

Abstract
We review select aspects of the biology of matrix metalloproteinases (MMPs) with a focus on the modulation of inflammatory responses by MMP-2. MMP-2 is a zinc- and calcium-dependent endoprotease with substrates including extracellular matrix proteins, vasoactive peptides and chemokines. Humans and mice with MMP-2 deficiency exhibit a predominantly inflammatory phenotype. Recent research shows that MMP-2 deficient mice display elevated activity of a secreted phospholipase A2 in the heart. Additionally, MMP-2 deficient mice exhibit abnormally high prostaglandin E2 levels in various organs (i.e., the heart, brain and liver), signs of inflammation and exacerbated lipopolysaccharide-induced fever. We briefly review the biology of sPLA2 enzymes to propose the existence of a heart-centric MMP-2/sPLA2 axis of systemic inflammation. Moreover, we postulate that PLA2 activation is induced by chemokines, whose ability to signal inflammation is regulated in a tissue-specific fashion by MMPs. Thus, genetic and pharmacologically induced MMP-deficiencies can be expected to perturb PLA2-mediated inflammatory mechanisms.
Keywords: MMPs, Inflammation, Chemokines, Secreted phospholipase A2
Relevant aspects of MMP-2 biology
The 1960s saw the demonstration that mammalian systems make collagenases,1,2 a discovery followed by the cloning of a superfamily of so-called extracellular matrix metalloproteinases (MMPs). Today, over 20 different zinc and calcium-dependent MMPs are known, each capable of degrading numerous substrates including (but not limited to) extracellular matrix components, and each involved in numerous biological processes and disease conditions.3
The focal point of this review is MMP-2, a ubiquitous and constitutive 72 kDa type IV collagenase. MMP-2 binds collagen through the fibronectin type II repeat in its catalytic domain and cleaves numerous extracellular matrix components including native collagens (types IV, V VII and X), denatured collagens, elastin and heparan-sulfate proteoglycans which are extracellular matrix receptors for signaling molecules. MMP-2 mediated cleavage of these extracellular components is important for the integrity of the matrix as well as for signaling events that impact cell behaviour. An illustrative example is MMP-2-mediated cleavage of decorin. Decorin is a multifunctional proteoglycan whose core protein has a serine covalently linked to a dermatan/chondroitin sulphate glycosaminoglycan chain. The integrity of extracellular decorin is important for normal collagen fibrillogenesis. Decorin is cleaved by MMP-2 which impairs decorin’s ability to sequester transforming growth factor–β, a master regulator of fibrosis signaling through the SMADs.4,5
A striking and most interesting feature of MMPs (and MMP-2 in particular) is its high promiscuity in terms of substrates which include non-extracellular matrix proteins and peptides.6,7 A relevant example with potential pathophysiological significance is the discovery that MMP-2 binds through its hemopexin-like domain and cleaves the CC-motif chemokine, monocyte chemoattractant protein-3 (MCP-3, ~9 kDa, encoded by Ccl7). Cleavage of MCP-3 at a Gly4/Ile5 bond converts it into a general CC-chemokine receptor antagonist which dampens inflammation.8 Interestingly, MCP-3 is also cleaved at the Gly4/Ile5 bond by MMP-1, 3, 13 and 14. MCP-1 is cleaved at its Ala4/Leu5 bond by MMP-1 and MMP-3, while MCP-2 is cleaved at its Ser4/Val5 bond by MMP-3, and MCP-4 is cleaved at its Ala4/Leu5 bond by MMP-1 and MMP-3.9 In addition to MCPs, MMP-2 is also known to cleave other substrates relating to inflammation. For example, MMP-2 cleaves CX3CL1, a pro-inflammatory protein, and possibly allowing for protective effects against rheumatoid arthritis through the suppression of macrophage influx. Cleavage of Ym1, S100A8 or S100A9 by MMP-2 or MMP-9 reduces the chemotactic activity of these proteins..7 Thus, MMPs and substrates such as chemokines likely have interrelated functions in the context of inflammation and innate immunity.9,10 In the context of cardiovascular regulation, the first MMP-2 substrates to be identified were vasoactive peptides, i.e., big endothelin (ET)-1,11 calcitonin gene-related peptide (CGRP) and adrenomedullin (ADM).12,13 A striking similarity between these peptides is that MMP-2 attacks them at Gly/Leu bonds; a peptide bond that is homologous to amino acid sequences targeted by MMP-2 in interstitial collagen (Gly-Leu or Gly-Ile), laminin-5 (Ala-Leu) and MCP-3 (Gly-Leu).
Another interesting observation is that the proteolytic cleavage of any of these peptides would be expected to impact vascular contractility. Possible mechanisms include promoting vasoconstriction or reducing vasodilation, although the promotion of vasodilation is a potential outcome in the case of endothelin-1.11–14 When experimental animals with spontaneous or agonist-induced hypertension are administered a MMP inhibitor such as doxycycline, the severity of their hypertension is consistently attenuated.15–19 MMP-2 activity may also impact vascular tone in normal physiological processes such as aging20–22 and pregnancy14,23,24 as well as in hypertensive conditions such as pregnancy-induced hypertension.14,20–22,25–27 Whether MMP-2 acts solely through the cleavage and regulation of vasoactive substrates or other mechanisms such as inflammation dampening (eg, the cleavage of pro-inflammatory chemokines that exert indirect vascular tone alterations) warrants further research.
Although there is a paucity of mechanisms, there is an emerging link between lipid metabolic gene expression and activity of MMPs – a topic we reviewed previously.3 For instance, MMP-2 may mediate a cardioprotective mechanism involving inhibition of the sterol regulatory element binding protein-2 (SREBP-2) pathway in the heart with absence of MMP-2 predisposing to hypertensive heart disease and resistance to statins.28 Mice lacking MMP-2 show resistance to high fat-induced obesity.29 Different metabolic phenotypes have been exposed in mice lacking MMP-2, MMP-9 or tissue inhibitors of MMPs.29–38 Lipoparticle receptor as well as apoplipoprotein cleavage has been demonstrated for MMP-2 and MMP-14, and could lead to lipoparticle dysregulation.39–46 We recently reported that MMP-2 can cleave proprotein convertase subtilisin/kexin type 9 (PCSK9, Pcsk9, ~70 kDa) in the pro-peptide and C-terminal domains.47 An important function of PCSK9 is to bind the receptor for low density lipoprotein (LDLR) and reroute it from the recycling to the lysosomal degradation pathway. Consequently, PCSK9 binding to LDLR reduces LDLR bioavailability and impairs hepatic uptake of plasma LDL-cholesterol. Mutations in PCSK9 can either cause or protect form hypercholesterolemia.48–50 The observation that MMP-2 binds and cleaves PCSK9 suggests a role for MMP-2 as an inhibitor of PCSK9-induced LDLR degradation and as a modulator of the metabolism of plasma LDL cholesterol and atherosclerosis development.47 Overall, there are puzzling links between MMPs (including MMP-2) and metabolism, making metalloproteinases attractive targets for changing the course of cardiometabolic diseases, whose etiology is notoriously complicated by co-morbidities such as obesity, atherosclerosis, diabetes and metabolic syndrome.3 The challenge lies in determining specific mechanisms of MMPs and establishing their context-dependent contribution to systemic metabolism.
