

Abstract
AIM: To determine the dynamic changes in the expression of matrix metalloproteinases (MMPs) and the endogenous tissue inhibitors of MMPs inhibitors (TIMPs) during hepatic fibrosis induced by alcohol.
METHODS: Male Sprague-Dawley rats were randomly divided into normal, 4 d, 2 wk, 4 wk, 9 wk and 11 wk groups, and the model rats were fed with a mixture of alcohol by gastric infusion at the designed time, respectively, then decollated and their livers were harvested for the examination of MMP-2, MMP-3, MMP-9, MMP-13, TIMP-1 and TIMP-2 by immunoh-istochemistry, zymograghy and Western blotting, respectively.
RESULTS: Normal rats had moderate expression of MMP-2, which was decreased in the model rats except in the 11 wk group, where MMP-2 expression slightly increased. MMP-3 had the similar changing pattern to MMP-2 despite weaker expression. MMP-9 expression decreased in the 4 d and 2 wk groups, rose in the 4 wk group, decreased again in the 9 wk group and returned to normal levels in the 11 wk group. MMP-13 expression decreased in the 4 d and 2 wk groups, and returned to normal levels in the 4 wk, 9 wk and 11 wk groups. TIMP-1 expression decreased in the 4 d and 2 wk groups, but sharply increased in the 4 wk group and sustained at a high level even after modeling was stopped for 2 wk. In normal rats TIMP-2 expression was strong. However, it decreased as soon as modeling began, and then gradually rose, but remained to a level lower than that in normal rats even after modeling was stopped for 2 wk.
CONCLUSION: MMP-2 may not always expresses at a high level during hepatic fibrosis. MMP-13 and MMP-3 are acutely affected by TIMP-1. In this model TIMP-1 is the most powerful factor imposed on capillarization and peri-sinusoidal fibrosis. TIMP-2 is the most effective regulator on the metabolism of type IV collagen located in the basement of sinus.
INTRODUCTION
China’s prosperity is accompanied with a dramatic rise in cases of alcoholic liver disease. Some scholars reported that hepatic pathological changes of alcoholic liver disease (ALD) included typical capillarization, peri-sinusoidal fibrosis, bridging fibrosis, and even cirrhosis which were often found in heavy drinkers who might have a drinking history over 10 to 20 years. In mild to moderate drinkers, however, the pathological changes which might be more popular at present were not thoroughly investigated. In order to probe into this field we improved a rat model of ALD induced by the gavage method, which is widely used among Chinese scholars to show the key mechanism of hepatic fibrosis. Liver fibrosis occurs as a consequence of net accumulation of matrix proteins (especially types I and III collagen) in response to liver injury. Liver fibrosis is underpinned by the activation of hepatic stellate cells (HSCs) to a myofibroblast like phenotype with a consequent increase in their synthesis of matrix proteins such as interstitial collagens that characterize fibrosis. The sinusoid and peri-sinusoidal space are the original site of fibrosis, which can badly affect the function of hepatocytes. The expression and ratio of matrix metalloproteinases/tissue inhibitor of matrix metalloproteinases (MMPs/TIMPs) are key factors to extra cellular matrix remodeling during hepatic fibrosis. The co-operation of urokinase and tissue plasminogen activator system is also very important. Considering that recent reports were almost all focused on MMPs/TIMPs, and that the dynamic changes of MMPs/TIMPs during ALD were not so clear, we decided to probe into such indexes. We have reported the early pathological changes of alcoholic liver disease located in sinusoid and peri-sinusoid (in another article), further elucidation of mechanism of such changes would depend on the investigation of the expression and ratio of MMPs/TIMPs during a dynamic course of fibrosis. So the dynamic changes of MMPs 2, 3, 9, 13, and TIMPs 1, 2 were worked out to show the mechanism of this new model.
MATERIALS AND METHODS
Materials
Male SD rats, weighing (150 ± 5) g, were purchased from Beijing Vital River Company , corn oil from Carafour Supermarket, xanthan gum and maltose from Beijing Chemical Agent Company, edible alcohol from Beijing General Alcohol Brewing Company, carbonyl iron and pirazole from Sigma, USA, First antibody to MMPs 2, 3, 9, 13 and TIMPs 1, 2 was purchased from Antibody Diagnostic Inc, ADI, USA with the help of its Chinese Dealer, Shanghai Long Island Antibody Company. PV-6001 kits were from PowerVision, USA. ZLI-9030 and ZLI9001 were from Beijing Zhongshan Company.
Methods
Rats grouping: Normal (6), 4 d (8), 2 wk (8), 4 wk (10), 9 wk (12), 11 wk (12).
Modeling: The model of ALD was induced by intragastric infusion ( gavage) of a mixture of alcohol ( 5 g/d·kg), pirazole (30 mg/d·kg), corn oil (3 mL/d·kg), and carbonyl iron (35 mg/d·kg, decreased to 15 mg after 4 wk), a little xanthan gum and maltose, one time a day for 5 d consecutively, with 2 d off per week, until the end of the ninth wk, and all rats were maintained on a standard diet with water ad libitum.
