

Abstract
Inflammatory bowel disease (IBD) is a complex, multi-factorial disease thought to arise from an inappropriate immune response to commensal bacteria in a genetically susceptible person that results in chronic, cyclical, intestinal inflammation. Dietary and environmental factors are implicated in the initiation and perpetuation of IBD; however, a singular causative agent has not been identified. As of now, the role of environmental priming or triggers in IBD onset and pathogenesis are not well understood, but these factors appear to synergize with other disease susceptibility factors. In previous work, we determined that the polysaccharide dietary additive, maltodextrin (MDX), impairs cellular anti-bacterial responses and suppresses intestinal anti-microbial defense mechanisms. In this addendum, we review potential mechanisms for dietary deregulation of intestinal homeostasis, postulate how dietary and genetic risk factors may combine to result in disease pathogenesis, and discuss these ideas in the context of recent findings related to dietary interventions for IBD.
Keywords: anti-microbial defense, Crohn's disease, diet, dietary additive, inflammatory bowel disease, intestinal homeostasis, maltodextrin, mucosal defense
Abbreviations
- AIEC
adherent-invasive Escherichia coli
- CD
Crohn's disease
- CMC
carboxymethyl cellulose
- DSS
dextran sulfate sodium
- GRAS
Generally Recognized As Safe
- FDA
Food and Drug Administration
- IBD
inflammatory bowel disease
- IBD-AID
inflammatory bowel disease-anti-inflammatory diet
- MDX
maltodextrin
- SCD
specific carbohydrate diet
- UC
ulcerative colitis
Host-Microbe Dynamics in Inflammatory Bowel Disease
Modern human health is plagued by a number of complex, chronic inflammatory states associated with altered dynamics between host anti-microbial defenses and commensal microbes. One such complex disease is inflammatory bowel disease (IBD), which is characterized by chronic, relapsing inflammation of the gastrointestinal tract. IBD is thought to arise after environmental priming or triggering of a genetically susceptible individual to initiate uncontrolled inflammation against commensal bacteria. The two main subtypes of IBD are ulcerative colitis (UC) and Crohn's disease (CD), which have similar symptoms but disparate histologic and clinical features.1 Over 160 IBD-associated genetic risk loci have been identified through meta-analyses of genome-wide association scans, and many of the genes in these loci modulate the interactions between the mucosal immune system and microbes.2 Although genetics are clearly linked to disease predisposition, carriage of these IBD risk alleles alone does not determine disease onset. This observation, combined with the recent, rapid, worldwide rise of IBD prevalence,3 suggests environmental factors are also key contributors to disease susceptibility. How diverse environmental risk factors4 contribute to IBD is not well understood; however, increasing evidence suggests that they may disrupt critical host-microbial dynamics in the gut.5
IBD patients have alterations in both composition and organization of the commensal microbiome, as well as enhanced mucosal permeability (reviewed in6). Overall, the bacterial diversity of the IBD microbiome is reduced, with notable decreases in Bacteroidetes and Firmicutes (especially in specific Clostridium species) and increases in Actinobacteria and Proteobacteria (including Escherichia coli).7 Additionally, E. coli strains with enhanced virulence have been identified in ileal CD patients.8 These strains, termed adherent-invasive E. coli (AIEC), have enhanced adhesive properties and the ability to invade and replicate within epithelial cells and macrophages. The IBD microbiome also has an altered metabolic activity, with decreases in butyrate-producing bacteria and increases in sulfate-reducing strains noted in multiple studies. The mucosal barrier of IBD patients is also more permeable than healthy individuals, leading to enhanced mucosal invasion and translocation of bacteria. This loss of mucosal barrier leads to bacterial colonization directly on the surface of the intestinal epithelium, increased bacterial translocation, and stimulation of the immune system. Of note, IBD patients exhibit anti-microbial serologic responses, with 80% of CD patients and 60% of UC patients positive for antibodies that react with enteric microbes.6 However, it is unclear how IBD risk factors contribute to these microbiome alterations and whether these microbial changes are sufficient to drive disease pathogenesis.
Maltodextrin (MDX) is a Common Food Additive That Alters Both Microbial Phenotype and Host Anti-Bacterial Defenses.
