

Abstract
AIM: A low vessel density is a common feature of malignant tumors. We suggested that the expansion of vessel diameter might reconstitute the oxygen and nutritient’s supply in this situation. The aim of the present study was to compare the number and diameter of blood vessels in pancreatic and liver carcinoma with normal tissue.
METHODS: Tumor induction of pancreatic (DSL6A) or hepatocellular (Morris-hepatoma) carcinoma was performed in male Lewis (pancreatic cancer) and ACI (hepatoma) rats by an orthotopic inoculation of solid tumor fragments (pancreatic cancer) or tumor cells (hepatoma). Six weeks (pancreatic cancer) or 12 d (hepatoma) after tumor implantation, the tumor microvasculature as well as normal pancreatic or liver blood vessels were investigated by intravital microscopy. The number of perfused blood vessels in tumor and healthy tissue was assessed by computer-assisted image analysis.
RESULTS: The vessel density in healthy pancreas (565 ± 89 n/mm2) was significantly higher compared to pancreatic cancer (116 ± 36 n/mm2) (P < 0.001). Healthy liver showed also a significantly higher vessel density (689 ± 36 n/mm2) compared to liver carcinoma (286 ± 32 n/mm2) (P < 0.01). The comparison of diameter frequency showed a significant increase of vessel diameter in both malignant tumors compared to normal tissue (P < 0.05).
CONCLUSION: The expansion of endothelial cells during tumor angiogenesis is accompanied to a large extent by an increase of vessel diameter rather than by formation of new blood vessels. This may be a possible adaptive mechanism by which experimental pancreatic and hepatocellular cancers expand their endothelial diffusion surface of endothelium to compensate for inadequate neoangiogenesis.
INTRODUCTION
Angiogenesis, defined as proliferation of endothelial cells and subsequent formation of new blood vessels from pre-existing vessels, is a characteristic feature of numerous pathological processes, including cancer[1,2]. Tumor growth depends on a continuous blood supply to cover increasing requirements for nutrients and oxygen. During tumor neovascularization, new capillaries are recruited from the existing microvasculature, a process that is controlled by positive and negative regulators of blood vessel growth[3]. In contrast to the angiogenesis occurring in wound healing, the tumor angiogenesis is characterized by a discordance between pro- and anti-angiogenic factors. This discordant process of tumor angiogenesis leads to the formation of a microvascular system with a typically distorted vessel architecture[4,5], and consequently irregular flow patterns and heterogeneous oxygen supply[6,7] resulting in hypoxia[8,9]. These parameters contribute to a unique tumor microenvironment which in turn modulates the therapeutic responsiveness of solid tumors, e.g., towards chemotherapeutic agents or radiation therapy[10,11].
Our previous studies reported that the increased vessel diameter was an important feature of tumor microangioarchitecture in pancreatic[12] and hepatocellular[13] cancer of the rat. This feature was not only specific for transplantable tumor models and was found in spontaneous pancreatic tumors in transgenic mice[14]. We showed that the vessel dilation was the first detectable stage of ongoing angiogenesis during multistep tumorigenesis which preceded vessel sprouting[14]. These observations encouraged us to start a detailed investigation of tumor blood vessels by a combination of different techniques including intravital microscopy and immunohistochemistry. We hypothesized that the vessel dilation indicated a possible adaptive process, which may help the tumor to expand their diffusion surface of endothelium and to compensate for the inadequate supply with oxygen and nutritients. In the present study, we used intravital microscopy and immunohistochemistry to investigate this phenomenon in normal and malignant pancreatic tissues as well as in hepatic tissue of rats. The present study demonstrated that the decrease of vessel density in experimental pancreatic and liver carcinoma was accompanied with an increase of vessel diameter.
MATERIALS AND METHODS
Tumor inoculation
Established cell lines of pancreatic duct-like (DSL6A)[15] and hepatocellular carcinoma (Morris-hepatoma MH-3924A)[13] of rats were used. Since the expression profile of tumor cells was strongly dependent on the site of growth, e.g. implantation[16,17] we decided to inoculate the tumors orthotopically which ensured the appropriate microenvironment for tumor cell growth. Twelve male Lewis rats (160-180 g) were used for the inoculation of pancreatic carcinoma. Hepatocellular carcinomas were inoculated on 12 male ACI rats (220-250 g). Each animal was anaesthetized with intramuscular injection of xylazin (10 mg/kg, Rompun®, Bayer, Leverkusen, Germany) and ketamin (40 mg/kg, Ketanest®, Parke Davis, Berlin, FRG).
