

Abstract
Rewarding cooperation is in many ways expected behaviour from social players. However, strategies that promote antisocial behaviour are also surprisingly common, not just in human societies, but also among eusocial insects and bacteria. Examples include sanctioning of individuals who behave prosocially, or rewarding of free-riders who do not contribute to collective enterprises. We therefore study the public goods game with antisocial and prosocial pool rewarding in order to determine the potential negative consequences on the effectiveness of positive incentives to promote cooperation. Contrary to a naive expectation, we show that the ability of defectors to distribute rewards to their like does not deter public cooperation as long as cooperators are able to do the same. Even in the presence of antisocial rewarding, the spatial selection for cooperation in evolutionary social dilemmas is enhanced. Since the administration of rewards to either strategy requires a considerable degree of aggregation, cooperators can enjoy the benefits of their prosocial contributions as well as the corresponding rewards. Defectors when aggregated, on the other hand, can enjoy antisocial rewards, but due to their lack of contributions to the public good they ultimately succumb to their inherent inability to secure a sustainable future. Strategies that facilitate the aggregation of akin players, even if they seek to promote antisocial behaviour, thus always enhance the long-term benefits of cooperation.
Keywords: cooperation, evolutionary games, rewarding, network reciprocity, social dilemmas
1. Introduction
The sustainability of modern human societies relies on cooperation among unrelated individuals [1]. Situations that require cooperative behaviour for socially beneficial outcomes abound, and range from taxpaying and voting to neighbourhood watch, recycling and climate change mitigation [2–6]. The crux of the problem lies in the fact that, while cooperation leads to group-beneficial outcomes, it is jeopardized by selfish incentives to free-ride on the contributions of others. Excessive short-term benefits to individuals who act as selfish maximizers create systemic risks that may nullify the long-term benefits of cooperation and lead to the tragedy of the commons [7]. Fortunately, we have strong predispositions to behave morally even when this is in conflict with our material interests [8]. The innate human drive to act prosocially is a product of our evolution as a species, as well as our unique capacity to internalize norms of social behaviour [9,10]. Yet it is also important to note that impaired recognition and absent cognitive skills are likewise potential triggers of antisocial rewarding, in particular since under such circumstances, the donor of the reward is likely to be unable to distinguish between cheaters and cooperators. As such, the concepts of mutualism and second-order free-riding are by no means limited to human societies, but apply just as well to certain eusocial insects, as well as to bacterial societies [11].
Despite favourable predispositions, however, cooperation is often subject to both positive and negative incentives [12–16]. Positive incentives typically entail rewards for behaving prosocially [17–21], while negative incentives typically entail punishing free-riding [22–30]. However, just as public cooperation incurs a cost for the well-being of the common good, so does the provisioning of rewards or sanctions incur a cost for the benefit or harm of the recipients. Individuals that abstain from dispensing such incentives therefore become second-order free-riders [31], and they are widely believed to be among the biggest impediments to the evolutionary stability of rewarding and punishing [32–36].
In addition to being costly, the success of positive and negative incentives is challenged by the fact that they can be applied to promote antisocial behaviour. Antisocial punishment—that is, the sanctioning of group members who behave prosocially—is widespread across human societies [37]. Moreover, antisocial rewarding is present in various interspecific social systems, where the host often rewards the parasitic species of a symbiont [38]. This phenomenon is due to the inability of the donor to distinguish defectors and cooperators. Recent theoretical work also indicates that antisocial punishment can prevent the coevolution of punishment and cooperation [39], just as antisocial rewarding can lead to the breakdown of cooperation if the latter is contingent on pool rewarding [40]. In theory, the resolution of such social traps involves rather complex set-ups, entailing the ability of second-order sanctioning, elevated levels of effectiveness of prosocial incentives in comparison with antisocial incentives, or the decreased ability to dispense antisocial incentives due to the limited production of public goods in environments with low levels of cooperation.
Here, we study what happens if both competing strategies are able to invest into a rewarding pool to support akin players. How does such a strategy-neutral intervention influence the evolutionary outcome of a public goods game? We consider a four-strategy game, where the traditional cooperators and defectors are joined by rewarding cooperators and rewarding defectors. Rewarding cooperators reward other rewarding cooperators, while rewarding defectors reward other rewarding defectors, thus representing prosocial and antisocial pool rewarding, respectively. Note that our set-up differs slightly from a recently studied model where rewarding players could be used directly by non-rewarding competitors [40]. In our case, we focus on the impact of the strategy-neutral intervention in the form of pool rewarding. In addition to the well-mixed game, we mainly study the game in a structured population, where everybody does not interact with everybody else, and the interactions that do exist are not random [41–43]. The importance of structured populations for the outcome of evolutionary social dilemmas was reported first by Nowak & May [44], and today the positive effects of spatial structure on the evolution of cooperation are well known as network reciprocity [45,46]. Several recent reviews are devoted to evolutionary games in structured populations [47–53].
The consideration of prosocial and antisocial pool rewarding in structured populations is thus an important step that promises to elevate our understanding of the impact of strategies that aim to promote antisocial behaviour in evolutionary games. As we will show, antisocial rewarding does not hinder the evolution of cooperation from a random state in structured populations, and in conjunction with prosocial rewarding, it still has positive consequences in that it promotes the spatial selection for cooperation in evolutionary social dilemmas. This counterintuitive outcome can be understood through pattern formation that facilitates the aggregation of players who adopt the same strategies, which in turn helps to reveal the long-term benefits of cooperation in structured populations.
2. Material and methods
The public goods game is a stylized model of situations that require cooperation to achieve socially beneficial outcomes despite obvious incentives to free-ride on the efforts of others. We suppose that players form groups of size G = 5, where they either contribute c = 1 or nothing to the common pool. After the sum of all contributions is multiplied by the synergy factor r1 > 1, the resulting public goods are distributed equally among all the group members irrespective of their contribution to the common pool. In parallel to this traditional version of the public goods game entailing cooperators (C) and defectors (D), two additional strategies run an independent pool rewarding scheme. These are rewarding cooperators (RC) and rewarding defectors (RD), who essentially establish a union-like support to aid akin players. Accordingly, rewarding cooperators contribute c = 1 to the prosocial rewarding pool. The sum of all contributions in this pool is subsequently multiplied by the synergy factor r2 > 1, and the resulting amount is distributed equally among all RC players in the group. Likewise, at each instance of the public goods game, all rewarding defectors contribute c = 1 to the antisocial rewarding pool. The sum of all contributions in this pool is subsequently multiplied by the same synergy factor r2 > 1 that applies to the prosocial rewarding pool, and the resulting amount is distributed equally among all RD players in the group. We are thus focusing on the consequences of union-like support to akin players, without considering second-order free-riding. It is therefore important that we consider strategy-neutral pool rewarding in that individual contributions to the prosocial and the antisocial rewarding pool are the same (c = 1), as is the multiplication factor r2 that is subsequently applied. Otherwise, if an obvious disadvantage would be given to either the prosocial or the antisocial rewarding pool, the outcome of the game would become predictable. We also emphasize that, in order to consider the synergistic consequence of mutual efforts and to avoid self-rewarding of a lonely player [54], we always apply r2 = 1 if only a single individual contributed to the rewarding pool.
