

Abstract
The complexity of Stone Age tool-making is assumed to have relied upon cultural transmission, but direct evidence is lacking. This paper reviews evidence bearing on this question provided through five related empirical perspectives. Controlled experimental studies offer special power in identifying and dissecting social learning into its diverse component forms, such as imitation and emulation. The first approach focuses on experimental studies that have discriminated social learning processes in nut-cracking by chimpanzees. Second come experiments that have identified and dissected the processes of cultural transmission involved in a variety of other force-based forms of chimpanzee tool use. A third perspective is provided by field studies that have revealed a range of forms of forceful, targeted tool use by chimpanzees, that set percussion in its broader cognitive context. Fourth are experimental studies of the development of flint knapping to make functional sharp flakes by bonobos, implicating and defining the social learning and innovation involved. Finally, new and substantial experiments compare what different social learning processes, from observational learning to teaching, afford good quality human flake and biface manufacture. Together these complementary approaches begin to delineate the social learning processes necessary to percussive technologies within the Pan–Homo clade.
Keywords: percussive technology, nut-cracking, stone tools, social learning, cultural transmission, chimpanzee
1. Introduction
If we take the beginnings of the Bronze Age, approximately 6000 years ago, as the transition from our species' reliance on lithic to metal cutting tools, and couple that with the known 2.6 Ma [1] (or perhaps now 3.3 Ma1) history of hominin stone tool making, we arrive at no less than 99.8% of this latest period of our evolutionary history having been spent in the manufacture of stone tools. Accordingly, percussive technology must have been a fundamental component in the evolutionary shaping of our species over this period, suggesting a substantial legacy in the associated mental and neural processes.
Study of the lithic remains illuminates not only the evolving skills necessary to make them, but also the evolution of human cultural capacities, insofar as material culture can be traced in the lithic archaeological record. Comparisons across time and space in Africa have delineated the slow beginnings of cumulative cultural evolution in the transition from elementary Oldowan tools to more sophisticated bilaterally symmetric Acheulian bifaces, and also within these major phases, with complexity and diversity rising progressively over the millennia [3–5]. However, this follows only so long as it is inferred that these changes were indeed culturally transmitted by forms of social learning. Unfortunately, such processes remain frustratingly opaque in the archaeological record: we simply cannot directly document their operation, as we can with living species.
A more direct empirical approach to this question of what cultural processes were necessary for the transmission of percussive technologies can be borrowed from the experimental study of social learning that has blossomed over recent decades, particularly in comparative [6,7] and developmental psychology [8,9]. These disciplines have developed a range of experimental designs to test and dissect the role of social learning and the many different forms it can take, in the acquisition of manipulative skills, including tool use [7–9].
Perhaps surprisingly, such experimental approaches to cultural transmission have only very recently been seriously applied to the acquisition of stone-knapping skills. Just two recent substantial experiments of this kind are available for review here [10,11; §6]. Perhaps their rarity is explicable by their practical demands, for these two studies analysed 18 000 and 6000 lithic products, respectively, the latter alone consuming two tons of flint.
Fortunately other, complementary sources of evidence can also be applied to understanding the evolution of percussive technology. A comparative approach has also become illuminating, particularly concerning our closest living relatives, the common chimpanzees and bonobos of the genus Pan, with whom we last shared a common ancestor around 6 Ma. The use of wooden and stone hammers and anvils for nut-cracking by West African chimpanzees has long been recognized for its similarities to human knapping and its consequent relevance for reconstructing the ancestral foundations of hominin percussive behaviour [12,13]. As in human communities displaying recent functional knapping cultures, chimpanzee nutcrackers typically sit truncally erect and with controlled force may use a hammer object held in one hand, to precisely target their objective. Because unlike the makers of ancient stone tools, these are living beings, rich observational studies have described the development of the requisite manipulative skills in the wild, including observations implicating cultural transmission [14–16]. However as in the case of human knapping, experimental approaches are particularly powerful when it comes to rigorously identifying a role for forms of social learning, and the experimental studies concerning the development of chimpanzee nut-cracking are reviewed in §2. The broader corpus of experimental studies dissecting the cultural transmission of chimpanzee tool use other than percussion per se is further examined in §3.
However nut-cracking does not exhaust the percussive repertoire of chimpanzees, who employ a wider range of forceful targeted tool use to a variety of foraging tasks in the wild [13]. To provide a more comprehensive account of chimpanzees' percussive and related tool use, these behaviours are briefly summarized in §5. The ape studies of §§2, 3 and 5 are valuable particularly because they concern our closest primate relatives, generating inferences about the origins of percussive technologies in our common ancestry [17]. Percussive technology by other animal taxa also contributes to our understanding because it illuminates convergent evolution that often highlights adaptive responses shared by the species concerned [18,19]. Such studies are discussed elsewhere in this issue [20].
A different rationale underlies studies that have specifically targeted stone tool making, and asked just what capacities apes possess and may display if given appropriate opportunities. Unlike the research of §§2, 3 and 5, this work has deliberately taken apes beyond their natural repertoire to address what pre-adaptations to stone tool knapping likely existed in ancestral apes. This research was begun with bonobos in the 1990s [21,22] but has recently been revisited with the focus shifted to functional outcomes achieved [23] and is reviewed in §4.
2. Experimental studies of nut-cracking by chimpanzees
(a). Background
Common chimpanzees (Pan troglodytes) have now been studied intensively in the field for over 50 years, and evidence for variations in behavioural profiles of communities across Africa has steadily accumulated. Building on a series of studies documenting such differences, collaboration between the leaders of nine long-term study sites achieved the first systematic analysis in 1999–2001 [24,25]. Setting aside regional variations in behaviour that were likely explicable by environmental differences, this analysis identified 39 putative traditions. One of the clearest is the percussive activity of nut-cracking, which occurs across a swathe of far-west Africa about 700 km from north to south [13], but not in central or East Africa (an earlier report based on cracked nuts in Cameroon [26] remains unconfirmed). This behaviour appears robust with respect to the potential difficulty of reliably excluding environmental explanations for the differences documented: two independent expeditions to non-nut-cracking locations confirmed that the raw materials of nuts and suitable hammers were readily available [27,28]. Potential genetic differences, whereby chimpanzees in the west have evolved innate dispositions that others lack [29], were harder to reject from field observations, but the experiments described below in §2(b,c) allow us to do this.
Clinching evidence of social transmission could in principle be achieved by experimentally translocating a nut-cracking expert into a naive wild population, but this has so far been judged logistically and probably ethically untenable. Instead, a suite of experiments have been completed with captive chimpanzees, including some wild born and resident in African sanctuaries. Those focused on nut-cracking are reviewed next (§2b–d).
(b). Experimental studies lacking a no-model control condition
I divide the experimental studies into two sets on the basis of the incorporation of control conditions. Perhaps the most basic experimental design in social learning research involves an experimental condition in which participants witness a model individual performing a novel act, and a control set of participants lacking such a model [6–9]. Comparing these can rigorously demonstrate any effect of learning from the model in the experimental condition. Unfortunately, often because of limited numbers of participants available, or where a no-model condition is impracticable, only one study has managed this experimental design and is described in the §2(c). The results of the others, outlined next, are all consistent with the operation of social learning, and compelling to different degrees, despite the lack of a clinching, between-subjects control condition.
Sumita et al. [30] exposed five chimpanzees to nut-cracking by both a conspecific and human demonstrator. Three acquired nut-cracking after this observation, by contrast with a prior baseline control condition in which presentation of raw materials elicited no nut-cracking. This was too small a sample to demonstrate a statistically significant change, and the ‘baseline followed by experimental condition’ design demands some caution with respect to whether we are seeing only the effect of more prolonged exposure, rather than social learning.
Hannah & McGrew [31] instead exploited a situation in which 16 wild-born chimpanzees were released onto a forested island in Liberia. After six weeks with no tool-use behaviour occurring, a newly introduced female began to actively crack nuts. Within 3 days, three others began to nut-crack and after two months 13 chimpanzees were nut-cracking. This appears a compelling contrast with the initial six weeks that provided a baseline control condition. However, the prior history of these chimpanzees was unknown and it remains possible, given the West African location, that they had some earlier experience of nut-cracking. The rapid take-up of nut-cracking after the first demonstration provides evidence for social learning, but uncertainty remains about whether this minimally involved the triggering of prior experience or whether instead, naive chimpanzees learned observationally how to nut-crack.