Tumor implantation was performed as previously described for pancreatic[12] and hepatocellular[13] carcinoma. For the inoculation of pancreatic cancer, two sterile polymethylmethacrylat (PMMA)-plates (∅ 11 mm, Glasflex, Stirling, NJ, USA) were applied in “sandwich-technique” on both sides of the pancreatic head. The tumor of approximately 1 mm3 was harvested from a subcutaneous parent tumor of a syngeneic rat and interposed intrapancreatically between the plates. Hepatocellular carcinomas were induced by a subcapsular injection of tumor cells (Morris hepatoma MH-3924A, 0.01 mL, 0.5 × 10 5 cells) into the left upper liver lobe. Major steps of both models are summarized in Table 1.
Table 1.
Summary of tumor models used in present study
| Pancreatic cance | Hepatocellular cancer | |
| Tumor cell line | DSL6A | Morris hepatoma 3924A |
| Syngeneic strain | Lewis | ACI |
| Tumor inoculation | Solid tumor fragment | Cell suspension |
| Time after inoculation | 6 wk | 12 d |
Intravital microscopy
Intravital microscopy was performed 12 d after inoculation for hepatocellular carcinoma and 6 wk after inoculation for pancreatic cancer when tumors reached a diameter of 8-10 mm. All animals were re-anaesthetized as described above. A teflon catheter (I.D. 0.5 mm, B.Braun AG, Melsungen, FRG) was inserted into the right internal jugular vein for venous access. Another catheter was placed into the left carotid artery for blood sampling and monitoring of cardiovascular parameters. The abdomen was opened by midline incision. The animal was placed on a special stage automatically maintained at 37 °C. The tumor was macroscopically identified. The tumor bearing pancreatic head was immobilized in a temperature-controlled (37 °C) immersion chamber containing Ringer’s solution. For the intravital microscopy of hepatocellular carcinoma, the tumor bearing liver lobe was placed on a rubber stage and superfused with Ringer’s solution (37 °C).
For intravital microscopy, the entire preparation was placed under a fluorescence microscope (Leica GmbH, Wetzlar, FRG). The fluorescence filter with excitation 450-490 nm and emission 520 nm was used. After the preparation was completed, all animals received an intravenous injection of 50 mg/kg of FITC-labeled albumin (Sigma Chemicals Co., St.Louis, MO, USA) dissolved in 1 mL saline. This fluorescent plasma marker ensured the maximal contrast of blood vessels and was utilized for the measurement of vessel diameter and vessel density. The microcirculatory images were transmitted by a video camera (CF 8/1, Kappa GmbH, Gleichen, FRG) to a monitor (PVM-1440 M, Sony, Tokyo, Japan) and recorded on a videorecorder (sVHS, AG-7 350-E, Panasonic, Osaka, Japan) for subsequent off-line analysis. After the experiment, the tumors and a fragment of normal tissue were harvested and frozen immediately in liquid nitrogen. The evaluation of vessel diameter and density in tumor microcirculation and in normal tissue was performed using special software (Capimage®, Zeintl GmbH, Heidelberg, FRG). The number of blood vessels was calculated on 3-4 randomly chosen fields and expressed as per 1 mm2 of tumor surface.
Immunohistochemistry
After intravital microscopy, the tumors were removed for further histological analysis. The tissue was snap frozen in liquid nitrogen. Five µm thick sections were cut, air-dried and fixed in aceton. Immunohistochemical staining of endothelium was performed using monoclonal antibodies recognising RECA-1 rat endothelial antigen (Clone HIS52, Serotec, Germany) and the LSAB-kit (Dako, Hamburg, Germany). In addition, the slides were counterstained with Mayer’s acid hemalum (Fluka, Steinheim, Germany). Quantitative analysis of immunohistochemical staining was performed by computer-assisted image analysis. For this aim, three microscopic fields of 3.18 mm2 were randomly chosen by light microscope (Leica DMRB, Leica GmbH, Germany), digitalized by a colour video camera (CF 20/4DX, Kappa GmbH, Gleichen, Germany) to histological images and saved on a computer. The number of blood vessels was counted and expressed as per 1 mm2 of surface.
Statistical analysis
All data were given as mean ± SD. Mann-Whitney-U test was used to compare the differences between groups as appropriate. P < 0.05 was considered statistically significant.