In addition to the well-mixed version of the game, we primarily consider the spatial game. We emphasize that the importance of a structured population is not restricted to human societies, but applies just as well to bacterial societies, where the interaction range is typically limited, especially in biofilms and in vitro experiments [55,56]. Biological mechanisms that are responsible for the population being structured rather than well mixed typically include limited mobility, time and energy constraints, as well as cognitive preferences in humans and higher mammals. In the corresponding model, the public goods game is staged on a square lattice with periodic boundary conditions where L2 players are arranged into overlapping groups of size G = 5, such that everyone is connected to its G − 1 nearest neighbours. Accordingly, each individual belongs to 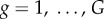 different groups. The square lattice is the simplest of networks that allows us to take into account the fact that the interactions among humans are inherently structured rather than well-mixed or random. Despite its simplicity, however, there exists ample evidence in support of the fact that the square lattice suffices to reveal all the feasible evolutionary outcomes for games that are governed by group interactions [57,58], and also that these outcomes are qualitatively independent of the details of the interaction structure [51]. As an alternative, and to explore the robustness of our findings, we nevertheless also consider regular small-world networks, where a fraction Q of all links is randomly rewired once before the start of the game [59].
different groups. The square lattice is the simplest of networks that allows us to take into account the fact that the interactions among humans are inherently structured rather than well-mixed or random. Despite its simplicity, however, there exists ample evidence in support of the fact that the square lattice suffices to reveal all the feasible evolutionary outcomes for games that are governed by group interactions [57,58], and also that these outcomes are qualitatively independent of the details of the interaction structure [51]. As an alternative, and to explore the robustness of our findings, we nevertheless also consider regular small-world networks, where a fraction Q of all links is randomly rewired once before the start of the game [59].
The considered evolutionary game in a structured population is studied by means of Monte Carlo simulations, which are carried out as follows. Initially, each player on site x is designated as a cooperator, defector, rewarding cooperator or rewarding defector with equal probability. Next, the following elementary steps are iterated repeatedly until a stationary solution is obtained. A randomly selected player x plays the public goods game with its G − 1 partners as a member of all the 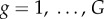 groups, whereby its overall pay-off
groups, whereby its overall pay-off 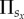 is thus the sum of all the pay-offs
is thus the sum of all the pay-offs 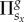 acquired in each individual group, as described in the preceding subsection. Next, player x chooses one of its nearest neighbours at random, and the chosen co-player y also acquires its pay-off
acquired in each individual group, as described in the preceding subsection. Next, player x chooses one of its nearest neighbours at random, and the chosen co-player y also acquires its pay-off 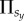 in the same way. Finally, player x enforces its strategy
in the same way. Finally, player x enforces its strategy 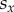 onto player y with a probability given by the Fermi function
onto player y with a probability given by the Fermi function 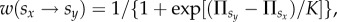 where K = 0.5 quantifies the uncertainty by strategy adoptions [57], implying that better-performing players are readily adopted, although it is not impossible to adopt the strategy of a player performing worse. Such errors in decision-making can be attributed to mistakes and external influences that adversely affect the evaluation of the opponent. Each full Monte Carlo step (MCS) gives a chance to every player to enforce its strategy onto one of the neighbours once on average.
where K = 0.5 quantifies the uncertainty by strategy adoptions [57], implying that better-performing players are readily adopted, although it is not impossible to adopt the strategy of a player performing worse. Such errors in decision-making can be attributed to mistakes and external influences that adversely affect the evaluation of the opponent. Each full Monte Carlo step (MCS) gives a chance to every player to enforce its strategy onto one of the neighbours once on average.
The average fractions of cooperators (fC), defectors (fD), rewarding cooperators (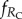 ) and rewarding defectors (
) and rewarding defectors (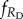 ) on the square lattice were determined in the stationary state after a sufficiently long relaxation time. Depending on the proximity to phase transition points and the typical size of emerging spatial patterns, the linear system size was varied from L = 400 to 1200, and the relaxation time was varied from 104 to 105 MCS to ensure that the statistical error is comparable with the line thickness in the figures.
) on the square lattice were determined in the stationary state after a sufficiently long relaxation time. Depending on the proximity to phase transition points and the typical size of emerging spatial patterns, the linear system size was varied from L = 400 to 1200, and the relaxation time was varied from 104 to 105 MCS to ensure that the statistical error is comparable with the line thickness in the figures.
3. Results
(a). Evolution in a well-mixed population
From the pairwise comparison of strategies, it follows that pool rewarding is dominant. Accordingly, the original four-strategy game can be reduced to a two-strategy game, where the RC and RD strategies compete. Designating by 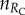 the number of rewarding cooperators, and by
the number of rewarding cooperators, and by 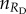 the number of rewarding defectors among other players in a group, the pay-offs of the two competing strategies are
the number of rewarding defectors among other players in a group, the pay-offs of the two competing strategies are
| 3.1 |
and
| 3.2 |
where
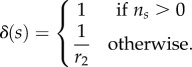 |
3.3 |
By designating the fraction of RC players as x, the corresponding replicator equation becomes
| 3.4 |
Here
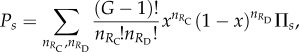 |
3.5 |
where  and
and 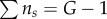 are always fulfilled.
are always fulfilled.
Starting from a random initial state, where both competing strategies are equally common (xi = 0.5), the solution of equation (3.4) indicates that the population will always terminate into the full RD state if r1 < G, and this independently of the value of r2. In other words, the introduction of strategy-neutral rewards cannot help cooperators if they are not already predominant in the initial population. Accordingly, the introduced rewards will not avert from the tragedy of the commons when the competing strategies start the evolutionary game equally strongly.
