

Abstract
AIM: To study the effect of integration of tandem aroG-pheA genes into the tyrA locus of Corynebacterium glutamicum (C. glutamicum) on the production of L-phenylalanine.
METHODS: By nitrosoguanidine mutagenesis, five p-fluorophenylalanine (FP)-resistant mutants of C.glutamicum FP were selected. The tyrA gene encoding prephenate dehydrogenase (PDH) of C.glutamicum was amplified by polymerase chain reaction (PCR) and cloned on the plasmid pPR. Kanamycin resistance gene (Km) and the PBF-aroG-pheA-T (GA) fragment of pGA were inserted into tyrA gene to form targeting vectors pTK and pTGAK, respectively. Then, they were transformed into C.glutamicum FP respectively by electroporation. Cultures were screened by a medium containing kanamycin and detected by PCR and phenotype analysis. The transformed strains were used for L-phenylalanine fermentation and enzyme assays.
RESULTS: Engineering strains of C.glutamicum (Tyr-) were obtained. Compared with the original strain, the transformed strain C. glutamicum GAK was observed to have the highest elevation of L-phenylalanine production by a 1.71-fold, and 2.9-, 3.36-, and 3.0-fold in enzyme activities of chorismate mutase, prephenate dehydratase and 3-deoxy-D-arabinoheptulosonate-7-phosphate synthase, respectively.
CONCLUSION: Integration of tandem aroG-pheA genes into tyrA locus of C. glutamicum chromosome can disrupt tyrA gene and increase the yield of L-phenylalanine production.
INTRODUCTION
L-phenylalanine is one of the essential amino acids for humans. Its applications range from feed to food,and pharmaceutical products. such as p-fluorophenylalanine are used as anti-tumour drugs. At present, L-phenylalanine biosynthesis genes have been well characterized and the enzymology of L-phenylalanine biosynthesis has been extensively investigated[1-10]. In bacteria, the biosynthesis of aromatic amino acids starts from condensation reaction of central carbon intermediates such as phosphoenol pyruvate (PEP) and erythrose-4-phosphate (E4P) to form 3-deoxy-D-arabino-heptulosonate-7-phosphate (DAHP), which is catalyzed by DAHP synthase (DS). DAHP is then converted to chorismate, the branch point of aromatic amino acid biosynthesis. L-phenylalanine is synthesized from chorismate by three continuous steps catalyzed by chorismate mutase (CM), prephenate dehydratase (PD) and aromatic-amino-acid transaminase (AT). aroG[1-5] and pheA[6-10] are two key genes in L-phenylalanine biosynthesis. In E. coli, they encode the 3-deoxy-D-arabinoheptulosonate-7-phosphate synthase (DS) and chorismate mutase/prephenate dehydratase (CM/PD), respectively. However, in Corynebacterium glutamicum (C. glutamicum), CM and PD are encoded by two different genes pheB and pheA[10]. The over-expression of aroG and pheA can increase the yield of L-phenylalanine biosynthesis in E. coli or in Corynebacteria by shuttle vectors[11-17]. tyrA encoding prephenate dehydrogenase(PDH) is a key gene in L-tyrosine biosynthesis branch pathway[18]. DS and CM of C. glutamicum can be synergistically inhibited by L-tyrosine and L-phenylalanine. Every step from DAHP to chorismate is repressed weakly by L-tyrosine. In addition, PD is strongly inhibited by L-phenylalanine and L-tryptophan, L-tyrosine stimulates PD activity and restores the enzyme activity inhibition by L-phenylalanine and L-tryptophan[19]. These regulations seem to result in balanced synthesis of L-tyrosine and L-phenylalanine (Figure 1). Therefore, elimination of feedback inhibition of L-phenylalanine on the key enzymes and deletion of tyrA gene can improve the yield of L-phenylalanine biosynthesis. There are successful examples by using homologous recombination technology to introduce genes into chromosome[20-23].
Figure 1.
