

Abstract
Many Gram-negative bacterial species release outer membrane vesicles (OMVs) that interact with the host by delivering virulence factors. Here, we report for the first time that RNA is among the wide variety of bacterial components that are associated with OMVs. To characterize the RNA profiles of bacterial OMVs, we performed RNA deep sequencing analysis using OMV samples isolated from a wild type Vibrio cholerae O1 El Tor strain. The results showed that RNAs originating from intergenic regions were the most abundant. Our findings reveal a hitherto unrecognised feature of OMVs mimicking eukaryotic exosomes and highlight a need to evaluate the potential role of RNA-containing bacterial membrane vesicles in bacteria-host interactions.
Outer membrane vesicles (OMVs) are constantly being discharged from the surface of Gram-negative bacteria during growth. OMVs typically range between 50–200 nm in diameter and contain outer membrane proteins, LPS, phospholipids, and some periplasmic constituents1.
Studies of bacterial OMVs have revealed some similarities with vesicles known as exosomes that are secreted by most mammalian cell types. OMVs and exosomes have similar size ranges and both carry payloads of proteins, lipids, and genetic material enclosed in membrane-bound spherical structures. Both types can deliver functional molecules to distant extracellular compartments and tissues. Exosomes are involved in: horizontal transfer of genetic material2; transport of mRNAs and miRNAs from mast cells to recipient mast cells3; delivery of miRNAs from T cells to antigen presenting cells to modulate gene expression4 and delivery of viral miRNAs from infected cells to non-infected cells5. These capabilities collectively suggest that exosomes, by being natural carriers and intercellular transporters of RNA, are suitable candidates for delivery of exogenous gene interfering compounds, such as siRNA.
The function of bacterial OMVs to aid in transport of a variety of bacterial products- such as proteins, enzymes, and DNA is well described. Extensive studies, reviewed elsewhere1,6,7, have proven the importance of a vesicle-mediated transport system and clearly describe the ways in which different bacteria make use of this delivery system. Several virulence-associated factors of pathogenic bacteria can be detected in association with OMVs8,9,10. OMVs likely facilitate direct delivery of virulence factors to target host cells rather than to the external milieu where diffusion could lessen the functional impact of the molecules.
Their similarity to eukaryotic exosomes and the fact that OMVs act as transporters of various bacterial constituents prompted us to investigate if an “exosome-like” RNA delivery system by bacterial OMVs exists.
Results and Discussion
Detection of RNA associated with V. cholerae A1552 vesicles
OMVs from the wild type V. cholerae strain A1552 were isolated as described earlier11,12. The ultrastructural analysis of OMVs was performed using transmission electron microscopy (TEM) and cryo-electron tomography. As shown in Fig. 1A, we observed OMVs with diameters ranging from 25–200 nm by TEM. The 3D volume of the purified vesicles was analysed by cryo-electron tomography and interestingly revealed that larger vesicles comprised double membranes (Fig. 1B). Morphologically these OMVs may appear similar to bacterial minicells which are achromosomal particles derived from bacterial cells with mutations inactivating genes controlling normal bacterial cell division13. However, OMVs are released during normal growth of bacteria and the V. cholerae strain used has no aberration of the normal process of cell division. Furthermore, the OMVs are distinctly smaller in size in comparison with minicells.
Figure 1. Vibrio cholerae O1 El Tor A1552 membrane vesicles.
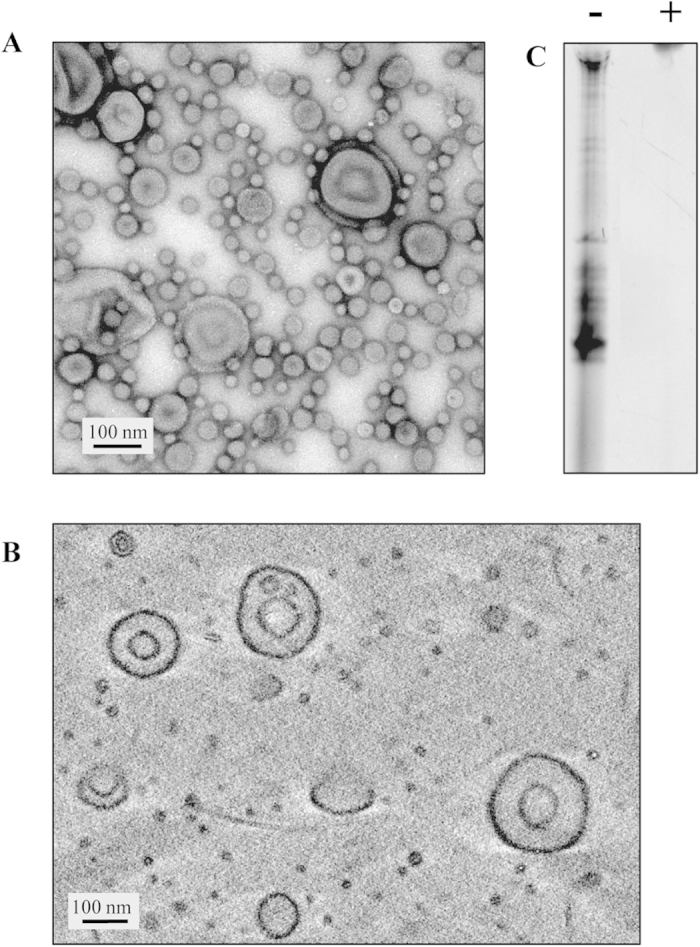
(A) Negative staining micrograph of purified vesicles before hot phenol treatment. Bar: 100 nm. (B) Cryo-electron tomogram of purified vesicles. The image shows a projection of 10 sections corresponding to 6 nm in the centre of the tomogram volume. (C) Polyacrylamide denaturing gel (12%) with 10 μg DNase-treated samples after (–); No RNase treatment, (+); RNase treatment.
