

Abstract
Sogatella furcifera (Horváth) is the most threatening migratory rice pest in Yunnan, China. S. furcifera overwinters in low- altitude basins and valleys in southern Yunnan and migrates northward in spring and summer of the following year, causing serious damage during migration. The overwintering distribution, areas, and spatial pattern of S. furcifera are relevant to the migration and outbreak of this pest. Based on a 4-yr field survey (2010–2013), this study projected areas suitable for S. furcifera to overwinter using a species distribution model, and analyzed the key influencing climatic factors using principal component analysis (PCA) and ecological niche factor analysis (ENFA). Our field survey showed that the northern latitudinal- and upper elevation limits of overwintering S. furcifera was 25.4° N and 1,608 m in western Yunnan and 24.2° N and 1,563 m in eastern Yunnan. The species distribution model produced a fragmented distribution pattern, with most of which in western Yunnan and only a few in eastern Yunnan. The PCA and ENFA analyses showed that the mean temperature of the driest quarter and the precipitation of the coldest quarter significantly influenced the distribution of S. furcifera in winter. The results suggested that the complex topography, spatial differences in winter temperatures, and host availability altogether determined the distribution of overwintering S. furcifera . Compared with previous surveys, the northern latitudinal- and upper elevation limits of overwintering S. furcifera were higher, while the population became rarer in some suitable areas due to change of farmland utilization in winter and possibly climate change.
Keywords: species distribution model (SDM), bioclimatic factor, ecological niche factor analysis (ENFA), maximum entropy model (MaxEnt), rice pest
The white-backed planthopper, Sogatella furcifera (Horváth) (Hemiptera: Delphacidae), is among the most devastating pests on Asian cultivated rice, Oryza sativa L., in southern China and northern Indochina (Catindig et al. 2009, Cheng 2009). The continuous outbreak of S. furcifera in the last decade has caused immeasurable yield loss to the rice production of this region. Yunnan province in southwestern China is one of the most impacted areas of S. furcifera infestation. Since the major outbreak of S. furcifera in 2007 (Fu et al. 2009), the situation of planthopper infestation remained intensified and then escaladed in the last few years.
S. furcifera migrates from the south to the north annually from late March to August, eventually covering all rice-planting areas in Yunnan (Hu et al. 1988, Gui et al. 2008). Severe damage occurs to rice along the course of migration. Previous studies have shown that migration and high reproduction rate are the main causes of population eruption (Gui et al. 2008, Cheng 2009, Hu et al. 2014). The distribution, population density, and habitat of overwintering S. furcifera has a direct influence on the range and density of migratory populations in the coming year (Denno and Roderick 1990). Therefore, studying overwintering S. furcifera will help understand its migratory dynamics and population eruptions.
S. furcifera is able to overwinter in southern Yunnan in vacant paddies with ratooning rice (Oryza sativa L.) and other Poaceae grasses (i.e., Leersia japonica Honda being the most common species) (Yang et al. 1982; Liu et al. 1991; Tao and Sogawa 2000; Hu et al. 2012, 2015). However, recent studies of overwintering S. furcifera were unable to identify the current distribution range and areas. This may be in part due to the significant change in winter farmland utilization in the region (YBS 1990, 2011). Hence, it is necessary to carry out a new survey for overwintering S. furcifera in Yunnan, to clarify its current spatial distribution, as well as to explain climatic factors that may affect these.
Distribution projections based on the occurrence data and bioclimate factors using various species distribution models (SDMs), for instance, CLIMEX (climate change experiment), ENFA (ecological niche factor analysis), GARP (genetic algorithm for rule-set prediction), and MaxEnt (maximum entropy), have been widely applied to many insects of agronomic interest, especially alien invasive species, to study their distribution patterns and predict their potential risks (Rafoss and Sæthre 2003, Sutherst and Maywald 2005, Peterson and Nakazawa 2008, Zeng et al. 2012, Liu et al. 2013). Among these analyses, ENFA and MaxEnt are the most frequently applied SDMs which project the suitable area of a species using the presence-only data without depending on the complicated bionomical parameters of the focal species (Hirzel et al. 2002, Elith and Graham 2009).
Based on the above-mentioned necessity to reevaluate the current distribution of overwintering S. furcifera in Yunnan and the recent advances in SDMs, this study was designed to project the suitable areas of overwintering S. furcifera in Yunnan based on the presence data obtained from a 4-yr field survey. It also analyzed the key influential climatic factors of overwintering S. furcifera . The results of this study could be useful for elucidating the distribution pattern of S. furcifera and key climatic factors influencing its distribution, to form a better understanding of the spatial and temporal dynamics of the population of S. furcifera and to provide information for the formulation of an outbreak assessment for this rice pest.