RESULTS
The intravital microscopy after injection of fluorescent plasma marker ensured the excellent contrast of all blood vessels in normal (Figure 1A, Figure 1E) and in tumoral (Figure 1B, Figure 1F) tissues. The microvascular system in healthy pancreas showed a capillary network of high density, the single afferent and efferent blood vessels were identified within normal pancreatic tissue (Figure 1A). Healthy liver showed a dense network of hepatic sinusoids which were drained by hepatic venules (Figure 1E). In contrast, the microangioarchtecture of both pancreatic and hepatocellular carcinomas was characterized by lost of normal vascular hierarchy “capillaries-arterioles (sinusoids) -venules”, chaotic arrangement of blood vessels, irregular vessel diameters and formation of lacunar blood vessels (Figure 1B, Figure 1F). In contrast to vascular systems of normal pancreatic and hepatic tissues, there were no specific features discriminating the microangioarchitecture of pancreatic cancer from that of hepatocellular carcinoma (Figure 1).
Figure 1.
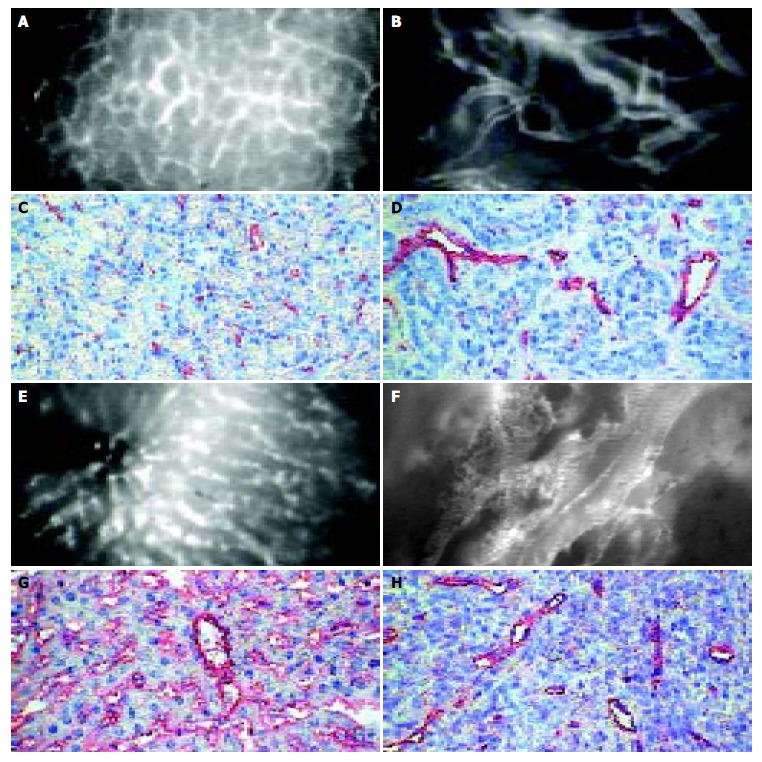
Microangioarchtecture investigated by intravital microscopy and immunohistochemical staining of endothelium of normal pancreas (A,B), liver (E,F), pancreatic (C,D) and hepatocellular (G,H) carcinoma: The microvascular system in healthy pancreas showed a dense network mainly consisting of capillaries in normal pancreas (A) and sinusoids in the liver (E). Both pancreatic (B) and hepatocellular (F) carcinomas showed a chaotic angioarchitecture with irregular diameter of blood vessels. Either intravital microscopy or immunohistochemical staining of endothelium turn up lower density and higher diameters of microvessels in both tumor types than in corresponding normal tissue (bar 50 µm).
The measurement of vessel density either by intravital microscopy or by immunohistochemistry demonstrated different results (Table 2, Table 3). The vessel density of all tissues measured by intravital microscopy was significantly higher than that measured by immunohistochemistry (P < 0.05, Table 2, Table 3). The vessel density in healthy pancreas was significantly higher compared to pancreatic cancer (P < 0.001, Table 2). Healthy liver showed also a significantly higher vessel density compared to liver carcinoma (P < 0.01, Table 3). The low vessel density of both pancreatic and hepatocellular carcinomas was accompanied with the development of tumor necrosis, which appeared frequently in the central area of the tumors and represented a characteristical feature of the tumors showing a cross-sectional diameter of more than 10 mm as opposed to smaller tumors.
Table 2.