However, if RC players are somehow able to aggregate, then a significantly new situation emerges. This condition can be reached by assuming xi > 0.5, when rewarding cooperators form the majority in the initial population. In this case, the full RC and the full RD state become attractor points, but the border of their basins depends sensitively on the values of xi. This effect is illustrated in figure 1, where we have plotted the border of the two stable solutions on the r1−r2 parameter plane.
Figure 1.

In a well-mixed population, when both competing strategies are initially equally common, the extinction of rewarding cooperators is unavoidable if r1 < G, independently of the value of r2. However, if RC players are initially in the majority, then a new stable state emerges, where rewarding cooperators are the only players remaining in the population. In this bistable case, the border of the attractive basin depends sensitively on the initial fraction xi of RC players. Lines in the figure show the border of the two basins, as obtained for xi = 0.51, 0.6, 0.7, 0.8 and 0.9, from top to bottom on the r1−r2 parameter plane. The bottom-most dashed grey line shows the border in the limiting case, when there is an infinitesimally small minority of rewarding defectors initially present in the well-mixed population. (Online version in colour.)
(b). Evolution in a structured population
The lesson learned from the preceding subsection is that rewarding cooperators should initially constitute the majority of the population to survive. Otherwise, if their strength in numbers is absent, rewarding defectors inevitably take over. In a structured population, however, this special initial condition can spontaneously emerge locally, during the course of evolution, without there being an obvious advantage given to rewarding cooperators at the outset. The fundamental question then is whether such a positive local solution is viable and able to spread across the whole population, or rather if it is unstable and folds back to the defector-dominated state. To clarify this, we perform systematic Monte Carlo simulations to obtain the phase diagram for the whole r1−r2 parameter plane, as shown in figure 2. Before addressing the details, we emphasize that the reported stationary states are highly stable and fully independent of the initial conditions, which is a fundamental difference from the well-mixed solutions we have reported above. Starting with the r2 = 1 line, which implies the absence of pool rewarding, we note that cooperators survive only if the critical value of r1 is 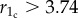 [57]. The fact that this value is still lower than the group size G = 5, which would be the threshold in a well-mixed population, is due to network reciprocity. The latter enables cooperators to form compact clusters and so protect themselves against being wiped out by defectors [44]. Taking this as a reference value, we can appreciate at a glance that, even in the presence of antisocial rewarding, prosocial rewarding still promotes the evolution of cooperation. However, neither defectors (D) nor cooperators (C) who abstain from pool rewarding can survive if r2 > 1. Indeed, as in the well-mixed case, only rewarding defectors (RD) and rewarding cooperators (RC) remain in the stationary state, depending on the value of r1 and r2. This outcome can be understood since players that do engage in pool rewarding collect pay-offs that exceed their initial contributions to the rewarding pool.
[57]. The fact that this value is still lower than the group size G = 5, which would be the threshold in a well-mixed population, is due to network reciprocity. The latter enables cooperators to form compact clusters and so protect themselves against being wiped out by defectors [44]. Taking this as a reference value, we can appreciate at a glance that, even in the presence of antisocial rewarding, prosocial rewarding still promotes the evolution of cooperation. However, neither defectors (D) nor cooperators (C) who abstain from pool rewarding can survive if r2 > 1. Indeed, as in the well-mixed case, only rewarding defectors (RD) and rewarding cooperators (RC) remain in the stationary state, depending on the value of r1 and r2. This outcome can be understood since players that do engage in pool rewarding collect pay-offs that exceed their initial contributions to the rewarding pool.
Figure 2.
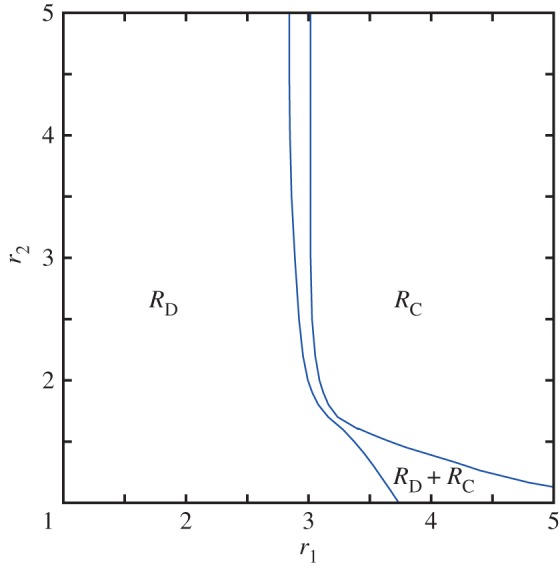
Phase diagram of the studied spatial public goods game, demonstrating that the presence of antisocial rewarding does not hinder prosocial rewarding to promote cooperation. Depicted are strategies that remain on the square lattice after sufficiently long relaxation times as a function of the multiplication factor for the public goods pool r1 and the multiplication factor for the antisocial and prosocial rewarding pool r2. Solid blue lines denote continuous phase transitions. Neither cooperators (C) nor defectors (D) who abstain from participating in pool rewarding are able to survive in the stationary state. Instead, for low values of r1 rewarding defectors (RD) dominate, while for sufficiently high values of r1 and r2 rewarding cooperators (RC) prevail. In-between is a rather narrow two-strategy RD + RC phase, where both rewarding strategies coexist. Interestingly, for example at r1 = 3.5, increasing solely the value of r2 can lead the population from a pure RD to a pure RC phase, thus indicating clearly that rewarding, even if applied to both strategies, still promotes cooperation. (Online version in colour.)
In terms of the relation between RD and RC players, it is interesting to note that the introduction of strategy-neutral pool rewarding unambiguously supports the cooperative strategy. In particular, as we increase the value of r2, and thus increase also the efficiency of rewarding, the critical value of r1 where RC players are able to survive decreases steadily. Also decreasing is the r1 threshold for complete dominance of the RD strategy. At specific values of r1, for example at r1 = 3.5, it is even possible to go from the pure RD phase to the pure RC phase solely by increasing the value of r2. Thus, indeed, even if the prosocial pool-rewarding scheme is accompanied by an equally effective antisocial pool-rewarding scheme, in structured populations the evolution of cooperation from a neutral or even from an adverse initial state is still promoted well past the boundaries imposed by network reciprocity alone.