The number of MMP-2 substrates has kept increasing steadily and the trend might continue over time. A recent proteomic approach revealed a number of new potential MMP-2 targets, including galectin-1 and insulin growth factor binding protein-4 among others involved in angiogenesis.6 MMP-2-mediated cleavage of plasminogen may also yield the angiogenesis inhibitor, angiostatin,51,52 The ever increasing number of substrates likely modulated by MMPs justifies posing the question of not ‘what MMPs do?’ but rather ‘what do they not do?!6,7,51
Relevant aspects of MMP-2 deficiency in humans and mice
Although rare, MMP2 deficiency is a pan-ethnic disorder, first reported in Saudi Arabian, Indian and Turkish families – all presenting multicentric osteolysis with nodulosis and arthropathy (a condition also known as MONA, Online Mendelian Inheritance in Man no. 259600).53 Many arthritic syndromes (eg, Torg, Winchester) have since been reported that co-develop with MMP-2 deficiency.53–55 This autosomal recessive condition is characterized by ‘vanishing bone’ syndrome, carpal and tarsal osteolysis and interphalangeal joint erosions, facial dysmorphia, fibrocollagenous nodules and congenic heart defects. Lack of MMP-2 in humans results from inactivating mutations. Two family-specific homoallelic MMP2 mutations: R101H and Y244X, result in deletion of the substrate-binding and catalytic sites, and the fibronectin type II-like and hemopexin/TIMP2 binding domains of MMP-2. Not surprisingly, the deletion of the terminal hemopexin domain of MMP-2 (comprising 83 C-terminal amino acids) also causes MONA and cardiac dysfunction. Human MMP-2 deficiency has no known mechanism, and remains without cure.53
Arguably, work with models of MMP deficiency can tell us much about the physiological and pathological functions of MMPs. Work with MMP-2 deficient mice has revealed a pronounced inflammatory phenotype while extracellular matrix accumulation due to a proteolytic defect seems to be limited.56–58 Mmp2−/− mice exhibit craniofacial abnormalities,54,59 are relatively small at birth and have delayed growth vs. age-matched wild type mice.29,36 These features are reminiscent of MMP-2 deficient humans that also exhibit dwarfism.53–55 Mmp2−/− mice also show propensity to hypertensive heart disease with cardiac inflammation and hugely elevated activity of a secretory phospholipase A2 (sPLA2) in myocardium,28,60 a hydrolase that targets the carbon 2 position of membrane glycerophospholipids to release fatty acids which are precursors of eicosanoids and other lipid mediators, as well as modulators of lipid metabolic gene transcription.61,62
Furthermore, absence of MMP-2 in mice impairs resolution of lung inflammation induced by allergens. This lack in MMP-2 causes the accumulation of eosinophils and TH2 cytokines in the lung, affecting lung fibroblasts and smooth muscle cells. Without the appropriate chemokines, the lung is unable to form a chemokine gradient that allows for luminal clearance of inflammatory cells.57 As a result, Mmp2−/− mice have a reduced ability to clear recruited immune cells from the lung, have robust asthmatic reactions and asphyxiate more easily than wild type mice when challenged with allergens.56–58 Therefore, MMP-2 deficiency in both humans and mice is pro-inflammatory in multiple organs as well as potentially having a metabolic component.
Why does MMP-2 deficiency cause inflammation?
We develop next a hypothetical answer to this question departing from the ability of MMP-2 to cleave extracellular matrix components and the chemokine, MCP-3, as well as being an endogenous inhibitor of cardiac sPLA2. We end up proposing that MMP-2, MCP-3 and sPLA2 activity are interrelated in a potentially novel pathway with ramifications for modulation of systemic inflammation and metabolism.
MMP-2 deficiency may cause an extracellular matrix breakdown defect. Paradoxically, studies of humans and mice with MMP-2 deficiency show a common predisposition to inflammation and arthritis;53–59,63 MMP-2 deficiency also predisposes to cardiac hypertrophy and fibrosis in response to angiotensin II but may protect against infarction.64–66 A study examined the cardiac expression of 56 metabolic and inflammatory genes in MMP-2 deficient mice.28 Results showed high levels of SREBP-2 and 3-hydroxy-3-methylglutarylcoenzyme A reductase (HMGCR), and elevated levels of pro-inflammatory genes such as: Ccl5, Ccl2 and Ccl6. MMP-2 was found to negatively regulate a mechanism involving SREBP-2 and HMGCR (the rate limiting enzyme in the synthesis of mevalonate) – factors predisposing to cardiac hypertrophy. Thus, MMP-2 expression during the development of agonist-induced hypertension may be cardioprotective, by preventing the upregulation of the SREBP-2/HMGCR pathway in the heart. Establishing how and when MMP-2 is protective vs. deleterious for cardiac function warrants further investigation.2,28,64–68
MMP-2 deficiency also impacts adipose tissue metabolism: Studies in mice indicate the formation of a fibrous cap around adipose tissue that might cause adipocyte hypotrophy and delayed growth.33,67 The delayed growth in mice may be present in humans, where the phenotype of MMP-2 deficiency is not ‘fibrosis’. Humans and mice with MMP-2 deficiency do not exhibit a generalized defect in extracellular matrix breakdown, but primarily crippling arthritis and dwarfism.53–55,59
A very attractive hypothesis is that MMP-2 primarily acts by cleaving and regulating chemokines, a characteristic shared by multiple MMPs (eg, MMP-2 and MMP-9).8,9,68–70 Although in vivo data are limited, chemokine cleavage and conversion from agonist into general chemokine receptor antagonist (as originally proposed for MCP-3) is an elegant mechanism whereby MMP-2 deficiency could exacerbate inflammation.8 Supporting studies include one where synthetic peptides with amino acid sequences corresponding to fragments of MCP-1, MCP-3 or MCP-4, equivalent to those that would be generated by MMP-mediated proteolysis, reduced carrageenan-induced paw swelling in rat.9 Similarly, Mmp2−/− mice infected with coxsackievirus develop mortal endocarditis within 2 weeks of infection.70 This exacerbated endocarditis is associated with elevated cardiac infiltration of neutrophils, macrophages and T cells (such as CD4+ and CD8+) and significantly reduced by MCP-3–neutralizing antibody treatment.70
Recently, we advanced the hypothesis that a heart-centric MMP-2-mediated mechanism may modulate blood pressure homeostasis, inflammatory responses and the severity of fever.60 We found hugely increased levels of sPLA2 in myocardium of Mmp2−/− mice. Tissue distribution analysis showed sPLA2 activity being much higher in the heart than in the liver, adipose tissue or kidney of Mmp2−/− mice. The enzyme is readily secreted from cardiac specimens ex vivo and has similar size, requirement of calicium and apparent MichaelisMenten constant for diheptanoyl thio-PC with plasma sPLA2, suggesting that the heart secretes sPLA2, which next acts on distal tissues (such as the liver) as well as on the heart to impact their inflammatory and metabolic state.
Linking MMP-2 with the biology of phospholipases A2
To illustrate the emerging connection between MMP-2 and cardiac sPLA2 activity and its potential pathophysiological significance, we review briefly some aspects concerning the various groups and functions ascribed to the PLA2 family. Phospholipases are membrane glycerophospholipid hydrolases which, depending on their site of action, are subdivided into type A1, A2, B, C or D. Type A1 releases the fatty acid esterified at carbon-1 (C-1) leaving behind a lysophospholipid. Similarly, type A2 releases the fatty acid esterified at C-2 leaving behind a lysophospholipid, while type B can release the fatty acid esterified at either C-1 or C-2. Type C phospholipase hydrolyses the bond between C-3 and the phosphate group to release diacylglycerol and phosphorylated head group. Phospholipase D cleaves the bond after the phosphate group releasing diacylglycerol phosphate and head group.
Current data is consistent with MMP-2 deficiency upregulating a type A2 phospholipase, an enzyme type whose existence was demonstrated back in the 19th century when it was found as a major component of snake venoms.61,71 Secreted PLA2 family has at least 11 isoforms in humans and mice:61,71,72 PLA2G1B, PLA2G2(A, C, D, E, F), PLA2G3, PLA2G5, PLA2G10, and PLA2G12(A, B). Members of the sPLA2 family have a molecular mass of 16–18 kDa, require calcium, have 6–8 disulphide bridges, and a histidine-aspartate dyad that catalyzes the hydrolysis and the release of fatty acids from the C-2 position of membrane glycerophospholipids. As a reflection of the multiple isoforms, sPLA2s target a spectrum of membrane phospholipids, with each individual phospholipase having distinct substrate selectivity and biological functions.72 This notion is best supported by studies72 with gene knock-out models and mass-spectrometry-based lipidomics to functionally characterize individual members of the sPLA2 family. The data gathered for PLA2G1B, PLA2G2(A, D, E), PLA2G3, PLA2G5, PLA2G10 now enables a functional subdivision of these isoforms as “digestive”, “inflammatory or bactericidal”, “resolving”, “metabolic”, “reproductive or anaphylactic”, “TH2-prone or metabolic”, and “asthmatic, reproductive or gastrointestinal”.72
The identity and amino acid sequence of the PLA2 upregulated in Mmp2−/− mice remains elusive.60 Enzyme inhibition assays with indoxam suggest that cardiac sPLA2 in Mmp2−/− mice is a mixture of sPLA2s (excluding PLA2G2A whose gene is disrupted in C57BL mice) or a completely novel enzyme with very interesting characteristics. For instance, the specific activity is elevated in excess of 102 fold in the heart regardless of sex and in a wide variety of dietary conditions vs. wild type (Mmp2+/+ C57BL/6) mice. Haplo-insufficiency (Mmp2−/+) lessens the specific activity of cardiac sPLA2, while pharmacological MMP inhibition increases both plasma and cardiac sPLA2 activity in wild type (Mmp2+/+) but not in Mmp2−/− mice. However, the activity is not elevated in many organs such as the liver or adipose tissue even though the Mmp2−/− mice are a whole body gene knock-out model. Therefore, a cardiac-specific mechanism (perhaps, an agonist), which is under MMP-2 control, must be responsible for the cardiac-specific upregulation of sPLA2 activity in MMP-2 deficient mice.