MDX is an easily digested, branched polysaccharide consisting of α(1→4) and α(1→6) linked D-glucose chains typically ranging from 3–20 glucose molecules in length generated by chemical and enzymatic processing of starch.9 Since the mid-1950s, MDX has been added to foods as a filler, thickener, texturizer, or coating agent10 and is generally recognized as safe (GRAS) by the Federal Drug Agency (FDA).11 We found in a survey of grocery store food items that ∼60% of all packaged items had “maltodextrin” or “modified (corn, wheat, etc.) starch” included in their ingredients list. Furthermore, results of a food frequency questionnaire indicated that 98.6% (210/213) of respondents routinely consume food items containing MDX, with an average consumption of 2.6 MDX-containing items per day. These surveys demonstrate that MDX is currently a ubiquitous and frequently consumed dietary polysaccharide additive in the general population.
Increasing evidence supports a modulatory relationship between commensal bacteria, host immune responses, and diet;6,12–15 therefore, we investigated the impact of MDX on bacteria, cellular anti-bacterial responses, and intestinal homeostasis. In the course of our studies, we uncovered disturbing parallels between the increasing dietary prevalence of MDX and a dramatic rise in CD incidence (Fig. 1).16 Additionally, MDX consumption by preterm piglets lead to the expansion of ileal E. coli populations17 and induced necrotizing enterocolitis in these animals, but not in fully developed pigs.18 Increased levels of E. coli and AIEC strains have been repeatedly observed in ileal CD patients, suggesting a role for E. coli in disease pathogenesis.19–21 In earlier work from our laboratory, we found that patients with ileal CD have a mucosal microbiome enriched for MDX metabolism, as compared to colonic CD patients and non-IBD controls.22 Additionally, we demonstrated that MDX has a direct effect on multiple E. coli strains, including AIEC, to enhance cellular adhesion and biofilm formation, mimicking the dense biofilms observed in the gut of CD patients21 (Fig. 2). These findings suggest that MDX consumption may promote E. coli colonization, as well as colonization of these microbes to a new region of the gastrointestinal tract.
Figure 1.
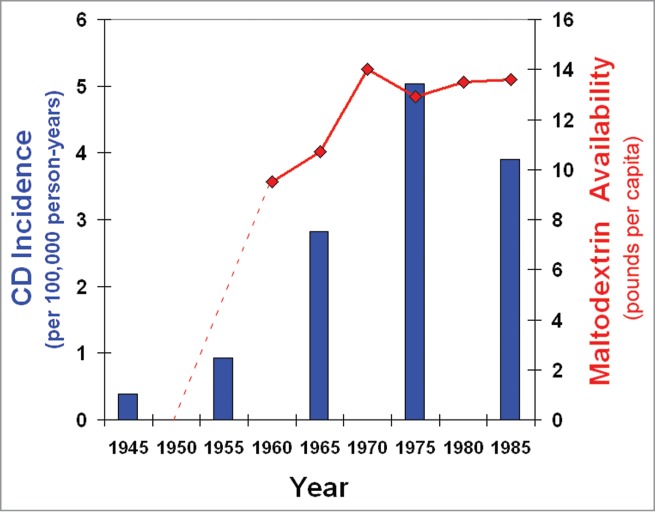
Concomitant increases in CD incidence and MDX in the American diet. The temporal trend of CD incidence in Rochester, NY16 (bars) correlates with the food availability of MDX10 (lines).
Figure 2.
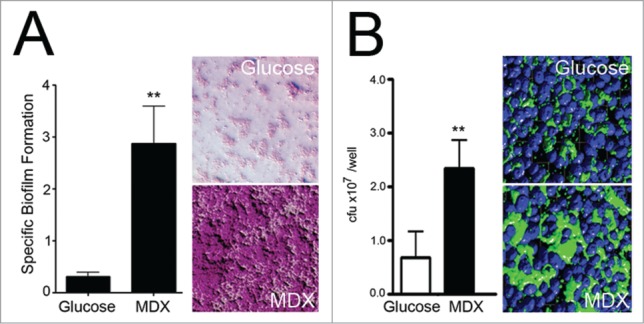
MDX enhances AIEC biofilm formation and cellular adhesion. (A) Specific biofilm formation assay of AIEC LF82 grown in minimal media supplemented with glucose or MDX for 24h (left) and images of crystal violet stained biofilms (right). (B) Adhesion of LF82 grown in glucose or MDX supplemented media to HT29 monolayers after 6h. Total adhered bacteria quantitated by colony plating (left) and immunofluorescent confocal images of wells stained for LF82 and nuclei (right). **p < 0.01. Figure modified with permission from.22
MDX consumption also influences cellular functions and shapes host-microbial interactions as demonstrated in our recent study.23 Cellular exposure to MDX in vitro impaired anti-bacterial responses, as demonstrated by the increased viability of intracellular Salmonella in macrophages and epithelial cells cultured in MDX-supplemented media. This increased bacterial viability was attributed mainly to 2 factors: (1) a diminished respiratory burst mediated by the NADPH oxidase system, and (2) alterations in bacterial trafficking to a protective niche in enlarged Rab7+ vesicles. Interestingly, genetic variants associated with CD risk have been described in NADPH complex genes24,25 and genes involved in vesicular trafficking/autophagy,2 which may affect similar processes.