Liver tissue management: The rats were executed at the end of 4 d, 2 wk, 4 wk, 9 wk and 11 wk, respectively. Harvested livers were split and fixed for electron microscopy, hematoxylin and eosin and Masson complex staining, and a portion was snap frozen for biochemical and molecular analysis. Further sections were cut from each liver, deparaffinized, HE and Masson staining. Histological analysis of each liver was undertaken, then the sections subjected to amylopsin antigen retrieval before stained with the two-step method for antigen MMPs 2, 3, 9, 13 and TIMPs 1 and 2. The semi-quantitive computations were conducted on the image analyzing system MIS-2000 from 3Y Company, USA, The activity of MMP-2 and MMP-9, Western blotting of TIMPs 1 and 2 were undertaken with Bio-Rad Electrophoresis System, pictures were taken and analyzed with Image master VDS from Pharmacia Biotech Company.
Statistical analysis
All values were expressed as mean ± SD. ANOVA was used to determine the significance of differences among the six groups. P < 0.05 was considered statistically significant.
RESULTS
The expression of MMP-2 in normal rats was at moderate level. However, during modeling it showed reducing curve changes and without recovery even after modeling was stopped for 2 wk. (Figure 1).
Figure 1.
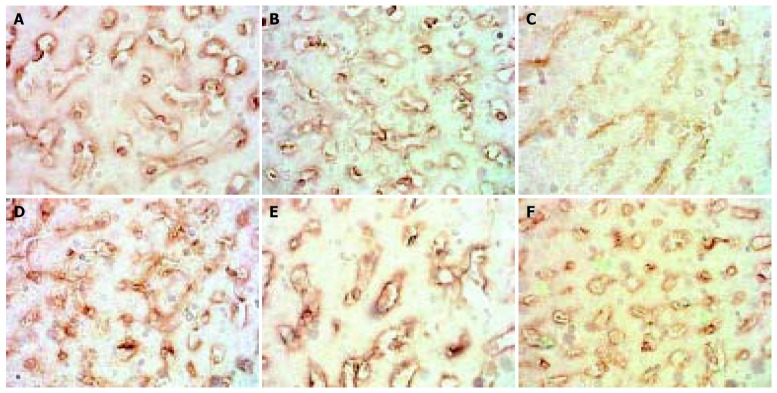
Dynamic changes of MMP-2. A: Normal rat, × 400 B: 4 d group, × 400 C: 2 wk group, × 400 D: 4 wk group, × 400. E: 9 wk group, × 400 F: 11 wk group, × 400.
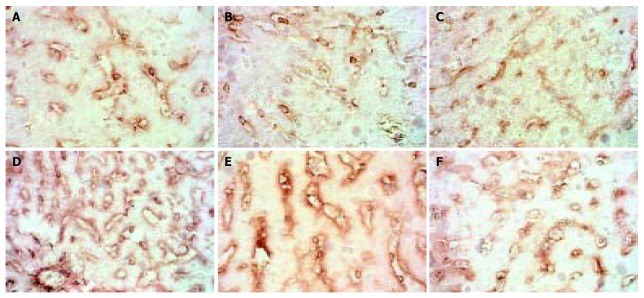
The expression of MMP-3 showed a similar pattern to that of MMP-2 but weaker, when modeling was stopped it recovered to normal level. (Figure 2).
Figure 2.
Dynamic changes of MMP-3. A: Normal rat, × 200 B: 4 d group, × 200 C: 2 wk group, × 200 D: 4 wk group, × 200. E: 9 wk group, × 200 F: 11 wk group, × 200.
The situation of MMP-9 was different from that of the two above. MMP-9 decreased in the 4 d and 2 wk groups, but in the 4 wk group and the following MMP-P showed higher expression than that of the normal rats with no significance. (Figure 3).
Figure 3.

Dynamic changes of MMP-9. A: Normal rat, × 400 B: 4 d group, × 400 C: 2 wk group, × 400 D: 4 wk group, × 400 E: 9 wk group, × 400 F: 11 wk group, × 400.
The expression of MMP-13 showed reduction in the 4 d and 2 wk groups, but recovered in the 4 wk group and then sustained, when modeling was stopped it showed higher expression than normal. (Figure 4).
Figure 4.

Dynamic changes of MMP-13. A: Normal rat, × 200 B: 4 d group, × 200 C: 2 wk group, × 200 D: 4 wk group, × 200 E: 9 wk group, × 200 F: 11 wk group, × 200.
