

Embryonic development is a highly sophisticated process whereby different cell types are produced in the correct spatio-temporal order and then organized into tissues of the growing embryo. As complexity of the developing embryo increases with time, different mechanisms have evolved at different times of development for generating cellular elements and positioning them in the correct spatial and cellular context.
Formation of the vertebrate peripheral nervous system (PNS) is an elegant example of how evolution has solved the logistical problem related to producing and positioning central nervous system (CNS)-derived cells far away from any CNS structures. The PNS is complex and contains functionally distinct classes of neurons organized into ganglia at discreet locations. Peripheral nerve projections target essentially all parts of the body starting from early stages of embryonic development. In large part, PNS is derived from multipotent migratory neural crest stem cells originating in the neural plate.1 The parasympathetic part of the PNS has perhaps been the most enigmatic. These neurons are crucial for "rest-and-digest" or "feed and breed" activities especially during eating, salivation, lacrimation, urination, digestion and defecation, also including sexual arousal. Unlike sensory and sympathetic PNS, the neurons of autonomic parasympathetic nervous system are widely dispersed in the body, often located inside of the organs which they innervate and, furthermore, appear late in embryonic development (after mouse embryonic day 12.5). Since neural crest stem cells migrate and differentiate into glia and neurons earlier during development (mouse embryonic days 9–10) the discrepancy in developmental timing has left unanswered questions. It has not been clear from where and through which developmental process the parasympathetic nervous system arises.
Recently 2 independent teams unveiled the developmental process of building the parasympathetic nervous system and identified the cellular origin of the neurons.2,3 Schwann cell precursors residing in outgrowing nerves convert into neurons and, through this process, generate parasympathetic ganglia in stereotypical locations at distal sites (Fig. 1). Both groups of researchers utilized selective ablations of individual nerves to demonstrate the importance of nerve fibers in transporting SCPs to peripheral parts of the body. The data suggest that preganglionic visceral motor nerves can program the position of their postganglionic target neurons. In line with this, Dyachuk et al. observed first Schwann cell precursors committed into Ascl1+ progenitors of parasympathetic neurons in association with the specific nerves, which later course through and innervate corresponding parasympathetic ganglia. Clonal tracing and genetic ablation of molecular interactions between SCPs and the nerve clearly confirmed the nerve-dwelling Schwann cell precursor cells as a source of parasympathetic neurons.
Figure 1.
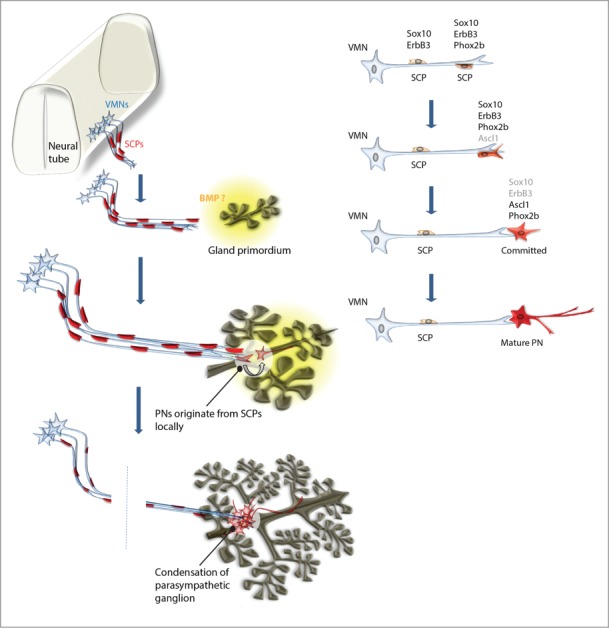
Nerves transport cellular sources of parasympathetic neurons to the locations of future ganglia. VMN - visceral motor neuron, SCP - Schwann cell precursor, PN - parasympathetic neuron.
The data adds to the growing list of cells building the embryo which arises from nerve associated multipotent SCPs, including myelinating and non-myelinating Schwann cells, endoneural fibroblasts, skin melanocytes and odontoblasts during development.4,5 The data also illuminates that SCPs are heterogeneous in their capacity to produce cell types. At least, as shown by Espinosa-Medina et al., only specific nerves which contain Phox2B+ SCPs give rise to parasympathetic neurons. This is in line with previous results demonstrating that SCPs proximal to the CNS or ganglia are different from those located more distally in nerves, as the more distal SCPs express diminishing contents of transcriptional factor Sox2, which defines their potential to convert into melanocytes.6 In a recent work Li et al. showed that some SCPs secrete CXCL12 that touches upon endothelial cells, attracting them and controlling nerve-vessel alignment. Special nerves, through this sophisticated and elegant mechanism, remodel the primary vessel network and scaffold future arteries and arterioles in the body,7 again highlighting a putative specialization of SCPs.
The discovery of multipotent stem-like progenitors dwelling in nerves may support an evolutionary hypothesis suggesting that proto-neural crest cells navigated along spinal and cranial nerves to reach their target destinations in the body of protochordate animals. Nerves, in this case, serve as primeval means of navigation since proto-neural crest may not have developed necessary pathfinding machinery for free migration. However, it is also possible that the nerve-associated niche of multipotent cells could be a later evolutionary innovation, opening for emergence of, for instance, parasympathetic neurons.
A number of questions are still left to be answered. What is the evolutionary origin of multipotent nerve-dwelling cells? What is the full competence of SCPs and how is the stem-like niche regulated? What are the mechanisms and molecules which patterns SCPs in nerves and bias them toward certain fates? Such mechanisms could involve the very neurons with which SCPs associate in nerves or may include signals from the cells of innervated developing tissues. Could multipotency be carried into adulthood, and, if so, how can it be exploited for development of regenerative therapies? We anticipate lots of interesting answers in forthcoming years.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- 1. Le Douarin NM, Dupin E. Curr Opin Genet Dev 2003; 13(5):529-36; PMID:14550420; http://dx.doi.org/ 10.1016/j.gde.2003.08.002 [DOI] [PubMed] [Google Scholar]
- 2. Dyachuk V, et al. Science 2014; 345(6192):82-87; PMID:24925909; http://dx.doi.org/ 10.1126/science.1253281 [DOI] [PubMed] [Google Scholar]
- 3. Espinosa-Medina I, et al. Science 2014; 345(6192):87-90; PMID:24925912; http://dx.doi.org/ 10.1126/science.1253286 [DOI] [PubMed] [Google Scholar]
- 4. Adameyko I, et al. Cell 2009; 139(2):366-79; PMID:19837037; http://dx.doi.org/ 10.1016/j.cell.2009.07.049 [DOI] [PubMed] [Google Scholar]
- 5. Kaukua N, et al. Nature 2014; PMID:25079316; http://dx.doi.org/ 10.1038/nature13536[Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- 6. Adameyko I, et al. Development 2012; 139(2):397-410; PMID:22186729; http://dx.doi.org/ 10.1242/dev.065581 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Li W, et al. Dev Cell 2013; 24(4):359-71; PMID:23395391; http://dx.doi.org/ 10.1016/j.devcel.2013.01.009 [DOI] [PMC free article] [PubMed] [Google Scholar]