

Abstract
The Wall Associated Kinases (WAKs) bind to both cross-linked polymers of pectin in the plant cell wall, but have a higher affinity for smaller fragmented pectins that are generated upon pathogen attack or wounding. WAKs are required for cell expansion during normal seedling development and this involves pectin binding and a signal transduction pathway involving MPK3 and invertase induction. Alternatively WAKs bind pathogen generated pectin fragments to activate a distinct MPK6 dependent stress response. Evidence is provided for a model for how newly generated pectin fragments compete for longer pectins to alter the WAK dependent responses.
Keywords: Arabidopsis, cell wall, plant defense, receptors, signaling
The cell walls of angiosperms are composed of a complex arrangement of cellulose, hemicellulose and pectin. The pectins can be selectively and locally modified to be cross-linked into a structural network that can have dramatic effects on cell enlargement,1-3 but numerous pathogens and mechanical disruptions fragment this pectin network, leading often to a plant stress response.4-6 The Wall Associated Kinases (WAKs) are receptor kinases that bind pectin in the cell wall, and span the plasma membrane to place a serine/threonine kinase in the cytoplasm.7-16 A number of studies have shown that WAKs are required for cell expansion during development, but also mediate a pectin fragment induced stress response.12,16,17 How these receptors can be involved in 2 distinct responses is not well understood, but the key perhaps lies in that WAKs bind to long polymers of pectin cross-linked in the cell wall of unchallenged plants, but also to pectin fragments or oligogalacturonides (OGs) generated by wounding or pathogens as they invade.12,18 What has been missing is a model for how WAKs might distinguish the 2 pectin states so as to trigger expansion versus a stress response. We proposed a model where newly generated OGs compete with native pectin for WAKs, and activate alternate signal transduction pathways.13
Background
Plant cell walls arise through a complex, developmentally regulated coordination of synthesis, turnover, and interactions between protein and carbohydrates.12 Screens for mutants in developmental processes have not surprisingly then revealed numerous alleles of cell wall biosynthesis genes, and conversely mutations in cell wall function have identified alleles in genes normally associated with a variety of metabolic and developmental pathways.21 These genes include receptor kinases such as THE1,FER, HERK, ANX, and RLP44 and have been termed cell wall sensors.19-26 Of the “wall sensors” only WAKs, also receptor kinases, are known to bind to a cell wall component, pectin.
Pectin is first made as methyl esterified α1–4 D-galacturonic acid in the golgi, secreted,1,3,27 and then modified and cross-linked in the extracellular space. Localized activity of Pectin Methylesterases (PME) expose an oxygen to bind calcium that mediates a crosslinking. It is thought that regulation of the location and extent of PME activity can influence wall structure and directionality of loosening thereby influencing cell growth.24 Because WAKs bind to pectin they long have been candidates for the pectin and thus wall sensors for both expansion and pathogen disturbances. WAKs are bound to native pectins in plants10,12,13,16 but in vitro binding assays demonstrate that WAKs have a higher binding of de-esterified over esterified pectins,9,28,29 and have a preference (competition assays) for short OGs of degree of polymerization.9-15 Pathogens tend to target de-esterified pectins. During seedling growth, WAKs are required for cell expansion and have been shown to be involved in the pectin activation of MPK3 and a vacuolar invertase that can increase turgor driven expansion.9,10,12 But WAKs are also required for a response to pathogen, are necessary for the OG stress response14 and bind and trigger a response to OGs in a transient assay.17 This stress response appears to have a distinct signaling pathway and includes a ROS burst, MPK6 activity, and the EDS1 and PAD4 dependent activation of numerous genes, including the ca. 1000 fold induction of a downstream target gene FADlox which serves as a robust indicator.13,14
A Model
We recently showed that a dominant WAK2 allele WAK2cTAP whose encoded protein requires a functional pectin binding domain and an active kinase induces a constitutive stress response.13,14 But importantly, this WAK allele is suppressed by a null allele of a pectin methyl esterase, pme3.13 This provides genetic evidence that WAKs are sensing the de-esterified form of pectin, consistent with the higher affinity in vitro of WAKs for de-esterified over esterified pectin. But we also found that the pme3/pme3 mutant plant is more responsive to OGs than WT plants. One explanation is that since WAK is bound less tightly to esterified pectin in the mutant, then more is available to receive incoming OGs. Collectively the data are consistent with a model (Fig. 1) where OGs are competing with native pectin for WAKs, and this provides a mechanism for WAKs to distinguish pectins, and activate alternate pathways.