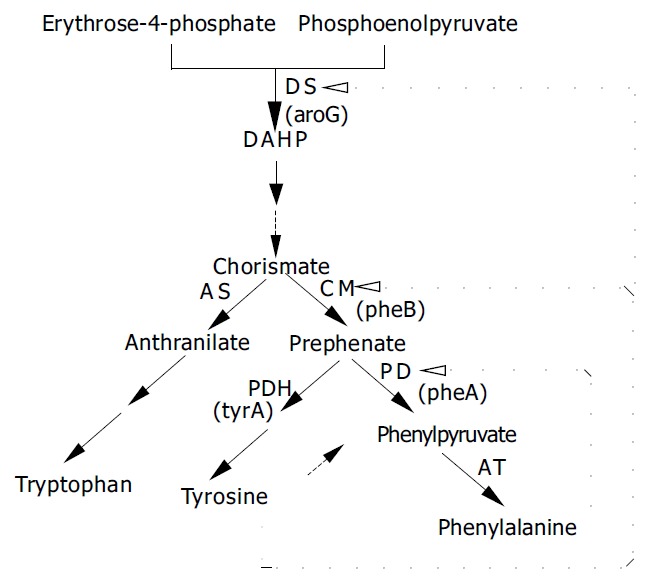
Pathway and primary regulations of phenylalanine biosynthesis in C. glutamicum ←---indicates feedback inhibition, ---→indicates activation.
In this study, we used nitrosoguanidine(NTG) to treat C. glutamicum. Mutants resistant to FP were isolated in a minimal medium plate with FP and named C.glutamicum FP. The tandem aroG-pheA[15] genes obtained from E. coli resistant to FP were inserted into the chromosome of C.glutamicum FP by a double-exchange homologous recombination between the fragments of tyrA gene on targeting vectors and the tyrA gene on the chromosome of C.glutamicum FP. Enzyme activities and L-phenylalanine biosynthesis in the engineered strains of C.glutamicum KM and C.glutamicum GAK were analyzed.
MATERIALS AND METHODS
Bacterial strains, media, and culture conditions
The bacterial strains and plasmids used in this study are listed in Table 1. Minimal medium[15] (MM) and NB (LB added with 10 g/L beef extract) were used for culture C. glutamicum. The medium for C. glutamicum fermentation was reported previously[16]. LB was used for growth of E. coli. When required, appropriate antibiotics were added to the suitable concentrations. Solid media were made by the addition of agar at the concentration of 15 g/L.
Table 1.
Strains and plasmids
| Strains or plasmids | Relevant characteristics | Sources or references |
| E.coli JM110 | Strain for vectors construction | Our laboratory |
| C.glutamicum | Nxr, original strainmutagenesis | ATCC |
| C.glutamicum FP | FPr, from C.glutamicum ATCC13032 | This work |
| C.glutamicum KM | C. glutamicum FP inserted with Km gene | This work |
| C.glutamicum GAK | C. glutamicum FP inserted with GAK | This work |
| pBluscript SK (pSK) | Apr, vector for gene cloning | Our laboratory |
| pPR | Apr, pBR322 derivative, suicidal vector | Our laboratory |
| pGA | Kmr, carrying PBF-aroG-pheA-T fragment | Ref.15 |
| pT | pPR carrying tyrA gene | This work |
| pTK | pT carrying Km gene(tyrA::Km) | This work |
| pSKGAK | pSK carrying GAK fragment | This work |
| pTGAK | pT carrying GAK fragment (tyrA::GAK) | This work |
FPr: Resistance to p-fluorophenylalanine; Nxr: Resistance to nalidixic acid; Apr: Resistance to ampicillin; Kmr: Resistance to kanamycin; ATCC: American Type Culture Collection. PBF: Promoter from C.glutamicum; T: Terminator of gene32 in T4 phage. GAK: PBF-aroG-pheA-T-Km fragment.
Chemicals
T4 DNA ligase, restriction enzymes, gel extract kit and DNA blunting kit were obtained from Takara Co. NTG and the reagents used in the enzyme assays were purchased from Sigma Co. They were used according to the instructions of their manufacturers.
Isolation of L-phenylalanine analogue resistant mutants
C. glutamicum cells were grown in NB supplemented with 20 mg/L nalidixic acid until log phase, and collected by centrifugation, followed by two washes with phosphate buffered saline (pH6.0).The cells were then resuspended in 500 µL of 9 g/L NaCl containing 5 mg of NTG and kept on shaking at 30 °C for another 30 min. The cells were collected by centrifugation followed by two washes before suspension in 9 g/L NaCl. Appropriate aliquots of the suspension were spread on the minimal agar plate containing different concentrations of p-fluorophenylalanine (p-FP) for mutant screening.