In order to investigate if OMVs may contain RNA, vesicle samples from V. cholerae strain A1552 were subjected to hot phenol RNA extraction and purification (Materials and Methods). After two DNase treatments, the sample was dissolved in DEPC-H2O. Two 10 μg-samples were prepared where one sample was subjected to RNase treatment. Both samples were run on a 12% denaturing PAGE gel, GelRed stained and visualized by UV light (Fig. 1C). The result strongly suggested that the separated bands from the un-treated sample mainly consisted of RNA since no bands in the RNase-treated sample were detected.
The OMV samples were prepared from dense overnight cultures of V. cholerae strain A1552 since the amount of OMVs obtained depends on the amount of bacteria used and gave a higher yield in comparison with OMV isolation from the exponentially growing bacteria. However, we also tested whether RNA can be observed in association with OMVs isolated from the exponential growth phase of bacteria. As shown in Supplementary Fig. S1A, lane 2, RNA was also associated with OMVs isolated from the exponential growth phase of bacteria. It suggested that the release of RNA from the bacterial cells in association with OMVs was not growth phase dependent. Furthermore, OMV-associated RNA was not likely due to contamination by nucleic acids from some lysis of bacterial cells. A live/dead staining analysis was performed for the estimation of the fraction dead bacterial cells as described in the Materials and Methods.
There was no apparent difference in the ratio of live and dead cells in bacterial populations when the log-phase bacterial culture and overnight culture were compared and in both cases there were very few dead cells detected (Supplementary Fig. S1C). Moreover, by Western immunoblot analysis we examined the possible presence of a cytoplasmic protein (RpoS) in samples of the culture supernatant before and after ultracentrifugation, and in OMV samples. As shown in Supplementary Fig. S1B, there was no RpoS detected in the supernatant and OMV samples although RpoS was observed in the whole cell lysate sample. The results indicated that there was little or no lysis of bacteria causing contamination of cytoplasmic components in the culture supernatants.
To clarify whether or not the detected RNA was tightly associated with the vesicles and not simply appeared there due to co-precipitation of somehow released RNA, e.g. from lysed bacterial cells during ultracentrifugation steps, OMVs were purified using an Optiprep density gradient centrifugation procedure as described in the Materials and Methods. Five distinct rings (fractions) could visually be detected in the centrifuged tube (Supplementary Fig. S2A, left panel). All five fractions were collected for further analyses. The presence of OMVs in these fractions was monitored by immunoblotting using polyclonal antiserum raised against the OmpU protein which is a major component of OMVs from V. cholerae strains and can be used as an OMV marker12. The OmpU protein was detected in the fractions 3, 4, and 5 indicating that these fractions contained OMVs (Supplementary Fig. S2A, right panel). RNA extraction was performed with each of the fractions and the presence of RNA was analysed after electrophoresis on a 12% denaturing PAGE gel by GelRed staining and visualization under UV light. As shown in Supplementary Fig. S2B, left panel, an appreciable amount of RNA was detected in the fraction 3 and a lesser amount was observed in the fraction 4 whereas there was no detectable RNA in the other fractions (Supplementary Fig. S2B, left panel, lanes 1 + 2, 3 and 4).
We concluded from these tests that RNA indeed was associated tightly with OMVs.
Sequence analysis of vesicle RNA
As a mean to directly identify the RNA detected in vesicles, we performed sequencing using MiSeq (Illumina) of rRNA-depleted RNA preparations obtained from the vesicles released by strain A1552. The RNA isolation kit was used to acquire ultrapure RNA from which most of the rRNA was removed by the procedure of the Ribo-Zero Magnetic Kit (Epicentre) before RNA library preparation as described in the Materials and Methods. After paired-end sequencing, the resulting 80 bp sequences were aligned to the sequenced chromosomal DNA of V. cholerae strain O1 El Tor N16961, a strain very closely related to the V. cholerae strain O1 El Tor A1552. A typical alignment view using Integrative Genomics Viewer (IGV) software is shown in Supplementary Fig. S3A. The following criteria were used to determine if alignments were of interest: i) that the number of short RNA sequences aligned to a specific DNA nucleotide (hit) was more than 200; ii) that these 200 DNA nucleotide hits included a homology stretch longer than 70 nucleotides. Regions of this size are presented in Supplementary Table 1A (chromosome I) and Supplementary Table 1B (chromosome II). Table 1 summarizes the total of 308 regions that fulfil the above-described criteria. Of these regions, 76 ppm (224/2,961,149 regions/bp) belong to chromosome I, whereas 77 ppm (83/1,072,315 regions/bp) belong to chromosome II. These figures indicate that the number of regions aligned to a chromosome is proportional to the size of that chromosome. This is also the case when comparing the numbers of intergenic regions. When comparing the number of regions in RNA coding for proteins with unknown functions almost half of the regions in chromosome II codes for parts of uncharacterized proteins, in contrast to one-sixth in chromosome I (Table 1). This can be explained by the fact that only half of the proteins encoded on chromosome II are characterized, while on chromosome I around five-sixths have known functions. Interestingly, the majority of the most abundant RNA detected corresponds to non-coding regions.