Materials and Methods
Field Survey
The survey range was set approximately between 25° N and the southern border of Yunnan Province, based on previous studies on the overwintering rice planthoppers (Liu et al. 1991, Hu 2009) (Supp Table 1 and Supp Fig. 1 [online only]). Supplementary survey areas were also set in valleys and basins that occur north of the survey range to ensure the identification of the northern-most boundary. The survey range was then divided into a 50 by 50 km grid, and field sites were established within each grid based on the following two criteria: 1) at least one site was below 1,500 m elevation; and 2) one site was between 1,500 and 1,700 m above sea level (a.s.l.) in order to determine the upper-elevation distribution limit of overwintering S. furcifera . Previous studies have found that S. furcifera cannot survive the winter in areas above 1,700 m (Liu et al. 1991), therefore, no field sites were set above this altitude level. In total, 116 field sites were established in this study, and the geographical coordinates of each site was recorded using a Garmin eTrex Vista GPS handset (Version 3.2; Garmin Ltd., Taiwan).
Field surveys were carried out in January and February, 2010–2013. Habitat types 1, 2, 3, and 5, described by Hu et al. (2012), were selected to represent the winter habitats due to the large numbers of S. furcifera that were captured from these habitats in that study. Since the size of ratooning rice and other related Poaceae grasses are shorter in winter and the population density of S. furcifera is generally lower, instead of using a 33 by 45 cm white plate, a sweeping net (50 cm in diameter, 85 cm in depth, 90-mesh soft nylon; AQSIQ and SAC 2009) was used to sample for planthoppers in three 2 by 2 m areas within each habitat type within each study site. During sampling, each 2 by 2 m area was swept 20 times to minimize sampling bias. All captured planthoppers (including both adults and nymphs) were preserved in 95% ethanol, and identified in the laboratory using the descriptions by Ballou et al. (1987), Asche and Wilson (1990), and Chen et al. (2000) .
Data Analysis
Presence-only data are commonly used in evaluating species suitability and potential distribution range (i.e., Pearson et al. 2007; Hengl et al. 2009; Liu et al. 2011, 2013), as it excludes absence data, which could be unreliable (Gu and Swihart 2004, Jiménez-Valverde et al. 2008, Lobo et al. 2010). Therefore, only field sites with the presence of S. furcifera were used as data points in subsequent analyses (Supp Table S1 [online only]; Fig. 1 A). Decimal coordinates (with four decimal places) of each data points were arranged according to the user manuals of Biomapper 4.0 (Hirzel et al. 2002) (www2.unil.ch/biomapper) and MaxEnt 3.3.3k (Phillips et al. 2004, 2006) (www.cs.princeton.edu/∼schapire/maxent).
Fig. 1.
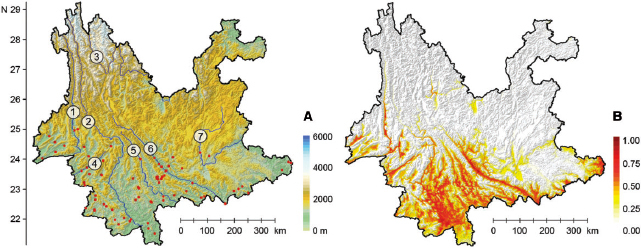
Distribution of presence sites (red dots) in field survey with major rivers (from the west to the east: 1. Nujiang River, 2. Lancang River, 3. Jinsha River, 4. Nanding River, 5. Lixian River, 6. Yuanjiang River, and 7. Nanpan River) in Yunnan (A) and the suitability maps of overwintering S. furcifera produced by MaxEnt (B), color scales indicate altitudes and suitability levels, respectively.
Data of ecogeographical variables (EGVs) were downloaded from WorldClim (www.worldclim.org). The bioclimatic (BioClim) dataset (averaged over 1950–2000) was used in the present research for its biological and ecological properties. The r29 dataset with 30 arc second resolution was obtained and then cropped by the political boundary of Yunnan Province in GlobalMapper 11.0 (www.globalmapper.com), both of the IDRISI (Eastman 1997) and the ASCII formats were used in accordance with Biomapper 4.0 and MaxEnt 3.3.3k, respectively. Host plant (O. sativa) and habitat are important environmental factors influencing the distribution of S. furcifera; however, this study could not include these parameters because the spatial data of rice distribution in Yunnan, especially the distribution of ratooning rice in winter, is not available to date.
In an attempt to analyze the distribution prevalence of overwintering S. furcifera, values of eight winter-related EGVs including, isothermality (Bio3), temperature seasonality (Bio4), minimum temperature of the coldest month (Bio6), mean temperature of the driest quarter (Bio9), mean temperature of the coldest quarter (Bio11), precipitation of the driest month (Bio14), precipitation of the driest quarter (Bio17), and precipitation of the coldest quarter (Bio19) in the overwintering region of S. furcifera, as well as the entire Yunnan Province were extracted using DIVA-GIS 5.7 (www.dive-gis.org) (Hijmans et al. 2012). Distribution frequencies were calculated in Biomapper 4.0. Then, key influencing factors were screened using both of principal component analysis (PCA) and ENFA function in Biomapper 4.0. The importance of factors was measured using the jackknife method in MaxEnt 3.3.3k with 1,000 iterations.