Preparation and manipulation of DNA
Chromosomal DNA of C. glutamicum was extracted as described preciously[10]. Manipulation of plasmid DNA was performed as described by Sambrook et al[24].
Construction of recombinant plasmids
Primers of tyrA and Km were designed according to the sequences in GenBank (NC_003450 and AF012346). Primer P1: 5’-GGCTGCAGCAGGCCATCCTCTTCAGTGTTCA-3’ and primerP2:5’-CCCAAGCTTCGAGGGTTTCAGCGTGGATG-3’ were designed for amplifying tyrA gene carrying PstI and Hind III restriction enzyme sites, respectively. Primer P3: 5’-GCGGAATTCACCGGAATTGCCAGCTG-3’ and primer P4: 5’-GCGGAATTCTGCAGTTATCAGAAGAACTCGTCAAG-3’weredesigned for amplifying Km gene carrying EcoRI and PstI sites, respectively. The primer P5 for PCR detection was 5’-CCCAAGCTTAGGAATCGCCTGAGAATCATCA-3’. Construction of recombinant plasmids was performed as shown in Figure 2.
Figure 2.
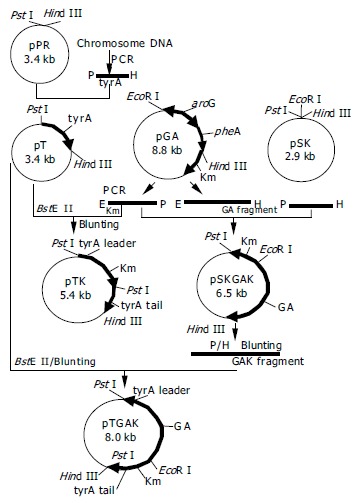
Construction of recombinant plasmids. E: EcoRI; P: PstI; H: Hind III; GA: PBF-aroG-pheA-T fragment; GAK: PBF-aroG-pheA-T-Km fragment.
Electroporation of linear DNA into C. glutamicum
Targeting vectors were digested by Hind III and the linear DNA was obtained by gel extraction. Then, they were denatured by 1 mol/L NaOH at 37 °C for 10 min, quickly placed on ice before the same volume of 1 mol/L HCl was added to neutralize the alkali. The method of Molenaar[25] was used to introduce denatured targeting vectors into C.glutamicum FP.
Selection and identification of the recombinant strain
After electroporation, C.glutamicum was incubated in 1mL NB medium containing 0.5 mol/L sucrose at 30 °C by shaking (100 r/min) for 1 h, then the culture was plated on selective NB medium containing 0.5 mol/L sucrose, 10 µg/mL nalidixic acid and 15 µg/mL kanamycin for 96 h. The strains on resistant plates were singled out and inoculated on NB with 100 µg/mL ampicillin, 15 µg/mL kanamycin and NB with 15 µg/mL kanamycin, respectively; Clones (Aps Kmr) were detected by PCR using primers pair P3 and P5. PCR products were sequenced.
Detection of Tyr- phenotype
The strain cultures were washed three times with sterile physiological saline and plated on MM plates containing 2 g/L glucose and 1 mol/L L-tyrosine or other amino acids to detect the Tyr- auxotroph.
Enzymatic activities assay
Crude cell lysates used for enzymatic activity assays were prepared as described previously[15]. Total protein level was determined according to the method of Bradford[26]. DS activity was assayed as described previously[15]. CM activity was determined as described by Xia et al[27]. PD activity was assayed as preciously described[28].
Fermentation and analysis of phenylalanine
Transformants of C. glutamicum were obtained by PCR and phenotype analysis. L-phenylalanine fermentation of them was carried out in the shaking flask and the yields of L-phenylalanine biosynthesis were determined as described by Zeng et al[15].
RESULTS
Isolation of L-phenylalanine analogue resistant mutants
Five mutants resistant to p-FP up to 3 mg/mL were picked out on MM plates after NTG mutagenesis. In those mutants, the feedback inhibition on the key enzymes in biosynthesis pathways was released from phenylalanine. The mutant with the highest yield of phenylalanine was named C.glutamicum FP and used as the recipient strain for electroporation (unpublished data). C.glutamicum FP was detected by PCR with primers P1 and P2. The sequence of PCR products was consistent with tyrA of C.glutamicum ATCC 13032.