The emerging picture is one where, given the same pathophysiological context, sPLA2s each have unique induction profiles and functions, despite catalyzing the same type of biochemical reaction. What then is the biological function of cardiac sPLA2 expressed in MMP-2 deficiency? Evidently, cardiac sPLA2 is both pro-inflammatory and metabolic in the heart.60 MMP-2 deficiency results in cardiac overexpression of inflammatory marker genes, eg, Tnfa and Il1b, which are downregulated by the pan-sPLA2 inhibitor, varespladib. Similarly, MMP-2 deficiency results in cardiac dysregulation of lipid metabolic genes, eg, elevated Srebf2 (encodes sterol regulatory binding protein-2) and target genes which are enzymes of the mevalonate pathway. The expression of these genes is normalized by varespladib. Moreover, cardiac sPLA2 pro-inflammatory effects appear to be systemic. MMP-2-deficient mice exhibit increased levels of prostaglandin E2 (PGE2) at baseline heart, brain and liver. Keeping in mind that cyclooxygenase and PGE2 synthase are important in the regulation of PGE2, 8-isoprostanes (prostaglandins not produced directly by cyclooxygenase) were also measured, once again showing elevated levels in Mmp2−/− mice compared to WT mice.60 Further, MMP-2 deficient mice develop exacerbated fever in response to low-dose bacterial lipopolysaccharide. This fever response is completely blunted by systemic administration of varespladib. In fact, the effect of cardiac sPLA2 on fever appears to be so dominant that only MMP-2 deficient mice, but not wild type mice, show blunted fever in response to varespladib. Thus, it is not surprising that MMP-2 deficient mice rely on prostanoids for blood pressure homeostasis and develop acute hypertension 4 hours after treatment with either varespladib or indomethacin. A corollary of these observations is that the inflammatory state of the heart impacts the inflammatory and metabolic state at systemic levels as well as blood pressure homeostasis, at least in part, through the MMP-2/sPLA2 axis. These observations bear potential significance for the understanding and clinical management of human conditions associated with reduced MMP-2 expression; particularly, human MMP-2 deficiency.60
The MMP-2/cardiac sPLA2 regulatory system is not an artefact of life-long MMP-2 deficiency. Cardiac sPLA2 activity is readily activated in mice treated with the pharmacological broad-spectrum MMP inhibitor doxycycline – the only MMP inhibitor with US Food and Drug Administration (FDA) approval. In these mice treated with doxycycline, plasma sPLA2 is significantly increased in 2–3 days and cardiac sPLA2 is hugely upregulated after 2 weeks. These observations also suggest mechanisms for cardiac specific effects of MMP-2 inhibitors, such as doxycycline, which currently has FAD-approval and is widely used for its many actions including as an antibiotic and a non-antibiotic/disease modifier drug with the capacity to inhibit the expression and activity of MMPs.73–77
Unanswered research questions
Beyond surmounting the challenge of identifying cardiac sPLA2, there are research opportunities on the biology of MMP-2 and the phenotype induced by its deficiency: What agonist(s) induce cardiac sPLA2 activity? How does MMP-2 inhibit such agonist(s)? What is the mechanism elicited by the agonist(s)? Why is sPLA2 activity so highly elevated in the MMP-2 deficient heart? Are cardiac and plasma sPLA2 the same enzyme (or enzyme mixtures)? How does the heart impact the inflammatory status of distal tissues and organs? If the heart impacts the inflammatory status of distal tissues through the secretion of sPLA2 from myocardium, the further question of how does cardiac sPLA2 affect inflammation on target sites remains to be answered. Many pathologies exhibit dysregulation of either MMP-2 or sPLA2, eg: arteriosclerosis, diabetes, obesity, arthritis, asthma, anaphylaxis and pain. It is tempting to speculate that the interaction between MMP-2 and sPLA2, which is released from myocardium, may contribute to the pathophysiology of these conditions. What is the actual contribution of the MMP-2/sPLA2 axis in these pathologies relative to other established mechanisms? Does the emergent link between MMP-2 and cardiac sPLA2 imply that the heart plays a more central role in systemic physiology and disease than previously suspected?
Explaining inflammation in MMP-2 deficiency
Previous research has shown that monocyte chemotaxis is significantly influenced by MCP-1.78–80 It has been shown that MCP-1 acts, at least in part, by binding to CC-chemokine receptor 2 to induce the intracellular translocation and activation of cytosolic calcium-dependent PLA2 (cPLA2) and calcium-independent PLA2 (iPLA2) isoforms.79 Interestingly, MCP-1 is cleaved and regulated by MMP-1 and MMP-3.9 Therefore, we hypothesize that MMPs, such as MMP-1 and MMP-3, may be important in the process of monocyte chemotaxis by cleaving MCP-1.79,80
A similar mechanism may apply to other MMPs and MCPs. MMP-2 cleaves MCP-3 and it is conceivable that MCP-3 shares with MCP-1 the ability to induce PLA2 activity in target cells (be it calcium-dependent, calcium-independent or secreted PLA2 activity). 9 If MCP-3 triggered the activity of secreted PLA2 in target cells, then cardiac MCP-3 could be an agonist of cardiac sPLA2 maturation and secretion in MMP-2 deficient hearts with MMP-2 acting as an inhibitor through MMP-2-mediated proteolysis of MCP-3.
We postulate (Figure, Table) that, since various MCP chemokines are susceptible to MMP-mediated proteolysis,9 tissue-specific MMP/MCP/PLA2 mechanisms might exist, whereby MCP triggers PLA2 activity under the control of MMP-mediated proteolysis of MCP. The existence of such mechanisms is supported by reports of MCP-1 actions in monocytes and regulatory loops, where cytokines produced downstream MCPs or PLA2s modify the expression of MCPs, PLA2s and MMPs, and PLA2 activity impacts MMP expression.81–84 Focusing on MCP-1, cytokines like TNF-α and IL-1β are major regulators of MCP-1 production in many systems, at least in part, via the nuclear factor-κB pathway.85,86 Similarly, cPLA2 activity via the prostanoid pathway is known to regulate MMP-9 production in macrophages, while sPLA2 activity induces MMP-2 and MMP-9 thus facilitating fibroblast proliferation and chondrocyte development, respectively.81–84 Thus, MMPs, MCPs and PLA2s depict a signaling network with the potential to affect inflammatory and metabolic responses through local and distal actions.
Figure.
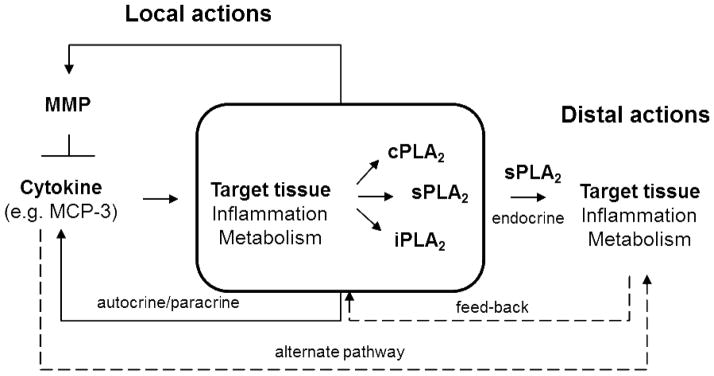
The postulated mechanism of PLA2 regulation by MMPs via cytokines gives rise to many plausible PLA2 mediated signaling events that can impact target tissues at short and long distances. Alternate signaling pathways and feed–back regulatory loops (dashed arrows) are likely part of the complex regulation of target tissues by MMPs.
