

Abstract
Vernalization establishes a memory of winter that must be maintained for weeks or months in order to promote flowering the following spring. The stability of the vernalized state varies among plant species and depends on the duration of cold exposure. In Arabidopsis thaliana, winter leads to epigenetic silencing of the floral repressor gene FLOWERING LOCUS C (FLC) and the duration of cold is measured through the dynamics of chromatin modifications during and after cold. The growing conditions encountered post-vernalization are thus critical for the maintenance of the vernalized state. We reported that high temperature leads to devernalization and, consistently, to FLC reactivation in Arabidopsis seedlings. Here we show that the repressive epigenetic mark H3K27me3 decreases at the FLC locus when vernalized seedlings are grown at 30°C, unless they were first exposed to a stabilizing period at 20°C. Ambient temperature thus controls the epigenetic memory of winter.
Keywords: Arabidopsis, devernalization, epigenetic modifications, FLOWERING LOCUS C, Flowering, H3K27me3, H3K4me3, vernalization
Vernalization-requiring plants are unable to flower or flower very late unless they are exposed to prolonged cold temperatures. The ecological significance of this trait is to block flowering prior to winter and permit the floral transition under the favorable conditions of following spring, when ambient temperature and daylength increase.1 In Arabidopsis thaliana, vernalization requirement is largely due to the high expression level of the FLOWERING LOCUS C (FLC) gene encoding a MADS-domain protein.2,3 FLC acts as a transcriptional inhibitor of the flowering integrator genes FLOWERING LOCUS T (FT) and SUPPRESSOR OF OVEREXPRESSION OF CO 1 (SOC1).4-7 Vernalization leads to stable silencing of FLC and so relieves FT and SOC1, which can then be activated by a range of environmental or endogenous signals stimulating flowering (reviewed in refs. 8–12).
The initial level of FLC expression depends on the balance between activation and repression activities of FRIGIDA (FRI) and the autonomous flowering pathway, respectively, which regulate antisense FLC transcripts (called COOLAIR) and histone modifications (reviewed in ref. 13). Allelic variation at FRI and FLC loci largely explain the differences in vernalization requirement between winter and summer accessions of Arabidopsis.14-18 Prior to vernalization, histone modifications associated with actively transcribed genes, such as H3K4me3 and H3K36me2/me3, are enriched at FLC whereas repressive marks such as H3K27me3 are present but low abundant.19-24 During vernalization, gradual accumulation of the repressive mark H3K27me3 occurs at the nucleation region, just downstream of the transcription initiation site, and thereafter spreads over the whole region of FLC upon return to warm temperature.19,25-27
In Arabidopsis as well as some other species such as Henbane, vernalization is both quantitative – short cold spells have no effect on flowering – and stable, since cold exposure and the subsequent floral transition may be separated by several months.8 There are, however, species in which vernalization is not stable.9,28 In species with a stable vernalized state, the memory of cold is only maintained after long cold exposure, and this behavior reflects the quantitative nature of the response. At the molecular level, it has been shown that the accumulation of H3K27me3 during vernalization reflects the length of the cold exposure and cell-autonomously regulates the transition from an ‘active’ to a ‘silenced’ state of FLC.25 After longer cold treatment, an increasing number of cells are switched off and thus the overall FLC level is gradually turned down29 and reflects the average state of a cell population.25 Then, upon return to warm temperature, fast deposition of repressive histone marks across the whole gene allows and is required for maintenance of the silenced state,30 which is limited to mitotically active tissues.31
At the physiological level though, the stability of vernalization does not only depend on its duration, but also on the growing conditions prevailing post-vernalization.32 ‘Warm temperatures’ - as used in experimental work on Arabidopsis - are around 20°C and stabilize the vernalized state. By contrast, ‘hot temperatures’ can devernalize the plants, as reported in early work on Arabidopsis.33,34 We showed previously that one week at 30°C given directly after a saturating vernalization period completely suppressed the vernalized state and reactivated FLC in Arabidopsis.28 By contrast, the same treatment given after 2 ‘stabilizing’ weeks of 20°C had no effect. As a complementary experiment, we analyzed the effect of one week at 30°C on the chromatin mark profiles at the FLC locus.