Similarly, Hayashi et al. [32] reported that already on the first day of observing a human cracking macadamia nuts, two of three human-reared chimpanzees placed a nut on an anvil stone and hammered it, with one succeeding on that day and the second on the next session. This is again consistent with social learning, yet there was no baseline or other control condition reported, weakening the strength of the evidence.
Finally, Hirata et al. [33] exposed four juvenile chimpanzees to a peer cracking macadamia nuts and all eventually succeeded in nut-cracking, by contrast with an extended baseline phase of 15 sessions in which none had done so. However, it took respectively 8, 11, 13 and 15 more sessions for success to occur, so the study suffers the same problems of small sample size and lack of between-subjects controls to reliably identify social learning. Nevertheless, this study is particularly valuable in the detailed analysis of a long, drawn out process of nut-cracking acquisition among juvenile chimpanzees, echoing the slow development of the skill in the wild [14–16]. For example, the numbers of bouts of conspecifics' nut-crackings observed before each individual's success was, respectively, 2958, 3114, 3184 and 5604 for the four participants. We are used to seeing examples of rapid social learning in our own species, but these results underline that other primates may require much more extended schedules of observation to support social learning, perhaps because of their more limited neural resources and associated cognitive processing limitations (a chimpanzee brain is only about one-third of the human size). In the wild, a young chimpanzee will probably observe many hundreds of expert nut-cracking episodes before it has the motor capacities to succeed itself.
The observation time before success clearly varies greatly in these studies. This and other comparisons across the experiments are best addressed after the final experiment in this corpus is outlined (§2c).
(c). A controlled experiment on social learning of nut-cracking in East African chimpanzees
Working in an African sanctuary housing juvenile chimpanzees, Marshall-Pescini & Whiten [34] had sufficient youngsters aged 3–6 years to compare an experimental group witnessing models cracking oil palm nuts with a control group who had no model. The latter were then also exposed to the model, thus fully exploiting the potential for statistical comparisons both between and within groups (figure 1). Experimental participants witnessed a skilled conspecific nut-cracking during a first phase of five consecutive daily sessions of 30–60 min. Control participants were provided with the same raw materials over the same period, then in a second phase all participants were exposed to a human experimenter cracking five nuts during 15-min sessions on each of five days, interspersed with opportunities for the participants to crack nuts themselves.
Figure 1.
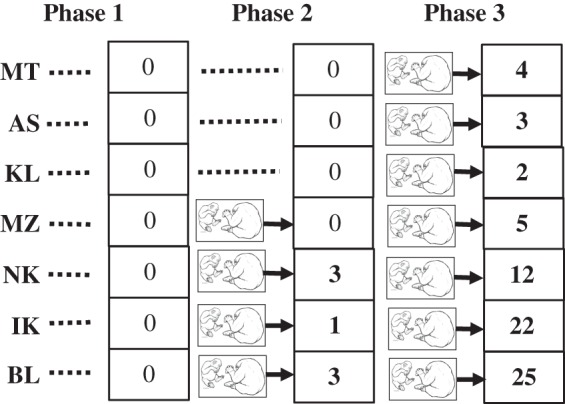
Social learning of nut-cracking by wild-born East African juvenile chimpanzees. In a baseline Phase 1, none cracked nuts when provided with raw materials; of four exposed to a conspecific model nut-cracking in Phase 2, three began to nutcrack (numbers in cells are numbers of successes) whereas no-model controls did not; when all witnessed nut-cracking in Phase 3, all began cracking, at higher frequencies for those who began in Phase 2 [33].
In the first learning phase, three of the four juveniles learned to nut-crack, whereas none of the control group did; moreover, after all participants witnessed modelled nut-cracking, all developed nut-cracking during the week (figure 1). There was thus convergent, statistically significant evidence for observational learning both between groups and within the initial control group. This result has additional value because the chimpanzees were from the wilds of East Africa, where chimpanzees do not crack nuts; that they did so in the experiment thus rules out a genetic explanation for east–west difference in nut-cracking [29], and likewise rejects environmentally constrained individual learning in favour of the cultural transmission hypothesis for the distribution of nut-cracking in the wild. New data on nut-cracking in the wild consistent with this conclusion have recently become available [35,36].
(d). Discussion: experimental studies of nut-cracking
(i). Innovation
In these social learning experiments what the model does is intentionally introduced by the experimenter, minimizing opportunities for innovation. However, an especially intriguing finding in the study of Hirata et al. [33] was that in the no-model baseline phase one chimpanzee hit a nut on the ground with a hammer stone and later did the same, twice, with a nut on an anvil, although without success. The history of these chimpanzees was well known so this was a true innovation. This is a very interesting observation because at some stage in the distant past at least one chimpanzee in the wild must be presumed to have invented nut-cracking, perhaps in West Africa where the behaviour has spread. That it is not seen elsewhere despite the presence of appropriate materials [27,28] underlines that initiating it is far from easy for chimpanzees, and makes the actions of this innovative chimpanzee a quite remarkable event to have witnessed. As the authors remark, it is consistent with the assumption that some similarly gifted wild chimpanzee invented the technique in Africa.
Other innovations have been recorded in the experimental studies, but can best be described as elaborations on what has been learned and are discussed in §2d(iii).
(ii). Variance in social learning experience required
Despite the methodological limitations in the experiments summarized above, the evidence is consistent across them and supports the hypothesis that for virtually all chimpanzees, social learning is crucial for mastery of nut-cracking. With the exception of the innovative female in the Hirata et al. study [33] (and possibly in [11]), no chimpanzee independently initiated use of a stone to hammer a nut, whereas chimpanzees that witnessed a competent nut-cracker typically went on to attempt the behaviour, mostly with eventual success. This conclusion is also consistent with the data and conclusions of field researchers who have studied the development of nut-cracking in the wild, documenting the close observation of experts by novices [14–16] and proposing it as ‘education by master–apprenticeship’ [37]. Inter-community differences in cracking techniques and raw material preferences in the wild have likewise implicated cultural transmission [35,36].
However, there are puzzling variations in the extent of observation apparently necessary for nut-cracking to develop. In the study of Hirata et al. [33], this involved several thousand bouts of cracking being observed. Age seems unlikely to explain this because the youngsters studied by Marshall-Pescini & Whiten [34] began to crack nuts successfully after much less experience. One explanation might lie in the use of relatively soft oil palm nuts in the latter study and hard macadamia nuts in the Hirata et al. study; however, macadamia nuts were also used in the study of Hayashi et al. [32] where two participants began cracking on the same day they witnessed the behaviour - although these were adults and were additionally highly ‘enculturated’ through rearing by humans. Other contributing factors may be differences in dominance and (in)tolerance between available models and learners.
The variation in observational experience apparently necessary to learn nut-cracking is certainly substantial across the studies. Field researchers have often emphasized the years of apprenticeship required to master nut-cracking, but the experiments suggest that this might reflect the initially slow development of motoric competencies, whereas in the experiments, participants already stronger and more skilled in general can sometimes learn quickly by observation, and master nut-cracking in a much shorter time-span.
(iii). Processes of social learning
A common theme across the experimental studies is that there is little sign of imitation, if by this is meant copying the actions of the nut-cracking model with high fidelity and reasonably promptly. Instead, success is typically preceded by prolonged manipulation and exploration of what can be done with the material, much of it playful and non-functional. For example, after observing a competent model cracking with hammer, nuts and anvil, young chimpanzees would spend many sessions manipulating only two of these items, or manipulating all three but in a non-functional configuration [33]. This has led to a common conclusion that the mode of social learning is best described as what Tomasello has called ‘emulation’ [38], replicating the desirable outcome observed (nuts cracked), but not necessarily using the means observed (which is what distinguishes imitation) [7,9]. Further observations consistent with an emulative form of learning include stamping the foot on a nut or hitting it with a hand, observed in early stages of development of the skill [33].