Table.
Possible MMP/Cytokine/PLA2 axis subtypes
| MMP | Cytokine substrate | Target Cell | PLA2 | Function | Refs. |
|---|---|---|---|---|---|
| MMP-1/3 | MCP-1 | Blood monocytes | iPLA2β cPLA2α |
Chemotaxis | 80,94 |
| MMP-1/3 | MCP-1 | Endothelial cells Macrophages | PLA2G5, Lp-PLA2 | Proatherogenic modification of LDL, Monocyte recruitment into arterial wall (atherosclerosis) | 95–97 |
| MMP-2 | MCP-3 | Cardiac cells | Cardiac sPLA2 | Blood pressure homeostasis, cardiac PGE2 production, cardiac lipid metabolism, fever | 60 |
| MMP-9/12 | IL-8 | Synovial fibroblast-like cells | cPLA2 | Rheumatoid arthritis (inflammation, joint destruction) | 98–102 |
Though this MMP/MCP/PLA2 axis is plausible, alternate mechanisms may be at play. Cytokines may act on distal organs directly to cause effects unrelated to the release of PLA2. Several MCPs can be cleaved by more than one subtype of MMP; for instance, MCP-3 cleavage can be brought about by MMP-1, MMP-3, MMP-13 and MMP-14 acting on their own or in concert, not just MMP-2.9 Thus, in principle, absence of MMP-2 may be compensated by the concurrent activity of other MMPs. However, given the overt inflammatory phenotype of MMP-2 deficient mice, one can only speculate that compensation is not a major mechanism at play in MMP-2 deficiency.
The MMP-2/cardiac sPLA2 system as a new endocrine function of the heart
In the 17th century, William Harvey established the heart as a pump – a concept now familiar to all. However, through further research during the century, new exciting discoveries were made pointing to the heart as a gland. The mammalian heart was shown to be able to both store and release norepinephrine when stimulated, causing sympathetic peripheral effects.87 This concept of the heart possibly being a neuroendocrine organ was proven when in 1983, atrial natriuretic factor (ANF) or atrial natriuretic peptide (ANP) was first isolated and sequenced in rat. Similar to other endocrine organs, the heart was also found to contain a high number of developed endoplasmic reticulum, Golgi complex and storage granules.88 This complex of systems allows the processing of peptides and then eventual release through the atrial-specific granules found in atrial cardiomyocytes. ANF and brain natriuretic peptide (BNP) are part of the family of cardiac natriuretic peptides (cNPs). These peptides are found primarily in the heart and when secreted, cause systemic effects such as hypotension, natriuresis and diuresis. On the other hand, peptides such as ADM and ET-1 are also found in the heart and have systemic effects, but are not truly endocrine products of the heart as they are found mainly in blood vessels rather than cardiomyocytes.88
The proposed MMP-2/sPLA2 axis mechanism points again to the heart as an endocrine organ. We propose that this function is normally masked by the omnipresence of MMP-2 in the myocardium but is readily exposed when MMP-2 deficiency is induced by either gene deletion or pharmacological blockade.60 Cardiac sPLA2 activity in mice deficient in MMP-2 is elevated by orders of magnitude. In addition, there is no difference in sPLA2 activity between WT and Mmp2−/− mice in liver, kidney and skeletal muscle. Thus, in addition to MMP-2, there must be heart-specific determinants of sPLA2 maturation and secretion from myocardium, whose discovery warrants further research. Similar to other heart-centric endocrine secretions where a peptide in the heart is activated by a protease and then secreted from storage granules to act on distal tissues and the heart itself, cardiac sPLA2 is negatively regulated by MMP-2 and secreted from the heart through the conventional endoplasmic reticulum-Golgi pathway to affect fever, blood pressure homeostasis and cardiac inflammatory/lipid metabolic gene expression.60 Notably, these functions of cardiac sPLA2 are different from those exerted by natriuretic peptides and other peptidic cardiac secretions.
MMP-2 overexpression as well as MMP-2 deficiency may cause inflammation
In principle, three scenarios can be distinguished: 1) Normal physiology, 2) MMP-2 deficiency and 3) MMP-2 overexpression with scenarios 2) and 3) contributing to inflammation.
In MMP-2 deficiency the likely cause of the inflammation is an effective excess of cytokines that would otherwise be kept at low levels through MMP-2-mediated proteolysis. This excess is a likely trigger of cardiac sPLA2 release, which may promote systemic inflammation (Figure).
In MMP-2 overexpression the cause of the inflammation may be excessive or disproportionate cleavage of extracellular matrix and, perhaps, other cellular components (such as membrane receptors and growth factors) by MMP-2.3,7 Many conditions including arthritis, obesity, diabetes, tissue injury, cardiomyopathies, and hypertension have been associated with an upregulation of MMP-2 as well as inflammation.3,7,10 However, there is a paucity of reports on the exact mechanisms by which MMP-2 promotes the inflammation in vivo. One suggestive example is, however, the setting of a vascular aneurysm where the overexpression of MMP-2 is deleterious.63,64 Purportedly, the pathological overexpression of MMP-2 creates an excess of collagen relative to elastin thus also increasing stiffness, decreasing distensibility in the vasculature and increasing the risk of aneurysm rupture.89,90 Additionally, a self-perpetuating loop may be created as smooth muscle cells release proteases that further break down the vascular intima. Chemoattractants attach on to the degraded intima and stimulate the migration of inflammatory and smooth muscle cells ultimately exacerbating aneurysm development.89
In situations of MMP-2 overexpression, MMP-2 pharmacological inhibition may protect from inflammation if MMP-2 activity is restored to normal physiological levels. However, if MMP-2 activity were decreased beyond physiological levels, inflammatory signals mediated by chemokines and cardiac sPLA2 (discussed earlier in the review, Figure) may dominate the phenotype induced by MMP-2 inhibitor treatment. This could explain part of the contradictions between reports pertaining to the use of pharmacological MMP inhibitors, such as doxycycline, for therapeutic purposes and emphasizes the need for titrating as opposed to completely inhibiting MMP-2 activity when treating inflammatory disease.60,91–93
Conclusion
Further research should refine the view of the heart as an endocrine organ that modulates systemic inflammatory responses through the secretion of sPLA2, a process that is under the control of MMP-2. The resultant knowledge should help to understand the mechanisms of MMP inhibitory drugs, such as doxycycline, as well as enhance the clinical management of human MMP gene deficiencies, which are rare but very debilitating and ineffectively treated conditions.53
Acknowledgments
This work was supported by summer studentships of the Natural Sciences and Engineering Council of Canada (NSERC) and Alberta Innovates Health Solutions (to DL) and operating grants from NSERC and the Canadian Institutes of Health Research (to CFP).
Contributor Information
Carlos Fernandez-Patron, Department of Biochemistry, Cardiovascular Research Group and Mazankowski Alberta Heart Institute, Faculty of Medicine and Dentistry, University of Alberta, Edmonton, Alberta, Canada.
Dickson Leung, Department of Biochemistry, Faculty of Medicine and Dentistry, University of Alberta, Edmonton, Alberta, Canada.