Table 1. Summary of OMV RNA sequence alignments to the V. cholerae N16961 genome.
| Chromosome I | Chromosome II | Bothchromosomes | |
|---|---|---|---|
| Size | 2961149 bp | 1072315 bp | 4033464 bp |
| Size/Total size | 73.4% | 26.6% | 100% |
| Regiona) | 224 | 83 | 307 |
| Region/Total size | 76 ppm | 77 ppm | 76 ppm |
| Intergenic regionb) | 11 | 3 | 14 |
| Intergenic region/Region | 4.9% | 3.6% | 4.5% |
| Up- or downstream region | 18 | 13 | 31 |
| Up- or downstream region/Region | 8.0% | 15.7% | 10.1% |
| Uncharacterizedc) | 38 | 38 | 74 |
| Uncharacterized/Region | 17.0% | 45.8% | 24.1% |
| Characterizedd) | 157 | 29 | 186 |
| Characterized/Region | 70.1% | 34.9% | 60.6% |
| Membrane associatede) | 70 | 13 | 83 |
| Membrane associated/Region | 31.1% | 15.7% | 27.0% |
| Membrane associated/Characterized | 44.6% | 44.8% | 44.6% |
| Cytoplasmicf) | 73 | 12 | 85 |
| Cytoplasmic/Region | 32.6% | 14.4% | 27.7% |
| Cytoplasmic/Characterized | 46.5% | 41.4% | 45.6% |
| sRNA | 17 | 11 | 28 |
| sRNA/Region | 7.6% | 13% | 9.1% |
a)Region: RNA-aligned DNA stretch >70 nucleotides and >200 hits.
b)Intergenic region: region situated downstream of both flanking genes.
c)Uncharacterized: region within an uncharacterized protein coding sequence.
d)Characterized: region within a characterized protein coding sequence.
e)Membrane associated: region within a membrane associated protein coding sequence.
f)Cytoplasmic: region within a cytoplasmic protein coding sequence.
Extensive studies of vesicles from Gram-negative bacteria over the last decades suggest that formation of vesicles occurs as the bacterial outer membrane creates a bleb with outer membrane proteins and lipids that surrounds the contents of the periplasm. However, a recent report by Pérez-Cruz et al. describes a sub-population of vesicles—outer-inner membrane vesicles (O-IMV) in which not only the outer but also the inner membrane is involved in the formation of the surface14. Such double-membrane vesicles would therefore have the ability to carry cytoplasmic constituents, leading to a few speculations on how RNA might enter the vesicles. One explanation might be via protein synthesis that occurs in the vicinity of the blebbing membrane. Translation machinery proteins are entrapped in vesicles of V. cholerae15 and E. coli vesicles16, thus these ribosomal proteins might simply bring their translating mRNA. Another explanation for how RNA enters vesicles is that free-floating RNA in the cytoplasm might be entrapped as well as some cytoplasmic constituents during vesicle formation. The actual localization of RNA in association with OMVs, whether or not the RNA is inside of the vesicles is as yet unclear. Attempts to use RNase treatments of OMVs as a means to reveal the localization have remained inconclusive. In our preliminary studies we observed an apparent loss of RNA after RNase treatment of OMVs. However, since it could not be determined how such treatments per se affected the integrity of the vesicles, or to what extent the RNase could enter vesicles, we need to employ additional approaches to assess the localization.
Analysis of highly transcribed RNA regions in vesicles
Our sequencing results further prompted us to investigate if the RNA sequences most frequently found in vesicles — high-hit regions — were transcribed at levels sufficient to be detected by Northern blotting. Three different probes were constructed corresponding to regions containing the highest peaks, the Highly Transcribed RNA Regions (HTRRs). These HTRRs are: the upstream region of vc2479 (uncharacterized protein) from chromosome I (1,090,000 hits), the intergenic region between vc0190 (DNA helicase II) and vc0191 (uncharacterized protein) from chromosome I (120,000 hits), and the intergenic region between vca0526 (H+/Cl- exchange transporter ClcA) and vca0527 (uncharacterized protein) from chromosome II (115,300 hits) (Supplementary Fig. S3B–D). Ultrapure RNA was extracted from A1552 whole cells and from vesicles, respectively, and analysed by Northern blot. Hybridization was done with probe vc2478,5 followed by stripping and re-probing of the same membrane using probes vc0190,5 and vca0526,5 (in that order). The Northern blot analysis clearly revealed that these high-hit regions were transcribed as the transcripts were detected in the whole cell RNA preparations and they were present at detectable levels in the vesicles (Supplementary Fig. S4A–D).
Furthermore, Northern blot analysis using probe vc2478,5 on extractions from the density gradient purified vesicle fractions also showed that fractions with RNA contained HTRRs (Supplementary Fig. S2B, right panel: lanes 3 and 4). To ensure that the DNAse I treatment could remove any contaminating DNA in the RNA preparations we performed a PCR test on extracted vesicle RNA before and after DNase I treatment using primers that would recognize DNA corresponding to the HTRR upstream of vc2479 and vc0633. Evidently the DNase I treatment successfully removed such DNA as no PCR bands were obtained after this treatment (Supplementary Fig. S5)
To verify that the vesicle RNA sequences aligned to the chromosomal V. cholerae DNA— corresponding to the HTRR regions — in fact give rise to the vesicle transcripts, we constructed three mutants (AES206, AES207, and AES208) in which the chromosomal DNA corresponding to each of these HTRRs was deleted and replaced by Km-cassettes. RNA from vesicle preparations and whole cell extracts of wild type A1552 and the three mutants were subjected to a 12% polyacrylamide denaturing gel electrophoresis and stained with GelRed (Fig. 2A). All vesicle-isolated RNA (v) samples exhibited a similar but unique band pattern, different from the likewise similar but unique whole cell extracted RNA (wc) sample pattern. Asterisks (*) in the AES207 and AES208 mutant lanes show where bands are missing in the samples from mutants. The gel-separated RNA transcripts were transferred to a membrane for Northern blotting and hybridized with probes vc2478,5 (Supplementary Fig. S4B), vc0190,5 (Supplementary Fig. S4C), and vca0526,5 (Supplementary Fig. S4D). Probing verified that these RNAs from HTRRs correspond to the identified DNA sequences on the chromosome (Fig. 2B–D). Interestingly, the size of the bands missing in the AES207 and AES208 RNA wc lanes are the same as that of the transcripts recognized by the vc2478,5 and vca0526,5 probes (Fig. 2A,B and D). Thus, the vc2478,5 and vca0526,5 region transcripts are apparently abundant enough to be visible by GelRed staining.