The PCA and the ENFA analyses calculated score matrices for all factors, among which the first n factors with a cumulative percentage of variance over 85% were taken as the most important, and the EGVs’ importance were determined by the absolute values of scores obtained in the analyses (Pearson 1901, Hirzel et al. 2002, Abdi and Williams 2010). Compared with the PCA analysis, results of the ENFA analysis bear ecological meanings, where absolute scores represent the extent of distribution preference of the focal species in comparison to environmental backgrounds, and the signs (+ or −) flag the focal species’ preference to extreme environmental backgrounds (Hirzel et al. 2002). This study applied both analyses for cross-validation, where 0.5 and 0.2 were taken as the boundary absolute scores in screening EGVs in the PCA and the ENFA analyses, respectively. However, the ecological interpretations of the EGVs entirely depended on the ENFA analysis.
The selected EGVs were assigned to the MaxEnt analysis to project suitable areas based on the presence data points, among which 25% were extracted for random testing. The logistic output method was selected to estimate the distribution (or presence) probability of S. furcifera considering certain assumptions of species’ prevalence and sampling effort (Elith et al. 2010). The resulting map was saved as ASCII format and then redrawn using Surfer 10.0 (Golden Software Inc., Golden, CO). Model robustness was evaluated using the receiver operation curve (ROC) and the area under the ROC curve (AUC), where an AUC value [AUC (0, 1)] that approaches 1.0 is usually considered acceptable and it should be rejected when approaching the random turquoise line of 0.5 (Fielding and Bell 1997).
Results
Overview of Presence Sites
The 4-yr field survey collected S. furcifera from 77 sites (presence sites) out of the total 116 field sites surveyed (Supp Table S1 [online only]). Presence of S. furcifera was inconsistent among sample years for 83.1% of the presence sites. Most of the presence sites (75.3%) were located on the western side of the Ailao Mountains, whereas the remaining sites were located on the eastern side. The northern limit of the 77 sites was at 25.4° N in western Yunnan (Wayao, Baoshan, Baoshan Prefecture), whereas the limit in the eastern portion was only 24.2° N (Panxi, Huaning, Yuxi Prefecture) (Fig. 1 A). Altitudes of the 77 presence sites ranged from 100 to 1,608 m a.s.l., and 93.5% of the total presence sites were situated below 1,500 m a.s.l. (Supp Table S1 [online only]). The site in Hekou (Honghe Prefecture) had the lowest altitude and the site in Longba (Mojiang, Pu’er Prefecture) had the highest
Key Influencing Factors
Most of the frequency distributions of the eight overwintering area EGVs of S. furcifera showed different deviations from the normal distributions of the same EGVs for the entire Yunnan Province. Among which, the temperature-related EGVs (Bio6, Bio9, and Bio11) deviated toward the upper limits as expected, and the water-related EGVs (Bio14, Bio17, and Bio19) showed slight deviation toward the upper limits (Fig. 2). For EGVs like Bio3 and Bio4, the frequency distributions of the overwintering area showed deviation in the upper and lower limits, respectively (Fig. 2).
Fig. 2.
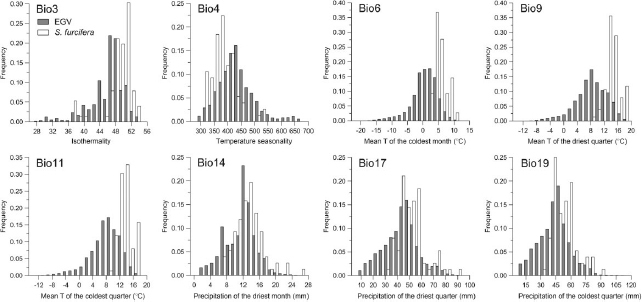
Frequency distribution of the EGVs involved in this study, gray bars: distribution of entire Yunnan Province, and white bars: distribution of overwintering S. furcifera .
In both the PCA and ENFA analyses, Bio9 was selected as the most important temperature-related EGV (−0.914 in PCA; 0.506 in ENFA) whereas Bio19 was the most important water-related EGV (−0.814 in PCA; 0.229 in ENFA) (Table 1). The scores of all other EGVs were listed in Table 1 . The jackknife analysis for importance produced similar results with the PCA and the ENFA analyses, demonstrating that Bio9 was the most important temperature-related EGV for estimating the distribution of S. furcifera in winter (1.047), and Bio19 was the most important water-related EGV (0.292) (Fig. 3 A).
Table 1.
Score matrices of the eight EGVs important for overwintering S. furcifera in PCA and the ENFA
| EGVs | PCA |
ENFA |
|||||
|---|---|---|---|---|---|---|---|
| C1 (65%) | C2 (21%) | C3 (12%) | F1 (59%) | F2 (14%) | F3 (9%) | F4 (8%) | |
| Bio3 | −0.526 | 0.686 | 0.480 | 0.247 | −0.366 | −0.126 | −0.288 |
| Bio4 | 0.754 | −0.525 | −0.343 | −0.303 | −0.086 | −0.410 | −0.254 |
| Bio6 | −0.870 | 0.067 | −0.479 | 0.462 | 0.149 | 0.339 | −0.440 |
| Bio9 | −0.914 | 0.233 | −0.318 | 0.506 | −0.067 | −0.239 | 0.248 |
| Bio11 | −0.909 | 0.218 | −0.350 | 0.485 | 0.041 | −0.166 | 0.113 |
| Bio14 | −0.772 | −0.535 | 0.132 | 0.201 | 0.419 | 0.207 | −0.072 |
| Bio17 | −0.799 | −0.526 | 0.242 | 0.222 | −0.719 | 0.309 | 0.611 |
| Bio19 | −0.814 | −0.481 | 0.268 | 0.229 | 0.370 | −0.691 | −0.454 |
Fig. 3.