One of the most striking observations in our study was a dose-dependent effect of MDX on Salmonella clearance in macrophages.23 While higher concentrations of MDX had greater effects on bacterial viability, any MDX exposure was sufficient to promote bacterial viability. This finding is significant because of the ubiquitous presence of MDX in food products and the absence of commercial usage regulation and tracking of consumption levels in the general population.
When the effects of MDX exposure were further examined in vivo using a murine Salmonella infection model, alterations in intestinal homeostasis were observed.23 In these experiments, the drinking water of mice was supplemented with an amount of MDX equivalent to levels commonly found in infant formulas (55.5 g/L) for 2 weeks. Oral Salmonella challenge of MDX-supplemented mice resulted in significantly higher cecal bacterial loads after 48 hours as compared to water-fed controls. Interestingly, this did not result in increased systemic bacterial dissemination or enhanced intestinal pathology, indicating that MDX consumption alone is not sufficient to induce disease in adult mice. These findings concur with the food industry's contention that MDX is a food additive safe for general consumption. However, when the robustness of the intestinal mucosal barrier was assessed in uninfected, MDX-supplemented mice by fluorescent in situ hybridization, commensal bacteria were aberrantly found within the mucous barrier and in direct contact with the intestinal epithelium (Fig. 3). These findings indicate that although MDX consumption does not cause intestinal disease in healthy, adult mice, it may prime the intestine for disease development through impairment of anti-bacterial cellular responses, decreases in mucosal barrier defenses, and promotion of E. coli strain adhesion. In the context of CD development, we envision that MDX consumption may (1) prime the intestine to be more sensitive to epithelial damage due to the increased proximity of bacteria to the epithelium and (2) combine with genetic risk factors that either also suppress microbial clearance or impair epithelial barrier function to result in an impaired ability to effectively and appropriately respond to bacteria breaching the intestinal barrier to restore intestinal homeostasis.
Figure 3.
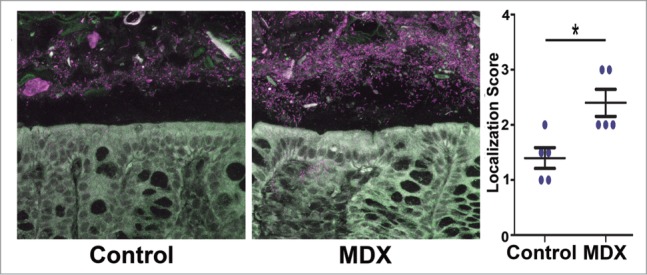
Consumption of MDX decreases the mucosal barrier of the intestine and increases the proximity of commensal bacteria to the epithelial layer. Confocal images of fluorescent in situ hybridization using a universal Eubacteria probe on uninfected proximal colon sections from mice that consumed either water or MDX-supplemented water for 2 weeks. Bacterial localization scores were assessed on a 4-point scale (1= preserved mucus layer and no bacterial contact with the epithelium, 2= bacterial penetration into mucus layer without epithelial contact, 3= some bacteria in contact with epithelium, and 4= extensive epithelial contact with the epithelium).45 Figure modified with permission from.23
Multiple Dietary Additives Alter Mucosal Homeostasis.
Extending our studies on the role of MDX in IBD development are observations that other dietary additives, such as emulsifying agents or thickeners, also have profound detrimental effects on intestinal homeostasis. These agents are used to stabilize prepared foods to extend shelf life, as well as achieve desired product viscosity and texture. Examples of dietary emulsifiers include carboxymethyl cellulose (CMC), carrageenan, xanthan gum, and MDX, which are derived from natural products and are classified as GRAS. However, carrageenan can be used to induce bacterially-driven intestinal inflammation in rodents26 and is now under re-evaluation by the FDA. Likewise, in interleukin-10-deficient mice that are genetically predisposed to colitis, CMC consumption synergizes with genetic risk to result in bacterial overgrowth and aggressive ileitis.27 Further evidence in humans demonstrates the pathogenic potential for these dietary additives when combined with other risk factors, as supplementation of infant formula with a xanthan gum-based thickener induced late-onset necrotizing enterocolitis in premature infants.28,29
Emulsifiers act directly on the mucosal barrier to decrease viscosity, permitting bacterial translocation and potentially driving inflammation.30 This is observed in the detergent-based dextran sulfate sodium (DSS) colitis model, where bacteria translocate the mucin layer soon after DSS consumption, preceding visible intestinal damage.31 In our studies, a similar alteration in mucosal integrity was observed after exposure to MDX, without subsequent damage to the intestine.23 We predict that the MDX-fed mice would be more sensitive to intestinal damaging agents (“a second hit”) to result in enhanced intestinal inflammation. As several dietary compounds have emulsifier properties, it may be important to consider the entire class of emulsifier compounds as environmental variables to examine in IBD initiation and perpetuation.