By contrast, Marshall-Pescini & Whiten [34] noted episodes in which a juvenile would closely match the form and even rhythm of a nut-cracking model's action, even when holding no hammer or nut, so this cannot be emulation and seems more consistent with motoric bodily matching. The relevant sequences have recently been re-analysed frame by frame (figure 2) and a series of times-series analyses has statistically demonstrated action matching and even synchrony between observer and model [39]. These patterns suggest that there is also a significant imitative element in the copying observed, even if of coarse-grained fidelity, mixed with exploratory experimentation and requiring a period of incubation to emerge.
Figure 2.

Entrainment of nut-cracking between observer and model. Example of frame-by-frame measures of the height of the hand shown for model (blue) and younger observer (red). Time-series analyses of such episodes confirmed matching and even synchronic entrainment of hitting actions [38].
3. Experimental studies of the cultural transmission of tool use in chimpanzees and other apes
(a). Cultural diffusion experiments
The studies above employed a ‘single generation’ design focused on the social learning of each participant from a model. However, cultural transmission is a larger phenomenon in which innovations diffuse horizontally within and/or between communities, or vertically (or obliquely) across generations. In recent years, additional experimental designs have been elaborated to address such phenomena, broadly described as diffusion experiments [40,41]. These have not so far included percussive acts, but they have included complex forms of tool use, so a brief overview is offered here. More comprehensive complementary reviews are available [17].
One important diffusion design is illustrated by its first use with chimpanzees, in which a high-ranking female in one group was taught one of two alternative stick-tool techniques to extract food from an artificial foraging device, and a similar female in a second group was taught a different technique [42]. This is ‘open diffusion’ because it is open as to whom (if any) will attend to the seeded technique and who will adopt it. In the event, each different technique spread differentially in the group seeded with it, showing that such technologies can produce incipient traditions through social learning, consistent with inferred regional variations in tool use across Africa [24,25].
An alternative design is the transmission chain. Here, individual B first watches A perform their trained action; if B then masters the task (whether using the seeded technique or not) they become the model for C, and so on along a chain of individuals. Implementing this with chimpanzees requires care in arranging pairings to be compatible rather than antagonistic, but Horner et al. [43] achieved this for chains of five to six individuals, in which the last chimpanzee (or child, in a comparative experiment) in each of two chains seeded with alternative techniques maintained the seeded option (in this case, alternative techniques to open an ‘artificial fruit’). This effectively simulated repeated inter-generational transmission that would span decades in the wild.
A combination of these two designs involved open diffusion within groups, together with a transmission chain design spanning a series of three groups [44]. Tool-use techniques that spread in one group then spread to the second allowed to watch it, and from them to the third group, with substantial fidelity to whichever of two techniques was seeded in the first group [44] (figure 3). Further diffusion studies with chimpanzees are reviewed elsewhere [17,40]. Together, they rigorously demonstrate a capacity to transmit and sustain tool-use techniques, consistent with analyses of spontaneous spreading of tool use in wild chimpanzees, that attribute the spread to social learning [45].
Figure 3.

Experimental test of cultural transmission of tool use. Alternative techniques for food extraction (a, turn-and-stab; b, lift-and-slide) were seeded in single individuals in groups B1 and B4. Graphs show differential spread of the two techniques across a first group and thence to further groups who were able to witness the methods used by neighbouring groups [43]. Bars represent the proportion of each technique shown by each chimpanzee, arranged in order of skill acquisition.
(b). Social learning processes in apes
The above experiments clearly demonstrate cultural transmission, yet are limited in their power to identify the underlying social learning processes. Elsewhere [7,13] I have described the results of experiments designed to dissect these as indicating that chimpanzees have a ‘portfolio’ of different social learning processes available, that are applied in context-dependent and largely adaptive ways. Those regarded as among the most sophisticated include imitation and emulation.
‘Emulation’ refers to learning by observation about desirable results of what others do, such as that nuts are crack-able, but generating the means to achieve this from one's own behavioural resources. In a nice example, chimpanzees shown that water could be poured from a bottle into a flask, making a peanut inside rise to the top and become accessible, when lacking a bottle themselves sucked water from their drinker and spat it into the flask to gain the nut [46]. Such emulation implies a significant degree of inventiveness.
Other research implicates imitation, in which the form of a model's actions is copied. ‘Do-as-I-do’ experiments in which apes learn to ‘Do this' through a battery of training actions and are then tested on a larger battery of novel actions has provided positive evidence of bodily imitation in orangutans and chimpanzees [47,48]. Although showing that they can imitate is not the same as showing they spontaneously do imitate, it would seem surprising if the capacity had evolved but was not used. The nut-cracking experiments described above give evidence of bodily matching consistent with this conclusion [39].
Alternatively in ‘ghost experiments' only the results of complex tool-based actions are presented, so only emulation (not imitation) is possible, and here chimpanzees have failed to master tasks based on this information alone; instead they appear to need to learn from an agent actually performing the act [49,50]. Other experiments, manipulating the perceptible evidence of causal relations in a tool-based task, have revealed a switch from relatively complete imitation of a programme or sequence of component acts in the opaque case to a more emulative response where transparency reveals that some action elements are redundant [51] (children and even human adults, by contrast, will imitate in more blanket fashion independently of such contextual variations [51,52], a phenomenon dubbed ‘over-imitation’ that we shall return to when examining experiments on humans learning to knap flint). Together this range of experiments and others reviewed more fully elsewhere [7,17,53] converge on the concept of a portfolio of context-dependent social learning processes available for learning percussive techniques.
4. Experimental studies on capacities for, and social learning of, stone tool making in Pan
The experiments described in §3 have aimed to create naturalistic tests in which the problems and tools investigated have clear analogies in the lives of wild apes. A different rationale underlies a research programme instead concerned to discover what potential for stone tool making is latent in living apes, and thus likely also in the ape ancestors from which stone tool making hominins evolved. These studies began as a collaboration between primatologists researching cognition and communication in bonobos (Pan paniscus), and archaeologists with expertise in stone tool making [21,22]. The following condensed account is based partly on these and more recent quantitative analyses [54] but also on an approximate 20 000-word, detailed narrative account by Savage-Rumbaugh (whose life's work has been with these bonobos) and a collaborating cultural anthropologist [55]. I divide this summary into a series of phases marked by apparent qualitative transitions punctuating the development of knapping in the initial learner. There have been just two individual participants, exposed to different regimes of experience, so we have no recourse to control conditions or statistical evaluation to rigorously distinguish social from personal learning. Nevertheless, there is much to be learned from this unique study and certain contingencies between events offer rather compelling evidence suggesting the learning processes at work [55].
(a). Kanzi
In a first experimental phase, an expert human (Nick Toth) knapped stone flakes using a freehand technique (hammer stone in one hand, target stone in the other) and used the flakes to cut a cord to release a flap on a container housing food rewards, while the nine-year-old, adolescent bonobo Kanzi watched. On the first day of testing, Kanzi displayed a quick facility to then select sharp flakes himself from those available, testing them orally before using them to cut the cord to obtain rewards. With verbal encouragement, he replicated the actions on stones but very crudely, clapping two stones together horizontally, somewhat weakly and ineffectively. This continued for two weeks. In a second phase, he shifted to repetitive hard hitting of this kind, creating many very small chips and flakes that could be used consecutively to cut the cord.
Several months later in a third phase, Kanzi developed more asymmetric use of the hands, holding one stone, acting as core, in his left hand, stabilized against his abdomen, and using his right hand to strike with what could now be called the hammer stone. This was more effective but appeared uncomfortable. Apparently as a result, Phase 4 saw the invention, after what the researchers described as a period of quiet inaction, of a radically different approach in which Kanzi reared up bipedally and used his right hand to smash down a stone on the hard tiled floor, producing many more useful flakes. Putt [56] has recently systematically evaluated the outcomes of three such primitive approaches, comparing them to knapping and proposing that they could plausibly have represented pre-knapping foundations of flake-making. She found that throwing a hammer stone onto a brittle core is highly efficient and bipolar flaking of a core on anvil/ground highly expedient, but knapping can create larger flakes with long useful cutting edges (investigated further in the human social learning experiments discussed in §6).