References
- 1.Gross J, Nagai Y. Specific degradation of the collagen molecule by tadpole collagenolytic enzyme. Proc Natl Acad Sci U S A. 1965 Oct;54(4):1197–1204. doi: 10.1073/pnas.54.4.1197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Schulz R. Intracellular targets of matrix metalloproteinase-2 in cardiac disease: rationale and therapeutic approaches. Annu Rev Pharmacol Toxicol. 2007;47:211–242. doi: 10.1146/annurev.pharmtox.47.120505.105230. [DOI] [PubMed] [Google Scholar]
- 3.Berry E, Bosonea AM, Wang X, Fernandez-Patron C. Insights into the activity, differential expression, mutual regulation, and functions of matrix metalloproteinases and a disintegrin and metalloproteinases in hypertension and cardiac disease. J Vasc Res. 2013;50(1):52–68. doi: 10.1159/000345240. [DOI] [PubMed] [Google Scholar]
- 4.Derynck R, Zhang YE. Smad-dependent and Smad-independent pathways in TGF-beta family signalling. Nature. 2003 Oct 9;425(6958):577–584. doi: 10.1038/nature02006. [DOI] [PubMed] [Google Scholar]
- 5.Imai K, Hiramatsu A, Fukushima D, Pierschbacher MD, Okada Y. Degradation of decorin by matrix metalloproteinases: identification of the cleavage sites, kinetic analyses and transforming growth factor-beta1 release. Biochem J. 1997 Mar 15;322(Pt 3):809–814. doi: 10.1042/bj3220809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Prudova A, auf dem Keller U, Butler GS, Overall CM. Multiplex N-terminome analysis of MMP-2 and MMP-9 substrate degradomes by iTRAQ-TAILS quantitative proteomics. Mol Cell Proteomics. 2010 May;9(5):894–911. doi: 10.1074/mcp.M000050-MCP201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rodriguez D, Morrison CJ, Overall CM. Matrix metalloproteinases: what do they not do? New substrates and biological roles identified by murine models and proteomics. Biochim Biophys Acta. 2010 Jan;1803(1):39–54. doi: 10.1016/j.bbamcr.2009.09.015. [DOI] [PubMed] [Google Scholar]
- 8.McQuibban GA, Gong JH, Tam EM, McCulloch CA, Clark-Lewis I, Overall CM. Inflammation dampened by gelatinase A cleavage of monocyte chemoattractant protein-3. Science. 2000 Aug 18;289(5482):1202–1206. doi: 10.1126/science.289.5482.1202. [DOI] [PubMed] [Google Scholar]
- 9.McQuibban GA, Gong JH, Wong JP, Wallace JL, Clark-Lewis I, Overall CM. Matrix metalloproteinase processing of monocyte chemoattractant proteins generates CC chemokine receptor antagonists with anti-inflammatory properties in vivo. Blood. 2002 Aug 15;100(4):1160–1167. [PubMed] [Google Scholar]
- 10.Morrison CJ, Butler GS, Rodriguez D, Overall CM. Matrix metalloproteinase proteomics: substrates, targets, and therapy. Curr Opin Cell Biol. 2009 Oct;21(5):645–653. doi: 10.1016/j.ceb.2009.06.006. [DOI] [PubMed] [Google Scholar]
- 11.Fernandez-Patron C, Radomski MW, Davidge ST. Vascular matrix metalloproteinase-2 cleaves big endothelin-1 yielding a novel vasoconstrictor. Circ Res. 1999 Nov 12;85(10):906–911. doi: 10.1161/01.res.85.10.906. [DOI] [PubMed] [Google Scholar]
- 12.Fernandez-Patron C, Stewart KG, Zhang Y, Koivunen E, Radomski MW, Davidge ST. Vascular matrix metalloproteinase-2-dependent cleavage of calcitonin gene-related peptide promotes vasoconstriction. Circ Res. 2000 Oct 13;87(8):670–676. doi: 10.1161/01.res.87.8.670. [DOI] [PubMed] [Google Scholar]
- 13.Martinez A, Oh HR, Unsworth EJ, et al. Matrix metalloproteinase-2 cleavage of adrenomedullin produces a vasoconstrictor out of a vasodilator. Biochem J. 2004 Nov 1;383(Pt. 3):413–418. doi: 10.1042/BJ20040920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jeyabalan A, Novak J, Danielson LA, Kerchner LJ, Opett SL, Conrad KP. Essential role for vascular gelatinase activity in relaxin-induced renal vasodilation, hyperfiltration, and reduced myogenic reactivity of small arteries. Circ Res. 2003 Dec 12;93(12):1249–1257. doi: 10.1161/01.RES.0000104086.43830.6C. [DOI] [PubMed] [Google Scholar]
- 15.Hao L, Du M, Lopez-Campistrous A, Fernandez-Patron C. Agonist-induced activation of matrix metalloproteinase-7 promotes vasoconstriction through the epidermal growth factor-receptor pathway. Circ Res. 2004 Jan 9;94(1):68–76. doi: 10.1161/01.RES.0000109413.57726.91. [DOI] [PubMed] [Google Scholar]
- 16.Friese RS, Rao F, Khandrika S, et al. Matrix metalloproteinases: discrete elevations in essential hypertension and hypertensive end-stage renal disease. Clin Exp Hypertens. 2009 Oct;31(7):521–533. doi: 10.3109/10641960802668730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Rodrigues SF, Tran ED, Fortes ZB, Schmid-Schonbein GW. Matrix metalloproteinases cleave the beta2-adrenergic receptor in spontaneously hypertensive rats. Am J Physiol Heart Circ Physiol. 2010 Jul;299(1):H25–35. doi: 10.1152/ajpheart.00620.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.DeLano FA, Schmid-Schonbein GW. Proteinase Activity and Receptor Cleavage: Mechanism for Insulin Resistance in the Spontaneously Hypertensive Rat. Hypertension. 2008;52(2):415–423. doi: 10.1161/HYPERTENSIONAHA.107.104356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Castro MM, Rizzi E, Figueiredo-Lopes L, et al. Metalloproteinase inhibition ameliorates hypertension and prevents vascular dysfunction and remodeling in renovascular hypertensive rats. Atherosclerosis. 2008 Jun;198(2):320–331. doi: 10.1016/j.atherosclerosis.2007.10.011. [DOI] [PubMed] [Google Scholar]
- 20.Merchant SJ, Narumiya H, Zhang Y, Guilbert LJ, Davidge ST. The effects of preeclampsia and oxygen environment on endothelial release of matrix metalloproteinase-2. Hypertens Pregnancy. 2004;23(1):47–60. doi: 10.1081/PRG-120028281. [DOI] [PubMed] [Google Scholar]
- 21.Lekontseva ON, Rueda-Clausen CF, Morton JS, Davidge ST. Ovariectomy in aged versus young rats augments matrix metalloproteinase-mediated vasoconstriction in mesenteric arteries. Menopause. 2010 May-Jun;17(3):516–523. doi: 10.1097/gme.0b013e3181c91f04. [DOI] [PubMed] [Google Scholar]
- 22.Lekontseva O, Jiang Y, Davidge ST. Estrogen replacement increases matrix metalloproteinase contribution to vasoconstriction in a rat model of menopause. J Hypertens. 2009 Aug;27(8):1602–1608. doi: 10.1097/HJH.0b013e32832c41b5. [DOI] [PubMed] [Google Scholar]
- 23.Conrad KP, Davison JM. The renal circulation in normal pregnancy and preeclampsia: is there a place for relaxin? Am J Physiol Renal Physiol. 2014 May 15;306(10):F1121–1135. doi: 10.1152/ajprenal.00042.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Jeyabalan A, Novak J, Doty KD, et al. Vascular matrix metalloproteinase-9 mediates the inhibition of myogenic reactivity in small arteries isolated from rats after short-term administration of relaxin. Endocrinology. 2007 Jan;148(1):189–197. doi: 10.1210/en.2006-0989. [DOI] [PubMed] [Google Scholar]