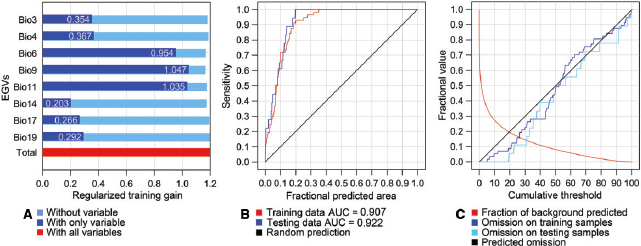
Statistical charts of MaxEnt analysis, (A) the jackknife of the regularized training gain of the EGVs, (B) the ROC and AUC of prediction, and (C) the omission and predicted area, where the values indicate the training gain with only variables.
Suitable Overwintering Areas of S. furcifera
Eight EGVs selected by the PCA and ENFA analyses were utilized in the MaxEnt projection of the suitable overwintering areas for S. furcifera . ROC and AUC were generated (Fig. 3 B and C), where the AUC value of training data was 0.907 and that of testing data was 0.922.
In the projection result, the northern limit of the suitable areas (suitability > 0.5, red colored) was 25.7° N in the west, situated at the middle portion of the Nujiang River Valley, whereas the northern limit was 23.9° N in the east, at the northeastern corner of Funing County (Wenshan Prefecture) (Fig. 1 B). The projected northern limit in the west was slightly higher than that of the presence data points (Fig. 1 A and B).
The suitable overwintering areas of S. furcifera showed similar distribution patterns with the presence data points, in that the suitable areas were aggregated to the west of Ailao Mountains, and most of the suitable areas were river valleys and low-altitudinal basins, i.e., the lower half of the Nujiang River valley and the Lancang River valley, most portions of the Yuanjiang River-, Lixian River-, and Nanding River valleys, and many basins scattered in Dehong, Licang, Pu’er, and Xishuangbanna prefectures in southwestern and southern Yunnan. However, the Nanpan River Valley was less suitable for S. furcifera in winter. It should be noted that though some river valleys (like the Jinsha River Valley and its connected branches) in northern Yunnan showed a low suitability rank (<0.5), no S. furcifera was ever collected during the 4-yr survey. Moreover, the suitability projection suggested that the overwintering areas of S. furcifera were extremely fragmented and scattered into low-altitude areas isolated by mountain ridges (Fig. 1 B).
Discussion
The geographical distribution of S. furcifera is often regarded as one of the key factors which influence the spatial and temporal change of population density and the eruption potential each year. Therefore, previous studies have focused on these factors in an attempt to understand the winter distribution of S. furcifera . In Yunnan, three winter surveys on S. furcifera, carried out in 1979–1981, 1982, and 2000 (Yang et al. 1982, Liu et al. 1991, Tao and Sogawa 2000), found that S. furcifera mostly overwintered in the low altitudinal river valleys and basins that lie to the south of 24.2–25.1° N and below 1,300–1,500 m a.s.l. The surveys also found that the latitudinal- and elevation limits of the distribution in the area west of Ailao Mountains were higher than those to the east of the Ailao Mountains (Yang et al. 1982, Liu et al. 1991, Tao and Sogawa 2000). Our survey showed that the northern limit of S. furcifera in winter was 25.4° N (Wayao, Baoshan) and the upper limit was 1,608 m a.s.l. (Longba, Mojiang) in the area west of Ailao Mountains, while the limits were 24.2° N (Panxi, Huaning) and 1,563 m a.s.l. (Beize, Yuanjiang) in the area east of the Ailao Mountains (Fig. 1 A; Supp Table S1 [online only]). Both of the northern and upper limits of overwintering S. furcifera in this study were higher than that reported by previous studies.
Winter temperature is the factor determining the northern and upper limits of overwintering S. furcifera . Lu et al. (2012) reported a northward shift of the limit in the overwintering brown planthopper, Nilaparvata lugens (Stål) (Hemiptera: Delphacidae), in China due to climate change. Similar cases were widely reported in other arthropods like Dendroctonus ponderosae (Coleoptera: Scolytinae), dragonflies (Odonata), Operophtera brumata (Lepidoptera: Geometridae), as well as Loxosceles reclusa (Araneae: Siraiidae) (Carroll et al. 2004, Hickling et al. 2005, Hagen et al. 2007, Saupe et al. 2011). Global climate change in the recent decades had influenced regional climate significantly. For example, the winter temperature in Yunnan has increased rapidly since 1987, and the winters since 2000 have been warmer than the past (winter temperature increment 0.24–0.26°C per decade) (Duan and Tao 2012, Cheng et al. 2014), resulting in northern tropical and southern subtropical areas expanding significantly (Cheng et al. 2014). Based on this, authors of this study believe that the increase of northern and upper limits of overwintering S. furcifera in Yunnan could be attributed to climate warming driven by global climate change.
