associated virus vector-mediated production of hepatocyte growth factor attenuates liver fibrosis in mice. Hepatol Int. 2008;2(1):80–8. doi: 10.1007/s12072-007-9042-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Ishikawa T, Factor VM, Marquardt JU, Raggi C, Seo D, Kitade M, et al. Hepatocyte growth factor/c-met signaling is required for stem-cell-mediated liver regeneration in mice. Hepatology. 2012;55(4):1215–26. doi: 10.1002/hep.24796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Zulehner G, Mikula M, Schneller D, van Zijl F, Huber H, Sieghart W, et al. Nuclear beta-catenin induces an early liver progenitor phenotype in hepatocellular carcinoma and promotes tumor recurrence. Am J Pathol. 2010;176(1):472–81. doi: 10.2353/ajpath.2010.090300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Wallace MC, Hamesch K, Lunova M, Kim Y, Weiskirchen R, Strnad P, et al. Standard operating procedures in experimental liver research: thioacetamide model in mice and rats. Lab Anim. 2015;49(1 Suppl):21–9. doi: 10.1177/0023677215573040. [DOI] [PubMed] [Google Scholar]
- 72.Fagan KJ, Pretorius CJ, Horsfall LU, Irvine KM, Wilgen U, Choi K, et al. ELF score >/=9.8 indicates advanced hepatic fibrosis and is influenced by age, steatosis and histological activity. Liver Int. 2014. doi:10.1111/liv.12760. [DOI] [PubMed]
- 73.Gadd VL, Melino M, Roy S, Horsfall L, O’Rourke P, Williams MR, et al. Portal, but not lobular, macrophages express matrix metalloproteinase-9: association with the ductular reaction and fibrosis in chronic hepatitis C. Liver Int. 2013;33(4):569–79. doi: 10.1111/liv.12050. [DOI] [PubMed] [Google Scholar]