ChIP analyses were performed on high-expressing FLC seedlings (Col-FRI)35 harvested 3 weeks after the end of a 6-week vernalization period or a 3-day stratification as a control (Fig. 1). Protocols for chromatin extraction and use of anti-H3K27me3 and -H3K4me3 antibodies followed manufacturer's instructions (www.diagenode.com).
Figure 1.
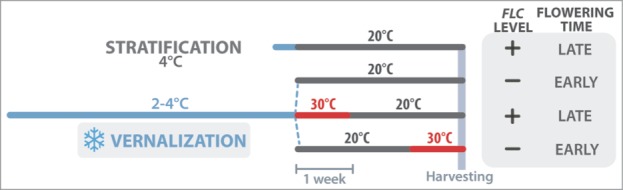
Experimental set-up. Col-FRI seeds were either stratified for 3 days at 4°C or vernalized for 6 weeks at 2–4°C. After the cold period, seedlings were transferred to 20°C (10:14 h light:dark cycles, 100 μE.m−2.s−1) or were first exposed to a devernalization treatment of 1 week at 30°C before transfer to 20°C conditions. The effects of these different post-vernalization conditions on FLC transcript level and flowering are summarized on the right according to Périlleux et al.28
We observed that the repressive H3K27me3 marks were much more abundant in vernalized than in non-vernalized seedlings (Fig. 2), and distributed across the FLC gene as previously reported.25,26 By contrast, vernalization had a very weak effect on the abundance of the H3K4me3 activation marks (Fig. 3). Most interestingly, the 30°C treatment given straight after vernalization completely prevented the accumulation of H3K27me3 marks at all the tested regions of the FLC gene, including the nucleation site (regions I to III in Fig. 2). A slight increase in H3K4me3 activation marks was also detected (Fig. 3). These results are fully consistent with the fact that this 30°C treatment reactivates FLC and inhibits flowering28 (Fig. 1). We can thus conclude that heat removes the epigenetic marks of vernalization when it occurs straight after cold. By contrast, when the heat treatment was given after 2 weeks at 20°C, the repressive H3K27me3 marks of vernalization remained on the chromatin and the vernalized state was maintained (Fig. 2). Angel et al.25 evaluated to 3 days the time taken after cold to achieve maximum H3K27me3 deposition at FLC locus, and hence the growing conditions occurring then are critically important for maintenance of the vernalized state. The erasing effect of heat (Fig. 4) highlights a risk for winter plants if we think of the extreme and erratic weather events that are becoming more common as the climate changes.
Figure 2.

H3K27me3 ChIP experiments. H3K27me3 profile across FLC, for non vernalized seedlings (NV) and after 6 weeks of vernalization (V) followed by 3 weeks of warm (20°C), or 1 week of heat (30°C) given before (30–20°C) or after (20–30°C) 2 weeks of warm. (A) H3K27me3 profile expressed as % of chromatin input retrieved for the V/20°C sample. (B) H3K27me3 quantifications (± SE of 3 technical replicates) relative to constitutive marks at AGAMOUS and SHOOT MERISTEMLESS genes. Two independent biological replicates are shown; similar results were obtained in a third experiment (Supplemental Fig. S1). Primers used are listed in Table S1.
Figure 3.
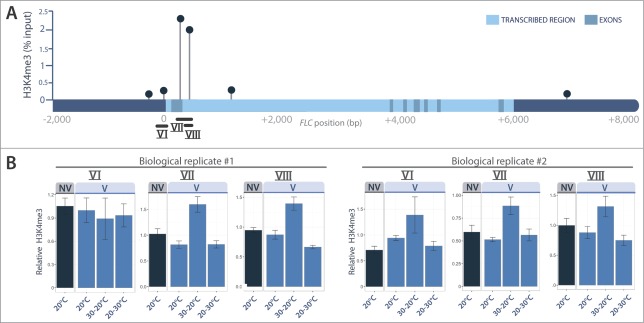
H3K4me3 ChIP experiments. H3K4me3 profile across FLC, for non vernalized seedlings (NV) and after 6 weeks of vernalization (V) followed by 3 weeks of warm (20°C), or 1 week of heat (30°C) given before (30–20°C) or after (20–30°C) 2 weeks of warm. (A) H3K4me3 profile expressed as % of chromatin input retrieved for the NV/20°C sample. (B) H3K4me3 quantifications (± SE of 3 technical replicates) relative to constitutive marks at ACTIN7 and UBIQUITIN10 genes. Two independent biological replicates are shown. Primers used are listed in Table S2.