These results are different from those obtained with random initial conditions in well-mixed populations, and they are likely to appear contradictory because there is no obvious advantage given to cooperators over defectors as the value of r2 increases. In fact, defectors benefit just as much given that they run an identical pool rewarding scheme as cooperators. So why is the evolution of cooperation promoted? The answer is rooted in the possible aggregation of cooperators, which can easily emerge spontaneously in a structured population. It is therefore instructive to monitor the evolution of the spatial distribution of strategies over time, as obtained for different values of r2. Results are presented in figure 3, where for clarity we have used a prepared initial state with only a stripe of rewarding cooperators (blue) and rewarding defectors (pale red) initially present, as illustrated in figure 3f. In all cases, the synergy factor for the main public goods game was set to r1 = 3.8.
Figure 3.

Evolution of the spatial distribution of strategies over time reveals that, even in the presence of equally effective antisocial rewarding, prosocial rewarding promotes the spatial selection for cooperation in the studied public goods game. Depicted are snapshots of the square lattice over time from left to right, as obtained for (a–e) r2 = 1, (f–j) r2 = 1.3 and (k–o) r2 = 2. For clarity, we have used a prepared initial state for all cases with only a stripe of rewarding cooperators (blue) and rewarding defectors (pale red) initially present in the population, as depicted in (f). It can be observed that in the absence of rewarding (a–e) the interface separating the two competing strategies is broken easily, and network reciprocity alone can ultimately sustain only small cooperative clusters. However, as the effectiveness of pool rewarding increases (f–o), the interface is strengthened, which makes the phalanx of cooperators more effective. The latter helps to reveal the benefit of aggregated cooperators in structured populations. In all three cases, the synergy factor for the main public goods game is r1 = 3.8. (Online version in colour.)
Figure 3a–e shows the evolution obtained at r2 = 1, which corresponds to the traditional, reward-free public goods game. It can be observed that the initially straight interface separating the two competing strategies disintegrates practically immediately. There is a very notable mixing of the two strategies, which ultimately helps defectors to occupy the larger part of the available space. Here, cooperators are able to survive solely due to network reciprocity, but at such a relatively small value of r1 only small cooperative clusters are sustainable. Nevertheless, we note that in a well-mixed population, defectors would wipe out all cooperators at such a small value of the synergy factor.
Snapshots depicted in figure 3f–j were obtained at r2 = 1.3, where thus both antisocial and prosocial pool rewarding mechanisms are at work. Here, the final state is still a mixed RC + RD phase (see also figure 2), but the fraction of cooperators is already significantly larger than in the absence of rewarding. Larger cooperative clusters are sustainable in the stationary state, which is due to an augmented interfacial stability between competing domains. In addition to traditional network reciprocity, clearly the formation of more compact cooperative clusters is further promoted by the introduction of pool rewarding, and this despite the fact that both antisocial and prosocial rewarding mechanisms are equally strong.
If an even higher value of r2 is applied, the interface that separates RC and RD players becomes impenetrable for defectors. The two strategies do not mix at all, which maintains the phalanx of cooperators [60]. Accordingly, the latter players simply spread into the region of defectors until they dominate completely. This scenario is demonstrated in figure 3k–o, where the final stationary state is indeed a pure RC phase.
As demonstrated in figure 3f–o, the introduction of pool rewarding supports the aggregation of akin players and results in more stable interfaces between competing domains. This fact enhances the positive impact of network reciprocity further and provides an even more beneficial condition for cooperation. This favourable consequence of rewarding can be studied directly by monitoring how the width w of the mixed zone—the stripe where both strategies are present—evolves over time when the evolution starts from the prepared initial state that is depicted in figure 3f. According to the definition of the width of the mixed zone, w = 0 in figure 3j, while it becomes w = L in figure 3e and j. The inset of figure 4 shows how w increases in time for different values of r2 increasing from top to the bottom curve. Clearly, as the effectiveness of rewarding increases, the width of the mixed zone increases slower and slower. While for low values of r2 the width of the mixed zone increases until eventually it covers the whole population (see figure 3e for a demonstration), for sufficiently large values of r2 the width remains finite, saturating and never exceeding a certain threshold. This result provides quantitative evidence that the interface between the two competing strategies remains intact, and that in fact the compact phalanx of cooperators cannot be broken by defectors. This in turn directly supports the evolution of cooperation to the point where defectors are wiped out completely, and this despite the fact that they are able support each other by means of antisocial rewarding.
Figure 4.
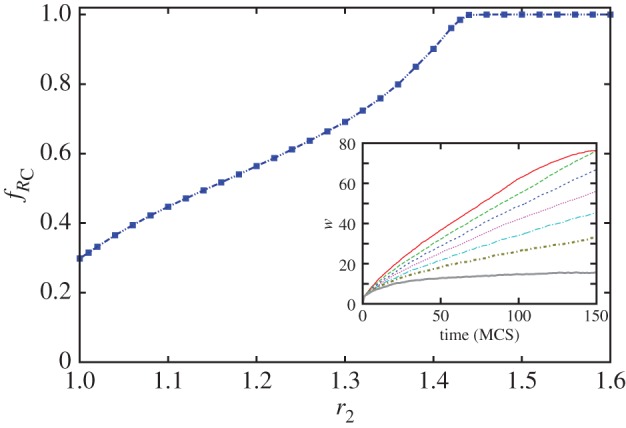
Quantitative evidence in support of enhanced spatial selection for cooperation in the studied spatial public goods game with antisocial and prosocial pool rewarding. The inset shows early stages of the evolution of the width w of the mixed zone, where both strategies are present after initially starting from a prepared initial state, as depicted in figure 3f. From top to bottom, the curves were obtained for r2 = 1, 1.1, 1.2, 1.3, 1.4, 1.5 and 2, and they correspond to the average over 100 independent runs at system size L × L = 100 × 100. In all cases, the synergy factor for the main public goods game is r1 = 3.8. The main panel shows the corresponding increase in the fraction of rewarding cooperators  as r2 increases, thus indicating that the favourable outcome is indeed due to the enhanced stability of interfaces in structured populations. This enables cooperators to dominate completely even at low values of r1, where in well-mixed populations they would not be able to survive, and where based only on network reciprocity they would fare poorly. (Online version in colour.)
as r2 increases, thus indicating that the favourable outcome is indeed due to the enhanced stability of interfaces in structured populations. This enables cooperators to dominate completely even at low values of r1, where in well-mixed populations they would not be able to survive, and where based only on network reciprocity they would fare poorly. (Online version in colour.)