Diet-Driven Mucosal Barrier Alterations Related to IBD Pathogenesis
Alterations of mucosal barrier integrity may be an important early event in the onset of IBD.32 The colonic and small intestinal epithelial cells are overlaid with structurally distinct mucin layers formed by a combination of goblet cell secreted mucins and cell-anchored mucin glycoproteins. Both cell surface-associated mucins and secretory mucins are highly modified structures decorated with elaborate glycosylation patterns important in delineating protein function and stability. For example, 2 major constituents of mucus are mucin-2 and immunoglobulin A, both of which are heavily glycosylated, rendering them resistant to proteolytic cleavage by bacteria.33 Consumption of carbohydrates and simple sugars directly alter cellular sugar concentrations, which can alter glycosylation patterns and amounts of glycosylated proteins produced by these cells. Modifications in mucosal glycosylation patterns can lead to enhanced degradation by mucolytic bacteria, and a survival advantage for these microbes in close proximity to the epithelium. In fact, these types of changes were observed in studies of spatial organization of the microbiome in IBD patients21 and in murine diet studies.34,35 Furthermore, it has been hypothesized that bacterial infection acts as an initiating event in the development of IBD, where inflammation is unresolved despite clearance of the offending pathogen. This is supported by epidemiologic studies linking prior enteric infection with increased IBD risk36 and in a AIEC-induced murine colitis model where inflammation persisted beyond times when viable AIEC could be detected.37,38 Although we did not formally measure mucin levels, glycosylation patterns, or barrier leakiness in our study of MDX-supplemented mice, we postulate that similar defects in mucosal barrier integrity would occur in these mice and that they would be more prone to expansion of pathobionts, such as AIEC. It may be that a diet rich in MDX leads to alterations in the commensal microbiome and cellular glycome which leads to increased susceptibility to enteric pathogen infection and IBD development.
Dietary Studies: From In Vitro Observations to Clinical Efficacy
Recent studies of dietary interventions in IBD patients have demonstrated promising results in promoting clinical remission of IBD, which include the Specific Carbohydrate Diet (SCD) and the IBD-Anti-Inflammatory Diet (IBD-AID).39–41 The SCD was developed in the 1920s to treat celiac disease, but has gained popularity in recent years as a potential IBD therapeutic regimen through word-of-mouth and discussion groups on the internet. The SCD eliminates consumption of complex carbohydrates, starches, grains, and dairy, while emphasizing consumption of specific vegetables, meats, homemade yogurt, and allowing some beans and hard cheeses.39 The IBD-AID diet also restricts many of the same foods, but differs from the SCD by the phased introduction of a wider variety of foods and textures as the patient's tolerance and absorption improves.40 Additionally, the IBD-AID encourages the use of pre- and probiotics and avoidance of food irritants, which include processing agents and flavorings. Interestingly, in context of our studies, both of these dietary paradigms exclude pre-packaged and commercially processed food products, effectively eliminating MDX (and other related emulsifiers and texturizers) from their diet as well.
Although these carbohydrate restriction diets have only been tested in small clinical studies so far, the results have been striking. The one case series report of adult IBD patients (11 subjects, 8 CD and 3 UC) on the IBD-AID for 4 or more weeks demonstrated symptom reduction and the discontinuation of at least one of their prior medications by all subjects that underwent full evaluation.40 Similar dramatic effects were also observed in small clinical studies of pediatric CD patients on the SCD.39,41 One study retrospectively evaluated 7 children on the diet for 3 months and observed symptom resolution and improvement of serum and fecal measures of inflammation in all participants.41 More dramatically, a small, prospective, pilot study of pediatric CD patients demonstrated significant improvement of multiple clinical indices and mucosal analyses by capsule endoscopy after 12 weeks on the SCD and further benefits (including mucosal healing) to subjects that continued the diet for up to 52 weeks.39 Therefore, although larger clinical studies are needed to confirm these initial results, both the SCD and IBD-AID appear to have promise as a therapeutic intervention for IBD.