Construction of recombinant plasmids
Km (950 bp) and tyrA (1067 bp) genes were amplified by PCR and they were verified by sequencing. pPR vector and tyrA gene were digested with PstI and Hind III and ligated to form pT. pT was digested with BstEII, and then blunted. Km gene and PBF-aroG-pheA-T fragment from pGA plasmids were subcloned into pSK vector, then digested with Pst I and Hind III to get GAK fragment (PBF-aroG-pheA-T-Km). This resulting fragment and the Km gene were inserted into the pT to form targeting vectors pTGAK and pTK, respectively. The detailed process refers to Figure 2.
Identification of transformants
Because primer P5 was located at the end of tyrA gene, it only existed on the chromosome of C.glutamicum, but not on the targeting vector. Only when the Km gene was inserted into the chromosome of C.glutamicum, could the product be obtained by PCR using the primer pair P3 and P5 (Figure 3). The expected PCR products of 1570 bp are shown in Lane 3 and Lane 5 of Figure 4. Since a PstI restriction site was introduced into Km gene in foreside (Figure 2), the obtained PCR products could be digested by PstI to produce two fragments whose sizes (950 bp and 620 bp) accorded to the sizes of Km gene and an expanded tyrA tail as shown in Lane 2 and Lane 4 of Figure 4. Sequencing of the 1570 bp fragment demonstrated it consisted of Km gene and a 620 bp fragment including tyrA tail (unpublished data). The strains obtained by PCR detection were named C. glutamicum KM (inserted with Km) and C. glutamicum GAK (inserted with GAK).
Figure 3.
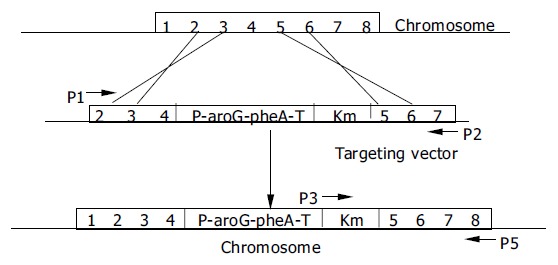
Insertion of PBF-aroG-pheA-T-Km fragment of pTGAK into chromosome tyrA locus of C. glutamicum by double-ex-change homologous recombination and detection of ligated DNA fragment Km-tyrA tail on chromosome by PCR ampli-fication with P3 and P5.
Figure 4.
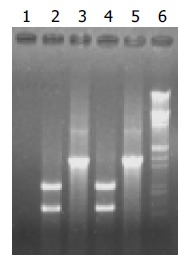
DNA detection of transformants by PCR amplifica-tion and PstI digested PCR products. Lane 1: control for PCR; lane 2: PstI digesting product of lane 3; lane 3: PCR detection of C. glutamicum KM; lane 4, PstI digesting product of lane 5; lane 5: PCR detection of C. glutamicum GAK; lane 6: Markers, EcoRI/Hind III digesting λ phage DNA.
Detection of Tyr- auxotroph
As shown in Figure 5, C.glutamicum GAK, in which tyrA was disrupted by inserting a GAK fragement, only grew on the MM with L-tyrosine, but not on MM and MM with L-tyrptophan or L-phenylalanine plates. This result indicated that C.glutamicum GAK was an auxotroph requiring L-tyrosine for growth. Here, we demonstrated that the pathway of tyrosine biosynthesis at steps from prephenate to 4-hydroxyphenlpyruvate was disrupted, and the function of prephenate dehydrogenase was inactivated successfully.
Figure 5.

Auxanogram detection of C. glutamicum (Tyr-). MM: C. glutamicum GAK on MM; Tyr: C.glutamicum GAK on MM + Tyr; Phe: C.glutamicum GAK on MM + Phe; Trp: C.glutamicum GAK on MM + Trp.