Within the geographical range of S. furcifera, overwintering populations were frequently recorded in the longitudinal range-gorge region (LRGR) located west of the Ailao Mountains, while far fewer were scattered on the altiplano located east of this mountain range forming a fragmented pattern restricted by river valleys and low-altitudinal basins (Fig. 1 B). Factor analyses demonstrated that winter temperatures, like the mean temperature of the driest quarter (Bio9) and the mean temperature of the coldest quarter (Bio11), significantly influenced the distribution of S. furcifera in winter (Table 1 and Fig. 3 A). Yunnan lies in the low-latitudinal plateau region of China (Qin et al. 1997, Xie and Liu 1998), with great mountains and deep river valleys that run parallel in the LRGR (He et al. 2005), forming many low-altitudinal basins and valleys which are warm and humid in winter (12–17°C of mean temperature in January) (Wang 2005), making them very suitable for S. furcifera to overwinter. In comparison, the eastern altiplano of Yunnan is much colder and drier in winter (6–14°C of mean temperature in January) (Wang 2005) due to lack of low-altitudinal basins and valleys and frequent exposure to cold currents, making this area much less suitable for S. furcifera to overwinter. This could partially explain why less S. furcifera populations were recorded in eastern Yunnan and the northern and upper limits were much lower in this area. Therefore, the large-scaled fragmented distribution pattern of overwintering S. furcifera is determined by the complex topography and distinct spatial distribution of winter temperature.
Host plant is another factor determining the distribution of S. furcifera in winter. The double-cropping rice areas in the low-altitudinal basins and valleys (<1,300 m) of the LRGR are warm and humid in winter, and the ratooning rice (O. sativa) and L. japonica could be commonly found in vacant or abandoned paddies after harvest (Chen et al. 1994, Zhu et al. 1999, Yu et al. 2009). The abundant ratooning rice and L. japonica provide sufficient food source for S. furcifera in winter. This was likely another reason why most S. furcifera populations were recorded from this area in winter. To the contrast, the single-cropping rice areas in the eastern altiplano as well as the medium-altitude hills (average 1,700–2,000 m) is colder and drier in winter, where ratooning rice and L. japonica are scarcer and restricted to only a few river valleys (Chen et al. 1994, Zhu et al. 1999, Yu et al. 2009). The limited food source in this area further reduced the distribution of overwintering S. furcifera . Our field survey also failed to collect any S. furcifera in winter from areas with very suitable temperature but absence of host plant. Hekou and Duotie in the Red-River Valley of southern Yunnan were good examples of this scenario. Liu et al. (1991) recorded a good number of S. furcifera while our survey did not capture any due to lack of host plant because rice has been completely substituted with banana in most of the valley. Similarly, the Jinsha River (upstream of the Yangtze River) valley of northern Yunnan is warm in winter but extremely dry (Wang 2005, Cheng et al. 2014), where ratooning rice and L. japonica could not survive without irrigation. Despite the mid to low level of suitable areas in this valley (Fig. 1 B), our 4-yr field survey did not find any hosts or collect any S. furcifera . Hence, host availability also determined the limited distribution in certain areas when temperature is suitable. The distribution of S. furcifera is strongly linked to host availability when climate is favorable. Therefore, reducing host availability by agricultural management in winter (i.e., plowing and replantating) is helpful in controlling the population density of overwintering S. furcifera .
This study did not include host plant in the factor analysis since such spatial data are still unavailable. Therefore the projection of suitable areas may bear a certain degree of bias. However, as suggested by many similar studies (Papeş and Gaubert 2007, Fløjgaard et al. 2009, Morueta-Holme et al. 2010, Pittman and Brown 2011, Reside et al. 2012, Yu et al. 2013), the SDM applied in this study is still useful because climate is the most important abiotic factor that determines and influences the distribution of organisms, especially for species where we have poor biological and distributional knowledge. Adding host plants and habitat data might improve the projection accuracy, and provide a more accurate prediction of the distribution of S. furcifera in Yunnan. Therefore, future endeavors are required to collect the spatial data regarding rice agroecosystems in Yunnan during different seasons, which could benefit the projection of S. furcifera as well as other rice pest. Moreover, based on the method applied herein, future studies using short-term weather data instead of long-term climate data (Reside et al. 2010) can be explored with S. furcifera to project more precise distributions under a rapidly changing climate.
Supplementary Data
Supplementary data are available at Journal of Insect Science online.
Supplementary Material
Acknowledgments
The authors wish to express their sincere thanks to the following persons for kind assistance during the course of field survey, data analysis, and coordination: Zhong-liang Han (Plant Protection and Quarantine Station of Yunnan Province), Yan Jin, Jin Xu, Xue-Yu Ma, Shu-Na Ge, Li-Min Dong, Yan-Ping Yu, and Xin Zhang (Laboratory of Biological Invasion and Ecosecurity, Yunnan University). The authors also wish to thank Toby R. Petrice (USDA Forest Service) for improving the earlier drafts of this article. This study was supported with the funds from the Natural Science Foundation of China (31160095), the Key Project of Yunnan Provincial Department of Science and Technology (2011FA003), and the Research Fund (Natural Science) of Yunnan University.