In phase 5, the researchers tried to curtail Kanzi's approach by carpeting the floor, but he peeled back a corner and continued smashing on the floor. Later on a bark floor, however, he began to throw stones onto other stones instead, thus in this respect returning to the original technique of hitting one stone on another. Over a period of days, he became progressively accurate in judging trajectories and thus more efficient in making more and larger flakes. Researchers then put all the stones into the apes' pool as another way of discouraging the existing non-knapping repertoire. Now in phase 6, a major change occurred in which Kanzi stood bipedally, and knapped. Perhaps benefitting from several months of aimed throwing of one rock at another, he was now described as using ‘glancing blows' more like Toth had originally demonstrated, with ‘considerably greater force and precision’ than earlier, and would more often ‘strike towards the edges of the core’ [55, p. 232]. Progress to glancing blows appears important; hitting at an acute angle is key to producing conchoidal fracture that experiments have shown to be among the principal ways in which expert knapping differs from novice efforts [57]. In phase 7 Kanzi developed a preference for knapping over throwing, producing and selecting his largest and most effective flakes, applied to cutting cords and slicing through leather coverings to obtain rewards.
Accordingly, Kanzi's repertoire now resembled much more closely than in his intermediate stages, the human knappers he had witnessed repeatedly through all stages of his developing skill. In those intermediate stages, it seems clear he had learned by observation that stones could be flaked and that flakes could be used to cut certain materials, but his actions were often emulative in their inventiveness, most dramatically in stone-smashing on the ground. The processes leading to his eventual closer matching to a human knapper are more ambiguous to identify; on the one hand, they may have been based on a degree of imitative matching, that required refinement of constituent skills of stone aiming, helped by the intermediate months of experience for its realization; on the other hand, the convergence of techniques may have resulted from individually learned refinements achieving a technique optimal for making the required flakes. However, the case of Kanzi's sister Panbanisha appears to implicate more direct observational learning from a model.
(b). Panbanisha
Unlike Kanzi, Panbanisha had experience in witnessing her brother knapping in a way comfortable to a bonobo, plus frequent knapping by the expert Toth, and less expert, familiar humans. However, when offered opportunities to knap flakes to use to gain rewards as Kanzi had done, she initially made weak attempts like he had initially done in his phase 1 and soon gave up, abandoning the stones. This continued for nearly a year until she saw a female expert (Kathy Schick) knapping. Later that day she switched to persevering in knapping and quickly graduated to employing ‘downward glancing blows' (again, cf. [57]) rather than the horizontal clapping that Kanzi had done, and ‘she began to rotate the core, looking for the best striking platform’; moreover ‘she did not move gradually from small to large flakes as had Kanzi, but produced a variety of sizes from the start, because she focused on the edges of the core’ [55, p. 233]. It was not clear why watching an expert female knapper appeared to have this effect (Savage-Rumbaugh offers some speculative explanations), but the result was that she appeared to make a quite rapid transition from an ineffectual phase to significant competence, rather equivalent to Kanzi's phase 7, perhaps expressing a crude imitative matching to what she had seen in the knapping of Schick and perhaps Kanzi.
(c). Recent follow-up studies: the functional potential of ape-made stone tools
Over a decade after the first suite of studies outlined above (§4a,b), the apes involved have been presented with other types of problems that could be tackled with stone tools beyond the cutting of cord and leather [23]. Experimenters split small logs longitudinally and glued them together encasing valued food items, so the logs needed to be split again to recover the rewards, somewhat analogously to splitting long bones by ancient hominins. The other problem presented was food buried under 60 cm of condensed sand covered by 20 cm of stone, so serious digging was needed for recovery. The apes were provided with tool-making materials, of which flint was preferred, but no further instruction.
These experiments do not address social learning per se; rather, they demonstrate what functional outcomes are created in apes consequent on the original acquisition of knapping skills through the social learning inferred in §4(a,b). A wide range of tools were made and used in different ways [23], 17 of which are illustrated in the article (e.g. figure 4). Thick cortical flakes knapped from core edges were described as being used as axes or cleavers, and small flakes were rotated drill-like or used as scrapers, all of these activities leaving corresponding wear patterns on the logs.
Figure 4.

Examples of flakes produced by Kanzi and used to split small logs containing food (reproduced with permission from [22]). (a) Thick cortical flake struck from edge of core. Thick edge was first used to hammer log, then sharp edge was further used as an axe to attack the split in the log. (b) Small flake used in drill-like rotational actions on splits. (c) Small flake used to scrape along split. For many more examples, see [22] and its supplementary videos.
This research programme has accordingly demonstrated in Pan (i) a considerable propensity to knap stone, making a range of tools whose functional utility for a variety of purposes is appreciated and exploited and (ii) a propensity for acquiring the skills by a complex interaction of observational learning and personal practice and invention. Some measure of such capacities therefore likely existed in our common ancestry and provided cognitive and motoric precursors that were exploited in the beginnings of knapping, once ecological circumstances created the potential for developing the new niches of early hominins [13].
It is somewhat curious that this research was done with bonobos, insofar as bonobos have not been reported to use many types of tools in the wild, whereas common chimpanzees use dozens, and more than any other non-human animal [24,25,58]; it would accordingly be valuable to extend such studies to Pan troglodytes, which might generate some different results.
Other notes of caution in interpreting this unusual set of studies is that, on the one hand, the ways in which Pan morphology differs greatly from humans', such as the relative uselessness of their small and differently placed thumbs, mean that their tool making is constrained in ways it was not in hominins as anatomy became more like our own [59–61]. On the other hand, the performances of the bonobos was greatly scaffolded by the efforts of existing human knappers to encourage their efforts over many months, on top of the extraordinary levels of human-focused enculturation that characterize the lives of these particular apes. The extent to which wild Pan of either species might display the propensities revealed here under appropriate selection pressures, remains relatively unclear.
5. Ape percussive technology in the wider context of force-based ‘power tools'
There is a tendency to focus only on the contribution made by comparative studies of nut-cracking and stone knapping to our understanding of the evolution of percussive technology. I suggest we gain a deeper understanding of the underlying behavioural and mental capacities by recognizing how these activities are embedded in broader repertoires of forceful and targeted tool use in wild chimpanzees. Whiten et al. [13] referred to these as ‘power tools'. In addition to nut-cracking they include clubbing, used against conspecifics and threats like snakes; pounding, used in such contexts as breaking into bees' nests; levering, to apply lateral force in such projects; stabbing, employing sticks with bitten ends (sometimes sharp and spear-like) to jab into tree holes to drive out and perhaps impale prey like bushbabies [62]; and puncturing, as in using a stout stick to create deep tunnels down which more delicate fishing probes are then inserted, to extract subterranean termites [63]. Some of these, like clubbing and pounding, appear universal, but others, like the stabbing and puncturing cited above, represent putative culturally transmitted local traditions. This whole variant repertoire means that chimpanzees develop a rich appreciation of the ‘folk physics' of forceful tool use and the effectiveness of certain trajectories and angles, that is likely to facilitate the fully percussive subset of actions that we see in nut-cracking.
6. Experimental studies of cultural transmission of human stone tool making
Having reviewed experimental approaches to the cultural transmission of nut-cracking, stone tool making and other forms of tool use in Pan, we now turn to related approaches in Homo. Although there is a long tradition of experimental (often described as ‘actualistic’) approaches to knapping per se, there exist only a handful of such experiments applied specifically to social learning of the skills involved, with just two substantial studies published very recently [10,11] and discussed in some detail below (§6a,b). A larger corpus of studies relevant to this topic includes those comparing novice and expert percussive skills [64], covering both nut-cracking and stone knapping [65]; identifying concordances in the knapping of novices and experts working in different sub-groups [66]; establishing the material artefact correlates of degrees of skill [67]; and longitudinal studies of skill development [68]. This larger literature is regrettably beyond the scope of this article but is referred to elsewhere in this Issue by the several papers focused on hominin knapping.