Based on the results presented thus far, it is possible to provide a clear rationale why a strategy-neutral intervention, like in this case the introduction of pool rewarding (which at least in principle ought to benefit cooperators and defectors equally) is able to have such a biased impact on the final evolutionary outcome. In particular, pool rewarding yields an additional pay-off to the players only if they aggregate and form at least partly uniform groups. This is beneficial for cooperators because it also helps them to obtain a competitive pay-off from the original public goods game. In other words, the long-term benefits of cooperation come into full effect. The fate of defectors, on the other hand, is under this assumption entirely different. They can benefit from the antisocial rewarding scheme if they aggregate into uniform groups, but then they are unable to exploit the efforts of cooperators in the main public goods game. If they do not aggregate, then the benefits from antisocial rewarding become void. Either way, unlike cooperators, defectors are unable to enjoy the rewards as well as maintain a sustainable level of public goods. Ultimately, this favours the evolution of cooperation even though the intervention on the game is strategy-neutral in that it does not favour one or the other strategy directly by granting it a higher pay-off. This argument also explains why the same positive outcome is not attainable from a random initial state in well-mixed populations, where it was concluded that the possibility of antisocial rewarding utterly shatters any evolutionary benefits to cooperators that might be stemming from prosocial rewards [40]. If the interactions among players are well mixed, then of course neither cooperators nor defectors can aggregate locally, which is a fundamental condition to reveal the long-term benefits of cooperation in a collective enterprise, even if the population contains strategies that seek to actively promote antisocial behaviour.
To corroborate our main arguments further, it is instructive to consider the studied spatial public goods game on alternative interaction networks, particularly where random mixing can be controlled and adjusted deliberately. To that effect, we randomly rewire a certain fraction Q of links that constitute the originally considered square lattice, so that for small values of Q, we obtain a regular small-world network, while in the Q → 1 limit, we obtain a regular random network, as described in [59]. Essentially, we thereby allow players to expand the range of their interactions to players that are well outside their local neighbourhood. In agreement with the above-outlined arguments, this randomness in the interaction structure ought to prevent defectors from suffering the negative consequences of aggregation with their like, thus allowing them to further exploit the cooperative efforts of others while still enjoying the benefits of antisocial pool rewarding. We note that at high values of Q, it is very likely that the direct neighbours of any given player are not strongly connected. The aggregation of players with the same strategies therefore loses effect. Defectors who are members in one group can also be members in completely different groups, where perhaps the exploitation of cooperators is still possible. We test this argument quantitatively in figure 5, where we show how the critical synergy factor rc of the main public goods game for which the population arrives to the pure RD phase increases as Q increases. Indeed, as we increase the fraction of random links, more and more defectors are able to enjoy the benefits of antisocial rewarding as well as the benefits of free-riding on the cooperative efforts of others. As a countermeasure, a higher synergy factor is needed to prevent defectors from taking over. Nevertheless, even at Q = 1, the required value of r1 is still below the survival threshold of cooperators in a well-mixed population, and up to Q = 0.5, when half of all the links are randomly rewired, there are still benefits to strategy-neutral pool rewarding that go beyond those offered solely by network reciprocity. We thus conclude that antisocial rewarding does not deter public cooperation in structured populations, even if the randomness of the interaction network is high. Detrimental effects of strategies that seek to promote antisocial behaviour appear to be significantly lessened if the assumption of a well-mixed population is replaced by a structured population.
Figure 5.
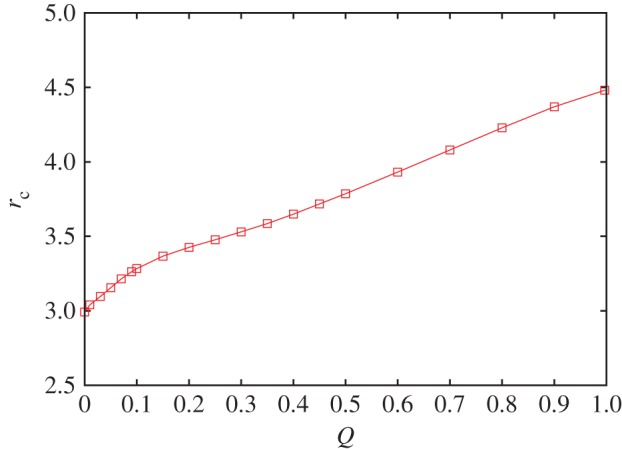
Random rewiring enables defectors to enjoy the benefits of antisocial rewarding and free-riding on the cooperative efforts of others, but relatively healthy conditions for the evolution of cooperation are maintained even if the randomness of the interaction network is high. Depicted is the critical value of r1 = rc at which the population arrives to a pure RD phase. It can be observed that rc-values increase steadily as Q increases, yet stay below the survival threshold of cooperators in a well-mixed population even at Q = 1. The synergy factor for the antisocial and prosocial pool rewarding scheme is r2 = 2. Qualitatively identical results are obtained also for other values of r2. (Online version in colour.)
4. Discussion
We have studied the joint impact of antisocial and prosocial pool rewarding in a public goods game, in particular focusing on potential detrimental effects on the evolution of public cooperation that may stem from strategies that seek to actively promote antisocial behaviour. We have been motivated by the fact that strategies that promote antisocial behaviour are surprisingly common in human societies [37] and in various interspecific social systems [38], as well as by the fact that recent research on a similar variant of the public goods game in a well-mixed population has shown that antisocial rewarding can lead to the breakdown of cooperation if the latter is contingent on pool rewarding [40]. By considering akin-like pool rewarding rather than peer rewarding, we also depart from the mainstream efforts to study the effects of rewards in structured populations [13,20,21] and join the recent [61–70] (and not so recent [71]) trend in recognizing the importance of institutions for the delivery of positive and negative incentives to cooperate in collective enterprises.