References Cited
- Abdi H., Williams L. J. . 2010. . Principal component analysis. Wiley Interdisciplinary Reviews : Computation. Stat. 2 : 433 – 459 . [Google Scholar]
- (AQSIQ) General Administration of Quality Supervision, Inspection and Quarantine of P.R.C., and (SAC) Standardization Administration Committee of P.R.C . 2009. . National Standard of the People’s Republic of China (GB/T 15794-2009) Rules of Investigation and Forecast for the Rice Planthopper ( Nilaparvata lugens Stål and Sogatella furcifera Horváth). Standard Press of China, Beijing, China . [Google Scholar]
- Asche M., Wilson M. R. . 1990. . The delphacid genus Sogatella and related groups: a revision with special reference to rice-associated species (Homoptera: Fulgoroidea) . Syst. Entomol. 15 : 1 – 42 . [Google Scholar]
- Ballou J. K., Tsai J. H., Wilson S. W. . 1987. . Delphacid planthoppers Sogatella kolophon and Delphacodes idonea (Homoptera: Delphacieae): descriptions of immature stages and notes on biology . Ann. Entomol. Soc. Am. 80 : 312 – 319 . [Google Scholar]
- Carroll A. L., Taylor S. W., Régnière J., Safranyik L. . 2004. . Effect of climate change on range expansion by the mountain pine beetle in British Columbia , pp. 223 – 232 . In Shore T. L., Brooks J. E., Stone J. E. (eds.), Mountain Pine Beetle Symposium: Challenges and Solutions, Information Report BC-X-339, 30–31 October 2003, Pacific Forest Center, Canadian Forest Service, Kelowna, BC . [Google Scholar]
- Catindig J.L.A., Arida G. S., Baehaki S. E., Bentur J. S., Cuong L. Q., Norowi M., Rattanakarn W., Sriratanasak W., Xia J., Lu Z. . 2009. . Situation of planthoppers in Asia , pp. 191 – 220 . In Heong K. L., Hardy B. (eds.), Planthoppers: new threats to the sustainability of intensive rice production systems in Asia . International Rice Research Institute; , Los Baños: . [Google Scholar]
- Chen G. H., Zhang H., Wang G. X., Ren C. F., Huang Y. Q., Liu S. L., Pang J. C. . 1994. . Climatic and ecological division of ratoon rice cultivation in Yunnan Province . J. Southwest Agri. Univ. 16 : 390 – 395 . [Google Scholar]
- Chen X.S., Li Z. Z., Jiang S.N. . 2000. . Taxonomic study on nymphs of Delphacidae (Homoptera: Fulgoroidea) from China II Sogatella . J. Mount. Agri. Biol. 19 : 108 – 112 . [Google Scholar]
- Cheng J. A. 2009. . Rice planthopper problems and relevant causes in China , pp. 157 – 178 . In Heong K. L., Hardy B. (eds.), Planthoppers: new threats to the sustainability of intensive rice production systems in Asia . International Rice Research Institute; , Los Baños: . [Google Scholar]
- Cheng J. G., Song L. C., Huang W., Xu Y. (eds). 2014. . Evaluation report on the projection of climate change in Yunnan and its impact in the next 10–30 Years . China Meteorological Press; , Beijing, China: . [Google Scholar]
- Denno R. F., Roderick G. K. 1990. . Population biology of planthoppers . Ann. Rev. Entomol. 35 : 489 – 520 . [Google Scholar]
- Duan X., Tao Y. . 2012. . The climate change of Yunnan over the last 50 years . J. Trop. Meteorol. 28 : 243 – 250 . [Google Scholar]
- Eastman J. R. 1997. . Idrisi for Windows 2.0 user’s guide. Clark University, Worcester, United Kingdom . [Google Scholar]
- Elith J., Graham C. H. . 2009. . Do they? How do they? Why do they differ? On finding reasons for differing performances of species distribution models . Ecography 32 : 66 – 77 . [Google Scholar]
- Elith J., Phillips S. J., Hastie T., Dudík M., Chee Y. E., Yates C. J. . 2010. . A statistical explanation of MaxEnt for ecologists . Divers. Distrib. 17 : 43 – 57 . [Google Scholar]
- Fielding A. H., Bell J. F. . 1997. . A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ . Conserv. 24 : 38 – 49 . [Google Scholar]
- Fløjgaard C., Normand S., Skov F., Svenning J.-C. . 2009. . Ice age distributions of European small mammals: insights from species distribution modelling . J. Biogeogr. 36 : 1152 – 1163 . [Google Scholar]
- Fu D. Y., Hu S. J., Ye H. . 2009. . Research achievements and prospects of the rice planthoppers in Yunnan Province . Zool. Res. 31 : 280 – 284 . [Google Scholar]
- Gu W. D., Swihart R. K. . 2004. . Absent or undetected? Effect of non-detection of species occurrence on wildlife-habitat models . Biol. Conserv. 116 : 195 – 203 . [Google Scholar]
- Gui F. R., Li Y. H., Han Z. L., Li Z. Y., Lü J. P. . 2008. . Occurrence, outbreak, and countermeasures of the rice plathoppers in Yunnan, China . China Plant Protect. 28 : 15 – 17 . [Google Scholar]