Figure 2. Analysis of OMV-associated RNA.
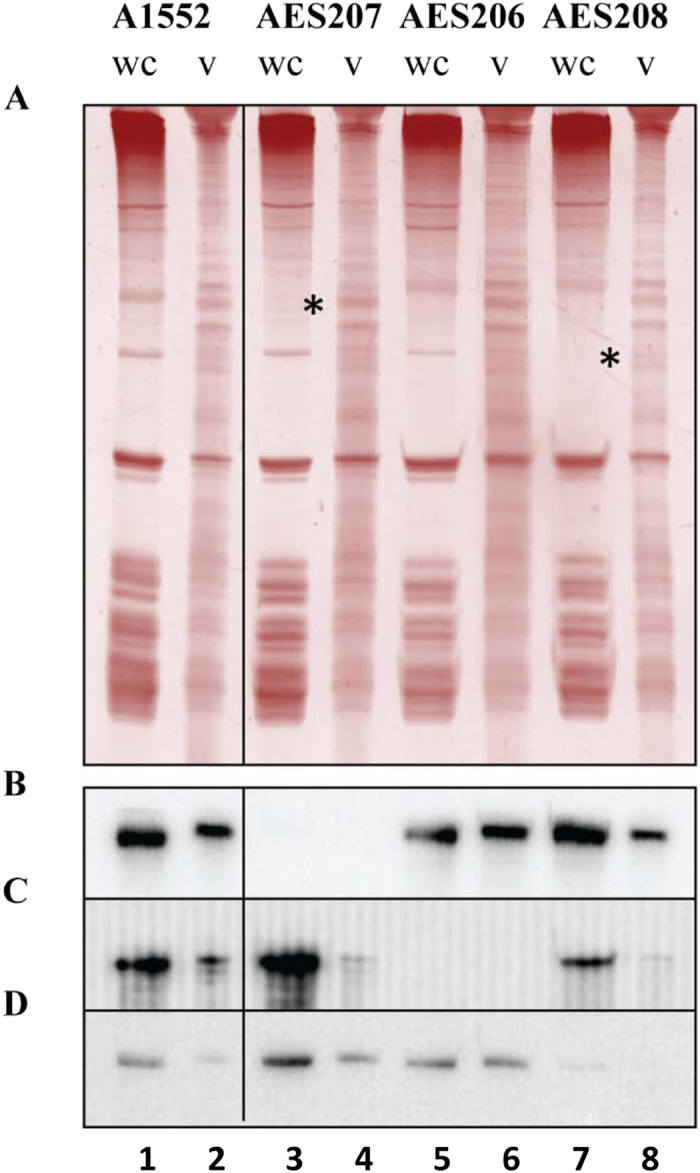
RNA extracted from overnight cultures of wt and HTRR mutant Vibrio cholerae O1 El Tor A1552 whole cells (wc) and vesicles (v) using the Total RNA Norgen kit. RNA (3.5 μg) was run on a 12% polyacrylamide denaturing gel and transferred to a membrane. (A) Gel stained with GelRed. Positions of bands missing in case of mutant strains are indicated by asterisks (*). (B) Northern blot membrane probed with probe vc2478,5. (C) Northern blot membrane re-probed with probe vc0190,5. (D) Northern blot membrane re-probed with probe vca0526,5.
As vesicles are known to be taken up by other bacteria and by eukaryotic cells12,17, we may hypothesize that such OMV-associated RNA might cause regulatory effects in the new “host” cells. OMV internalization into eukaryotic cells has been demonstrated to mediate intracellular antigen exposure and recent studies with vesicles from the bacterium Aggregatibacter actinomycetemcomitans showed that they acted as strong inducers of cytoplasmic peptidoglycan sensor NOD1- and NOD2-dependent NF-κB activation in human embryonic kidney cells18.
Interestingly, earlier studies demonstrated the successful siRNA inhibition of the Staphylococcus aureus coagulase gene and suggested that siRNAs could effectively modulate virulence and drug resistance of S. aureus19. Subsequent studies revealed that siRNAs-mediated regulation of recipient bacterial genes expression was dependent upon the RNA-binding protein Hfq20,21. In these studies, synthetic siRNA was introduced into the recipient bacterial cells using electroporation. We suggest that OMV-associated sRNA with the ability to modulate the expression of target genes could be introduced into the recipient bacterial cells using OMVs as a RNA delivery vehicle. From earlier studies, there is evidence that the OMV membrane can fuse with the outer membrane of a recipient bacterium and thereby introduce the OMV luminal contents into the recipient bacteria, including peptidoglycan hydrolase causing the lysis of recipient bacterial cells22. On the basis of earlier and present findings, we propose that it is highly likely that OMV-associated RNA can be delivered into either or both prokaryotic and eukaryotic host cells and then can cause modulation of the expression of some of their genes.
In conclusion, we report for the first time the detection and successful isolation of bacterial RNA released via OMVs from V. cholerae, providing a proof of principle that bacterial RNA can be released from bacteria via membrane vesicles. The origin of three selected sRNA was confirmed by Northern blotting analysis using the wild type strain and its respective mutant construct. Our findings show that we need to consider the potential role(s) of RNA-containing bacterial membrane vesicles in bacteria-host and bacteria-bacteria interactions.
Material and Methods
Bacterial strains, plasmids, and growth conditions
The strains and plasmids used in the present work are described in Table 2 and primers are described in Table 3. Unless otherwise stated, the strains were grown at 37 °C in either Luria–Bertani (LB) broth with vigorous shaking or on LB agar. When necessary, media was supplemented at the following concentrations: sucrose (Suc), 10% (w/v); Carbenicillin (Cb), 50 μgml−1; Kanamycin (Km), 25 μgml−1; and Rifampicin (Rif), 100 μgml−1.