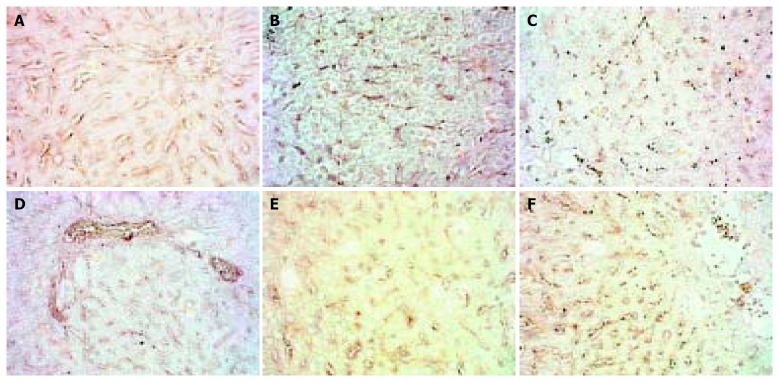
The expression of TIMP-1 in the 4 d and 2 wk groups showed reduction, after then it gave distinct higher expression and did not recover to normal level even after modeling was stopped for 2 wk. (Figure 5).
Figure 5.
Dynamic changes of TIMP-1. A: Normal rat, × 400 B: 4 d group, × 400 C: 2 wk group, × 400 D: 4 wk group, × 400 E: 9 wk group, × 400 F: 11 wk group, × 400.
The expression of TIMP-2 showed different type from that of TIMP-1. In normal rats it had a high level with some hepatocytes expressing, but as soon as modeling began it fell rapidly, then gradually ascended as modeling went on, but after modeling was stopped for 2 wk it was still lower than normal. (Figure 6).
Figure 6.
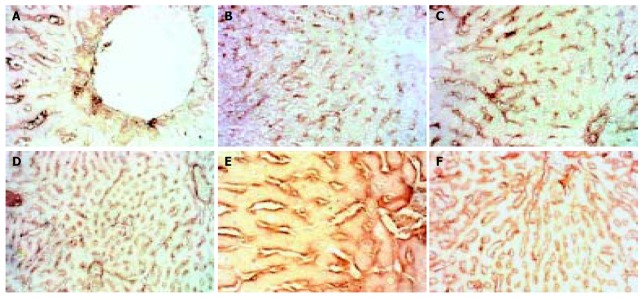
Dynamic changes of TIMMP-2. A: Normal rat, × 200 B: 4 d group, × 100 C: 2 wk group, × 200 D: 4 wk group, × 100 E: 9 wk group, × 400 F: 11 wk group, × 100.
We failed in this model to find any activities of MMP-2 and MMP-9 with zymography.
Western blotting tests showed that in the 4 wk and 9 wk groups there were strong expressions of TIMP-1, but in other groups there were not. As to TIMP-2 we did not find its expression. (Figure 7, Figure 8).
Figure 7.
Expression of TIMP-1 in the 4 wk and 9 wk groups.
Figure 8.
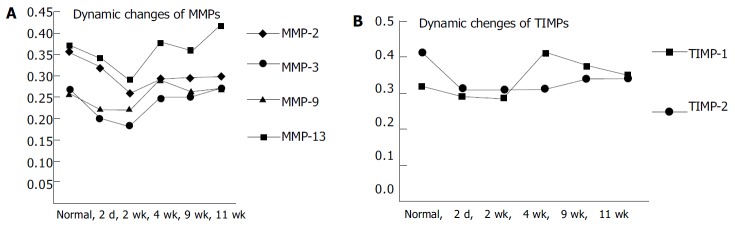
Dynamic changes of MMPs 2, 3, 9, 13 and TIMPs 1, 2 A: Dynamic changes of MMPs 2, 3, 9, 13. B: Dynamic changes of TIMPs 1, 2.
DISCUSSION
China’s prosperity is accompanied with cases of alcoholic liver disease growing day by day. Alcohol can stimulate hepatic stellate cells (HSCs) to proliferate and produce various fibrosis cytokines, which in turn result in hepatic fibrosis in the end. Abstinence itself can not reverse hepatic fibrosis, and more over, temperance is usually very difficult. So it is important to probe into the mechanism of hepatic fibrosis reversion in ALD research. Most of the recent reports[1-11] reveal that the formation and prognosis of hepatic fibrosis were all related with MMPs/TIMPs, and this viewpoint has been accepted as a common sense. So we probed into the important factors among MMPs/TIMPs, and respective discussions were as follows.
Matrix Metallopretinase-2 (MMP-2) is an important factor in the metabolism of type IV collagen in sinusoid basement. It has a peculiar activation course taking place on the membrane and forms a triple complex bridged with MT1-MMP by TIMP-2. Deactivation could be triggered by more competitive TIMP-2 to conjugate with MMP-2 to restrain the formation of triple complex. Because of local expression of TIMP-2 it is hypothesized that MMP-2 could not be activated on a long trip, and it is only transient and local. Activated MMP-2 could decompose type IV collagen in the basement, resulting in broken micro circumstances of HSC and promoting HSC to activate, proliferate and migrate. Activated MMP-2 also could play an important role in reversion of hepatic fibrosis, metastasis of tumors, MOF induced by endotoxin, apoptosis of HSC, etc[12-26]. MMP-2 is mainly composed and secreted by HSC in liver. Activation induced by various pathogens might need different time, and strong stimulation would result in rapid activation[27]. It was reported that[28] during reversion period activated MMP-2 was in favor of fibrosis reversion, so it was thought that MMP-2 played another role at this time because activated MMP-2 could activate HSC and promote fibrosis[29,30].