Our research reveals that, in structured populations, the detrimental effects of antisocial rewarding are significantly more benign than in well-mixed populations. Even if the interaction network lacks local structure and has many long-range links, and in this sense approaches conditions that one might hope to adequately describe by a well-mixed population, antisocial rewarding still fails to upset the effectiveness of prosocial rewarding in promoting public cooperation. We have shown that the rationale behind this rather surprising result is rooted in spatial pattern formation, and in particular in the necessity of alike strategies to aggregate if they want to enjoy the benefits of rewarding. While this condition is actually beneficial for cooperators because it helps them to obtain a competitive pay-off from the original public goods game, defectors suffer significantly because they are no longer able to free-ride on the cooperative efforts of others. The situation for defectors is thus a lot like Sophie's choice, in that they can either enjoy the benefits of antisocial rewarding or the benefits of free-riding on the public goods, but they cannot do both simultaneously—and just one of the two options is not sufficient to grant them evolutionary superiority over cooperators. Therefore, even in the presence of antisocial rewarding, prosocial rewarding still offers benefits to cooperators that go well beyond network reciprocity alone.
An interesting alternative interpretation of the studied public goods game is to consider the introduction of antisocial and prosocial pool rewarding as a strategy-neutral interference on the original rules of the social dilemma [48]. We emphasize that neither defectors nor cooperators gain an obvious evolutionary advantage from the introduction of pool rewarding—in fact, both strategies benefit exactly the same. It is therefore puzzling why, in the long run, cooperators turn out as the favoured strategy. This is in fact different from what was reported before for punishment, where available results indicate that antisocial punishment prevents the coevolution of punishment and cooperation [39], unless individuals have a reputation to lose [35], or if individuals have the freedom to leave their group and become loners [72]. Nevertheless, the results presented in our study add to the favourable aspects that positive incentives to promote cooperation have over negative incentives [17,73]. The likely unwanted consequences of punishment are well known, including failure to lead to higher total earning, damage to reputation and invitation to retaliation [17,37,74].
Summarizing, we have shown that antisocial rewarding does not necessarily deter public cooperation in structured populations, even if the randomness of the interaction network is high. This is because the delivery of rewards is contingent on the aggregation of alike strategies, which effectively prevents defectors from free-riding on the public goods. At the same time, the aggregation enhances the spatial selection for cooperation in evolutionary social dilemmas, and thus helps to expose the long-term benefits of cooperative behaviour.
Authors' contributions
A.S. and M.P. designed and performed the research, and as wrote the paper.
Competing interests
The authors declare no competing financial interests.
Funding
This research was supported by the Hungarian National Research Fund (grant no. K-101490), the Slovenian Research Agency (grant no. P5-0027) and the Deanship of Scientific Research, King Abdulaziz University (grant no. 76-130-35-HiCi).
References
- 1.Ostrom E. 1990. Governing the commons: the evolution of institutions for collective action. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 2.Wang R-W, Sun B-F, Zheng Q, Shi L, Zhu L. 2011. Asymmetric interaction and indeterminate fitness correlation between cooperative partners in the fig–fig wasp mutualism. J. R. Soc. Interface 8, 1487–1496. ( 10.1098/rsif.2011.0063) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Rand DA, Nowak MA. 2013. Human cooperation. Trends Cogn. Sci. 17, 413–425. ( 10.1016/j.tics.2013.06.003) [DOI] [PubMed] [Google Scholar]
- 4.Wu Z-X, Rong Z, Chen MZQ. 2015. Diverse roles of the reduced learning ability of players in the evolution of cooperation. EPL 110, 30002 ( 10.1209/0295-5075/110/30002) [DOI] [Google Scholar]
- 5.Pacheco JM, Vasconcelos VV, Santos FC. 2014. Climate change governance, cooperation and self-organization. Phys. Life Rev. 11, 573–586. ( 10.1016/j.plrev.2014.02.003) [DOI] [PubMed] [Google Scholar]
- 6.Rong Z, Wu Z-X, Hao D, Chen MZQ, Zhou T. 2015. Diversity of timescale promotes the maintenance of extortioners in a spatial prisoner's dilemma game. New J. Phys. 17, 033032 ( 10.1088/1367-2630/17/3/033032) [DOI] [Google Scholar]
- 7.Hardin G. 1968. The tragedy of the commons. Science 162, 1243–1248. ( 10.1126/science.162.3859.1243) [DOI] [PubMed] [Google Scholar]
- 8.Nowak MA, Highfield R. 2011. SuperCooperators: altruism, evolution, and why we need each other to succeed. New York, NY: Free Press. [Google Scholar]
- 9.Hrdy SB. 2011. Mothers and others: the evolutionary origins of mutual understanding. Cambridge, MA: Harvard University Press. [Google Scholar]
- 10.Bowles S, Gintis H. 2011. A cooperative species: human reciprocity and its evolution. Princeton, NJ: Princeton University Press. [Google Scholar]
- 11.He J-Z, Wang R-W, Li Y-T. 2014. Evolutionary stability in the asymmetric volunteer's dilemma. PLoS ONE 9, e103931 ( 10.1371/journal.pone.0103931) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Andreoni J, Harbaugh W, Vesterlund L. 2003. The carrot or the stick: rewards, punishments, and cooperation. Am. Econ. Rev. 93, 893–902. ( 10.1257/000282803322157142) [DOI] [Google Scholar]
- 13.Szolnoki A, Perc M. 2013. Correlation of positive and negative reciprocity fails to confer an evolutionary advantage: phase transitions to elementary strategies. Phys. Rev. X 3, 041021. [Google Scholar]