Table 2. Strains and plasmids used in this study.
| Strain/Plasmid | Description/Relevant characteristics | Reference/Source |
|---|---|---|
| V. cholerae Strains | ||
| A1552 | O1 El Tor, Inaba, RifR | 27 |
| A1552∆rpoS | ∆rpoS derivative of A1552 | 28 |
| AES206 | A1552, replacement of inter-genic region VC0190 - VC0191 with Km-cassette | This study |
| AES207 | A1552, replacement of upstream region of VC2479 with Km-cassette | This study |
| AES208 | A1552, replacement of inter-genic region VCA0526 - VCA0527 with Km-cassette | This study |
| E. coli Strains | ||
| SM10λpir | thi thr leu tonA lacY supE recA::RP4-2 Tc::Mu Km λpir | 29 |
| Plasmids | ||
| pJET1.2 | Cloning vector, CbR | Fermenta |
| pKD4 | Carrying frt-flanking Km-cassette | 30 |
| pCVD442 | Suicide vector, CbR | 31 |
| pCVD-190,5::Km | pCVD442 carrying Δvc0190-1::Km | This study |
| pCVD-2478,5::Km | pCVD442 carrying Δvc2478-9::Km | This study |
| pCVD-526,5::Km | pCVD442 carrying Δvca0526-7::Km | This study |
Table 3. Primers used in this study.
| Primer name | Primer sequence (5′ to 3′) | Bold letters |
|---|---|---|
| Primers for cloning | ||
| FRTup | GTGTAGGCTGGAGCTGCTT | |
| FRTdo | CATATGAATATCCTCCTTAG | |
| vc2478upA | GCTCGCGACAGTTGGTGCTTAGCTTTTGC | NruI site |
| vc2478doB | CTAAGGAGGATATTCATATGAGGGTAAATCAATGCTGTGTTTATTGC | Complement to FRTdo |
| vc2479upC | GAAGCAGCTCCAGCCTACACGGGGTTCTCTCTATGACAGATTCTCG | Complement to FRTup |
| vc2479doD | GCTCGCGAGCAGTTGTTCGACTTGTTGGC | NruI site |
| vc2478upAA | CCAATCCTACCACTTCATTGCC | |
| vc2479doDD | GTAACGGAATATCCCAGCTTGC | |
| vc0190upA | GCTCGCGAAAGCTCTACATCACCTACG | NruI site |
| vc0190doB | CTAAGGAGGATATTCATATGCGAGTAAGACTCTAACCCC | Complement to FRTdo |
| vc0191C | GAAGCAGCTCCAGCCTACACGAAATTTTGAACA | Complement to FRTup |
| vc0191D | GCTCGCGACTTCACAAAAGGCATGATGA | NruI site |
| vc0190upAA | GAAGAGGCGGGGCGTCTCGAAG | |
| vc0191DD | CAAAAAAACGCTAGCACACCGACGC | |
| vca0526A | GCTCGCGACCATTTGTACTGTAGGACCTTC | NruI site |
| vca0526B | CTAAGGAGGATATTCATATGCCAATTCATCAATAGGTG | Complement to FRTdo |
| vca0527upC | GAAGCAGCTCCAGCCTACACGGATATACTTCGGCCAAAAGG | Complement to FRTup |
| vca0527doD | GCTCGCGACACTGATCGACTTTGTTCCCC | NruI site |
| vca0526AA | GCGCGTATCTTCATTTTTGACACG | |
| vca0527doDD | GGCATCAATGTCACTACTGCTG | |
| Primers for Northern blot probes | ||
| vc2478,5up | CCCTGGGGTGTTCGTCAGCGGAT | |
| vc2478,5do | CGTTGAGGATCGTAGCCCTTGAAC | |
| vc0190,5up | CCTTGATAGTACGCTGCAATCCGTC | |
| vc0190,5do | GGACATTGAACGGACGCAATCG | |
| vca0526,5up | GATGTGCCGGAATTATACGCTGC | |
| vca0526,5do | GGTTCCATATCCTCCATCAGAAATG | |
| ompUup1 | GCGTCGACGCTTGATGCATCACCTATTTCG | |
| ompUdo3 | GTCAACACGGTCTGCTACAGC | |
Construction of plasmids for Km cassette replacement mutagenesis
Mutant plasmid constructs were designed with a 1500 bp Km-cassette flanked by 500-bp sequences corresponding to upstream and downstream regions of the target gene. The Km-cassette and flanking regions co-amplified as one unit by PCR. The upstream (A and B primers) and downstream (C and D primers) flanking regions were PCR amplified (Kapa HiFi HotStart Polymerase, Kapa Biosystems) from A1552 chromosomal DNA and cloned into pJET1.2 for sequencing and amplification purposes. Correct pJET1.2 constructs were digested with XhoI and XbaI and the desired fragments were gel purified. The Km-cassette was PCR amplified from plasmid pKD4 (FRTup and FRTdo primers, DreamTaq Green PCR Master Mix (Fermenta)) and gel-purified. Fifty to 90 ng of the flanking region DNA and 200 ng of the Km-cassette DNA were used for cross-over PCR (Kapa HiFi HotStart Polymerase, Kapa Bisystems) with the following conditions: anneal 43 °C–20 s, elongation 69 °C–1.45 min). The reaction was purified on a gel where the resulting 2500-bp fragments were excised and cloned into pJET1.2 for sequencing and amplification purposes. Correct constructs were digested with NruI and the gel-purified fragments were cloned into SmaI-digested pCVD442 yielding plasmids pCVD-190,5::Km, pCVD-2478,5::Km, and pCVD-526,5::Km.