(a). The effects of different social learning opportunities on Oldowan-like flake-making
Morgan et al. [11] set up diffusion chains of the kind discussed in §2, in which participants were provided with one of five different grades of information about flaking by an expert, and then after a practice phase passed this grade of knowledge on to another novice, and so on along a chain of either five or ten individuals. The grades of information we provided were either: (i) observation of knapped flakes only, so that learners had to imagine how to create these (‘reverse engineering’); (ii) observation of making flakes, permitting imitation or emulation; (iii) ‘basic’ teaching, making the modelling clearly visible to the observer and allowing the teacher to mould their hands as necessary; (iv) gestural teaching in which gestures could be added but not vocalizations; or (v) the addition of verbal teaching (figure 5). Six main outcome measures were collected, including an index of flake quality that involved a complex computed function that took account of the length of the cutting edge and the flakes' mass and diameter, such that it effectively indexed a good cutting edge (which is what participants were asked to attempt to maximize) and penalized very small sizes.
Figure 5.

Experimental tests of social learning of flake-making (adapted from [10], with permission). Five different grades of information were tested; for definitions see text.
Given all the potential contrasts between pairs of the five conditions described above and the six available comparison measures, results were complex, but consistent findings emerged across the analyses. There was little evidence that observational learning improved success in generating a cache of good cutting flakes, but the different measures repeatedly showed graded improvements across the three different teaching conditions, with verbal teaching being particularly effective. For example, gestural teaching doubled the probability of striking producing a flake viable for cutting compared to the baseline of reverse engineering, and verbal teaching quadrupled it, whereas observation alone had no such effect. The authors acknowledge that a longer period of observation of an expert might well create a more substantial improvement associated with observation relative to reverse engineering (just 5 min learning was allowed in each generation, followed by up to 20 min of flaking), but the key conclusion drawn was that characteristics of the process of knapping, even at this simple Oldowan flake level, would have created a selection pressure for the benefits of active teaching. Given evidence that the three grades of teaching were progressively effective, such a sequence could plausibly have emerged during the Stone Age through a cumulative, step-wise process of gene–culture coevolution. These results will be discussed further in §6(b) after the other recent experimental study is outlined. It generated some apparent contradictions to the above, as well as convergent findings.
(b). The effects of different social learning opportunities on Acheulian-like biface making
Putt et al. [10] targeted the making of symmetric bifaces, characteristic of Acheulian tools. Just two conditions were compared: a verbal instruction group (equivalent to condition 5 in our study, perhaps including the gestural condition 4) and a non-verbal group (most obviously equivalent to our condition 4). Both regimes of learning coupled with practice occupied an hour a week for five weeks.
By contrast with the Oldowan study, Putt et al. found no difference between the verbal teaching and non-verbal conditions in subjective ratings of symmetry or in an objective index of symmetry, nor in a complex numerically based index of overall shape. Examples rated 8–10 on a 10-point scale of symmetry are illustrated in figure 6. The authors conclude that ‘bifacial knapping can be transmitted quite successfully in a non-verbal learning condition’ [10, p. 107]. This may seem to contradict the conclusions drawn in the Oldowan flake study about the superior effects of verbal teaching, despite the more sophisticated products examined in the biface study. However, the flake study found no differences between conditions in the morphology of individual flakes (e.g. size, amount of cutting edge); the differences were in aggregate properties such as total flake quality (the sum of the quality indices across all viable flakes made). Thus, the two studies may not conflict as much as might at first be imagined. Additionally, the symmetry that Putt et al. focus on analysing and illustrating is in plan view (figure 6), which is likely easier to attain by novice flaking than is symmetry requiring thinning in the orthogonal plane.
Figure 6.
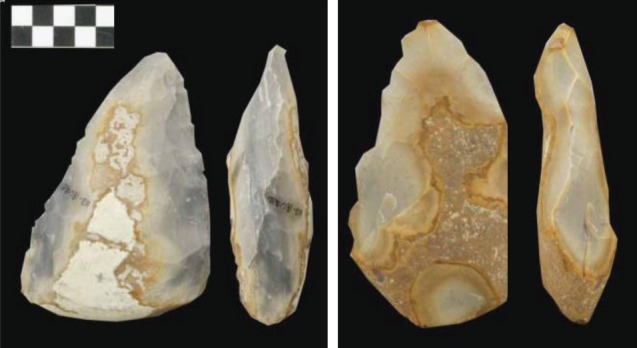
Examples of bifaces generated in social learning experiments (adapted from [9], with permission). Examples rated as of high quality; from non-verbal instruction condition on left; from verbal instruction condition on right.
Analysis of the detached flakes also revealed more consistency between the studies. The learners in the verbal biface group more accurately reproduced the approach of their tutor, setting up larger striking platforms, detaching larger flakes and so taking off fewer flakes to reach their end goal of a biface. Thus, like the Oldowan flaking study, this demonstrates the specific effects of teaching in focusing the attention of the learner on critical aspects of the skill. However, by contrast, the non-verbal learners ‘produced arguably more efficient flakes in that they were large and thin’, and moreover the non-verbal group ‘showed marked improvement in flake efficiency week to week, while the verbal group showed only gradual improvement and some backsliding’; they also generated more incipient cones in the debitage, meaning they had more failed attempts to remove flakes.
The authors attempt to makes sense of this intriguing mixture of contrasts by suggesting that ‘These results present an interesting parallel with current comparative and developmental psychology literature on the issues of emulation vs imitative learning and their role in cultural transmission among human and non-human apes', thus linking directly with our research discussed in §§2 and 3 [7,9,34]. Their interpretation is that ‘the verbal participants more faithfully imitated the instructor, to the point of over-imitation, by devoting more time to setting up ambitious platforms that, in the end, were too difficult to execute at this early a stage of learning. Thus, their task efficiency was reduced, as is evidenced by their higher frequency of missed hits and thick flakes'. By contrast, the non-verbal learners ‘focused more on emulating the process to reach the goal of a large core biface by detaching small, thin flakes until they were satisfied that their product resembled those produced by the instructor. Through their experimentation, they learned from their mistakes and improved their flake production each week’ [10, p. 107]. These interpretations are of course highly relevant to our focus on cultural transmission and identify an interesting contrast between the effects of the two conditions. However, I suggest that an alternative view of the response of the non-verbal learners is that they were engaged in imitation also, but with looser fidelity, lacking a focus on the shaping of platforms to which the attention of the other group was being drawn by verbal teaching.
That the group lacking verbal teaching achieved some quite impressive bifaces like those shown in figure 6 after less than 5 h of practice (tiny compared to the childhood apprenticeship of an early hominin Acheulian biface maker: and see [69]) raises the question of how adequate extended observational learning alone might be; however, Putt et al. [10] had no such condition, nor a baseline no-model condition. Morgan et al. [11] did have these conditions, but offered even shorter learning experiences. A question thus still remains on how adequate such inputs could be, if intertwined in long sequences of practice alternating with observation—a scenario discussed in §7.
7. Concluding discussion
The experimental studies of social learning of nut-cracking and other forms of tool use in apes provide rigorous evidence of a capacity for cultural transmission of such behaviour within and across groups. Other evidence further indicates that such transmission is achieved through a portfolio of social learning mechanisms that include the relatively sophisticated processes of emulation and imitation. In the case of apes' stone tool knapping specifically, there is also clear evidence of social learning from expert (human) models, but with so few participants involved one does not have the same precision of diagnosing the particular mechanisms involved, and can only provisionally assume they correspond to those identified in nut-cracking and other tool-use scenarios.
An important observation threaded through all of these studies of the acquisition of complex percussive technological skills is the great importance of individual learning, exploration and practice, intertwined with social learning. In figure 7, I outline a simple model that captures these observations. In this ‘helical curriculum’ model, individuals (particularly developing juveniles) are exposed repeatedly to models, who may, for example, be nut-cracking, or knapping. Each bout of observation may afford learning a little, but to assimilate more, learners need to take another ‘turn’ round the helix, indulging in practice, play and exploration, the effect of which is that when they see models later, they can now perceive more relevant aspects than they could earlier. Social and individual learning thus each progress step-wise, with interactions between them racking up the levels of sophistication in both. This may have taken on particular significance in the context of hominin knapping, as suggested by Stout et al. [70,71] remarking that ‘important parameters (e.g. kinetic energy) are not perceptually available to naïve observers … Thus the observer must begin by (incorrectly) imitating the observed gesture, checking the outcome against the predicted (desired) outcome, and then embarking on a lengthy process of goal oriented exploration and practice to (re)discover the relevant task constraints and develop corresponding internal models' [71, p. 167].