The results of our experiment were different from those of others. MMP-2 showed moderate expression in SD rat liver slides along the sinus in normal, began to fall during modeling, instead of rising up as reported, meanwhile zymograghy tests did not show any activity of it. So we thought that it was peculiar that lowly expressed MMP-2 could also break down type IV collagen in sinus basement markedly. Capillarization of this model was not typical because of lower contents of type IV collagen than normal. This might imply lowly expressed MMP-2 not only decomposed parts of type IV collagen in sinus basement but also obstructed its accumulation, resulting in atypical capillarization in the end. It could not achieve success unless TIMP-2 markedly decreased its expression so as to alleviate MMP-2, and our experiment showed an authentic result just like this. This phenomenon may exclude the possibility that only high activity of MMP-2 could result in breakage of sinus basement and progression of fibrosis, and it also may imply that MMP-2 is not the most important factor imposed on the metabolism of type IV collagen in sinus basement. Our results also showed that the cooperation of MMP-2/TIMP-2 could make the contents of type IV collagen in sinus basement become normal after the modeling was stopped for 2 wk. This was not consistent with other scholar’s report[28] that apoptosis of HSC could decrease the expression of TIMP-2 and promote the activity of MMP-2. The differences between species, modeling, stages and types of pathology may be related with these diversities. The fast and accurate adjusting mechanism of MMP-2’s function influenced heavily by TIMP-2, rather than TIMP-1, might be a commonfeature of different models.
MMP-3 also called stromelysin 1, originates from almost the same cells as MMP-1 but does not show synchronization in expression. MMP-3 has a wide range of substrates but important one may be laminin. it can also activate MMP-1, but there are not many reports about it. It has been reported that the measurement of serum MMP-3 was of little use for assessing fibrolysis in chronically diseased livers. However, because of the distinct pathological changes in sinusoid and peri-sinusoid in alcoholic liver disease, the value of MMP-3 rose to some extent. We found that accumulation of laminin in basement was quite prominent during capillarization. It was evident that the sustained low expression during modeling of MMP-3 would be in favor of laminin’s accumulation. The accumulated laminin in basement was still high after the modeling was stopped for 2 wk in spite of the normal expression of MMP-3, which might imply not only that the strong inhibiting effect of TIMP-1 on MMP-3 could result in delayed fashion of metabolism of laminin compared to that of type IV collagen, but also that sinusoid pathological changes were not entirely modulated by MMP-2/TIMP-2, and this is important.
MMP-9 is also called gelatinase B, the majority of which is secreted by kupffer cells in liver. MMP-9 could also decompose type IV collagen in basement, and this might happen a little earlier than that of MMP-2, implying MMP-9 might take part in the early events of fibrosis. In this model, however, MMP-9 had lower expression than MMP-2, and continued after the modeling began. It could be thought that MMP-9 did not play role in this model entirely. It was reported[31] that widespread capillarization in cirrhosis suggested no MMP-2 activity but low MMP-9 activity, implying potent importance of MMP-9 in reversion of fibrosis. In this model in the 4 wk and 11 wk groups MMP-9 showed little higher expression than normal but still lower than that of MMP-2, actually it might not affect the accumulation of type IV collagen, so it was not an important factor. This result did not show any consistency with other reports[29,30].
MMP-1, an interstitial collagenase (in rats it is MMP-13) is the most important enzyme secreted by kupffer cells and HSC in liver, and after secretion it is activated and the course may be related to plasminogen activator system. MMP-1/MMP-13 could be deactivated by forming a complex with TIMP-1 at a ratio 1:1. It was reported that kupffer cells in liver were one of the main originations of MMPs, including MMP-13. It was also reported recently that[32] the rise of activity of MMP-13 would significantly improve cirrhosis, which could be realized by stimulating kupffer cells. This implied another way to reverse fibrosis. The activity of MMP-13 was also needed to ensure normal metabolism of hepatocytes[33-35]. We found in our experiment that MMP-13 expressed themselves moderately mainly along sinusoid in normal rats which would support such a kind of viewpoints. MMP-13 decreased to some extent when the modeling began, in the 2 wk group some hepatocytes and kupffer cells showed active expression of MMP-13, then the expression along sinusoid again showed an ascending trend. In the 4 wk group when fibrosis was most severe the expression of MMP-13 was almost the same as normal, meaning that MMP-13 itself could not affect the course of peri-sinusoidal fibrosis. In the reversion phase the expression of MMP-13 was significantly higher than normal and the active stains implied that kupffer cells might restart their expression, which would be helpful to fibrosis reversion, but the accumulation of type I collagen was not affected and became more severe with the modeling, evidently the inhibiting effect of TIMP-1 on MMP-13 should be the cause. The higher expression of MMP-13 in the reversion phase actually could not help reverse peri-sinusoidal fibrosis, demonstrating once more that TIMP-1 was the most important factor during hepatic fibrosis.