Figure 1.
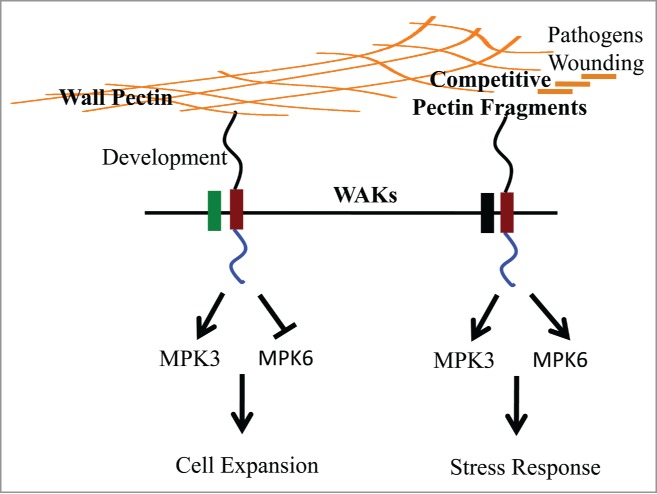
(A) Model for Pectin and OG (pectin fragment) Activation of 2 Responses through WAK. Orange lines represent cell wall pectin, cross-linked or fragmented by pathogens or wounding. OGs outcompete longer polymers for activation of a WAK (red boxes) dependent stress response. WAKs are predicted to associate with co-receptors (green or black) to mediate different responses. In the absence of pathogen, WAKs monitor cell wall pectin and are required for cell expansion. See text for details.
Future Questions
The question now remains as to how the 2 different types of pectins can trigger one receptor to activate different paths. It is possible that part of the mechanism lies in the heterogeneity of the WAK family, as there are 5 WAKs tightly clustered in a 30 KB Arabidopsis locus.30 But it is also possible that WAKs associate with different co-receptors to distinguish the pectin, and these receptor complexes have different downstream partners. Analysis of the components of the 2 pathways should help to answer this next question.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- 1.Somerville C, Bauer S, Brininstool G, Facette M, Hamann T, Milne J, Osborne E, Paredez A, Persson S, Raab T, et al.. Toward a systems approach to understanding plant cell walls. Science 2004; 306:2206-11; PMID:15618507; http://dx.doi.org/ 10.1126/science.1102765 [DOI] [PubMed] [Google Scholar]
- 2.Kohorn BD. Plasma membrane-cell wall contacts. Plant Physiol 2009; 124:31-8; http://dx.doi.org/ 10.1104/pp.124.1.31 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Caffall KH, Mohnen D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr Res 2009; 344:1879-900; PMID:19616198; http://dx.doi.org/ 10.1016/j.carres.2009.05.021 [DOI] [PubMed] [Google Scholar]
- 4.Mohnen D. Pectin structure and biosynthesis. Curr Opin Plant Biol 2008. 11, 266-277; PMID:18486536; http://dx.doi.org/ 10.1016/j.pbi.2008.03.006 [DOI] [PubMed] [Google Scholar]
- 5.Wolf S, Mouille G, Pelloux J. Homogalacturonan methyl-esterification and plant development. Mol Plant 2009; 2:851-60; PMID:19825662; http://dx.doi.org/ 10.1093/mp/ssp066 [DOI] [PubMed] [Google Scholar]
- 6.Peaucelle A, Braybrook S, Höfte H. Cell wall mechanics and growth control in plants: the role of pectins revisited. Front Plant Sci 2012; 3:121; PMID:22685449; http://dx.doi.org/ 10.3389/fpls.2012.00121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.He ZH, Fujiki M, Kohorn BD. A cell wall-associated, receptor-like protein kinase. J Biol Chem 1996; 271:19789-93; PMID:8702686; http://dx.doi.org/ 10.1074/jbc.271.33.19789 [DOI] [PubMed] [Google Scholar]
- 8.Kohorn BD. WAKs; cell wall associated kinases. Curr Opin Cell Biol 2001; 13:529-33; PMID:11544019; http://dx.doi.org/ 10.1016/S0955-0674(00)00247-7 [DOI] [PubMed] [Google Scholar]
- 9.Kohorn BD, Johansen S, Shishido A, Todorova T, Martinez R, Defeo E, Obregon P. Pectin activation of MAP kinase and gene expression is WAK2 dependent. Plant J 2009; 60:974-82; PMID:19737363; http://dx.doi.org/ 10.1111/j.1365-313X.2009.04016.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kohorn BD, Kobayashi M, Johansen S, Friedman HP, Fischer A, Byers N. Wall-associated kinase 1 (WAK1) is crosslinked in endomembranes, and transport to the cell surface requires correct cell-wall synthesis. J Cell Sci 2006; 119:2282-90; PMID:16723734; http://dx.doi.org/ 10.1242/jcs.02968 [DOI] [PubMed] [Google Scholar]