- 25.Narumiya H, Zhang Y, Fernandez-Patron C, Guilbert LJ, Davidge ST. Matrix metalloproteinase-2 is elevated in the plasma of women with preeclampsia. Hypertens Pregnancy. 2001;20(2):185–194. doi: 10.1081/PRG-100106968. [DOI] [PubMed] [Google Scholar]
- 26.Myers JE, Merchant SJ, Macleod M, Mires GJ, Baker PN, Davidge ST. MMP-2 levels are elevated in the plasma of women who subsequently develop preeclampsia. Hypertens Pregnancy. 2005;24(2):103–115. doi: 10.1081/PRG-200059836. [DOI] [PubMed] [Google Scholar]
- 27.Brennan LJ, Morton JS, Davidge ST. Vascular dysfunction in preeclampsia. Microcirculation. 2014 Jan;21(1):4–14. doi: 10.1111/micc.12079. [DOI] [PubMed] [Google Scholar]
- 28.Wang X, Berry E, Hernandez-Anzaldo S, Takawale A, Kassiri Z, Fernandez-Patron C. Matrix Metalloproteinase-2 Mediates a Mechanism of Metabolic Cardioprotection Consisting of Negative Regulation of the Sterol Regulatory Element-Binding Protein-2/3-Hydroxy-3-Methylglutaryl-CoA Reductase Pathway in the Heart. Hypertension. 2015 Apr;65(4):882–888. doi: 10.1161/HYPERTENSIONAHA.114.04989. [DOI] [PubMed] [Google Scholar]
- 29.Van Hul M, Lijnen HR. A functional role of gelatinase A in the development of nutritionally induced obesity in mice. J Thromb Haemost. 2008 Jul;6(7):1198–1206. doi: 10.1111/j.1538-7836.2008.02988.x. [DOI] [PubMed] [Google Scholar]
- 30.Lijnen HR, Demeulemeester D, Van Hoef B, Collen D, Maquoi E. Deficiency of tissue inhibitor of matrix metalloproteinase-1 (TIMP-1) impairs nutritionally induced obesity in mice. Thromb Haemost. 2003 Feb;89(2):249–255. [PubMed] [Google Scholar]
- 31.Lee SW, Song KE, Shin DS, et al. Alterations in peripheral blood levels of TIMP-1, MMP-2, and MMP-9 in patients with type-2 diabetes. Diabetes Res Clin Pract. 2005 Aug;69(2):175–179. doi: 10.1016/j.diabres.2004.12.010. [DOI] [PubMed] [Google Scholar]
- 32.Jaworski DM, Sideleva O, Stradecki HM, et al. Sexually dimorphic diet-induced insulin resistance in obese tissue inhibitor of metalloproteinase-2 (TIMP-2)-deficient mice. Endocrinology. 2011 Apr;152(4):1300–1313. doi: 10.1210/en.2010-1029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Van Hul M, Piccard H, Lijnen HR. Gelatinase B (MMP-9) deficiency does not affect murine adipose tissue development. Thromb Haemost. 2010 Jul;104(1):165–171. doi: 10.1160/TH09-10-0739. [DOI] [PubMed] [Google Scholar]
- 34.Van Hul M, Lijnen HR. Effect of weight loss on gelatinase levels in obese mice. Clin Exp Pharmacol Physiol. 2011 Sep;38(9):647–649. doi: 10.1111/j.1440-1681.2011.05552.x. [DOI] [PubMed] [Google Scholar]
- 35.Van Hul M, Bauters D, Lijnen RH. Differential effects of a gelatinase inhibitor on adipocyte differentiation and adipose tissue development. Clin Exp Pharmacol Physiol. 2013 Oct;40(10):689–697. doi: 10.1111/1440-1681.12154. [DOI] [PubMed] [Google Scholar]
- 36.Van Hul M, Bauters D, Himmelreich U, et al. Effect of gelatinase inhibition on adipogenesis and adipose tissue development. Clin Exp Pharmacol Physiol. 2012 Jan;39(1):49–56. doi: 10.1111/j.1440-1681.2011.05635.x. [DOI] [PubMed] [Google Scholar]
- 37.Lijnen HR, Silence J, Lemmens G, Frederix L, Collen D. Regulation of gelatinase activity in mice with targeted inactivation of components of the plasminogen/plasmin system. Thromb Haemost. 1998 Jun;79(6):1171–1176. [PubMed] [Google Scholar]
- 38.Bauters D, Van Hul M, Lijnen HR. Gelatinase B (MMP-9) gene silencing does not affect murine preadipocyte differentiation. Adipocyte. 2014 Jan 1;3(1):50–53. doi: 10.4161/adip.26966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Park JY, Park JH, Jang W, et al. Apolipoprotein A-IV is a novel substrate for matrix metalloproteinases. Journal of Biochemistry. 2011;151(3):291–298. doi: 10.1093/jb/mvr137. [DOI] [PubMed] [Google Scholar]
- 40.Park JH, Park S-M, Park S-H, Cho K-H, Lee S-T. Cleavage and functional loss of human apolipoprotein E by digestion of matrix metalloproteinase-14. Proteomics. 2008;8(14):2926–2935. doi: 10.1002/pmic.200700487. [DOI] [PubMed] [Google Scholar]
- 41.Park JH, Park S-M, Park K-H, Cho K-H, Lee S-T. Analysis of apolipoprotein A-I as a substrate for matrix metalloproteinase-14. Biochemical and Biophysical Research Communications. 2011;409(1):58–63. doi: 10.1016/j.bbrc.2011.04.105. [DOI] [PubMed] [Google Scholar]
- 42.Marcel YL, Kiss RS. Structure-function relationships of apolipoprotein A-I: a flexible protein with dynamic lipid associations. Curr Opin Lipidol. 2003 Apr;14(2):151–157. doi: 10.1097/00041433-200304000-00006. [DOI] [PubMed] [Google Scholar]
- 43.Mahley RW, Innerarity TL, Rall SC, Jr, Weisgraber KH. Plasma lipoproteins: apolipoprotein structure and function. J Lipid Res. 1984 Dec 1;25(12):1277–1294. [PubMed] [Google Scholar]
- 44.Kim SY, Park SM, Lee S-T. Apolipoprotein C-II is a novel substrate for matrix metalloproteinases. Biochemical and Biophysical Research Communications. 2006;339(1):47–54. doi: 10.1016/j.bbrc.2005.10.182. [DOI] [PubMed] [Google Scholar]
- 45.Beisiegel U, Weber W, Ihrke G, Herz J, Stanley KK. The LDL-receptor-related protein, LRP, is an apolipoprotein E-binding protein. Nature. 1989 Sep 14;341(6238):162–164. doi: 10.1038/341162a0. [DOI] [PubMed] [Google Scholar]
- 46.Aoki T, Sato D, Li Y, Takino T, Miyamori H, Sato H. Cleavage of apolipoprotein E by membrane-type matrix metalloproteinase-1 abrogates suppression of cell proliferation. J Biochem. 2005 Jan;137(1):95–99. doi: 10.1093/jb/mvi009. [DOI] [PubMed] [Google Scholar]
- 47.Wang X, Berry E, Hernandez-Anzaldo S, et al. MMP-2 inhibits PCSK9-induced degradation of the LDL receptor in Hepa1-c1c7 cells. FEBS Lett. 2015 Feb 13;589(4):490–496. doi: 10.1016/j.febslet.2015.01.007. [DOI] [PubMed] [Google Scholar]
- 48.Horton JD, Cohen JC, Hobbs HH. PCSK9: a convertase that coordinates LDL catabolism. J Lipid Res. 2009 Apr;50(Suppl):S172–177. doi: 10.1194/jlr.R800091-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Benjannet S, Rhainds D, Essalmani R, et al. NARC-1/PCSK9 and its natural mutants: zymogen cleavage and effects on the low density lipoprotein (LDL) receptor and LDL cholesterol. J Biol Chem. 2004 Nov 19;279(47):48865–48875. doi: 10.1074/jbc.M409699200. [DOI] [PubMed] [Google Scholar]
- 50.Abifadel M, Varret M, Rabes JP, et al. Mutations in PCSK9 cause autosomal dominant hypercholesterolemia. Nat Genet. 2003 Jun;34(2):154–156. doi: 10.1038/ng1161. [DOI] [PubMed] [Google Scholar]
- 51.Doucet A, Overall CM. Protease proteomics: revealing protease in vivo functions using systems biology approaches. Mol Aspects Med. 2008 Oct;29(5):339–358. doi: 10.1016/j.mam.2008.04.003. [DOI] [PubMed] [Google Scholar]
- 52.O’Reilly MS, Wiederschain D, Stetler-Stevenson WG, Folkman J, Moses MA. Regulation of angiostatin production by matrix metalloproteinase-2 in a model of concomitant resistance. J Biol Chem. 1999 Oct 8;274(41):29568–29571. doi: 10.1074/jbc.274.41.29568. [DOI] [PubMed] [Google Scholar]