- Hagen S. B., Jepsen J. U., Ims R. A., Yoccoz N. G. . 2007. . Shifting altitudinal distribution of outbreak zones of winter moth Operophtera brumata in sub-arctic birch forest: a response to current climate warming? Ecography 30 : 299 – 307 . [Google Scholar]
- He D. M., Wu S. H., Peng H., Yang Z. F., Ou X. K., Cui B. S. . 2005. . A study of ecosystem changes in Longitudinal Range-Gorge Region and transboundary eco-security in southwestern China . Adv. Earth Sci. 20 : 932 – 943 . [Google Scholar]
- Hengl T., Sierdsema H., Radović A., Dilo A. . 2009. . Spatial prediction of species’ distribution from occurrence-only records: combining point pattern analysis, ENFA and regression-kriging . Ecol. Model. 220 : 3499 – 3511 . [Google Scholar]
- Hickling R., Roy D. B., Hill J. K., Thomas C. D. . 2005. . A northward shift of range margins in British Odonata . Glob. Change Biol. 11 : 502 – 506 . [Google Scholar]
- Hijmans R. J., Guarino L., Mathur P. . 2012. . DIVA-GIS version 7.5 manual. ( http://www.diva-gis.org/documentation ) (accessed 13 October 2013) . [Google Scholar]
- Hirzel A. H., Hausser J., Chessel D., Perrin N. . 2002. . Ecological-niche factor analysis: how to compute habitat-suitability maps without absence data? Ecology 83 : 2027 – 2036 . [Google Scholar]
- Hu G. W., Xie M. X., Wang Y. C. . 1988. . A suggestion for the delimitation of the incidence areas of white-backed planthopper in China . Acta Entomol. Sin. 31 : 42 – 49 . [Google Scholar]
- Hu S. J. 2009. . Preliminary prediction of suitable area for overwintering planthoppers in Yunnan based on DIVA-GIS . Zool. Res. 30 : 122 . [Google Scholar]
- Hu S. J., Fu D. Y., Liu X. J., Zhao T., Han Z. L., Lü J. P., Wan H. L., Ye H. . 2012. . Diversity of planthoppers associated with the winter rice agroecosystems in southern Yunnan, China . J. Insect Sci. 12 : 1 – 11 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu S. J., Fu D. Y., Han Z. L., Ye H. . 2015. . Density, demography, and influential environmental factors on overwintering populations of Sogatella furcifera (Hemiptera: Delphacidae) in southern Yunnan, China . J. Insect Sci. 15 : 58 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu Y., Cheng J. A., Zhu Z. R., Heong K. L., Fu Q., He J. C. . 2014. . A comparative study on population development patterns on Sogatella furcifera between tropical and subtropical areas . J. Asia-Pacific Entomol. 17 : 845 – 851 . [Google Scholar]
- Jiménez-Valverde A., Lobo J. M., Hortal J. . 2008. . Not as good as they seem: the importance of concepts in species distribution modelling . Divers. Distrib. 14 : 885 – 890 . [Google Scholar]
- Liu. J. H., Xiong X. Z., Pan Y. Z., Xiong Z. P., Deng Z. J., Yang L. Y. . 2011. . Predicting potential distribution of oriental fruit fly, Bactrocera dorsalis in Jiangxi Province, South China based on maximum entropy model . Sci. Res. Essays 6 : 2888 – 2894 . [Google Scholar]
- Liu X. F., Jin Y., Ye H. . 2013. . Recent spread and climatic ecological niche of the invasive guava fruit fly, Bactrocera correcta , in mainland China . J. Pest Sci. 86 : 449 – 458 . [Google Scholar]
- Liu Y. B., Yang J. L., Lin L., Kong F. F. . 1991. . The occurrence characteristics of Sogatella furcifera and Nilaparvata lugens in Yunnan . Chin. Bull. Entomol. 28 : 257 – 261 . [Google Scholar]
- Lobo J. M., Jiménez-Valverde A., Hortal J. . 2010. . The uncertain nature of absences and their importance in species distribution modelling . Ecography 33 : 103 – 114 . [Google Scholar]
- Lu X. F., Huo Z. G., Shen S. H., Huang D. P., Wang L., Xiao J. J., Yu C. X. . 2012. . Effects of climate warming on the northern distribution boundary of brown planthopper ( Nilaparvata lugens (Stål)) overwintering in China . Chinese J. Ecol. 31 : 1977 – 1983 . [Google Scholar]
- Morueta-Holme N., Fløjgaard C., Svenning J.-C. . 2010. . Climate change risks and conservation implications for a threatened small-range mammal species . PLoS One 5 : e10630 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papeş M., Gaubert P. . 2007. . Modelling ecological niches from low numbers of occurrence: assessment of the conservation status of poorly known viverrids (Mammalia, Canivora) across two continents . Divers. Distrib. 13 : 890 – 902 . [Google Scholar]