Cultivating a Greater Understanding of the Effect of Diet on Human Health
As food technology has advanced to produce increasingly shelf-stable products through the addition of dietary additives, we are observing a corresponding increase in chronic inflammatory diseases associated with intestinal barrier dysfunction and bacterial dysbiosis.42 Although these additives have been designated as GRAS by the FDA, more and more studies suggest that these agents may not be safe for individuals with other risk factors for chronic disease. It is also unknown whether these GRAS additives have a pathogenic threshold, as consumption of these agents is not formally measured by any standard diet questionnaire. Notable studies have demonstrated a direct effect of these compounds on mucosal barrier integrity, which translates to exacerbated intestinal inflammation or increased bacterial burdens in animal models.17,18,27 Likewise, restrictive diets that eliminate processed foods are showing clinical promise in eliminating disease symptoms and promoting mucosal healing in both adult and pediatric IBD patients.39–41 Although preliminary analyses suggest that one beneficial effect of the SCD is to increase bacterial diversity in the intestinal microbiome,43 it is still unknown which of the eliminated dietary components promote disease or their mechanism(s) of action. We postulate that dietary additives, such as MDX, are potentiators of disease and, if true, could explain the suggested efficacy of seemingly disparate diets (i.e. enteral nutrition, parenteral nutrition, gluten-free diets, elemental diets, etc.) on intestinal inflammation (reviewed in44). Uncovering these mechanisms of disease and examining how they interact with other IBD risk factors, such as genetics, would provide us with opportunities to more effectively resolve existing disease and the exciting possibility to prevent it in susceptible individuals.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Gail Cresci, Jackie Rigutto, Kelsey Rodgers, and Liz DeJuliea for their assistance with the estimates of MDX levels in products and dietary consumption.
Funding
Research in our laboratory is supported by grants from the National Institutes of Health (R01DK082437, R01DK050984, and UL1TR000439) and the Department of Defense (PR110887), the Howard Hughes Medical Institute “Med into Grad” Initiative, as well as the generosity of Gerald and Nancy Goldberg.
References
- 1. Kaser A, Zeissig S, Blumberg RS. Inflammatory bowel disease. Annu Rev Immunol 2010; 28:573-621; PMID:20192811; http://dx.doi.org/ 10.1146/annurev-immunol-030409-101225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Jostins L, Ripke S, Weersma RK, Duerr RH, McGovern DP, Hui KY, Lee JC, Schumm LP, Sharma Y, Anderson CA. et al. Host-microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature 2012; 491:119-24; PMID:23128233; http://dx.doi.org/ 10.1038/nature11582 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Molodecky NA, Soon IS, Rabi DM, Ghali WA, Ferris M, Chernoff G, Benchimol EI, Panaccione R, Ghosh S, Barkema HW., et al. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review. Gastroenterology 2012; 142:46-54 e42; quiz e30; PMID:22001864; http://dx.doi.org/ 10.1053/j.gastro.2011.10.001 [DOI] [PubMed] [Google Scholar]
- 4. Rogler G, Vavricka S. Exposome in IBD: recent insights in environmental factors that influence the onset and course of IBD. Inflamm Bowel Dis 2014; 21:400-8; PMID:25358064; http://dx.doi.org/ 10.1097/MIB.0000000000000229 [DOI] [PubMed] [Google Scholar]
- 5. Spooren CE, Pierik MJ, Zeegers MP, Feskens EJ, Masclee AA, Jonkers DM. Review article: the association of diet with onset and relapse in patients with inflammatory bowel disease. Aliment Pharmacol Ther 2013; 38:1172-87; PMID:24118051; http://dx.doi.org/ 10.1111/apt.12501 [DOI] [PubMed] [Google Scholar]