TIMP-1 is produced by kupffer cells, HSC and myofibroblasts in liver, but most of it is mainly produced by activated HSC, It could be induced by TGF beta, interferon or pentoxifylline. TIMP-1 could inhibit most of MMPs except MMP-14 and MMP-19 by integrating them at a ratio 1:1 to form a complex. Injured liver tissues showed more expression of TIMP-1, and in turn, this would help interstitial fibrils to accumulate[33-36]. Serum TIMP-1 had a close relation to hepatic pathology, serum TIMP-1 could forecast hepatic pathological changes and prognosis, and could also be used as an index to evaluate the treatment. The transgenic model lacking TIMP-1 gene and anti-oligonucleotide treatment model aiming at TIMP-1 were established to find that without injury TIMP-1 itself could not result in fibrosis, but if there were injuries TIMP-1 would significantly accelerate the course in which fibrosis was formed. This might imply omni directional and all target treatment for hepatic fibrosis should be considered, nevertheless targeted and usual treatment still took TIMP-1 as a comprehensive index for evaluation[37-41]. Overexpression of TIMP-1 could keep hepatic fibrosis by holding back the apoptosis of HSC, otherwise the down-regulation of TIMP-1 would promote MMPs to be activated, and in turn, fibrils decomposed, HSC apoptosis took place and fibrosis reversed. All this emphasized the importance of TIMP-1.
We found in this model that at the early phase TIMP-1 showed a relatively low expression, in the 4 wk and 9 wk groups TIMP-1 showed evident expression mainly along sinusoid, which might be attributed to the widely activated HSCs. The over-expression of TIMP-1 would inhibit MMP-13 and MMP-3, resulting in accumulation of type I collagen and laminin, which in turn would be in favor of the formation of capillarization and peri-fibrosis. This discovery supported the importance of TIMP-1. It was also underpinned by Western blotting tests which showed TIMP-1 expressed itself most evidently in the 4 wk and 9 wk groups. In this model after the stimulus was ceased for 2 wk TIMP-1 still showed high expression, which might imply that activated HSCs were difficult to start spontaneous apoptosis, and in turn, TIMP-1 would not cease its expression by itself, which again showed it was a key step to reduce the expression of TIMP-1 in order to reverse fibrosis. Our results demonstrated that even if MMP-13 expressed highly it still would be inhibited by TIMP-1 to result in accumulation of type I collagen. It was reasonable that normal-level MMP-3 was inhabited by over-expressed TIMP-1, resulting in high content of laminin in basement of sinusoid, and capillarization and peri-sunusoidal fibrosis were not drastically improved. So this should be attributed to the high expression of TIMP-1.
The ability of TIMP-2 to integrate MMP-2 is 7-9 fold higher than that of TIMP-1, so if it is true it is the most important factor imposed on the metabolism of type IV collagen in basement of sinus. MMP-2 itself could not affect the course of capillarization as found in this experiment. We also found in normal SD rat livers that TIMP-2 was highly expressed in hepatocytes and kupffer cells, but transformed to HSCs with few active stains along sinusoid as the modeling began, which might imply activated HSCs would be the main origin of TIMP-2. During the modeling decreased expression of TIMP-2 might weaken the inhibition on MMP-2 and result in the decomposition of type IV collagen in basement and could not accumulate to help form typical capillarization. This was a typical feature of our model. After the modeling was stopped for 2 wk, the expression of TIMP-2 ascended but was little lower than normal, and the content of type IV collagen returned to normal level, which implied the gradually rising TIMP-2 could show its suppressing function on MMP-2, so that type IV collagen was protected and became normal. This demonstrated the significant effect of TIMP-2 and MMP-2 on the metabolism of type IV collagen in basement. However, TIMP-2 was apparently the more important.
In conclusion, the pathological changes of ALD model with fibrosis grades I-II can be controlled by TIMPs, especially the expression of TIMP-1 and TIMP-2 has significant effects on the prognosis of capillarization and peri-sinusoidal fibrosis, and MMPs are always dominated by TIMP-1 and TIMP-2. So the fatal step to reverse fibrosis might be to block or modulate the expression of TIMP-1 and TIMP-2.
ACKNOWLEDGEMENTS
The data of this article were obtained and analyzed with the help of Zhi-Gao Jin Professor and Ms. Hong Cai working in Academy of TCM. Many thanks to them!