Figure 7.
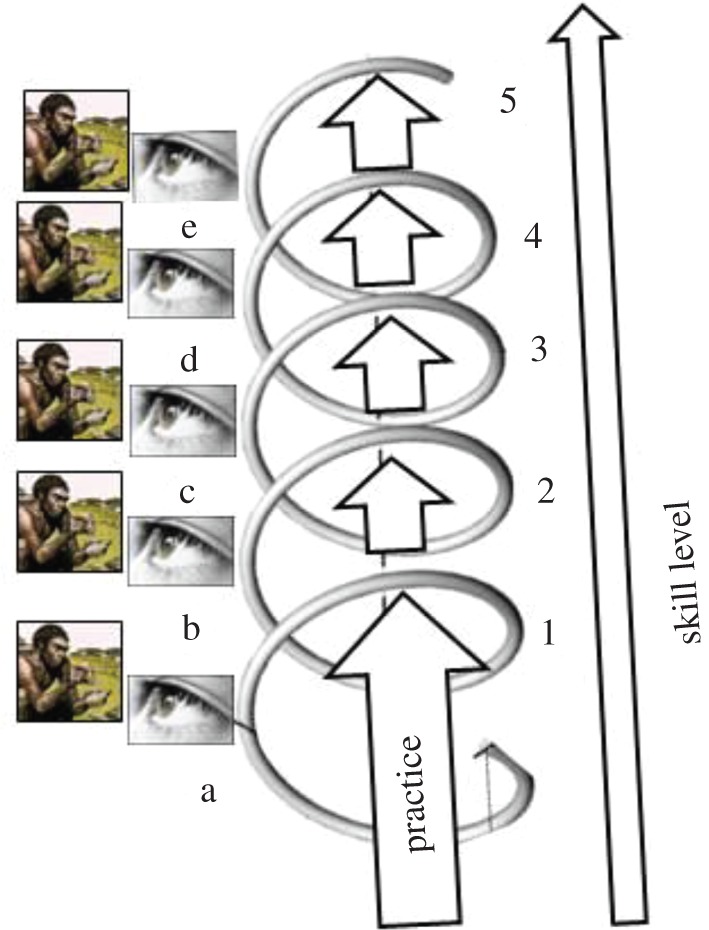
‘Helical curriculum’ model of social learning of complex skills (educationalists talk of a ‘spiral curriculum’ in which topics are revisited at increasingly higher levels, but representing the developmental time dimension creates a three-dimensional helix, not a two-dimensional spiral). At each turn of the helix, an observer watches a model and learns from them. However, in between such observational episodes is a crucial period of exploration and practice, as a result of which the learner extracts additional aspects in consecutive observational periods a–e, aspects that it could not assimilate earlier. Corresponding skill levels are thence able to rise progressively, indicated by 1–5. (Online version in colour.)
Taken together the studies reviewed in §§2–5 delineate several important features shared between humans, chimpanzees and often other great apes too, and therefore, we infer, with our ancient shared ancestors: significant capacities for cultural transmission of complex tool use, including percussive and other ‘power tools' that rely on the controlled use of force directed at very specific targets, be they nuts or stone tool cores. These would have offered important and quite elaborate foundations for the hominin Stone Age that followed.
The recent substantive experiments on human stone tool making are instead so far relatively focused on teaching, and converge in demonstrating the power and particular effects of both gestural and verbal teaching in relation to the specific demands of stone knapping, which presents difficulties for purely observational learning in its details, including effective striking trajectories, platform preparation and platform angles and edges. However, the other side of this coin is that the Putt et al. paper [10] reveals how much can be achieved without verbal input, given a modest amount of practice. The Morgan et al. [11] and Putt et al. [10] papers have illuminated much, but can be regarded largely as initial explorations and proof-of-concept studies that lay the foundations for further studies that fill the gaps they inevitably show—notably a need for more realistically longer observation and practice sequences, and for observational and other no-model control conditions coupled with these. The above discussion and figure 7 suggest that more realistic longer term experiments will repay the effort, although this may be practical for only limited sample sizes. The variance entailed in these can perhaps be ameliorated in other ways, such as by standardization in raw materials, like the use of regular bricks [58] or porcelain casts [72] as blanks.
Acknowledgements
For comments on and discussion of drafts of this paper I am most grateful to Satoshi Hirata, Mary Marzke, Bill McGrew, Tom Morgan, Shelby Putt, Itai Roffman, Sue Savage-Rumbaugh, Natalie Uomini and particularly Dietrich Stout. Thanks to Jason Zampol for chimpanzee and human illustrations in figures 1, 2 and 5.
Endnote
Recent finds appear to convincingly extend the date back to 3.3 Ma [2], but these fresh finds naturally await further similar, reinforcing discoveries and considered appraisal by the archaeological community.
Competing interests
We declare we have no competing interests.
Funding
The author was supported during the preparation of this paper by John Templeton Foundation research grant no. ID/40128 to K. Laland and A.W.
References
- 1.Semaw S, et al. 2003. 2.6-million-year-old stone tools and associated bones from OGS-6 and OGS-7, Gona, Afar, Ethiopia. J. Hum. Evol. 45, 169–177. ( 10.1016/S0047-2484(03)00093-9) [DOI] [PubMed] [Google Scholar]
- 2.Harmand S, et al. 2015. 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature 521, 310–315. ( 10.1038/nature14464) [DOI] [PubMed] [Google Scholar]
- 3.Whiten A, Hinde RA, Stringer CB, Laland KN. 2011. Culture evolves. Phil. Trans. R. Soc. B 366, 938–948. ( 10.1098/rstb.2010.0372) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Stout D. 2011. Stone toolmaking and the evolution of human culture and cognition. Phil. Trans. R. Soc. B 366, 1050–1059. ( 10.1098/rstb.2010.0369) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Gowlett JAJ. 2015. Variability in an early hominin percussive tradition: the Acheulean versus cultural variation in modern chimpanzee artefacts. Phil. Trans. R. Soc. B 370, 20140358 ( 10.1098/rstb.2014.0358) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hopper LM, Whiten A. 2012. The comparative and evolutionary psychology of social learning and culture. In The Oxford handbook of comparative evolutionary psychology (eds Vonk J, Shackelford T), pp. 451–473. Oxford, UK: Oxford University Press. [Google Scholar]
- 7.Whiten A, Horner V, Litchfield CA, Marshall-Pescini S. 2004. How do apes ape? Learn. Behav. 32, 36–52. ( 10.3758/BF03196005) [DOI] [PubMed] [Google Scholar]
- 8.Want SC, Harris PL. 2002. How do children ape? Applying concepts from the study of non-human primates to the developmental study of 'imitation' in children. Dev. Sci. 5, 1–13. ( 10.1111/1467-7687.00194) [DOI] [Google Scholar]
- 9.Whiten A, McGuigan H, Hopper LM, Marshall-Pescini S. 2009. Imitation, over-imitation, emulation and the scope of culture for child and chimpanzee. Phil. Trans. R. Soc. B 364, 2417–2428. ( 10.1098/rstb.2009.0069) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Putt SS, Woods AD, Franciscus RD. 2014. The role of verbal interaction during experimental bifacial stone tool manufacture. Lithic Technol. 39, 96–112. ( 10.1179/0197726114Z.00000000036) [DOI] [Google Scholar]
- 11.Morgan TJH, et al. 2015. Evidence for the co-evolution of hominin tool-making, teaching and language. Nat. Comm. 6, 6029 ( 10.1038/ncomms7029) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Boesch C, Boesch H. 1984. Mental map in wild chimpanzees: an analysis of hammer transports for nut-cracking. Primates 25, 160–170. ( 10.1007/BF02382388) [DOI] [Google Scholar]
- 13.Whiten A, Schick K, Toth N. 2009. The evolution and cultural transmission of percussive technology: integrating evidence from paleoanthropology and primatology. J. Hum. Evol. 57, 420–435. ( 10.1016/j.jhevol.2008.12.010) [DOI] [PubMed] [Google Scholar]
- 14.Biro D, Inoue-Nakamura N, Tonooka R, Yamakoshi G, Sousa C, Matasuzawa T. 2003. Cultural innovation and transmission of tool use in wild chimpanzees: evidence from field experiments. Anim. Cogn. 6, 213–223. ( 10.1007/s10071-003-0183-x) [DOI] [PubMed] [Google Scholar]