Enzyme assay
As shown in Figure 1, in the pathway of L-phenylalanine biosynthesis, there were three key enzymes, PD, CM and DS, encoded by pheA, pheB and aroG, respectively in C.glutamicum. The results of enzyme activity in C.glutamicum are listed in Table 2. The activity of the original strain was used as the control. Compared to the original strain, the relative activities of three enzymes (CM, PD and DS) in C.glutamicum FP were 1.73-, 2.37- and 1.93-fold higher and in C.glutamicum GAK were 2.9-, 3.36- and 3.0-fold higher respectively. The relative activities of these enzymes in C.glutamicum KM (Kmr FPr) were similar to those in C.glutamicum FP (FPr) respectively. The results indicated that the enzyme activities were increased differently by inhibiting the release of L-phenylalanine feedback using FPr selection and the integration of tandem genes aroG-pheA into chromosome of C.glutamicum. At the same time, it also proved that the homologous recombination was successful.
Table 2.
Relative activities of CM, PD and DS compared with the original strains
| Strains |
Enzymes |
||
| CM | PD | DS | |
| C. glutamicum | 1.0 | 1.0 | 1.0 |
| C. glutamicum FP | 1.73 | 2.37 | 1.93 |
| C. glutamicum KM | 1.74 | 2.30 | 1.95 |
| C. glutamicum GAK | 2.90 | 3.36 | 3.00 |
The enzyme activity of C.glutamicum was used as the standard, i.e. the relative activity of 1. CM: Chorismate mutase; PD: Prephenate dehydratase; DS: 3-deoxy-D-arabinoheptulosonate-7-phosphate synthase.
Measurement of phenylalanine yields
Recombinant strains were shaked in 25 mL of fermentation medium with 0.5 mol/L L-tyrosine in 250 mL flask at 30 °C for 72 h. The average yield of three measurements is shown in Table 3. From these data, we could find inhibition of the release of feedback contributed to increase the yield of L-phenylalanine obviously, the yield was increased 42%. On the other hand, interruption of the L-tyrosine branch pathway also increased 9% of the yield. Integration of the tandem genes aroG-pheA made 20% improvement. The total yield was increased 71%. The highest yield was added up to 3.97 g/L.
Table 3.
Yields of L-phenylalanine biosynthesis in flask fer-mentation (mean ± SD)
| Strains | C.glutamicum | C.glutamicum FP | C.glutamicum KM | C.glutamicum GAK |
| Yields (g/L) | 2.20 ± 0.15 | 3.12 ± 0.17 | 3.32 ± 0.12 | 3.76 ± 0.21 |
| Relative yields | 1 | 1.42 | 1.51 | 1.71 |
The phenylalanine yield of the original stain was used as the standard, i.e. the relative yield of 1.
DISCUSSION
The results of this study showed that interruption of the branch pathway of L-tyrosine biosynthesis could accumulate intermediates for L-phenylalanine synthesis and integration of the tandem aroG-pheA encoding DS, CM and PD from E. coli into the C.glutamicum chromosome was also favorable to L-phenylalanine synthesis due to the improvement of key enzyme activities in biosynthesis pathways. These two factors contributed to the increase of 30% yield in L-phenylalanine biosynthesis. By increasing the precursors for phenylalanine synthesis, interruption of the tyrosine branch pathway could decrease L-tyrosine synthesis, and inhibit L-phenylalanine synthesis because L-tyrosine could stimulate prephenate dehydratase activity and restore the enzyme activity inhibited by L-phenylalanine and L-tryptophan. Ours results also indicated that mutagenesis by NTG was still an effective method for screening phenylalanine-produing strains. This method could effectively to improve the amino acid production in this study. The strains integrated with Km gene kept their resistance to kanamycin after cultured in medium without antibiotics for 96 h (unpublished data). Compared to the strains carrying plasmids[15-17], strains integrated with heterologous genes had lower yields of L-phenylalanine but a higher stability. Further mutagenesis and application of strong promoters may be useful to increase the yield of L-phenylalanine of engineering strains. Further investigations are necessary to understand the mechanism of aromatic amino acid biosynthesis.
ACKNOWLEDGEMENTS
We thank professor Jiang Zhong, associate professor Han-Ying Yuan and De-Qiang Xu from Fudan University for their helps on this work.