- 14.Okada I, Yamamoto H, Toriumi F, Sasaki T. 2015. The effect of incentives and meta-incentives on the evolution of cooperation. PLoS Comput. Biol. 11, e1004232 ( 10.1371/journal.pcbi.1004232) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rand DG, Kraft-Todd G, Gruber J. 2015. The collective benefits of feeling good and letting go: Positive emotion and (dis) inhibition interact to predict cooperative behavior. PLoS ONE 10, e0117426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.He J-Z, Wang R-W, Jensen CXJ, Li Y-T. 2015. Asymmetric interaction paired with a super-rational strategy might resolve the tragedy of the commons without requiring recognition or negotiation. Sci. Rep. 5, 7715 ( 10.1038/srep07715) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Dreber A, Rand DG, Fudenberg D, Nowak MA. 2008. Winners don't punish. Nature 452, 348–351. ( 10.1038/nature06723) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hilbe C, Sigmund K. 2010. Incentives and opportunism: from the carrot to the stick. Proc. R. Soc. B 277, 2427–2433. ( 10.1098/rspb.2010.0065) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hauert C. 2010. Replicator dynamics of reward & reputation in public goods games. J. Theor. Biol. 267, 22–28. ( 10.1016/j.jtbi.2010.08.009) [DOI] [PubMed] [Google Scholar]
- 20.Szolnoki A, Perc M. 2010. Reward and cooperation in the spatial public goods game. EPL 92, 38003 ( 10.1209/0295-5075/92/38003) [DOI] [Google Scholar]
- 21.Szolnoki A, Perc M. 2012. Evolutionary advantages of adaptive rewarding. New J. Phys. 14, 093016 ( 10.1088/1367-2630/14/9/093016) [DOI] [Google Scholar]
- 22.Fehr E, Gächter S. 2000. Cooperation and punishment in public goods experiments. Am. Econ. Rev. 90, 980–994. ( 10.1257/aer.90.4.980) [DOI] [Google Scholar]
- 23.Gardner A, West SA. 2004. Cooperation and punishment, especially in humans. Am. Nat. 164, 753–764. ( 10.1086/425623) [DOI] [PubMed] [Google Scholar]
- 24.Henrich J, et al. 2006. Costly punishment across human societies. Science 312, 1767–1770. ( 10.1126/science.1127333) [DOI] [PubMed] [Google Scholar]
- 25.Sigmund K. 2007. Punish or perish? Retaliation and collaboration among humans. Trends Ecol. Evol. 22, 593–600. ( 10.1016/j.tree.2007.06.012) [DOI] [PubMed] [Google Scholar]
- 26.Raihani NJ, Thornton A, Bshary R. 2012. Punishment and cooperation in nature. Trends Ecol. Evol. 27, 288–295. ( 10.1016/j.tree.2011.12.004) [DOI] [PubMed] [Google Scholar]
- 27.Rand DG, Nowak MA. 2011. The evolution of antisocial punishment in optional public goods games. Nat. Commun. 2, 434 ( 10.1038/ncomms1442) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Powers ST, Taylor DJ, Bryson JJ. 2012. Punishment can promote defection in group-structured populations. J. Theor. Biol. 311, 107–116. ( 10.1016/j.jtbi.2012.07.010) [DOI] [PubMed] [Google Scholar]
- 29.Hauser OP, Nowak MA, Rand DG. 2014. Punishment does not promote cooperation under exploration dynamics when anti-social punishment is possible. J. Theor. Biol. 360, 163–171. ( 10.1016/j.jtbi.2014.06.041) [DOI] [PubMed] [Google Scholar]
- 30.McCabe C, Rand DG. 2014. Coordinated punishment does not proliferate when defectors can also punish cooperators. In Antisocial behavior: etiology, genetic and environmental influences and clinical management (ed. Gallo JH.), pp. 1–14. Hauppauge, NY: Nova Publisher. [Google Scholar]
- 31.Fehr E. 2004. Don't lose your reputation. Nature 432, 449–450. ( 10.1038/432449a) [DOI] [PubMed] [Google Scholar]
- 32.Panchanathan K, Boyd R. 2004. Indirect reciprocity can stabilize cooperation without the second-order free rider problem. Nature 432, 499–502. ( 10.1038/nature02978) [DOI] [PubMed] [Google Scholar]
- 33.Hauert C, Traulsen A, Brandt H, Nowak MA, Sigmund K. 2007. Via freedom to coercion: the emergence of costly punishment. Science 316, 1905–1907. ( 10.1126/science.1141588) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Helbing D, Szolnoki A, Perc M, Szabó G. 2010. Evolutionary establishment of moral and double moral standards through spatial interactions. PLoS Comput. Biol. 6, e1000758 ( 10.1371/journal.pcbi.1000758) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hilbe C, Traulsen A. 2012. Emergence of responsible sanctions without second order free riders, antisocial punishment or spite. Sci. Rep. 2, 458 ( 10.1038/srep00458) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chen X, Szolnoki A, Perc M. 2014. Probabilistic sharing solves the problem of costly punishment. New J. Phys. 16, 083016 ( 10.1088/1367-2630/16/8/083016) [DOI] [Google Scholar]
- 37.Herrmann B, Thoni C, Gächter S. 2008. Antisocial punishment across societies. Science 319, 1362–1367. ( 10.1126/science.1153808) [DOI] [PubMed] [Google Scholar]
- 38.Wang R-W, Sun B-F, Zheng Q. 2010. Diffusive coevolution and mutualism maintenance mechanisms in a fig–fig wasp system. Ecology 91, 1308–1316. ( 10.1890/09-1446.1) [DOI] [PubMed] [Google Scholar]
- 39.Rand DG, Armao JJ, Nakamaru M, Ohtsuki H. 2010. Anti-social punishment can prevent the co-evolution of punishment and cooperation. J. Theor. Biol. 265, 624–632. ( 10.1016/j.jtbi.2010.06.010) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.dos Santos M. 2015. The evolution of anti-social rewarding and its countermeasures in public goods games. Proc. R. Soc. B 282, 20141994 ( 10.1098/rspb.2014.1994) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wasserman S, Faust K. 1994. Social network analysis. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 42.Christakis NA, Fowler JH. 2009. Connected: the surprising power of our social networks and how they shape our lives. New York, NY: Little Brown. [Google Scholar]
- 43.Apicella CL, Marlowe FW, Fowler JH, Christakis NA. 2012. Social networks and cooperation in hunter-gatherers. Nature 481, 497–501. ( 10.1038/nature10736) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Nowak MA, May RM. 1992. Evolutionary games and spatial chaos. Nature 359, 826–829. ( 10.1038/359826a0) [DOI] [Google Scholar]