- 6. Sartor RB. Microbial influences in inflammatory bowel diseases. Gastroenterology 2006; 134:577-94; PMID:18242222; http://dx.doi.org/ 10.1053/j.gastro.2007.11.059 [DOI] [PubMed] [Google Scholar]
- 7. Walters WA, Xu Z, Knight R. Meta-analyses of human gut microbes associated with obesity and IBD. FEBS Lett 2014; 588:4223-33; PMID:25307765; http://dx.doi.org/ 10.1016/j.febslet.2014.09.039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Darfeuille-Michaud A, Boudeau J, Bulois P, Neut C, Glasser AL, Barnich N, Bringer MA, Swidsinski A, Beaugerie L, Colombel JF. High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohn's disease. Gastroenterology 2004; 127:412-21; PMID:15300573; http://dx.doi.org/ 10.1053/j.gastro.2004.04.061 [DOI] [PubMed] [Google Scholar]
- 9. Chronakis IS. On the molecular characteristics, compositional properties, and structural-functional mechanisms of maltodextrins: a review. Crit Rev Food Sci Nutr 1998; 38:599-637; PMID:9813736; http://dx.doi.org/ 10.1080/10408699891274327 [DOI] [PubMed] [Google Scholar]
- 10. Economic Research Service (ERS), US Department of Agriculture (USDA). Food Availability (Per Capita) Data System, 2010.. Retrieved from http://ers.usda.gov/data-products/food-availability-(per-capita)-data-system.aspx [Google Scholar]
- 11. FDA Title 21, volume 3, part 184, subpart B, section 184.1444. Three Code of Federal Regulations. [Google Scholar]
- 12. Brown K, DeCoffe D, Molcan E, Gibson DL. Diet-induced dysbiosis of the intestinal microbiota and the effects on immunity and disease. Nutrients 2012; 4:1095-119; PMID:23016134; http://dx.doi.org/ 10.3390/nu4081095 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Flint HJ, Scott KP, Louis P, Duncan SH. The role of the gut microbiota in nutrition and health. Nat Rev Gastroenterol Hepatol 2012; 9:577-89; PMID:22945443; http://dx.doi.org/ 10.1038/nrgastro.2012.156 [DOI] [PubMed] [Google Scholar]
- 14. Ivanov II, Littman DR. Segmented filamentous bacteria take the stage. Mucosal Immunol 2010; 3:209-12; PMID:20147894; http://dx.doi.org/ 10.1038/mi.2010.3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Kunisawa J, Kiyono H. Vitamins mediate immunological homeostasis and diseases at the surface of the body. Endocr Metab Immune Disord Drug Targets 2014; Oct 21 [Epub ahead of print]; PMID:25335990; http://dx.doi.org/ 10.2174/1871530314666141021114651 [DOI] [PubMed] [Google Scholar]
- 16. Stowe SP, Redmond SR, Stormont JM, Shah AN, Chessin LN, Segal HL, Chey WY. An epidemiologic study of inflammatory bowel disease in rochester, new york. hospital incidence. Gastroenterology 1990; 98:104-10; PMID:2293567. [DOI] [PubMed] [Google Scholar]
- 17. McDonald DE, Pethick DW, Mullan BP, Hampson DJ. Increasing viscosity of the intestinal contents alters small intestinal structure and intestinal growth, and stimulates proliferation of enterotoxigenic escherichia coli in newly-weaned pigs. Br J Nutr 2001; 86:487-98; PMID:11591236; http://dx.doi.org/ 10.1079/BJN2001416 [DOI] [PubMed] [Google Scholar]
- 18. Thymann T, Moller HK, Stoll B, Stoy AC, Buddington RK, Bering SB, Jensen BB, Olutoye OO, Siggers RH, Molbak L., et al. . Carbohydrate maldigestion induces necrotizing enterocolitis in preterm pigs. Am J Physiol Gastrointest Liver Physiol 2009; 297:G1115-25; PMID:19808655; http://dx.doi.org/ 10.1152/ajpgi.00261.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Lapaquette P, Glasser AL, Huett A, Xavier RJ, Darfeuille-Michaud A. Crohn's disease-associated adherent-invasive E. coli are selectively favoured by impaired autophagy to replicate intracellularly. Cell Microbiol 2010; 12:99-113; PMID:19747213; http://dx.doi.org/ 10.1111/j.1462-5822.2009.01381.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Martinez-Medina M, Aldeguer X, Lopez-Siles M, Gonzalez-Huix F, Lopez-Oliu C, Dahbi G, Blanco JE, Blanco J, Garcia-Gil LJ, Darfeuille-Michaud A. Molecular diversity of escherichia coli in the human gut: new ecological evidence supporting the role of adherent-invasive E. coli (AIEC) in crohn's disease. Inflamm Bowel Dis 2009; 15:872-82; PMID:19235912; http://dx.doi.org/ 10.1002/ibd.20860 [DOI] [PubMed] [Google Scholar]