Footnotes
Supported by the National Natural Science Foundation of China, No. 30070020
Edited by Kumar M and Wang XL Proofread by Xu FM
References
- 1.Davies WD, Pittard J, Davidson BE. Cloning of aroG, the gene coding for phospho-2-keto-3-deoxy-heptonate aldolase(phe), in Escherichia coli K-12, and subcloning of the aroG promoter and operator in a promoter-detecting plasmid. Gene. 1985;33:323–331. doi: 10.1016/0378-1119(85)90240-9. [DOI] [PubMed] [Google Scholar]
- 2.Kikuchi Y, Tsujimoto K, Kurahashi O. Mutational analysis of the feedback sites of phenylalanine-sensitive 3-deoxy-D-arabino-heptulosonate-7-phosphate synthase of Escherichia coli. Appl Environ Microbiol. 1997;63:761–762. doi: 10.1128/aem.63.2.761-762.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ger YM, Chen SL, Chiang HJ, Shiuan D. A single Ser-180 mutation desensitizes feedback inhibition of the phenylalanine-sensitive 3-deoxy-D-arabino-heptulosonate 7-phosphate (DAHP) synthetase in Escherichia coli. J Biochem. 1994;116:986–990. doi: 10.1093/oxfordjournals.jbchem.a124657. [DOI] [PubMed] [Google Scholar]
- 4.Jiang PH, Shi M, Qian ZK, Li NJ, Huang WD. Effect of F209S Mutation of Escherichia coli AroG on Resistance to Phenylalanine Feedback Inhibition. Shengwu Huaxue Yu Shengwu Wuli Xuebao (Shanghai) 2000;32:441–444. [PubMed] [Google Scholar]
- 5.Hu C, Jiang P, Xu J, Wu Y, Huang W. Mutation analysis of the feedback inhibition site of phenylalanine-sensitive 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase of Escherichia coli. J Basic Microbiol. 2003;43:399–406. doi: 10.1002/jobm.200310244. [DOI] [PubMed] [Google Scholar]
- 6.Zhang S, Wilson DB, Ganem B. Probing the catalytic mechanism of prephenate dehydratase by site-directed mutagenesis of the Escherichia coli P-protein dehydratase domain. Biochemistry. 2000;39:4722–4728. doi: 10.1021/bi9926680. [DOI] [PubMed] [Google Scholar]
- 7.Nelms J, Edwards RM, Warwick J, Fotheringham I. Novel mutations in the pheA gene of Escherichia coli K-12 which result in highly feedback inhibition-resistant variants of chorismate mutase/prephenate dehydratase. Appl Environ Microbiol. 1992;58:2592–2598. doi: 10.1128/aem.58.8.2592-2598.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gavini N, Davidson BE. pheAo mutants of Escherichia coli have a defective pheA attenuator. J Biol Chem. 1990;265:21532–21535. [PubMed] [Google Scholar]
- 9.Hsu SK, Lin LL, Lo HH, Hsu WH. Mutational analysis of feedback inhibition and catalytic sites of prephenate dehydratase from Corynebacterium glutamicum. Arch Microbiol. 2004;181:237–244. doi: 10.1007/s00203-004-0649-5. [DOI] [PubMed] [Google Scholar]
- 10.Chan MS, Hsu WH. Cloning of m-fluorophenylalanine-resistant gene and mutational analysis of feedback-resistant prephenate dehydratase from Corynebacterium glutamicum. Biochem Biophys Res Commun. 1996;219:537–542. doi: 10.1006/bbrc.1996.0269. [DOI] [PubMed] [Google Scholar]
- 11.Ozaki A, Katsumata R, Oka T, Furuya A. Functional expression of the genes of Escherichia coli in gram-positive Corynebacterium glutamicum. Mol Gen Genet. 1984;196:175–178. doi: 10.1007/BF00334113. [DOI] [PubMed] [Google Scholar]
- 12.Ikeda M, Ozaki A, Katsumata R. Phenylalanine production by metabolically engineered Corynebacterium glutamicum with the pheA gene of Escherichia coli. Appl Microbiol Biotechnol. 1993;39:318–323. doi: 10.1007/BF00192085. [DOI] [PubMed] [Google Scholar]
- 13.Jiang PH, Liu AM, Ge HP, Zhang YY, Fan CS, Huang WD. Cloning and Expression of aroG Gene of E. coli and Its Co-expression with pheA and tyrB Genes. Shengwu Huaxue Yu Shengwu Wuli Xuebao (Shanghai) 1998;30:593–596. [PubMed] [Google Scholar]