Footnotes
Edited by Wang XL and Hu DK Proofread by Xu FM
References
- 1.Reeves HL, Friedman SL. Activation of hepatic stellate cells--a key issue in liver fibrosis. Front Biosci. 2002;7:d808–d826. doi: 10.2741/reeves. [DOI] [PubMed] [Google Scholar]
- 2.Smart DE, Vincent KJ, Arthur MJ, Eickelberg O, Castellazzi M, Mann J, Mann DA. JunD regulates transcription of the tissue inhibitor of metalloproteinases-1 and interleukin-6 genes in activated hepatic stellate cells. J Biol Chem. 2001;276:24414–24421. doi: 10.1074/jbc.M101840200. [DOI] [PubMed] [Google Scholar]
- 3.Boeker KH, Haberkorn CI, Michels D, Flemming P, Manns MP, Lichtinghagen R. Diagnostic potential of circulating TIMP-1 and MMP-2 as markers of liver fibrosis in patients with chronic hepatitis C. Clin Chim Acta. 2002;316:71–81. doi: 10.1016/s0009-8981(01)00730-6. [DOI] [PubMed] [Google Scholar]
- 4.Vaillant B, Chiaramonte MG, Cheever AW, Soloway PD, Wynn TA. Regulation of hepatic fibrosis and extracellular matrix genes by the th response: new insight into the role of tissue inhibitors of matrix metalloproteinases. J Immunol. 2001;167:7017–7026. doi: 10.4049/jimmunol.167.12.7017. [DOI] [PubMed] [Google Scholar]
- 5.Watanabe T, Niioka M, Ishikawa A, Hozawa S, Arai M, Maruyama K, Okada A, Okazaki I. Dynamic change of cells expressing MMP-2 mRNA and MT1-MMP mRNA in the recovery from liver fibrosis in the rat. J Hepatol. 2001;35:465–473. doi: 10.1016/s0168-8278(01)00177-5. [DOI] [PubMed] [Google Scholar]
- 6.Lichtinghagen R, Michels D, Haberkorn CI, Arndt B, Bahr M, Flemming P, Manns MP, Boeker KH. Matrix metalloproteinase (MMP)-2, MMP-7, and tissue inhibitor of metalloproteinase-1 are closely related to the fibroproliferative process in the liver during chronic hepatitis C. J Hepatol. 2001;34:239–247. doi: 10.1016/s0168-8278(00)00037-4. [DOI] [PubMed] [Google Scholar]
- 7.Okazaki I, Watanabe T, Hozawa S, Niioka M, Arai M, Maruyama K. Reversibility of hepatic fibrosis: from the first report of collagenase in the liver to the possibility of gene therapy for recovery. Keio J Med. 2001;50:58–65. doi: 10.2302/kjm.50.58. [DOI] [PubMed] [Google Scholar]
- 8.Ninomiya T, Yoon S, Nagano H, Kumon Y, Seo Y, Kasuga M, Yano Y, Nakaji M, Hayashi Y. Significance of serum matrix metalloproteinases and their inhibitors on the antifibrogenetic effect of interferon-alfa in chronic hepatitis C patients. Intervirology. 2001;44:227–231. doi: 10.1159/000050052. [DOI] [PubMed] [Google Scholar]
- 9.Williams EJ, Benyon RC, Trim N, Hadwin R, Grove BH, Arthur MJ, Unemori EN, Iredale JP. Relaxin inhibits effective collagen deposition by cultured hepatic stellate cells and decreases rat liver fibrosis in vivo. Gut. 2001;49:577–583. doi: 10.1136/gut.49.4.577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Benyon RC, Arthur MJ. Extracellular matrix degradation and the role of hepatic stellate cells. Semin Liver Dis. 2001;21:373–384. doi: 10.1055/s-2001-17552. [DOI] [PubMed] [Google Scholar]
- 11.Murphy FR, Issa R, Zhou X, Ratnarajah S, Nagase H, Arthur MJ, Benyon C, Iredale JP. Inhibition of apoptosis of activated hepatic stellate cells by tissue inhibitor of metalloproteinase-1 is mediated via effects on matrix metalloproteinase inhibition: implications for reversibility of liver fibrosis. J Biol Chem. 2002;277:11069–11076. doi: 10.1074/jbc.M111490200. [DOI] [PubMed] [Google Scholar]
- 12.Ueberham E, Löw R, Ueberham U, Schönig K, Bujard H, Gebhardt R. Conditional tetracycline-regulated expression of TGF-beta1 in liver of transgenic mice leads to reversible intermediary fibrosis. Hepatology. 2003;37:1067–1078. doi: 10.1053/jhep.2003.50196. [DOI] [PubMed] [Google Scholar]
- 13.Le Pabic H, Bonnier D, Wewer UM, Coutand A, Musso O, Baffet G, Clément B, Théret N. ADAM12 in human liver cancers: TGF-beta-regulated expression in stellate cells is associated with matrix remodeling. Hepatology. 2003;37:1056–1066. doi: 10.1053/jhep.2003.50205. [DOI] [PubMed] [Google Scholar]