Genetic manipulation to obtain Km replacement mutants
Wild type V. cholerae A1552 (recipient) was cross streaked with SM10λ/pCVD-190,5::Km, SM10λ/pCVD-2478,5::Km, and SM10λ/pCVD-526,5::Km (donors), respectively, on non-selective LB plates and incubated at 37 °C for 6 h. A1552/pCVD-construct integrants (cross sections) were then selected for on LB + Km + Rif plates. To cure the integrants of transfer plasmids and to select for homologous recombination of Km-cassette replacement mutations, KmR + RifR-colonies were suspended in LB, serially diluted, and spread on LB + Km + Suc plates. Successful mutagenesis of plasmid-free (CbS) Km-cassette replacement clones was verified by PCR using AA and DD primers. Mutagenesis gave rise to strains AES206 (A1552Δvc0190 - vc0191), AES207 (A1552Δvc02478 - vc2479), and AES208 (A1552Δvca0527 - vca527).
Vesicle isolation
OMVs were isolated from bacterial culture supernatants essentially as described previously8. Briefly, bacteria were inoculated in 1 litre flasks containing 280 ml LB. Cultures were grown with shaking at 37 °C for 16 h and then centrifuged at 8,250 × g for 15 min at 4 °C. Pellets were discarded and supernatant re-centrifuged twice. This supernatant was filtered twice through vacuum driven 0.2 μm pore size stericup filters (Millipore Express™ Plus) and ultra-centrifuged at 100,000 × g for 2 h at 4 °C in a 45 Ti rotor (Beckman). The vesicle pellets were re-suspended in 600 μl TE buffer and kept at −80 °C until further use.
Density gradient centrifugation
Isolated vesicles suspended in TE buffer were fractionated by the density gradient centrifugation method slightly modified from previously described12. Shortly, a gradient of Optiprep (Sigma-Aldrich) was created in a 4 ml ultracentrifugation tube (0.4 ml 45%, 0.5 ml 35%, 0.6 ml 30%, 0.6 ml 25%, 0.6 ml 20%, 0.5 ml 15% and 0.6 ml 10%). The vesicle sample was added and the tube was centrifuged (180, 000 × g, 3 h, 4 °C) in a SW60Ti rotor (Beckman). After centrifugation visual fraction bands were extracted (100–300 μl), washed in 60 ml TE buffer, and ultra-centrifuged at 100,000 × g for 2 h at 4 °C in a 45 Ti rotor (Beckman). The resulting fraction pellets were re-suspended in 20–80 μl TE buffer. 1 μl of the suspended fractions were taken for immunoblot analysis with anti OmpU and the rest kept at −80 °C until further use.
For tests with RNase treatment, vesicles obtained from an overnight culture were suspended in 300 μl TE buffer and 50 μg RNase A (QIAgen) was added. The sample was kept at RT for 15 min, washed in 70 ml TE buffer, and ultra-centrifuged at 100,000 × g for 2 h at 4 °C in a 45 Ti rotor (Beckman). The resulting vesicle pellet was re-suspended in 60 μl SUPERase• In™ RNase Inhibitor buffer (20 mM Tris-HCl, pH 7.5, 50 mM NaCl, 1 mM EDTA) and 100U inhibitor (Life Technologies) was added before analysis.
Live/dead staining and fluorescence microscopy analyses
Live/dead staining was performed using the LIVE/DEAD® BacLightTM Bacterial Viability Kit L13152 according to the manual. In short, V. cholerae O1 El Tor strain A1552 cultures grown to OD600 = 0.7 (log-phase) or OD600 = 4.6 (overnight culture) were harvested by centrifugation and suspended in 0.85% NaCl. After additional centrifugation and washing, bacterial suspensions of 106 bacteria/ml were subjected to staining as described by the manufacturer (Molecular Probes). Treatment of a sample with isopropanol was done to obtain a population of only dead bacteria for comparison. The staining was examined and visualized in a Nikon fluorescence microscope with bandpass filters for Texas Red and Fluorescein, respectively.
Electron microscopy
For negative staining 3.5 μl of sample was adsorbed for 2 min onto glow-discharged formvar and carbon-coated copper grids, washed in H2O and immediately negatively stained in 50 μl of 1.5% uranyl acetate solution for 30 s. Negative-stained samples were examined on a JEOL JEM1230 TEM operating at 80 kV. Micrographs were recorded with a Gatan MSC 600CW CCD camera using Digital Micrograph software. For cryo-electron tomography 4 μl sample was vitrified on a holey carbon film using FEI Vitrobot. The tomography data was collected on a JEOL 2200FS TEM with SerialEM software and a pixel size of 0.57 nm at the Caesar institute in Bonn, Germany. The 3D reconstruction was performed with SIRT and visualized using the IMOD package from University of Colorado at Boulder23.
RNA extraction
RNA was extracted from whole cells (1 ml culture) or vesicles isolated from 2.5–5 litre cultures. Extraction was initially made by modified hot phenol extraction methods described by von Gabain et al.24. Briefly, vesicles were disrupted in 3% SDS followed by RNA hot phenol extraction. After DNase I (Fermenta) treatment, phenol::chloroform purification, and ethanol precipitation the resulting RNA pellet was dissolved in 100 μl DEPC-treated water.
Less yield but cleaner RNA was obtained using the Total RNA Purification Plus Kit (Norgen Biotek corp) according to the manufacturer’s manual followed by treatment with DNase I. RNA was kept at −80 °C until further use.
To check if the DNase I treatment efficiently removed any contaminating DNA, the RNA preparations were subjected to PCR amplification using DreamTaq Green PCR Master Mix (2×) (Fermenta) and two primer pairs, ompU1up + ompU3do and vc2478,5up + vc2478,5do, respectively. The templates for each set were 100ng vesicle RNA isolated with the Norgen kit and samples without or with DNase I treatment were tested.