- 53.Martignetti JA, Aqeel AA, Sewairi WA, et al. Mutation of the matrix metalloproteinase 2 gene (MMP2) causes a multicentric osteolysis and arthritis syndrome. Nat Genet. 2001 Jul;28(3):261–265. doi: 10.1038/90100. [DOI] [PubMed] [Google Scholar]
- 54.Mosig RA, Dowling O, DiFeo A, et al. Loss of MMP-2 disrupts skeletal and craniofacial development and results in decreased bone mineralization, joint erosion and defects in osteoblast and osteoclast growth. Hum Mol Genet. 2007 May 1;16(9):1113–1123. doi: 10.1093/hmg/ddm060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Tuysuz B, Mosig R, Altun G, Sancak S, Glucksman MJ, Martignetti JA. A novel matrix metalloproteinase 2 (MMP2) terminal hemopexin domain mutation in a family with multicentric osteolysis with nodulosis and arthritis with cardiac defects. Eur J Hum Genet. 2009 May;17(5):565–572. doi: 10.1038/ejhg.2008.204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Greenlee KJ, Werb Z, Kheradmand F. Matrix metalloproteinases in lung: multiple, multifarious, and multifaceted. Physiol Rev. 2007 Jan;87(1):69–98. doi: 10.1152/physrev.00022.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Corry DB, Rishi K, Kanellis J, et al. Decreased allergic lung inflammatory cell egression and increased susceptibility to asphyxiation in MMP2-deficiency. Nat Immunol. 2002 Apr;3(4):347–353. doi: 10.1038/ni773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Corry DB, Kiss A, Song LZ, et al. Overlapping and independent contributions of MMP2 and MMP9 to lung allergic inflammatory cell egression through decreased CC chemokines. FASEB J. 2004 Jun;18(9):995–997. doi: 10.1096/fj.03-1412fje. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Mosig RA, Martignetti JA. Loss of MMP-2 in murine osteoblasts upregulates osteopontin and bone sialoprotein expression in a circuit regulating bone homeostasis. Dis Model Mech. 2013 Mar;6(2):397–403. doi: 10.1242/dmm.007914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Berry E, Hernandez-Anzaldo S, Ghomashchi F, et al. Matrix metalloproteinase-2 negatively regulates cardiac secreted phospholipase A2 to modulate inflammation and fever. J Am Heart Assoc. 2015;4(4) doi: 10.1161/JAHA.115.001868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Lambeau G, Gelb MH. Biochemistry and physiology of mammalian secreted phospholipases A2. Annu Rev Biochem. 2008;77:495–520. doi: 10.1146/annurev.biochem.76.062405.154007. [DOI] [PubMed] [Google Scholar]
- 62.Ou J, Tu H, Shan B, et al. Unsaturated fatty acids inhibit transcription of the sterol regulatory element-binding protein-1c (SREBP-1c) gene by antagonizing ligand-dependent activation of the LXR. Proc Natl Acad Sci U S A. 2001 May 22;98(11):6027–6032. doi: 10.1073/pnas.111138698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Hartl D, Krauss-Etschmann S, Koller B, et al. Infiltrated neutrophils acquire novel chemokine receptor expression and chemokine responsiveness in chronic inflammatory lung diseases. J Immunol. 2008 Dec 1;181(11):8053–8067. doi: 10.4049/jimmunol.181.11.8053. [DOI] [PubMed] [Google Scholar]
- 64.Monden Y, Kubota T, Inoue T, et al. Tumor necrosis factor-alpha is toxic via receptor 1 and protective via receptor 2 in a murine model of myocardial infarction. Am J Physiol Heart Circ Physiol. 2007 Jul;293(1):H743–753. doi: 10.1152/ajpheart.00166.2007. [DOI] [PubMed] [Google Scholar]
- 65.Matsumura S, Iwanaga S, Mochizuki S, Okamoto H, Ogawa S, Okada Y. Targeted deletion or pharmacological inhibition of MMP-2 prevents cardiac rupture after myocardial infarction in mice. J Clin Invest. 2005 Mar;115(3):599–609. doi: 10.1172/JCI22304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Hayashidani S, Tsutsui H, Ikeuchi M, et al. Targeted deletion of MMP-2 attenuates early LV rupture and late remodeling after experimental myocardial infarction. Am J Physiol Heart Circ Physiol. 2003 Sep;285(3):H1229–1235. doi: 10.1152/ajpheart.00207.2003. [DOI] [PubMed] [Google Scholar]
- 67.Lijnen HR, Collen D. Matrix metalloproteinase system deficiencies and matrix degradation. Thromb Haemost. 1999 Aug;82(2):837–845. [PubMed] [Google Scholar]
- 68.Pinto YM, Heymans S. Letter by Pinto and Heymans regarding article, “Ablation of matrix metalloproteinase-9 increases severity of viral myocarditis in mice”. Circulation. 2008 Nov 11;118(20):e697. doi: 10.1161/CIRCULATIONAHA.108.787697. [DOI] [PubMed] [Google Scholar]
- 69.Cheung C, Marchant D, Walker EK, et al. Ablation of matrix metalloproteinase-9 increases severity of viral myocarditis in mice. Circulation. 2008 Mar 25;117(12):1574–1582. doi: 10.1161/CIRCULATIONAHA.107.733238. [DOI] [PubMed] [Google Scholar]
- 70.Westermann D, Savvatis K, Lindner D, et al. Reduced degradation of the chemokine MCP-3 by matrix metalloproteinase-2 exacerbates myocardial inflammation in experimental viral cardiomyopathy. Circulation. 2011 Nov 8;124(19):2082–2093. doi: 10.1161/CIRCULATIONAHA.111.035964. [DOI] [PubMed] [Google Scholar]
- 71.Dennis EA, Cao J, Hsu YH, Magrioti V, Kokotos G. Phospholipase A2 enzymes: physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem Rev. 2011 Oct 12;111(10):6130–6185. doi: 10.1021/cr200085w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Murakami M, Sato H, Miki Y, Yamamoto K, Taketomi Y. A New Era of Secreted Phospholipase A2 (sPLA2) J Lipid Res. 2015 Mar 24; doi: 10.1194/jlr.R058123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Seftor RE, Seftor EA, De Larco JE, et al. Chemically modified tetracyclines inhibit human melanoma cell invasion and metastasis. Clin Exp Metastasis. 1998 Apr;16(3):217–225. doi: 10.1023/a:1006588708131. [DOI] [PubMed] [Google Scholar]
- 74.Greenwald RA, Moak SA, Ramamurthy NS, Golub LM. Tetracyclines suppress matrix metalloproteinase activity in adjuvant arthritis and in combination with flurbiprofen, ameliorate bone damage. J Rheumatol. 1992 Jun;19(6):927–938. [PubMed] [Google Scholar]
- 75.Golub LM, McNamara TF, D’Angelo G, Greenwald RA, Ramamurthy NS. A non-antibacterial chemically-modified tetracycline inhibits mammalian collagenase activity. J Dent Res. 1987 Aug;66(8):1310–1314. doi: 10.1177/00220345870660080401. [DOI] [PubMed] [Google Scholar]
- 76.Golub LM, Wolff M, Lee HM, et al. Further evidence that tetracyclines inhibit collagenase activity in human crevicular fluid and from other mammalian sources. J Periodontal Res. 1985 Jan;20(1):12–23. doi: 10.1111/j.1600-0765.1985.tb00405.x. [DOI] [PubMed] [Google Scholar]
- 77.Gu Y, Lee HM, Sorsa T, et al. Non-antibacterial tetracyclines modulate mediators of periodontitis and atherosclerotic cardiovascular disease: a mechanistic link between local and systemic inflammation. Pharmacol Res. 2011 Dec;64(6):573–579. doi: 10.1016/j.phrs.2011.06.023. [DOI] [PubMed] [Google Scholar]