- 14.Yang C, Zeisberg M, Mosterman B, Sudhakar A, Yerramalla U, Holthaus K, Xu L, Eng F, Afdhal N, Kalluri R. Liver fibrosis: insights into migration of hepatic stellate cells in response to extracellular matrix and growth factors. Gastroenterology. 2003;124:147–159. doi: 10.1053/gast.2003.50012. [DOI] [PubMed] [Google Scholar]
- 15.Gardi C, Arezzini B, Fortino V, Comporti M. Effect of free iron on collagen synthesis, cell proliferation and MMP-2 expression in rat hepatic stellate cells. Biochem Pharmacol. 2002;64:1139–1145. doi: 10.1016/s0006-2952(02)01257-1. [DOI] [PubMed] [Google Scholar]
- 16.Kobayashi H, Li ZX, Yamataka A, Lane GJ, Miyano T. Clinical evaluation of serum levels of matrix metalloproteinases and tissue inhibitors of metalloproteinases as predictors of progressive fibrosis in postoperative biliary atresia patients. J Pediatr Surg. 2002;37:1030–1033. doi: 10.1053/jpsu.2002.33836. [DOI] [PubMed] [Google Scholar]
- 17.Giannelli G, Bergamini C, Marinosci F, Fransvea E, Quaranta M, Lupo L, Schiraldi O, Antonaci S. Clinical role of MMP-2/TIMP-2 imbalance in hepatocellular carcinoma. Int J Cancer. 2002;97:425–431. doi: 10.1002/ijc.1635. [DOI] [PubMed] [Google Scholar]
- 18.Sawada S, Murakami K, Murata J, Tsukada K, Saiki I. Accumulation of extracellular matrix in the liver induces high metastatic potential of hepatocellular carcinoma to the lung. Int J Oncol. 2001;19:65–70. [PubMed] [Google Scholar]
- 19.Lee HS, Huang GT, Miau LH, Chiou LL, Chen CH, Sheu JC. Expression of matrix metalloproteinases in spontaneous regression of liver fibrosis. Hepatogastroenterology. 2001;48:1114–1117. [PubMed] [Google Scholar]
- 20.Chen PS, Zhai WR, Zhou XM, Zhang JS, Zhang Y, Ling YQ. [Effects of different causes on the expression of matrix metalloproteinase 2 in hepatic stellate cells] Zhonghua Ganzangbing Zazhi. 2002;10:279. [PubMed] [Google Scholar]
- 21.Preaux AM, D'ortho MP, Bralet MP, Laperche Y, Mavier P. Apoptosis of human hepatic myofibroblasts promotes activation of matrix metalloproteinase-2. Hepatology. 2002;36:615–622. doi: 10.1053/jhep.2002.35279. [DOI] [PubMed] [Google Scholar]
- 22.Lu X, Wang B, Xie Y, Liu C, Fu B. [Dynamic change and expression of matrix metalloproteinase-2, -9 in alcoholic liver disease in rats] Zhonghua Ganzangbing Zazhi. 2001;9:268–270. [PubMed] [Google Scholar]
- 23.Xie YH, Li L, Wang LE, Liu CR, Fu BY. Matrix metalloprotinase-2, -9 and hepatic fibrosis. Chin J Med. 2002;82:1172. [Google Scholar]
- 24.Dudás J, Kovalszky I, Gallai M, Nagy JO, Schaff Z, Knittel T, Mehde M, Neubauer K, Szalay F, Ramadori G. Expression of decorin, transforming growth factor-beta 1, tissue inhibitor metalloproteinase 1 and 2, and type IV collagenases in chronic hepatitis. Am J Clin Pathol. 2001;115:725–735. doi: 10.1309/J8CD-E9C8-X4NG-GTVG. [DOI] [PubMed] [Google Scholar]
- 25.Sakaida I, Hironaka K, Terai S, Okita K. Gadolinium chloride reverses dimethylnitrosamine (DMN)-induced rat liver fibrosis with increased matrix metalloproteinases (MMPs) of Kupffer cells. Life Sci. 2003;72:943–959. doi: 10.1016/s0024-3205(02)02342-1. [DOI] [PubMed] [Google Scholar]
- 26.Lee HS, Huang GT, Chen CH, Chiou LL, Lee CC, Yang PM, Chen DS, Sheu JC. Less reversal of liver fibrosis after prolonged carbon tetrachloride injection. Hepatogastroenterology. 2001;48:1312–1315. [PubMed] [Google Scholar]
- 27.Woo SW, Lee SH, Kang HC, Park EJ, Zhao YZ, Kim YC, Sohn DH. Butein suppresses myofibroblastic differentiation of rat hepatic stellate cells in primary culture. J Pharm Pharmacol. 2003;55:347–352. doi: 10.1211/002235702658. [DOI] [PubMed] [Google Scholar]
- 28.Gagliano N, Arosio B, Grizzi F, Masson S, Tagliabue J, Dioguardi N, Vergani C, Annoni G. Reduced collagenolytic activity of matrix metalloproteinases and development of liver fibrosis in the aging rat. Mech Ageing Dev. 2002;123:413–425. doi: 10.1016/s0047-6374(01)00398-0. [DOI] [PubMed] [Google Scholar]