- 11.Kohorn BD, Kobayashi M, Johansen S, Riese J, Huang LF, Koch K, Fu S, Dotson A, Byers N. An Arabidopsis cell wall-associated kinase required for invertase activity and cell growth. Plant J 2006; 46:307-16; PMID:16623892; http://dx.doi.org/ 10.1111/j.1365-313X.2006.02695.x [DOI] [PubMed] [Google Scholar]
- 12.Kohorn BD, Kohorn SL. The cell wall-associated kinases, WAKs, as pectin receptors. Front Plant Sci 2012; 3:88; PMID:22639672; http://dx.doi.org/ 10.3389/fpls.2012.00088 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kohorn BD, Kohorn SL, Saba NJ, Martinez VM. Requirement for pectin methyl esterase and preference for fragmented over native pectins for wall-associated kinase-activated, EDS1/PAD4-dependent stress response in Arabidopsis. J Biol Chem 2014; 289, 18978-86; http://dx.doi.org/ 10.1074/jbc.M114.567545 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kohorn BD, Kohorn SL, Todorova T, Baptiste G, Stansky K, McCullough M. A dominant allele of Arabidopsis pectin-binding wall-associated kinase induces a stress response suppressed by MPK6 but not MPK3 mutations. Mol Plant 2012; 5:841-51; PMID:22155845; http://dx.doi.org/ 10.1093/mp/ssr096 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kohorn BD, Lane S, Smith TA. An Arabidopsis serine/threonine kinase homologue with an epidermal growth factor repeat selected in yeast for its specificity for a thylakoid membrane protein. Proc Natl Acad Sci USA 1992; 89:10989-92; PMID:1438303; http://dx.doi.org/ 10.1073/pnas.89.22.10989 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wagner TA, Kohorn BD. Wall-associated kinases are expressed throughout plant development and are required for cell expansion. Plant Cell 2001; 13, 303-18; PMID:11226187; http://dx.doi.org/ 10.1105/tpc.13.2.303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Brutus A, Sicilia F, Macone A, Cervone F, De Lorenzo G. A domain swap approach reveals a role of the plant wall-associated kinase 1 (WAK1) as a receptor of oligogalacturonides Proc Natl Acad Sci USA 2010; 107:9452-7; PMID:20439716; http://dx.doi.org/ 10.1073/pnas.1000675107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ferrari S, Savatin DV, Sicilia F, Gramegna G, Cervone F, Lorenzo GD. Oligogalacturonides: plant damage-associated molecular patterns and regulators of growth and development. Front Plant Sci 2013; 4:49; PMID:23493833; http://dx.doi.org/ 10.3389/fpls.2013.00049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hematy K, Höfte H. Novel receptor kinases involved in growth regulation. Curr Opin Plant Biol 2008; 11:321-8; PMID:18395489; http://dx.doi.org/ 10.1016/j.pbi.2008.02.008 [DOI] [PubMed] [Google Scholar]
- 20.Hematy K, Sado PE, Van Tuinen A, Rochange S, Desnos T, Balzergue S, Pelletier S, Renou JP, Höfte H. A receptor-like kinase mediates the response of Arabidopsis cells to the inhibition of cellulose synthesis. Curr Biol 2007; 17:922-31; PMID:17540573; http://dx.doi.org/ 10.1016/j.cub.2007.05.018 [DOI] [PubMed] [Google Scholar]