Vessel density in normal and malignant tissue
| Vessel density (n/mm2) | Healthy pancreas | Pancreatic cancer | P |
| Intravital microscopy | 565 ± 89 | 116 ± 36 | < 0.001 |
| Immunohistochemistry | 91 ± 22 | 35 ± 10 | < 0.001 |
Table 3.
Vessel density in normal and malignant liver tissue
| Vessel density (n/mm2) | Healthy liver | Liver carcinoma | P |
| Intravital microscopy | 689 ± 36 | 286 ± 32 | < 0.01 |
| Immunohistochemistry | 196 ± 30 | 49 ± 13 | < 0.01 |
The frequency analysis of vessel diameters showed that the most frequent diameters in normal pancreas and liver were considerably smaller compared to the most frequent diameters in pancreatic and hepatocellular carcinomas (Figure 2, Figure 3). Ninety-four percent of blood vessels in normal pancreas, 41% of blood vessels in pancreatic carcinoma and less than 9% of blood vessels in non-malignant and malignant hepatocellular tissues showed a diameter less than 10 µm. The most frequent diameter was between 10 and 20 µm (59%) in pancreatic carcinoma and more than 20 µm in hepatocellular carcinoma (59%).
Figure 2.
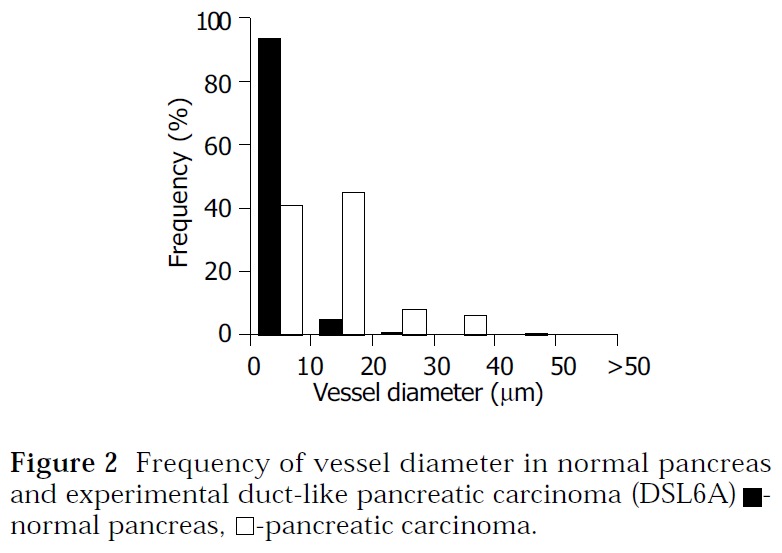
Frequency of vessel diameter in normal pancreas and experimental duct-like pancreatic carcinoma (DSL6A) ■- normal pancreas, □-pancreatic carcinoma.
Figure 3.
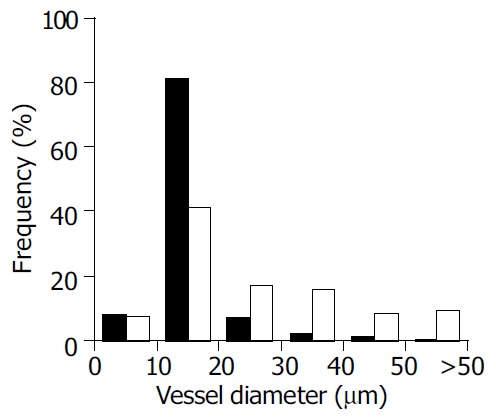
Frequency of vessel diameter in normal liver and experimental hepatocellular carcinoma (Morris-hepatoma) ■- normal liver, □-hepatocellular carcinoma.
DISCUSSION
Using intravital microscopy, we investigated the microvascular systems of pancreatic and liver carcinoma and performed a quantitative analysis of vessel density and vessel diameter. The present study demonstrated that the mean vessel density in both types of experimental malignant tumors was lower than that in the corresponding normal tissues. This hypovascularity was regarded as a main cause of hypoxia in tumor tissue which is responsible for the development of necrotic areas within solid tumors[8,11] as it has been observed in the present models of pancreatic and hepatocellular carcinoma once diameters exceeded 10 mm. However, as it was shown in the present study, tumor angiogenesis could compensate in part for this insufficient vascularisation by other mechanisms than growth of new blood vessels. Although vessel sprouting has been accepted as the main mechanism of angiogenesis, some pathological processes might display an angiogenic phenotype characterized by an extensive vessel dilatation[18,19]. Previous data demonstrated in a model of multistep tumorigenesis in mouse malignant insulinoma[14] and in the model of thyroid hyperplasia[18] that vessel dilatation was the first detectable stage of ongoing angiogenesis which preceded vessel sprouting and extended transformation of the cells. The results of the present study allowed to transfer these findings to solid murine tumors and demonstrated that the “non-sprouting” angiogenesis might accompany vessel sprouting during growth of solid pancreatic and hepatocellular cancer in rats. We suggested that the observed vessel dilatation during tumor development and growth could cause an increase of endothelial surface for the diffusion of oxygen and nutrition, representing a possible adaptive mechanism to compensate for the insufficient vascularisation.