RNA preparation for the library construction
Isolated vesicle RNA was treated with the Ribo-Zero Magnetic Kit for Gram-Negative Bacteria (Epicentre) according to the manufacturer’s instructions to reduce the amount of rRNA. This rRNA-depleted RNA was used for library construction using the TruSeq RNA sample prep kit v2 (Illumina) according to the manufacturer’s protocol (#15026495 revD).
Briefly, the rRNA-depleted RNA was fragmented and the RNA was reverse transcribed into cDNA using random-sequence primers and SuperScript III reverse transcriptase. The cDNA was purified using AMPure XP beads (Beckman Coulter) and amplified for 15 cycles of PCR with barcoded primers, followed by purification using AMPure beads.
Cluster generation and sequencing
An 18-pM solution of RNA was subjected to cluster generation and paired-end sequencing with 80-bp read length on the MiSeq system (Illumina Inc.) using the v3 chemistry according to the manufacturer’s protocols.
Base calling was done on the instrument by RTA 1.18.42 and the resulting .bcl files were demultiplexed and converted to fastq format with tools provided by CASAVA 1.8.2 (Illumina Inc.), allowing for one mismatch in the index sequence. Additional statistics on sequence quality were compiled with an in-house script from the fastq-files, RTA and CASAVA output files. Sequencing was performed by the SNP&SEQ Technology Platform in Uppsala, Sweden www.sequencing.se.
RNA sequencing analysis
RNA sequences were aligned to the N16961 Vibrio cholerae chromosomes using Integrative Genomics Viewer (IGV) software25,26
Polyacrylamide gel electrophoresis (PAGE) and Northern blot analyses
Whole cell RNA and vesicle RNA from the strains were separated on a 12% acrylamide:bis-acrylamide 19:1/8 M urea gel. The gel was stained with GelRed dye (Biotium) for visualization and then the RNA was transferred onto an Amersham Hybond™-XL membrane (GE Healthcare) for 1 h at 120 mA, 15 V on a semi-dry transfer unit (EBU-4000, C. B. S Scientific). The blotted membrane was cross-linked with UV. 30 ng of specific probes generated by PCR were [α-32P]ATP-labelled using the Prime-a-Gene® Labeling System (Promega) according to the manufacturer’s instructions. Unincorporated nucleotides were removed with MicroSpin™ G-25 Columns (GE Healthcare). Prehybridization (4 h) and hybridization (16 h) was performed in Roti-Hybri Solution (Roth) at 58 °C. Bands were visualized using a Phospho-Imager, Storm 860 (Molecular Dynamics).
Stripping of the membrane was done by 2 × 15 min washing on shaker at RT in Wash 1 (0.01 M EDTA, 0.05 × SSC, 0.1% SDS—heated to 100 °C) followed by 2 × 15 min washing on shaker at RT in Wash 2 (0.02 × SSC).
SDS-PAGE and Western blotting analyses
Protein extracts were separated by SDS-polyacrylamide gel electrophoresis and thereafter transferred to a polyvinylidene fluoride (PVDF) micro-porus membrane. OmpU polyclonal antibodies were used to detect outer membrane constituents (vesicles) in the Optiprep fractions. RpoS was used as possible cytoplasmic hallmark for lysed bacteria in the supernatants recognizable by anti-RpoS polyclonal antibodies. Anti-rabbit HRP-conjugated antibody (Agrisera) was used as secondary antiserum at a final dilution of 1:25,000. Further visualization was carried out using ClarityTM Western ECL substrate as described by the manufacturer (Bio-Rad). Bands were detected using the ChemiDoc XRS system (Bio-Rad).
Additional Information
How to cite this article: Sjöström, A. E. et al. Membrane vesicle-mediated release of bacterial RNA. Sci. Rep. 5, 15329; doi: 10.1038/srep15329 (2015).
Supplementary Material
Acknowledgments
Sequencing was performed by the SNP&SEQ Technology Platform in Uppsala which is part of the National Genomics Infrastructure (NGI) Sweden and Science for Life Laboratory. The SNP&SEQ Platform is also supported by the Swedish Research Council and the Knut and Alice Wallenberg Foundation. Special acknowledgements for cryo-EM data collection to Dr. Stephan Irsen at Caesar in Bonn, Germany and JEOL GmbH, Germany. This work was performed within the Umeå Centre for Microbial Research (UCMR) Linnaeus Program and was supported by project grants to SNW and to BEU from the Swedish Research Council and to LS from the Swedish Research Council and the Kempe Foundations.
Footnotes
Author Contributions A.E.S., L.S., B.E.U. and S.N.W. conceived the study and designed experiments, analyzed the data, and wrote the manuscript. A.E.S. and L.S. performed experiments. L.S., B.E.U. and S.N.W. contributed materials and analysis tools. All authors reviewed the manuscript.
