

Communication between cells is a vital process that governs cellular organization and coordination in an organism. Thus, multiple mechanisms of cell communication have evolved to respond to all the needs of an organism. The modes of cell-cell communication range from the ones that require direct contact between cells, such as cytoplasmic bridges (CBs) and gap junctions, to others that may take place over long distances throughout the organism, such as hormone signaling. Precise integration of data acquired from all these different modes of communication maintains tissue homeostasis and allows optimal adaptation of an organism to various stresses.
Cellular senescence represents one of the outcomes of cellular response to stress.1 Senescent cells execute essential functions in different physiological and pathophysiological conditions. They are present in pre-malignant lesions, sites of tissue damage, aging tissues and even during embryonic development. In all these places senescent cells communicate with cells in their surroundings and modulate the function of these cells.2-5 The effect of senescent cells on nearby cells is commonly attributed to the secretion of cytokines, chemokines and matrix metalloproteinases.4,6 We have recently demonstrated that in addition to secretion, senescent cells affect neighboring cells by direct intercellular protein transfer (IPT).2 Proteins from senescent cells are directly transferred to recipient neighboring cells, such as immune and cancer cells, triggering activation of signaling pathways in these cells, ultimately leading to changes in cellular behavior.
We have detected IPT from senescent cells and demonstrated that these cells form CBs with other cells, including immune cells, cancer and non-cancer epithelial cells.2 CBs, which in some conditions are called tunneling nanotubes, are open on both edges and allow transfer of cytoplasmic content to neighboring cells. Interestingly, we also identified mitochondria and lysosomes in CBs, implying that organelles might be transferred between the cells. Transfer of these organelles was indeed identified in several other systems. Since dysfunctional mitochondria are thought to play a role in cellular senescence, it can be speculated that transfer of the dysfunctional mitochondria to healthy neighboring cells may induce senescence in these cells. Transfer of cytoplasmic content or whole organelles might serve therefore, for communication of cellular stress.
The functional consequences of cell communication of senescent cells through IPT are not completely understood and could be cell type dependent. For instance, we have reported that IPT facilitates elimination of senescent cells by NK cells since transfer of proteins to NK cells correlated with NK cell activation and cytotoxicity. Remarkably, inhibition of CDC42 in senescent cells resulted in decreased IPT and impaired elimination of the senescent cells by NK cells.2 In addition to interaction with NK cells senescent cells also communicate with other components of the innate and adaptive immune systems.2,5,6 Thus, it is possible that senescent cells transfer proteins to other types of immune cells and alter their behavior. To fulfill their function immune cells respond to cytokines and chemokines using receptor-ligand interactions to activate cellular responses (Fig. 1). IPT can serve as an additional mechanism regulating activity of immune cells following contact mediated interaction with resident cells and thereby fine-tune the immune response.
Figure 1.
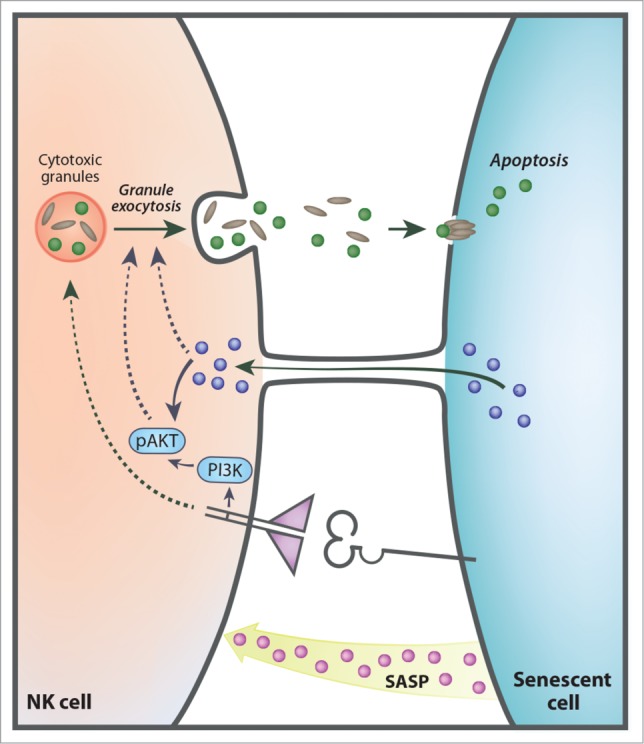
Senescent cells communicate with NK cells by multiple mechanisms. Senescent cells influence NK cells by (i) senescent associated secretory phenotype (SASP); (ii) specific receptor ligand interactions; (iii) cytoplasmic bridges. These diverse mechanisms trigger PI3K and p-AKT pathways, resulting in activation of the granule exocytosis in the NK cells and elimination of the senescent target cells by activation of apoptosis.
Senescent cells can impact tumorigenesis, mainly by secretion of pro-inflammatory cytokines.3,5 Our data demonstrate that senescent cells form CBs with cancer cells, thereby implementing IPT in interaction of senescent and cancer cells. Since the interaction of these cells is long lasting, in contrast to the interaction with NK cells, higher amount and number of proteins could potentially be transferred. The functional impact of IPT from senescent cells to cancer cells is unknown, as it could potentially promote or restrain cancer cells. Senescent cells could transfer cell cycle inhibitors (i.e. p16, p15 and p21) to cancer cells, thereby inhibiting their proliferation. Conversely, cancer cells may also receive organelles and proteins to support their rapid proliferation.
In addition to pathological conditions, senescent cells modulate physiological processes, including embryonic development.4 During embryonic development human placental syncytiotrophoblast exhibit features and molecular markers of cellular senescence.7 The syncytiotrophoblast is a multinucleate epithelium which supports fetal growth by creating an interface between maternal and fetal circulation. The syncytiotrophoblast cells communicate with subjacent layer of mono-nucleated cytotrophoblasts to promote division and fusion of cytotrophoblasts into the syncytiotrophoblast. These cells also communicate with immune cells to control immune tolerance at the maternal-fetal interface. Therefore, senescent syncytiotrophoblast cells might also form CBs to facilitate these non-cell autonomous effects which are essential for embryonic development.
Overall, IPT from senescent cells represents a pivotal mode of cellular communication with possible physiological outcomes on cancer progression, tissue repair, embryonic development, aging and immune modulation.
References
- 1.Burton DG, Krizhanovsky V. Cell Mol Life Sci 2014; 71:4373-86. PMID:25080110; http://dx.doi.org/ 10.1007/s00018-014-1691-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Biran A, et al.. Genes Dev 2015; 29:791-802. PMID:25854920; http://dx.doi.org/ 10.1101/gad.259341.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kuilman T, et al.. Genes Dev 2010; 24:2463-79. PMID:21078816; http://dx.doi.org/ 10.1101/gad.1971610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ovadya Y, Krizhanovsky V. Biogerontology 2014; 15(6):627-42. PMID:25217383; http://dx.doi.org/ 10.1007/s10522-014-9529-9. [DOI] [PubMed] [Google Scholar]
- 5.Salama R, et al.. Genes Dev 2014; 28:99-114. PMID:24449267; http://dx.doi.org/ 10.1101/gad.235184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sagiv A, Krizhanovsky V. Biogerontology 2013; 14:617-28. PMID:24114507; http://dx.doi.org/ 10.1007/s10522-013-9473-0. [DOI] [PubMed] [Google Scholar]
- 7.Chuprin A, et al.. Genes Dev 2013; 27:2356-66. PMID:24186980; http://dx.doi.org/ 10.1101/gad.227512.113. [DOI] [PMC free article] [PubMed] [Google Scholar]