In the present study, both tumor types were induced by an implantation of tumor cells and did not arise spontaneously. It is known that tumor blood vessels in transplantable tumor models develop from surrounding tissues[20,21], whereas the growth of autochtonous blood vessels forms the vascular system in spontaneous tumors[22,23]. However, the vessel dilatation as an initial step of the vascular transformation has been reported previously in spontaneous tumor model also and seems to be a feature which is likely to be a common feature in both transplanted and spontaneous tumors.
In a previous study we analysed the percentage of vascular surface of pancreatic cancer DSL6A which was performed on histological sections using intravascular perfusion with a fluorescent-labeled plasma marker[12]. In the present study, the number of blood vessels per mm2 was investigated by intravital microscopy and immunohistochemistry. The values of vessel density obtained by these analyses corresponded well and both showed a significantly lower microvascularity of malignant tumors compared to normal tissues. The vessel density investigated by intravital microscopy was higher than that by immunohistochemistry. Since the optical depth of tissue accessable for the intravital microscopy (30-50 µm) is considerably higher than the thickness of histological sections (5 µm), the difference between the results of intravital microscopy and immunohistochemistry finds its logical explanation.
In summary, we compared the vessel density and vessel diameter of two experimental tumors by intravital microscopy. The present study demonstrates that the vessel dilatation is an integral part of the angiogenic activity which represents a possible mechanism by which the tumors expand their diffusion surface of endothelium to compensate for the inadequate neoangiogenesis.
Footnotes
Edited by Wang XL Proofread by Xu FM
References
- 1.Carmeliet P, Jain RK. Angiogenesis in cancer and other diseases. Nature. 2000;407:249–257. doi: 10.1038/35025220. [DOI] [PubMed] [Google Scholar]
- 2.Folkman J. Angiogenesis in cancer, vascular, rheumatoid and other disease. Nat Med. 1995;1:27–31. doi: 10.1038/nm0195-27. [DOI] [PubMed] [Google Scholar]
- 3.Hanahan D, Folkman J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell. 1996;86:353–364. doi: 10.1016/s0092-8674(00)80108-7. [DOI] [PubMed] [Google Scholar]
- 4.Konerding MA, van Ackern C, Steinberg F, Streffer C. Com-bined morphological approaches in the study of network for-mation in tumor angiogenesis. Steiner R, Weisz PB, Langer R eds. Angiogenesis: key principles-science-technology-medicine. Basel Birkhauser; 1992. pp. 40–58. [Google Scholar]
- 5.Steinberg F, Konerding MA, Sander A, Streffer C. Vascularization, proliferation and necrosis in untreated human primary tumours and untreated human xenografts. Int J Radiat Biol. 1991;60:161–168. doi: 10.1080/09553009114551751. [DOI] [PubMed] [Google Scholar]
- 6.Vaupel P, Thews O, Kelleher DK, Hoeckel M. Oxygenation of human tumors: the Mainz experience. Strahlenther Onkol. 1998;174(Suppl 4):6–12. [PubMed] [Google Scholar]
- 7.Dewhirst MW, Tso CY, Oliver R, Gustafson CS, Secomb TW, Gross JF. Morphologic and hemodynamic comparison of tumor and healing normal tissue microvasculature. Int J Radiat Oncol Biol Phys. 1989;17:91–99. doi: 10.1016/0360-3016(89)90375-1. [DOI] [PubMed] [Google Scholar]
- 8.West CM, Cooper RA, Loncaster JA, Wilks DP, Bromley M. Tumor vascularity: a histological measure of angiogenesis and hypoxia. Cancer Res. 2001;61:2907–2910. [PubMed] [Google Scholar]