- 15.Inoue-Nakamura N, Matsuzawa T. 1997. Development of stone tool use by wild chimpanzees (Pan troglodytes). J. Comp. Psychol. 11, 159–173. ( 10.1037/0735-7036.111.2.159) [DOI] [PubMed] [Google Scholar]
- 16.Boesch C. 2012. Wild cultures: a comparison between chimpanzee and human cultures. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 17.Whiten A. 2011. The scope of culture in chimpanzees, humans and ancestral apes. Phil. Trans. R. Soc. B 366, 997–1007. ( 10.1098/rstb.2010.0334) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Visalberghi E, Haslam M, Spagnoletti N, Fragaszy D. 2013. Use of stone hammer tools and anvils by bearded capuchin monkeys over time and space: construction of an archaeological record of tool use. J. Archaeol. Sci. 40, 3222–3232. ( 10.1016/j.jas.2013.03.021) [DOI] [Google Scholar]
- 19.Gumert MD, Kluck M, Malaivijitnond S. 2009. The physical characteristics and usage patterns of stone axe and pounding hammers used by long-tailed macaques in the Andaman Sea region of Thailand. Am. J. Primatol. 71, 594–608. ( 10.1002/ajp.20694) [DOI] [PubMed] [Google Scholar]
- 20.Visalberghi E, Sirianni G, Fragaszy D, Boesch C. 2015. Percussive tool use by Taï Western chimpanzees and Fazenda Boa Vista bearded capuchin monkeys: a comparison. Phil. Trans. R. Soc. B 370, 20140351 ( 10.1098/rstb.2014.0351) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Toth N, Schick KD, Savage-Rumbaugh ES, Sevcik RA, Rumbaugh DM. 1993. Pan the tool-maker: investigations into the stone tool-making and tool-using capabilities of a bonobo (Pan paniscus). J. Archaeol. Sci. 20, 81–91. ( 10.1006/jasc.1993.1006) [DOI] [Google Scholar]
- 22.Schick KD, Toth N, Garufi G, Savage-Rumbaugh ES, Rumbaugh D, Sevcik R. 1999. Continuing investigations into the stone tool-making and tool-using capabilities of a bonobo (Pan paniscus). J. Archaeol. Sci. 26, 821–832. ( 10.1006/jasc.1998.0350) [DOI] [Google Scholar]
- 23.Roffman I, Savage-Rumbaugh S, Rubert-Pugh E, Ronen A, Nevo E. 2012. Stone tool production and utilization by bonobo-chimpanzees (Pan paniscus). Proc. Natl Acad. Sci. USA 109, 14 500–14 503. ( 10.1073/pnas.1212855109) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Whiten A, Goodall J, McGrew WC, Nishida T, Reynolds V, Sugiyama Y, Tutin CEG, Wrangham RW, Boesch C. 1999. Cultures in chimpanzees. Nature 399, 682–685. ( 10.1038/21415) [DOI] [PubMed] [Google Scholar]
- 25.Whiten A, Goodall J, McGrew WC, Nishida T, Reynolds V, Sugiyama Y, Tutin CEG, Wrangham RW, Boesch C. 2001. Charting cultural variation in chimpanzees. Behaviour 138, 1489–1525. ( 10.1163/156853901317367717) [DOI] [Google Scholar]
- 26.Morgan BJ, Abwe EE. 2006. Chimpanzees use stone hammers in Cameroon. Curr. Biol. 16, R632–R633. ( 10.1016/j.cub.2006.07.045) [DOI] [PubMed] [Google Scholar]
- 27.Boesch C, Marchesi P, Marchesi N, Fruth B, Joulian F. 1994. Is nutcracking in wild chimpanzees a cultural behaviour? J. Hum. Evol. 26, 325–338. ( 10.1006/jhev.1994.1020) [DOI] [Google Scholar]
- 28.McGrew WC, Ham RM, White LJT, Tutin CEG, Fernandez M. 1997. Why don't chimpanzees in Gabon crack nuts? Int. J. Primatol. 18, 353–374. ( 10.1023/A:1026382316131) [DOI] [Google Scholar]
- 29.Langergraber KE, et al. 2011. Genetic and ‘cultural’ similiarity in chimpanzees. Proc. R. Soc. B 278, 408–416. ( 10.1098/rspb.2010.1112) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Sumita K, Kitahara-Frisch J, Norikoshi K. 1985. The acquisition of stone tool use in captive chimpanzees. Primates 26, 168–181. ( 10.1007/BF02382016) [DOI] [Google Scholar]
- 31.Hannah A, McGrew WC. 1987. Chimpanzees using stones to crack open oil palm nuts in Liberia. Primates 28, 31–46. ( 10.1007/BF02382181) [DOI] [Google Scholar]
- 32.Hayashi M, Mizuno Y, Matsuzawa T. 2005. How does stone-tool use emerge? Introduction of stones and nuts to naive chimpanzees in captivity. Primates 46, 91–102. ( 10.1007/s10329-004-0110-z) [DOI] [PubMed] [Google Scholar]
- 33.Hirata S, Morimura N, Houki C. 2009. How to crack nuts: acquisition process in captive chimpanzees (Pan troglodytes) observing a model. Anim. Cogn. 12(Suppl 1), S87–S101. ( 10.1007/s10071-009-0275-3) [DOI] [PubMed] [Google Scholar]
- 34.Marshall-Pescini S, Whiten A. 2008. Social learning of nut-cracking behaviour in East African sanctuary-living chimpanzees (Pan troglodytes schweinfurthii). J. Comp. Psychol. 122, 186–194. ( 10.1037/0735-7036.122.2.186) [DOI] [PubMed] [Google Scholar]
- 35.Luncz LV, Boesch C. 2014. Tradition over trend: neighboring chimpanzee communities maintain differences in cultural behaviour despite frequent immigration of adult females. Am. J. Primatol. 76, 649–657. ( 10.1002/ajp.22259) [DOI] [PubMed] [Google Scholar]
- 36.Luncz LV, Wittig RM, Boesch C. 2015. Primate archaeology reveals cultural transmission in wild chimpanzees (Pan troglodytes verus). Phil. Trans. R. Soc. B 370, 20140348 ( 10.1098/rstb.2014.0348) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Matsuzawa T, Biro D, Humle T, Inoue-Nakamura N, Tonooka R, Yamakoshi G. 2001. Emergence of culture in wild chimpanzees: education by master-apprenticeship. In Primate origins of human cognition and behaviour (ed. Matsuzawa T.), pp. 557–574. Berlin, Germany: Springer. [Google Scholar]
- 38.Tomasello M. 1990. Cultural transmission in the tool use and communicatory signalling of chimpanzees? In ‘Language’ and intelligence in monkeys and apes: comparative developmental perspectives (eds Parker S, Gibson K), pp. 274–311. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 39.Fuhrmann D, Ravignani A, Marshall-Pescini S, Whiten A. 2014. Synchrony and motor mimicking in chimpanzee observational learning. Sci. Rep. 4, 5283 ( 10.1038/srep05383) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Whiten A, Mesoudi A. 2008. Establishing an experimental science of culture: animal social diffusion experiments. Phil. Trans. R. Soc. B 363, 3477–3488. ( 10.1098/rstb.2008.0134) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Mesoudi A, Whiten A. 2008. The multiple roles of cultural transmission experiments in understanding human cultural evolution. Phil. Trans. R. Soc. B 363, 3489–3501. ( 10.1098/rstb.2008.0129) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Whiten A, Horner V, de Waal FBM. 2005. Conformity to cultural norms of tool use in chimpanzees. Nature 437, 737–740. ( 10.1038/nature04047) [DOI] [PubMed] [Google Scholar]
- 43.Horner V, Whiten A, Flynn E, de Waal FBM. 2006. Faithful replication of foraging techniques along cultural transmission chains by chimpanzees and children. Proc. Natl Acad. Sci. USA 103, 13 878–13 883. ( 10.1073/pnas.0606015103) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Whiten A, Spiteri A, Horner V, Bonnie KE, Lambeth SP, Schapiro SJ, de Waal FBM. 2007. High-fidelity transmission of multiple traditions within and between groups of chimpanzees. Curr. Biol. 17, 1038–1043. ( 10.1016/j.cub.2007.05.031) [DOI] [PubMed] [Google Scholar]