- 78.Han KH, Hong KH, Park JH, et al. C-reactive protein promotes monocyte chemoattractant protein-1--mediated chemotaxis through upregulating CC chemokine receptor 2 expression in human monocytes. Circulation. 2004 Jun 1;109(21):2566–2571. doi: 10.1161/01.CIR.0000131160.94926.6E. [DOI] [PubMed] [Google Scholar]
- 79.Carnevale KA, Cathcart MK. Calcium-independent phospholipase A(2) is required for human monocyte chemotaxis to monocyte chemoattractant protein 1. J Immunol. 2001 Sep 15;167(6):3414–3421. doi: 10.4049/jimmunol.167.6.3414. [DOI] [PubMed] [Google Scholar]
- 80.Mishra RS, Carnevale KA, Cathcart MK. iPLA2beta: front and center in human monocyte chemotaxis to MCP-1. J Exp Med. 2008 Feb 18;205(2):347–359. doi: 10.1084/jem.20071243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Ii H, Hontani N, Toshida I, Oka M, Sato T, Akiba S. Group IVA phospholipase A2-associated production of MMP-9 in macrophages and formation of atherosclerotic lesions. Biol Pharm Bull. 2008 Mar;31(3):363–368. doi: 10.1248/bpb.31.363. [DOI] [PubMed] [Google Scholar]
- 82.Gorovetz M, Schwob O, Krimsky M, Yedgar S, Reich R. MMP production in human fibrosarcoma cells and their invasiveness are regulated by group IB secretory phospholipase A2 receptor-mediated activation of cytosolic phospholipase A2. Front Biosci. 2008;13:1917–1925. doi: 10.2741/2811. [DOI] [PubMed] [Google Scholar]
- 83.Choi YA, Kim DK, Bang OS, Kang SS, Jin EJ. Secretory phospholipase A2 promotes MMP-9-mediated cell death by degrading type I collagen via the ERK pathway at an early stage of chondrogenesis. Biol Cell. 2010 Feb;102(2):107–119. doi: 10.1042/BC20090073. [DOI] [PubMed] [Google Scholar]
- 84.Choi YA, Lim HK, Kim JR, et al. Group IB secretory phospholipase A2 promotes matrix metalloproteinase-2-mediated cell migration via the phosphatidylinositol 3-kinase and Akt pathway. J Biol Chem. 2004 Aug 27;279(35):36579–36585. doi: 10.1074/jbc.M314235200. [DOI] [PubMed] [Google Scholar]
- 85.Ping D, Jones PL, Boss JM. TNF regulates the in vivo occupancy of both distal and proximal regulatory regions of the MCP-1/JE gene. Immunity. 1996 May;4(5):455–469. doi: 10.1016/s1074-7613(00)80412-4. [DOI] [PubMed] [Google Scholar]
- 86.Andoh A, Takaya H, Saotome T, et al. Cytokine regulation of chemokine (IL-8, MCP-1, and RANTES) gene expression in human pancreatic periacinar myofibroblasts. Gastroenterology. 2000 Jul;119(1):211–219. doi: 10.1053/gast.2000.8538. [DOI] [PubMed] [Google Scholar]
- 87.Braunwald E, Harrison DC, Chidsey CA. The Heart as an Endocrine Organ. Am J Med. 1964 Jan;36:1–4. doi: 10.1016/0002-9343(64)90144-5. [DOI] [PubMed] [Google Scholar]
- 88.Ogawa T, de Bold AJ. The heart as an endocrine organ. Endocr Connect. 2014;3(2):R31–44. doi: 10.1530/EC-14-0012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Chakraborti S, Chowdhury A, Alam MN, et al. Vascular aneurysms: a perspective. Indian J Biochem Biophys. 2014 Dec;51(6):449–456. [PubMed] [Google Scholar]
- 90.Jackson CL, Raines EW, Ross R, Reidy MA. Role of endogenous platelet-derived growth factor in arterial smooth muscle cell migration after balloon catheter injury. Arterioscler Thromb. 1993 Aug;13(8):1218–1226. doi: 10.1161/01.atv.13.8.1218. [DOI] [PubMed] [Google Scholar]
- 91.Zeng S, Zhou X, Tu Y, et al. Long-term MMP inhibition by doxycycline exerts divergent effect on ventricular extracellular matrix deposition and systolic performance in stroke-prone spontaneously hypertensive rats. Clin Exp Hypertens. 2011;33(5):316–324. doi: 10.3109/10641963.2010.549262. [DOI] [PubMed] [Google Scholar]
- 92.Hori Y, Kunihiro S, Sato S, et al. Doxycycline attenuates isoproterenol-induced myocardial fibrosis and matrix metalloproteinase activity in rats. Biol Pharm Bull. 2009 Oct;32(10):1678–1682. doi: 10.1248/bpb.32.1678. [DOI] [PubMed] [Google Scholar]
- 93.Vinet L, Rouet-Benzineb P, Marniquet X, et al. Chronic doxycycline exposure accelerates left ventricular hypertrophy and progression to heart failure in mice after thoracic aorta constriction. Am J Physiol Heart Circ Physiol. 2008 Jul;295(1):H352–360. doi: 10.1152/ajpheart.01101.2007. [DOI] [PubMed] [Google Scholar]
- 94.Yen H, Zhang Y, Penfold S, Rollins BJ. MCP-1-mediated chemotaxis requires activation of non-overlapping signal transduction pathways. J Leukoc Biol. 1997 Apr;61(4):529–532. doi: 10.1002/jlb.61.4.529. [DOI] [PubMed] [Google Scholar]
- 95.Sonoki K, Iwase M, Ohdo S, Ieiri I, Takata Y, Kitazono T. Statin inhibits the expression of secretory phospholipase A2 and subsequent monocyte chemoattractant protein-1 in human endothelial cells. J Cardiovasc Pharmacol. 2014 Dec;64(6):489–496. doi: 10.1097/FJC.0000000000000147. [DOI] [PubMed] [Google Scholar]
- 96.Hu MM, Zhang J, Wang WY, et al. The inhibition of lipoprotein-associated phospholipase A2 exerts beneficial effects against atherosclerosis in LDLR-deficient mice. Acta Pharmacol Sin. 2011 Oct;32(10):1253–1258. doi: 10.1038/aps.2011.127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Goncalves I, Edsfeldt A, Ko NY, et al. Evidence supporting a key role of Lp-PLA2-generated lysophosphatidylcholine in human atherosclerotic plaque inflammation. Arterioscler Thromb Vasc Biol. 2012 Jun;32(6):1505–1512. doi: 10.1161/ATVBAHA.112.249854. [DOI] [PubMed] [Google Scholar]
- 98.Dean RA, Cox JH, Bellac CL, Doucet A, Starr AE, Overall CM. Macrophage-specific metalloelastase (MMP-12) truncates and inactivates ELR+ CXC chemokines and generates CCL2, -7, -8, and -13 antagonists: potential role of the macrophage in terminating polymorphonuclear leukocyte influx. Blood. 2008 Oct 15;112(8):3455–3464. doi: 10.1182/blood-2007-12-129080. [DOI] [PubMed] [Google Scholar]
- 99.Sommerfelt RM, Feuerherm AJ, Skuland T, Johansen B. Cytosolic phospholipase A2 modulates TLR2 signaling in synoviocytes. PLoS One. 2015;10(4):e0119088. doi: 10.1371/journal.pone.0119088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Kazantseva MG, Hung NA, Highton J, Hessian PA. MMP expression in rheumatoid inflammation: the rs11568818 polymorphism is associated with MMP-7 expression at an extra-articular site. Genes Immun. 2013 Apr;14(3):162–169. doi: 10.1038/gene.2012.65. [DOI] [PubMed] [Google Scholar]
- 101.McGarry T, Veale DJ, Gao W, Orr C, Fearon U, Connolly M. Toll like receptor 2 (TLR2) induces migration and invasive mechanisms in rheumatoid arthritis. Arthritis Res Ther. 2015 Jun 9;17(1):153. doi: 10.1186/s13075-015-0664-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Silosi I, Cojocaru M, Foia L, et al. Significance of circulating and crevicular matrix metalloproteinase-9 in rheumatoid arthritis-chronic periodontitis association. J Immunol Res. 2015;2015:218060. doi: 10.1155/2015/218060. [DOI] [PMC free article] [PubMed] [Google Scholar]