- 21.Wolf S, Mravec J, Greiner S, Mouille G, Höfte H. Plant cell wall homeostasis is mediated by brassinosteroid feedback signaling. Curr Biol 2012; 22:1732-7; PMID:22885061; http://dx.doi.org/ 10.1016/j.cub.2012.07.036 [DOI] [PubMed] [Google Scholar]
- 22.Guo H, Ye H, Li L, Yin Y. A family of receptor-like kinases are regulated by BES1 and involved in plant growth in Arabidopsis thaliana. Plant Signal Behav 2009; 4:784-6; PMID:19820315; http://dx.doi.org/ 10.4161/psb.4.8.9231 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Miyazaki S, Murata T, Sakurai-Ozato N, Kubo M, Demura T, Fukuda H, Hasebe M. ANXUR1 and 2, sister genes to FERONIA/SIRENE, are male factors for coordinated fertilization. Curr Biol 2009; 19:1327-31; PMID:19646876; http://dx.doi.org/ 10.1016/j.cub.2009.06.064 [DOI] [PubMed] [Google Scholar]
- 24.Wolf S, Höfte H. Growth control: a saga of cell walls, ROS, and peptide receptors. Plant Cell 2014; 26:1848-56; PMID:24808052; http://dx.doi.org/ 10.1105/tpc.114.125518 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wolf S, van der Does D, Ladwig F, Sticht C, Kolbeck A, Schurholz AK, Augustin S, Keinath N, Rausch T, Greiner S, et al.. A receptor-like protein mediates the response to pectin modification by activating brassinosteroid signaling. Proc Natl Acad Sci USA 2014; 111:15261-6; PMID:25288746; http://dx.doi.org/ 10.1073/pnas.1322979111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Haruta M, Sabat G, Stecker K, Minkoff BB, Sussman MR. A peptide hormone and its receptor protein kinase regulate plant cell expansion. Science 2014; 343:408-11; PMID:24458638; http://dx.doi.org/ 10.1126/science.1244454 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Harholt J, Suttangkakul A, Vibe Scheller H. Biosynthesis of pectin. Plant Physiol 2010; 153:384-95; PMID:20427466; http://dx.doi.org/ 10.1104/pp.110.156588 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Decreux A, Messiaen J. Wall-associated kinase WAK1 interacts with cell wall pectins in a calcium-induced conformation. Plant Cell Physiol 2005; 46:268-78; PMID:15769808; http://dx.doi.org/ 10.1093/pcp/pci026 [DOI] [PubMed] [Google Scholar]
- 29.Decreux A, Thomas A, Spies B, Brasseur R, Van Cutsem P, Messiaen J. In vitro characterization of the homogalacturonan-binding domain of the wall-associated kinase WAK1 using site-directed mutagenesis. Phytochemistry 2006; 67:1068-79; PMID:16631829; http://dx.doi.org/ 10.1016/j.phytochem.2006.03.009 [DOI] [PubMed] [Google Scholar]
- 30.He ZH, Cheeseman I, He D, Kohorn BD. A cluster of five cell wall-associated receptor kinase genes, Wak1-5, are expressed in specific organs of Arabidopsis. Plant Mol Biol 1999; 39:1189-96; PMID:10380805; http://dx.doi.org/ 10.1023/A:1006197318246 [DOI] [PubMed] [Google Scholar]
- 31.Kohorn BD, Kohorn SL, Saba NJ, Meco Martinez V. Requirement for pectin methyl esterase and preference for fragmented over native pectins for Wall Associated Kinase Activated, EDS1/PAD4 dependent stress response in Arabidopsis. J Biol Chem 2014; 289:18978-86; PMID:24855660; http://dx.doi.org/ 10.1074/jbc.M114.567545 [DOI] [PMC free article] [PubMed] [Google Scholar]