- 9.Koong AC, Mehta VK, Le QT, Fisher GA, Terris DJ, Brown JM, Bastidas AJ, Vierra M. Pancreatic tumors show high levels of hypoxia. Int J Radiat Oncol Biol Phys. 2000;48:919–922. doi: 10.1016/s0360-3016(00)00803-8. [DOI] [PubMed] [Google Scholar]
- 10.Kerbel RS. Tumor angiogenesis: past, present and the near future. Carcinogenesis. 2000;21:505–515. doi: 10.1093/carcin/21.3.505. [DOI] [PubMed] [Google Scholar]
- 11.Brown JM, Le QT. Tumor hypoxia is important in radiotherapy, but how should we measure it. Int J Radiat Oncol Biol Phys. 2002;54:1299–1301. doi: 10.1016/s0360-3016(02)03936-6. [DOI] [PubMed] [Google Scholar]
- 12.Schmidt J, Ryschich E, Daniel V, Herzog L, Werner J, Herfarth C, Longnecker DS, Gebhard MM, Klar E. Vascular structure and microcirculation of experimental pancreatic carcinoma in rats. Eur J Surg. 2000;166:328–335. doi: 10.1080/110241500750009195. [DOI] [PubMed] [Google Scholar]
- 13.Maksan SM, Paulo H, Ryschich E, Kuntz C, Gebhard MM, Klar E, Schmidt J. In vivo assessment of angioarchitecture and microcirculation in experimental liver cancer: a new model in rats. Dig Dis Sci. 2003;48:279–290. doi: 10.1023/a:1021919224972. [DOI] [PubMed] [Google Scholar]
- 14.Ryschich E, Schmidt J, Hämmerling GJ, Klar E, Ganss R. Transformation of the microvascular system during multistage tumorigenesis. Int J Cancer. 2002;97:719–725. doi: 10.1002/ijc.10074. [DOI] [PubMed] [Google Scholar]
- 15.Pettengill OS, Faris RA, Bell RH, Kuhlmann ET, Longnecker DS. Derivation of ductlike cell lines from a transplantable acinar cell carcinoma of the rat pancreas. Am J Pathol. 1993;143:292–303. [PMC free article] [PubMed] [Google Scholar]
- 16.Gullino PM. Microenvironment and angiogenic response. Steiner R, Weisz PB, Langer R eds. Angiogenesis: key principles-sci-ence-technology-medicine. Basel Birkhauser; 1992. pp. 125–128. [Google Scholar]
- 17.Fukumura D, Yuan F, Monsky WL, Chen Y, Jain RK. Effect of host microenvironment on the microcirculation of human colon adenocarcinoma. Am J Pathol. 1997;151:679–688. [PMC free article] [PubMed] [Google Scholar]
- 18.Many MC, Denef JF, Haumont S. Precocity of the endothelial proliferation during a course of rapid goitrogenesis. Acta Endocrinol (Copenh) 1984;105:487–491. doi: 10.1530/acta.0.1050487. [DOI] [PubMed] [Google Scholar]
- 19.Bull RH, Bates DO, Mortimer PS. Intravital video-capillaroscopy for the study of the microcirculation in psoriasis. Br J Dermatol. 1992;126:436–445. doi: 10.1111/j.1365-2133.1992.tb11815.x. [DOI] [PubMed] [Google Scholar]
- 20.Sckell A, Safabakhsh N, Dellian M, Jain RK. Primary tumor size-dependent inhibition of angiogenesis at a secondary site: an intravital microscopic study in mice. Cancer Res. 1998;58:5866–5869. [PubMed] [Google Scholar]
- 21.Vajkoczy P, Thurnher A, Hirth KP, Schilling L, Schmiedek P, Ullrich A, Menger MD. Measuring VEGF-Flk-1 activity and consequences of VEGF-Flk-1 targeting in vivo using intravital microscopy: clinical applications. Oncologist. 2000;5(Suppl 1):16–19. doi: 10.1634/theoncologist.5-suppl_1-16. [DOI] [PubMed] [Google Scholar]
- 22.Folkman J, Watson K, Ingber D, Hanahan D. Induction of angiogenesis during the transition from hyperplasia to neoplasia. Nature. 1989;339:58–61. doi: 10.1038/339058a0. [DOI] [PubMed] [Google Scholar]
- 23.Bergers G, Javaherian K, Lo KM, Folkman J, Hanahan D. Effects of angiogenesis inhibitors on multistage carcinogenesis in mice. Science. 1999;284:808–812. doi: 10.1126/science.284.5415.808. [DOI] [PubMed] [Google Scholar]