- Pearson K. 1901. . On lines and planes of closest fit to systems of points in space . Philos. Mag. 2 : 559 – 572 . [Google Scholar]
- Pearson R. G., Raxworthy C. J., Nakamura M., Peterson A. T. . 2007. . Predicting species distributions from a small number of occurrence records: a test case using cryptic geckos in Madagascar . J. Biogeogr. 34 : 102 – 117 . [Google Scholar]
- Peterson A. T., Nakazawa Y. 2008. . Environmental data sets matter in ecological niche modelling: an example with Solenopsis invicta and Solenopsis richteri . Glob. Ecol. Biogeogr. 17 : 135 – 144 . [Google Scholar]
- Phillips S. J., Dudík M., Schapire R. E. . 2004. . A maximum entropy approach to species distribution modelling , pp. 655 – 662 . In Brodley C. E. (ed.), Proceedings of the 21st International Conference on Machine Learning . Banff: . [Google Scholar]
- Phillips S. J., Anderson R. P., Schapire R. E. . 2006. . Maximum entropy modeling of species geographic distributions . Ecol. Model. 190 : 231 – 259 . [Google Scholar]
- Pittman S.J., Brown K. A. . 2011. . Multi-scale approach for predicting fish species distributions across coral reef seaspaces . PLoS One 6 : e20583 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin J., Ju J.H., Xie M. E. (eds.). 1997. . Climate in low latitudinal plateau . Meteorology Press; , Beijing, China: . [Google Scholar]
- Rafoss T., Sæthre M.-G. . 2003. . Spatial and temporal distribution of bioclimatic potential for the Codling moth and the Colorado potato beetle in Norway: model predictions versus climate and field data from the 1990s . Agri. For. Entomol. 5 : 75 – 85 . [Google Scholar]
- Reside A. E., VanDerWal J. J., Kutt A. S., Perkins G. C. . 2010. . Weather, not climate, defines distribution of vagile bird species . PLoS One 5 : e13569 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reside A. E., VanDerWal J. J., Kutt A. S. . 2012. . Projection changes in distribution of Australian tropical savanna birds under climate change using three dispersal scenarios . Ecol. Evol. 2 : 705 – 718 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saupe E. E., Papeş M., Selden P. A., Vetter R. S. . 2011. . Tracking a medically important spider: climate change, ecological niche modeling, and the brown recluse ( Loxosceles reclusa ) . PLoS One 6 : e17731 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sutherst R. W., Maywald G. . 2005. . A climate model of the red imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae): implications for invasion of new regions, particularly Oceania . Environ. Entomol. 34 : 317 – 335 . [Google Scholar]
- Tao Y. M., Sogawa K. . 2000. . The survey of winter source of rice planthoppers in Yunnan, China . Yunnan Agri. Sci. Technol. 29 : 3 – 6 . [Google Scholar]
- Wang Y. 2005. . The mountane climate of Yunnan. Yunnan Science & Technology Press; , Kunming, China: . [Google Scholar]
- Xie M. E., Liu Y. . 1998. . Climatic features primary study of global low latitude plateau regions . Yunnan Geogr. Environ. Res. 10 : 25 – 33 . [Google Scholar]
- Yang J. L., Liu Y. B., Kong F. F., Lin L. . 1982. . Winter survey of rice planthoppers . Yunnan Agri. Sci. Technol. 11 : 31 – 35 . [Google Scholar]
- (YBS) Yunnan Bureau of Statistics . 1990. . Yunnan statistic yearbook 1990 . China Statistics Press; , Beijing, China: . [Google Scholar]
- (YBS) Yunnan Bureau of Statistics . 2011. . Yunnan statistic yearbook 2011 . China Statistics Press; , Beijing, China: . [Google Scholar]
- Yu D., Chen M., Zhou Z. C., Eric R., Tang Q. Y., Lu H. Z. . 2013. . Global climate change will severely decrease potential distribution of the East Asian coldwater fish Rhynchocypris oxycephalus (Actinopterygii, Cyprinidae) . Hydrobiologia 700 : 23 – 32 . [Google Scholar]
- Yu L. Q., Guo Y. Q., Yin F. Y., Lu Y. L., Zhang J. P., Xuan S. N., Zhou Y. J. . 2009. . The rice production and weed prevention in Yunnan . China Rice 16 : 70 – 73 . [Google Scholar]
- Zeng X. L., Wang P., Cheng W. J., Wang X. P., Lei C. L. . 2012. . Projecting overwintering regions of the beet armyworm, Spodoptera exigua in China using the CLIMEX model . J. Insect Sci. 12 : 1 – 13 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu Y., Duan C. C., Wang P. Y. . 1999. . The agroclimatic advantages to growing hybrid rice and its zoning in Yunnan Province . Chin. J. Agrometeorol. 20 : 21 – 25 . [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.