- 45.Nowak MA. 2006. Five rules for the evolution of cooperation. Science 314, 1560–1563. ( 10.1126/science.1133755) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rand DG, Nowak MA, Fowler JH, Christakis NA. 2014. Static network structure can stabilize human cooperation. Proc. Natl Acad. Sci. USA 111, 17 093–17 098. ( 10.1073/pnas.1400406111) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Szabó G, Fáth G. 2007. Evolutionary games on graphs. Phys. Rep. 446, 97–216. ( 10.1016/j.physrep.2007.04.004) [DOI] [Google Scholar]
- 48.Perc M, Szolnoki A. 2010. Coevolutionary games: a mini review. BioSystems 99, 109–125. ( 10.1016/j.biosystems.2009.10.003) [DOI] [PubMed] [Google Scholar]
- 49.Roca CP, Cuesta JA, Sánchez A. 2009. Evolutionary game theory: temporal and spatial effects beyond replicator dynamics. Phys. Life Rev. 6, 208–249. ( 10.1016/j.plrev.2009.08.001) [DOI] [PubMed] [Google Scholar]
- 50.Santos FC, Pinheiro F, Lenaerts T, Pacheco JM. 2012. Role of diversity in the evolution of cooperation. J. Theor. Biol. 299, 88–96. ( 10.1016/j.jtbi.2011.09.003) [DOI] [PubMed] [Google Scholar]
- 51.Perc M, Gómez-Gardeñes J, Szolnoki A, Flora Y, Moreno LM. 2013. Evolutionary dynamics of group interactions on structured populations: a review. J. R. Soc. Interface 10, 20120997 ( 10.1098/rsif.2012.0997) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Szolnoki A, Mobilia M, Jiang L-L, Szczesny B, Rucklidge AM, Perc M. 2014. Cyclic dominance in evolutionary games: a review. J. R. Soc. Interface 11, 20140735 ( 10.1098/rsif.2014.0735) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Wang Z, Wang L, Szolnoki A, Perc M. 2014. Evolutionary games on multilayer networks: a colloquium. Eur. Phys. J. B 88, 124 ( 10.1140/epjb/e2015-60270-7) [DOI] [Google Scholar]
- 54.Brandt H, Hauert C, Sigmund K. 2006. Punishing and abstaining for public goods. Proc. Natl Acad. Sci. USA 103, 495–497. ( 10.1073/pnas.0507229103) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kerr B, Riley MA, Feldman MW, Bohannan BJM. 2002. Local dispersal promotes biodiversity in a real-life game of rock–paper–scissors. Nature 418, 171–174. ( 10.1038/nature00823) [DOI] [PubMed] [Google Scholar]
- 56.Drescher K, Nadell CD, Stone HA, Wingreen NS, Bassler BL. 2014. Solutions to the public goods dilemma in bacterial biofilms. Curr. Biol. 24, 50–55. ( 10.1016/j.cub.2013.10.030) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Szolnoki A, Perc M, Szabó G. 2009. Topology-independent impact of noise on cooperation in spatial public goods games. Phys. Rev. E 80, 056109 ( 10.1103/PhysRevE.80.056109) [DOI] [PubMed] [Google Scholar]
- 58.Szolnoki A, Perc M. 2011. Group-size effects on the evolution of cooperation in the spatial public goods game. Phys. Rev. E 84, 047102 ( 10.1103/PhysRevE.84.047102) [DOI] [PubMed] [Google Scholar]
- 59.Szabó G, Szolnoki A, Izsák R. 2004. Rock-scissors-paper game on regular small-world networks. J. Phys. A Math. Gen. 37, 2599–2609. ( 10.1088/0305-4470/37/7/006) [DOI] [Google Scholar]
- 60.Nowak MA, Sigmund K. 2000. Games on grids. In The geometry and ecological interactions: simplifying spatial complexity (eds Dieckmann U, Law R, Metz JAJ), pp. 135–150. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 61.Henrich J. 2006. Cooperation, punishment, and the evolution of human institutions. Science 312, 60–61. ( 10.1126/science.1126398) [DOI] [PubMed] [Google Scholar]
- 62.Gurerk O, Irlenbusch B, Rockenbach B. 2006. The competitive advantage of sanctioning institutions. Science 312, 108–111. ( 10.1126/science.1123633) [DOI] [PubMed] [Google Scholar]
- 63.Sigmund K, De Silva H, Traulsen A, Hauert C. 2010. Social learning promotes institutions for governing the commons. Nature 466, 861–863. ( 10.1038/nature09203) [DOI] [PubMed] [Google Scholar]
- 64.Szolnoki A, Szabó G, Perc M. 2011. Phase diagrams for the spatial public goods game with pool punishment. Phys. Rev. E 83, 036101 ( 10.1103/PhysRevE.83.036101) [DOI] [PubMed] [Google Scholar]
- 65.Traulsen A, Röhl T, Milinski M. 2012. An economic experiment reveals that humans prefer pool punishment to maintain the commons. Proc. R. Soc. B 279, 3716–3721. ( 10.1098/rspb.2012.0937) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Cressman R, Song J-W, Zhang B-Y, Tao Y. 2012. Cooperation and evolutionary dynamics in the public goods game with institutional incentives. J. Theor. Biol. 299, 144–151. ( 10.1016/j.jtbi.2011.07.030) [DOI] [PubMed] [Google Scholar]
- 67.Perc M. 2012. Sustainable institutionalized punishment requires elimination of second-order free-riders. Sci. Rep. 2, 344 ( 10.1038/srep00344) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Vasconcelos VV, Santos FC, Pacheco JM. 2013. A bottom-up institutional approach to cooperative governance of riskly commons. Nat. Clim. Change 3, 797–801. ( 10.1038/nclimate1927) [DOI] [Google Scholar]
- 69.Zhang B, Li C, De Silva H, Bednarik P, Sigmund K. 2014. The evolution of sanctioning institutions: an experimental approach to the social contract. Exp. Econ. 17, 285–303. ( 10.1007/s10683-013-9375-7) [DOI] [Google Scholar]
- 70.Sasaki T, Uchida S. 2014. Rewards and the evolution of cooperation in public good games. Biol. Lett. 10, 20130903 ( 10.1098/rsbl.2013.0903) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Yamagishi T. 1986. The provision of a sanctioning system as a public good. J. Pers. Soc. Psychol. 51, 110–116. ( 10.1037/0022-3514.51.1.110) [DOI] [Google Scholar]
- 72.Garcia J, Traulsen A. 2012. Leaving the loners alone: evolution of cooperation in the presence of antisocial punishment. J. Theor. Biol. 307, 168–173. ( 10.1016/j.jtbi.2012.05.011) [DOI] [PubMed] [Google Scholar]
- 73.Rand DG, Dreber A, Ellingsen T, Fudenberg D, Nowak MA. 2009. Positive interactions promote public cooperation. Science 325, 1272–1275. ( 10.1126/science.1177418) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Rockenbach B, Milinski M. 2011. To qualify as a social partner, humans hide severe punishment, although their observed cooperativeness is decisive. Proc. Natl Acad. Sci. USA 108, 18 307–18 312. ( 10.1073/pnas.1108996108) [DOI] [PMC free article] [PubMed] [Google Scholar]