References
- Kulp A. & Kuehn M. J. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annual review of microbiology 64, 163–184, 10.1146/annurev.micro.091208.073413 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zomer A. et al. Exosomes: Fit to deliver small RNA. Communicative & integrative biology 3, 447–450, 10.4161/cib.3.5.12339 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valadi H. et al. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nature cell biology 9, 654–659, 10.1038/ncb1596 (2007). [DOI] [PubMed] [Google Scholar]
- Zhong H. et al. Induction of a tumour-specific CTL response by exosomes isolated from heat-treated malignant ascites of gastric cancer patients. International journal of hyperthermia: the official journal of European Society for Hyperthermic Oncology, North American Hyperthermia Group 27, 604–611, 10.3109/02656736.2011.564598 (2011). [DOI] [PubMed] [Google Scholar]
- Pegtel D. M. et al. Functional delivery of viral miRNAs via exosomes. Proceedings of the National Academy of Sciences of the United States of America 107, 6328–6333, 10.1073/pnas.0914843107 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berleman J. & Auer M. The role of bacterial outer membrane vesicles for intra- and interspecies delivery. Environmental microbiology 15, 347–354, 10.1111/1462-2920.12048 (2013). [DOI] [PubMed] [Google Scholar]
- Bonnington K. E. & Kuehn M. J. Protein selection and export via outer membrane vesicles. Biochimica et biophysica acta 1843, 1612–1619, 10.1016/j.bbamcr.2013.12.011 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wai S. N. et al. Vesicle-mediated export and assembly of pore-forming oligomers of the enterobacterial ClyA cytotoxin. Cell 115, 25–35 (2003). [DOI] [PubMed] [Google Scholar]
- Wai S. N., Takade A. & Amako K. The release of outer membrane vesicles from the strains of enterotoxigenic Escherichia coli. Microbiology and immunology 39, 451–456 (1995). [DOI] [PubMed] [Google Scholar]
- Lindmark B. et al. Outer membrane vesicle-mediated release of cytolethal distending toxin (CDT) from Campylobacter jejuni. BMC microbiology 9, 220, 10.1186/1471-2180-9-220 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duperthuy M. et al. Role of the Vibrio cholerae matrix protein Bap1 in cross-resistance to antimicrobial peptides. PLoS pathogens 9, e1003620, 10.1371/journal.ppat.1003620 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elluri S. et al. Outer membrane vesicles mediate transport of biologically active Vibrio cholerae cytolysin (VCC) from V. cholerae strains. PloS one 9, e106731, 10.1371/journal.pone.0106731 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Boer P. A., Crossley R. E. & Rothfield L. I. A division inhibitor and a topological specificity factor coded for by the minicell locus determine proper placement of the division septum in E. coli. Cell 56, 641–649 (1989). [DOI] [PubMed] [Google Scholar]
- Pérez-Cruz C. et al. New type of outer membrane vesicle produced by the Gram-negative bacterium Shewanella vesiculosa M7T: implications for DNA content. Applied and environmental microbiology 79, 1874–1881, 10.1128/AEM.03657-12 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altindis E., Fu Y. & Mekalanos J. J. Proteomic analysis of Vibrio cholerae outer membrane vesicles. Proceedings of the National Academy of Sciences of the United States of America 111, E1548–1556, 10.1073/pnas.1403683111 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee E. Y. et al. Global proteomic profiling of native outer membrane vesicles derived from Escherichia coli. Proteomics 7, 3143–3153, 10.1002/pmic.200700196 (2007). [DOI] [PubMed] [Google Scholar]
- Thay B., Damm A., Kufer T. A., Wai S. N. & Oscarsson J. Aggregatibacter actinomycetemcomitans outer membrane vesicles are internalized in human host cells and trigger NOD1- and NOD2-dependent NF-kappaB activation. Infection and immunity 82, 4034–4046, 10.1128/IAI.01980-14 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thay B., Damm A., Kufer T. A., Wai S. N. & Oscarsson J. Aggregatibacter actinomycetemcomitans outer membrane vesicles are internalized in human host cells and trigger NOD1- and NOD2-dependent NF-kappaB activation. Infection and immunity 82, 4034–4046, 10.1128/IAI.01980-14 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yanagihara K. et al. Effects of short interfering RNA against methicillin-resistant Staphylococcus aureus coagulase in vitro and in vivo. The Journal of antimicrobial chemotherapy 57, 122–126, 10.1093/jac/dki416 (2006). [DOI] [PubMed] [Google Scholar]
- Aiba H. Mechanism of RNA silencing by Hfq-binding small RNAs. Current opinion in microbiology 10, 134–139, 10.1016/j.mib.2007.03.010 (2007). [DOI] [PubMed] [Google Scholar]
- Sonnleitner E. & Haas D. Small RNAs as regulators of primary and secondary metabolism in Pseudomonas species. Applied microbiology and biotechnology 91, 63–79, 10.1007/s00253-011-3332-1 (2011). [DOI] [PubMed] [Google Scholar]
- Li Z., Clarke A. J. & Beveridge T. J. Gram-negative bacteria produce membrane vesicles which are capable of killing other bacteria. Journal of bacteriology 180, 5478–5483 (1998). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kremer J. R., Mastronarde D. N. & McIntosh J. R. Computer visualization of three-dimensional image data using IMOD. J Struct Biol 116, 71–76, 10.1006/jsbi.1996.0013 (1996). [DOI] [PubMed] [Google Scholar]
- von Gabain A., Belasco J. G., Schottel J. L., Chang A. C. & Cohen S. N. Decay of mRNA in Escherichia coli: investigation of the fate of specific segments of transcripts. Proceedings of the National Academy of Sciences of the United States of America 80, 653–657 (1983). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson J. T. et al. Integrative genomics viewer. Nature biotechnology 29, 24–26, 10.1038/nbt.1754 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thorvaldsdóttir H., Robinson J. T. & Mesirov J. P. Integrative Genomics Viewer (IGV): high-performance genomics data visualization and exploration. Briefings in bioinformatics 14, 178–192, 10.1093/bib/bbs017 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yildiz F. H. & Schoolnik G. K. Role of rpoS in stress survival and virulence of Vibrio cholerae. Journal of bacteriology 180, 773–784 (1998). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valeru S. P. et al. Role of melanin pigment in expression of Vibrio cholerae virulence factors. Infection and immunity 77, 935–942, 10.1128/IAI.00929-08 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller V. L. & Mekalanos J. J. A novel suicide vector and its use in construction of insertion mutations: osmoregulation of outer membrane proteins and virulence determinants in Vibrio cholerae requires toxR. Journal of bacteriology 170, 2575–2583 (1988). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Datsenko K. A. & Wanner B. L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proceedings of the National Academy of Sciences of the United States of America 97, 6640–6645, 10.1073/pnas.120163297 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donnenberg M. S. & Kaper J. B. Construction of an eae deletion mutant of enteropathogenic Escherichia coli by using a positive-selection suicide vector. Infection and immunity 59, 4310–4317 (1991). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.