- 29.Nie Q, Zhou Y, Xie Y. [Expression and significance of tissue inhibitors of metallproteinases-1 and -2 in serum and liver tissue of patients with liver cirrhosis] Zhonghua Yixue Zazhi. 2001;81:805–807. [PubMed] [Google Scholar]
- 30.Nie QH, Cheng YQ, Xie YM, Zhou YX, Bai XG, Cao YZ. Methodologic research on TIMP-1, TIMP-2 detection as a new diagnostic index for hepatic fibrosis and its significance. World J Gastroenterol. 2002;8:282–287. doi: 10.3748/wjg.v8.i2.282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mastroianni CM, Liuzzi GM, D'Ettorre G, Lichtner M, Forcina G, Di Campli NF, Riccio P, Vullo V. Matrix metalloproteinase-9 and tissue inhibitors of matrix metalloproteinase-1 in plasma of patients co-infected with HCV and HIV. HIV Clin Trials. 2002;3:310–315. doi: 10.1310/U9LJ-MFF9-ARE1-257H. [DOI] [PubMed] [Google Scholar]
- 32.Flisiak R, Maxwell P, Prokopowicz D, Timms PM, Panasiuk A. Plasma tissue inhibitor of metalloproteinases-1 and transforming growth factor beta 1--possible non-invasive biomarkers of hepatic fibrosis in patients with chronic B and C hepatitis. Hepatogastroenterology. 2002;49:1369–1372. [PubMed] [Google Scholar]
- 33.Yoshiji H, Kuriyama S, Yoshii J, Ikenaka Y, Noguchi R, Nakatani T, Tsujinoue H, Yanase K, Namisaki T, Imazu H, et al. Tissue inhibitor of metalloproteinases-1 attenuates spontaneous liver fibrosis resolution in the transgenic mouse. Hepatology. 2002;36:850–860. doi: 10.1053/jhep.2002.35625. [DOI] [PubMed] [Google Scholar]
- 34.Melgert BN, Olinga P, Van Der Laan JM, Weert B, Cho J, Schuppan D, Groothuis GM, Meijer DK, Poelstra K. Targeting dexamethasone to Kupffer cells: effects on liver inflammation and fibrosis in rats. Hepatology. 2001;34:719–728. doi: 10.1053/jhep.2001.27805. [DOI] [PubMed] [Google Scholar]
- 35.Raetsch C, Jia JD, Boigk G, Bauer M, Hahn EG, Riecken EO, Schuppan D. Pentoxifylline downregulates profibrogenic cytokines and procollagen I expression in rat secondary biliary fibrosis. Gut. 2002;50:241–247. doi: 10.1136/gut.50.2.241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.García L, Hernández I, Sandoval A, Salazar A, Garcia J, Vera J, Grijalva G, Muriel P, Margolin S, Armendariz-Borunda J. Pirfenidone effectively reverses experimental liver fibrosis. J Hepatol. 2002;37:797–805. doi: 10.1016/s0168-8278(02)00272-6. [DOI] [PubMed] [Google Scholar]
- 37.Bruck R, Genina O, Aeed H, Alexiev R, Nagler A, Avni Y, Pines M. Halofuginone to prevent and treat thioacetamide-induced liver fibrosis in rats. Hepatology. 2001;33:379–386. doi: 10.1053/jhep.2001.21408. [DOI] [PubMed] [Google Scholar]
- 38.Jia JD, Bauer M, Cho JJ, Ruehl M, Milani S, Boigk G, Riecken EO, Schuppan D. Antifibrotic effect of silymarin in rat secondary biliary fibrosis is mediated by downregulation of procollagen alpha1(I) and TIMP-1. J Hepatol. 2001;35:392–398. doi: 10.1016/s0168-8278(01)00148-9. [DOI] [PubMed] [Google Scholar]
- 39.Dubuisson L, Desmoulière A, Decourt B, Evadé L, Bedin C, Boussarie L, Barrier L, Vidaud M, Rosenbaum J. Inhibition of rat liver fibrogenesis through noradrenergic antagonism. Hepatology. 2002;35:325–331. doi: 10.1053/jhep.2002.31166. [DOI] [PubMed] [Google Scholar]
- 40.Wang A, Yang X, Wang W, Zuo F, Wang Q, He F. [Effect of recombinant human augmenter of liver regeneration on gene expression of tissue inhibitor of metalloproteinase-1 in rat with experimental liver fibrosis] Zhonghua Yixue Zazhi. 2002;82:610–612. [PubMed] [Google Scholar]
- 41.Spira G, Mawasi N, Paizi M, Anbinder N, Genina O, Alexiev R, Pines M. Halofuginone, a collagen type I inhibitor improves liver regeneration in cirrhotic rats. J Hepatol. 2002;37:331–339. doi: 10.1016/s0168-8278(02)00164-2. [DOI] [PubMed] [Google Scholar]