- 21. Swidsinski A, Loening-Baucke V, Herber A. Mucosal flora in crohn's disease and ulcerative colitis - an overview. J Physiol Pharmacol 2009; 60 6:61-71; PMID:20224153 [PubMed] [Google Scholar]
- 22. Nickerson KP, McDonald C. Crohn's disease-associated adherent-invasive escherichia coli adhesion is enhanced by exposure to the ubiquitous dietary polysaccharide maltodextrin. PLoS One 2012; 7:e52132; PMID:23251695; http://dx.doi.org/ 10.1371/journal.pone.0052132 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Nickerson KP, Homer CR, Kessler SP, Dixon LJ, Kabi A, Gordon IO, Johnson EE, de la Motte CA, McDonald C. The dietary polysaccharide maltodextrin promotes Salmonella survival and mucosal colonization in mice. PLoS One 2014; 9:e101789; PMID:25000398; http://dx.doi.org/ 10.1371/journal.pone.0101789 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Muise AM, Xu W, Guo CH, Walters TD, Wolters VM, Fattouh R, Lam GY, Hu P, Murchie R, Sherlock M., NADPH oxidase complex and IBD candidate gene studies: identification of a rare variant in NCF2 that results in reduced binding to RAC2. Gut 2012; 61:1028-35; PMID:21900546; http://dx.doi.org/ 10.1136/gutjnl-2011-300078 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Roberts RL, Hollis-Moffatt JE, Gearry RB, Kennedy MA, Barclay ML, Merriman TR. Confirmation of association of IRGM and NCF4 with ileal crohn's disease in a population-based cohort. Genes Immun 2008; 9:561-5; PMID:18580884; http://dx.doi.org/ 10.1038/gene.2008.49 [DOI] [PubMed] [Google Scholar]
- 26. Moyana TN, Lalonde JM. Carrageenan-induced intestinal injury in the rat–a model for inflammatory bowel disease. Ann Clin Lab Sci 1990; 20:420-6; PMID:2073092. [PubMed] [Google Scholar]
- 27. Swidsinski A, Ung V, Sydora BC, Loening-Baucke V, Doerffel Y, Verstraelen H, Fedorak RN. Bacterial overgrowth and inflammation of small intestine after carboxymethylcellulose ingestion in genetically susceptible mice. Inflamm Bowel Dis 2009; 15:359-64; PMID:18844217; http://dx.doi.org/ 10.1002/ibd.20763 [DOI] [PubMed] [Google Scholar]
- 28. Beal J, Silverman B, Bellant J, Young TE, Klontz K. Late onset necrotizing enterocolitis in infants following use of a xanthan gum-containing thickening agent. J Pediatr 2012; 161:354-6; PMID:22575248; http://dx.doi.org/ 10.1016/j.jpeds.2012.03.054 [DOI] [PubMed] [Google Scholar]
- 29. Woods CW, Oliver T, Lewis K, Yang Q. Development of necrotizing enterocolitis in premature infants receiving thickened feeds using simplythick(R). J Perinatol 2012; 32:150-2; PMID:22289705; http://dx.doi.org/ 10.1038/jp.2011.105 [DOI] [PubMed] [Google Scholar]
- 30. Swidsinski A, Sydora BC, Doerffel Y, Loening-Baucke V, Vaneechoutte M, Lupicki M, Scholze J, Lochs H, Dieleman LA. Viscosity gradient within the mucus layer determines the mucosal barrier function and the spatial organization of the intestinal microbiota. Inflamm Bowel Dis 2007; 13:963-70; PMID:17455202; http://dx.doi.org/ 10.1002/ibd.20163 [DOI] [PubMed] [Google Scholar]
- 31. Johansson ME, Gustafsson JK, Sjoberg KE, Petersson J, Holm L, Sjovall H, Hansson GC. Bacteria penetrate the inner mucus layer before inflammation in the dextran sulfate colitis model. PLoS One 2010; 5:e12238; PMID:20805871; http://dx.doi.org/ 10.1371/journal.pone.0012238 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Johansson ME. Mucus layers in inflammatory bowel disease. Inflamm Bowel Dis 2014; 20:2124-31; PMID:25025717; http://dx.doi.org/ 10.1097/MIB.0000000000000117 [DOI] [PubMed] [Google Scholar]