- 14.Eikmanns BJ, Kleinertz E, Liebl W, Sahm H. A family of Corynebacterium glutamicum/Escherichia coli shuttle vectors for cloning, controlled gene expression, and promoter probing. Gene. 1991;102:93–98. doi: 10.1016/0378-1119(91)90545-m. [DOI] [PubMed] [Google Scholar]
- 15.Zeng XB, Fan CS, Jiang PH, Chen YQ, Huang WD. Co-expression of heterologous genes pheA, aroG and tyrB for biosynthesis path-ways of L-phenylalanine. Chin J Biotechnol. 2000;16:671–674. [Google Scholar]
- 16.Wu YQ, Jiang PH, Fan CS, Wang JG, Shang L, Huang WD. Co-expression of five genes in E coli for L-phenylalanine in Brevibacterium flavum. World J Gastroenterol. 2003;9:342–346. doi: 10.3748/wjg.v9.i2.342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Fan C, Zeng X, Chai Y, Jiang P, Huang W. [Expression of genes aroG and pheA in phenylalanine biosynthesis] Weishengwu Xuebao. 1999;39:430–435. [PubMed] [Google Scholar]
- 18.http: //www ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=nucleoti de&val=23308765
- 19.Follettie MT, Sinskey AJ. Molecular cloning and nucleotide sequence of the Corynebacterium glutamicum pheA gene. J Bacteriol. 1986;167:695–702. doi: 10.1128/jb.167.2.695-702.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Schäfer A, Tauch A, Jäger W, Kalinowski J, Thierbach G, Pühler A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene. 1994;145:69–73. doi: 10.1016/0378-1119(94)90324-7. [DOI] [PubMed] [Google Scholar]
- 21.Fitzpatrick R, O'Donohue M, Joy J, Heery DM, Dunican LK. Construction and characterization of recA mutant strains of Corynebacterium glutamicum and Brevibacterium lactofermentum. Appl Microbiol Biotechnol. 1994;42:575–580. doi: 10.1007/BF00173923. [DOI] [PubMed] [Google Scholar]
- 22.Ikeda M, Katsumata R. A novel system with positive selection for the chromosomal integration of replicative plasmid DNA in Corynebacterium glutamicum. Microbiology. 1998;144(Pt 7):1863–1868. doi: 10.1099/00221287-144-7-1863. [DOI] [PubMed] [Google Scholar]
- 23.Kirchner O, Tauch A. Tools for genetic engineering in the amino acid-producing bacterium Corynebacterium glutamicum. J Biotechnol. 2003;104:287–299. doi: 10.1016/s0168-1656(03)00148-2. [DOI] [PubMed] [Google Scholar]
- 24.Sambrook J, Russell DW. Molecular cloning: a laboratory Manual. 3thed. New York: Cold Spring Harbor Laboratory Press; 2001. pp. 48–51. [Google Scholar]
- 25.van der Rest ME, Lange C, Molenaar D. A heat shock following electroporation induces highly efficient transformation of Corynebacterium glutamicum with xenogeneic plasmid DNA. Appl Microbiol Biotechnol. 1999;52:541–545. doi: 10.1007/s002530051557. [DOI] [PubMed] [Google Scholar]
- 26.Fong DS, Frederick AR, Blumenkranz MS, Walton DS. Exudative retinal detachment in X-linked retinoschisis. Ophthalmic Surg Lasers. 1998;29:332–335. [PubMed] [Google Scholar]
- 27.Xia T, Zhao G, Jensen RA. Loss of allosteric control but retention of the bifunctional catalytic competence of a fusion protein formed by excision of 260 base pairs from the 3' terminus of pheA from Erwinia herbicola. Appl Environ Microbiol. 1992;58:2792–2798. doi: 10.1128/aem.58.9.2792-2798.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zhang S, Wilson DB, Ganem B. Probing the catalytic mechanism of prephenate dehydratase by site-directed mutagenesis of the Escherichia coli P-protein dehydratase domain. Biochemistry. 2000;39:4722–4728. doi: 10.1021/bi9926680. [DOI] [PubMed] [Google Scholar]