- 45.Hobaiter C, Poiso T, Zuberbühler K, Hoppit W, Gruber T. 2014. Social network analysis shows direct evidence for social learning of tool use in wild chimpanzees. PLoS Biol. 12, e1001960. ( 10.1371/journal.pbio.1001960) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tennie C, Call J, Tomasello M. 2010. Evidence for emulation in chimpanzees in social settings using the floating peanut task. PLoS ONE 5, e10544. ( 10.1371/journal.pone.0010544) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Custance DM, Whiten A, Bard KA. 1995. Can young chimpanzees imitate arbitrary actions? Hayes and Hayes (1952) revisited. Behaviour 132, 839–858. ( 10.1163/156853995X00036) [DOI] [Google Scholar]
- 48.Call J. 2001. Body imitation in an enculturated orangutan (Pongo pygmaeus). Cybernet. Syst. 32, 97–119. ( 10.1080/019697201300001821) [DOI] [Google Scholar]
- 49.Hopper LM, Lambeth SP, Schapiro SJ, Whiten A. 2008. Observational learning in chimpanzees and children studied through ‘ghost’ conditions. Proc. R. Soc. B 275, 835–840. ( 10.1098/rspb.2007.1542) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Hopper LM, Lambeth SP, Schapiro SJ, Whiten A. 2015. The importance of witnessed agency in chimpanzee social learning of tool use. Behav. Proc. 112, 120–129. ( 10.1016/j.beproc.2014.10.009) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Horner V, Whiten A. 2005. Causal knowledge and imitation/emulation switching in chimpanzees (Pan troglodytes) and children. Anim. Cogn. 8, 164–181. ( 10.1007/s10071-004-0239-6) [DOI] [PubMed] [Google Scholar]
- 52.McGuigan N, Makinson J, Whiten A. 2011. From over-imitation to super-copying: adults imitate causally irrelevant aspects of tool use with higher fidelity than young children. Br. J. Psychol. 102, 1–18. ( 10.1348/000712610X493115) [DOI] [PubMed] [Google Scholar]
- 53.Galef BG, Whiten A. In press The comparative psychology of social learning. In APA handbook of comparative psychology (ed. Call J.). Washington, DC: American Psychological Association. [Google Scholar]
- 54.Toth N, Schick K, Semaw S. 2006. A comparative study of the stone tool-making skills of Pan, Australopithecus, and Homo sapiens. In The Oldowan: case studies into the earliest Stone Age (eds Toth N, Schick K), pp. 155–222. Gosport, IN: Stone Age Institute Press. [Google Scholar]
- 55.Savage-Rumbaugh S, Fields WM. 2006. Rules and tools: beyond anthropomorphism. In The Oldowan: case studies into the earliest Stone Age (eds Toth N, Schick K), pp. 223–241. Gosport, IN: Stone Age Institute Press. [Google Scholar]
- 56.Putt SS. 2015. The origins of stone tool reduction and the transition to knapping: an experimental approach. J. Arch. Sci. Rep. 2, 51–60. [Google Scholar]
- 57.Geribas N, Mosquera M, Verges JM. 2010. What novice knappers have to learn to become expert stone toolmakers. J. Arch. Sci. 37, 2857–2870. ( 10.1016/j.jas.2010.06.026) [DOI] [Google Scholar]
- 58.McGrew WC. 2013. Is primate tool use special? Chimpanzee and New Caledonian crow compared. Phil. Trans. R. Soc. B 368, 20120422 ( 10.1098/rstb.2012.0422) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Marzke M. 2013. Toolmaking, hand morphology and fossil hominins. Phil. Trans. R. Soc. B 368, 20120414 ( 10.1098/rstb.2012.0414) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Skinner MM, Stephens NB, Tsegai ZJ, Foote AC, Nguyen NH, Gross T, Pahr DD, Hublin J-J, Kivell T. 2015. Human-like hand-use in Australopithecus africanus. Science 347, 395–398. ( 10.1126/science.1261735) [DOI] [PubMed] [Google Scholar]
- 61.Kivell TL. 2015. Evidence in hand: recent discoveries and the early evolution of human manual manipulation. Phil. Trans. R. Soc. B 370, 20150105 ( 10.1098/rstb.2015.0105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Pruetz JD, Bertolani P. 2007. Savanna chimpanzees, Pan troglodytes verus, hunt with tools. Curr. Biol. 17, 1–6. ( 10.1016/j.cub.2006.12.042) [DOI] [PubMed] [Google Scholar]
- 63.Sanz C, Call J, Morgan D. 2009. Design complexity in termite-fishing tools of chimpanzees (Pan troglodytes). Biol. Lett. 5, 293–296. ( 10.1098/rsbl.2008.0786) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Nonaka T, Bril B, Rein R. 2010. How do stone knappers predict and control the outcome of flaking? Implications for understanding early stone tool technology. J. Hum. Evol. 59, 155–167. ( 10.1016/j.jhevol.2010.04.006) [DOI] [PubMed] [Google Scholar]
- 65.Bril B, Smaers J, Steele J, Rein R, Nonaka T, Dietrich G, Biryukova E, Hirata S, Roux V. 2012. Functional mastery of percussive technology in nut-cracking and stone-flaking actions: experimental comparison and implications for the evolution of the human brain. Phil. Trans. R. Soc. B 367, 59–74. ( 10.1098/rstb.2011.0147) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Rein R, Nonaka T, Bril B. 2014. Movement pattern variability in stone knapping: implications for the development of percussive traditions. PLoS ONE 9, e113567. ( 10.1371/journal.pone.0113567) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Hecht EE, Gutman DA, Khreisheh N, Taylor SV, Kilner J, Faisal AA, Bradley BA, Chaminade T, Stout D. 2014. Acquisition of Paleolithic toolmaking abilities involves structural remodeling to inferior frontoparietal regions. Brain Struct. Func. 222, 2315–2331. ( 10.1007/s00429-014-07896) [DOI] [PubMed] [Google Scholar]
- 68.Stout D, Apel J, Commander J, Roberts M. 2014. Late Acheulean technology and cognition at Boxgrove, UK. J. Arch. Sci. 41, 576–590. ( 10.1016/j.jas.2013.10.001) [DOI] [Google Scholar]
- 69.Högberg A. 2008. Playing with flint: tracing a child's imitation of adult work in a lithic assemblage. J. Archaeol. Method Theory 15, 112–131. ( 10.1007/s10816-007-9050-4) [DOI] [Google Scholar]
- 70.Stout D, Passingham R, Frith C, Apel J, Chaminade T. 2011. Technology, expertise and social cognition in human evolution. Eur. J. Neurosci. 33, 1328–1338. ( 10.1111/j.1460-9568.2011.07619.x) [DOI] [PubMed] [Google Scholar]
- 71.Stout D. 2013. Neuroscience of technology. In Cultural evolution: society, technology, language, and religion (eds Richerson PJ, Christiansen M), pp. 157–173. Cambridge, MA: MIT Press. [Google Scholar]
- 72.Khreisheh NN, Davies D, Bradley BA. 2013. Extending experimental control: the use of porcelain in flaked stone experimentation. Adv. Arch. Prac. 1, 37–46. [Google Scholar]