- 33. van der Post S, Subramani DB, Backstrom M, Johansson ME, Vester-Christensen MB, Mandel U, Bennett EP, Clausen H, Dahlen G, Sroka A., Site-specific O-glycosylation on the MUC2 mucin protein inhibits cleavage by the porphyromonas gingivalis secreted cysteine protease (RgpB). J Biol Chem 2013; 288:14636-46; PMID:23546879; http://dx.doi.org/ 10.1074/jbc.M113.459479 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Mastrodonato M, Mentino D, Portincasa P, Calamita G, Liquori GE, Ferri D. High-fat diet alters the oligosaccharide chains of colon mucins in mice. Histochem Cell Biol 2014; 142:449-59; PMID:24770665; http://dx.doi.org/ 10.1007/s00418-014-1221-2 [DOI] [PubMed] [Google Scholar]
- 35. Martinez-Medina M, Denizot J, Dreux N, Robin F, Billard E, Bonnet R, Darfeuille-Michaud A, Barnich N. Western diet induces dysbiosis with increased E coli in CEABAC10 mice, alters host barrier function favouring AIEC colonisation. Gut 2014; 63:116-24; PMID:23598352; http://dx.doi.org/ 10.1136/gutjnl-2012-304119 [DOI] [PubMed] [Google Scholar]
- 36. Garcia Rodriguez LA, Ruigomez A, Panes J. Acute gastroenteritis is followed by an increased risk of inflammatory bowel disease. Gastroenterology 2006; 130:1588-94; PMID:16697722; http://dx.doi.org/ 10.1053/j.gastro.2006.02.004 [DOI] [PubMed] [Google Scholar]
- 37. Chassaing B, Koren O, Carvalho FA, Ley RE, Gewirtz AT. AIEC pathobiont instigates chronic colitis in susceptible hosts by altering microbiota composition. Gut 2014; 63:1069-80; PMID:23896971; http://dx.doi.org/ 10.1136/gutjnl-2013-304909 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Small CL, Reid-Yu SA, McPhee JB, Coombes BK. Persistent infection with crohn's disease-associated adherent-invasive escherichia coli leads to chronic inflammation and intestinal fibrosis. Nat Commun 2013; 4:1957; PMID:23748852; http://dx.doi.org/ 10.1038/ncomms2957 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Cohen SA, Gold BD, Oliva S, Lewis J, Stallworth A, Koch B, Eshee L, Mason D. Clinical and mucosal improvement with specific carbohydrate diet in pediatric crohn disease. J Pediatr Gastroenterol Nutr 2014; 59:516-21; PMID:24897165; http://dx.doi.org/ 10.1097/MPG.0000000000000449 [DOI] [PubMed] [Google Scholar]
- 40. Olendzki BC, Silverstein TD, Persuitte GM, Ma Y, Baldwin KR, Cave D. An anti-inflammatory diet as treatment for inflammatory bowel disease: a case series report. Nutr J 2014; 13:5; PMID:24428901; http://dx.doi.org/ 10.1186/1475-2891-13-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Suskind DL, Wahbeh G, Gregory N, Vendettuoli H, Christie D. Nutritional therapy in pediatric crohn disease: the specific carbohydrate diet. J Pediatr Gastroenterol Nutr 2014; 58:87-91; PMID:24048168; http://dx.doi.org/ 10.1097/MPG.0000000000000103 [DOI] [PubMed] [Google Scholar]
- 42. Cordain L, Eaton SB, Sebastian A, Mann N, Lindeberg S, Watkins BA, O'Keefe JH, Brand-Miller J. Origins and evolution of the western diet: health implications for the 21st century. Am J Clin Nutr 2005; 81:341-54; PMID:15699220. [DOI] [PubMed] [Google Scholar]
- 43. Kakodkar SM, Engen P, Mutlu E. The bacterial microbiome of IBD patients on the specific carbohydrate diet (SCD). Am J Gastroenterol 2013; 108:S552; http://dx.doi.org/ 10.1038/ajg.2012.470 [DOI] [Google Scholar]
- 44. Dixon LE, Kabi A, Nickerson KP, McDonald C. Combinatorial effects of diet and genetics on inflammatory bowel disease pathogenesis. Inflamm Bowel Dis 2015; Jan 9 [Epub ahead of print]; PMID: 25581832; http://dx.doi.org/24448096 10.1097/MIB/0000000000000289 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Korn LL, Thomas HL, Hubbeling HG, Spencer SP, Sinha R, Simkins HM, Salzman NH, Bushman FD, Laufer TM. Conventional CD4+ T cells regulate IL-22-producing intestinal innate lymphoid cells. Mucosal Immunol 2014; 7:1045-57; PMID:24448096; http://dx.doi.org/ 10.1038/mi.2013.121 [DOI] [PMC free article] [PubMed] [Google Scholar]