Figure 4.
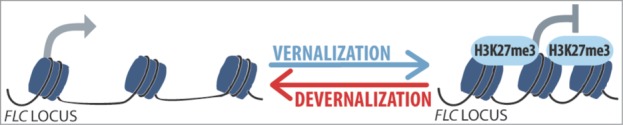
Model of the devernalization process. Vernalization represses the expression of FLC through chromatin modifications, mainly an increase in H3K27me3. Heat occurring just after vernalization restores FLC activity and this is at least partly due to the removal of H3K27me3 repressive marks.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
FB is grateful to the F.R.S.-FNRS for the award of a Ph.D. Fellowship (FC 87200). This research was funded by the Interuniversity Attraction Poles Program, Belgian State, Belgian Science Policy, P7/29. The authors are grateful to Prof. R. Amasino (Department of Biochemistry, University of Wisconsin, Madison, WI) for providing the Col-FRI seeds and to Diagenode for the early access to their Plant ChIP-seq kit (C01010150) and to their Bioruptor® Pico sonicator for chromatin shearing.
Supplemental Material
Primers are listed in Tables S1 and S2. Supplemental Fig. S1 shows a third biological replicate of H3K27me3 analyses.
Supplemental data for this article can be accessed on the publisher's website.
References
- 1. Chouard P. Vernalization and its relations to dormancy. Annu Rev Plant Physiol 1960; 11:191–238; http://dx.doi.org/ 10.1146/annurev.pp.11.060160.001203 [DOI] [Google Scholar]
- 2. Michaels SD, Amasino RM. FLOWERING LOCUS C encodes a novel MADS domain protein that acts as a repressor of flowering. Plant Cell 1999; 11:949-56; PMID:10330478; http://dx.doi.org/ 10.1105/tpc.11.5.949 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Sheldon CC, Burn JE, Perez PP, Metzger J, Edwards JA, Peacock WJ, Dennis ES. The FLF MADS box gene: a repressor of flowering in Arabidopsis regulated by vernalization and methylation. Plant Cell 1999; 11:445-58; PMID:10072403; http://dx.doi.org/ 10.1105/tpc.11.3.445 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Helliwell CA, Wood CC, Robertson M, James Peacock W, Dennis ES. The Arabidopsis FLC protein interacts directly in vivo with SOC1 and FT chromatin and is part of a high-molecular-weight protein complex. Plant J 2006; 46:183-92; PMID:16623882; http://dx.doi.org/ 10.1111/j.1365-313X.2006.02686.x [DOI] [PubMed] [Google Scholar]
- 5. Hepworth SR, Valverde F, Ravenscroft D, Mouradov A, Coupland G. Antagonistic regulation of flowering-time gene SOC1 by CONSTANS and FLC via separate promoter motifs. EMBO J 2002; 21:4327-37; PMID:12169635; http://dx.doi.org/ 10.1093/emboj/cdf432 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Michaels SD, Himelblau E, Kim SY, Schomburg FM, Amasino RM. Integration of flowering signals in winter-annual Arabidopsis. Plant Physiol 2005; 137:149-56; PMID:15618421; http://dx.doi.org/ 10.1104/pp.104.052811 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Searle I, He Y, Turck F, Vincent C, Fornara F, Krober S, Amasino RA, Coupland G. The transcription factor FLC confers a flowering response to vernalization by repressing meristem competence and systemic signaling in Arabidopsis. Genes Dev 2006; 20:898-912; PMID:16600915; http://dx.doi.org/ 10.1101/gad.373506 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Amasino R. Seasonal and developmental timing of flowering. Plant J 2010; 61:1001-13; PMID:20409274; http://dx.doi.org/ 10.1111/j.1365-313X.2010.04148.x [DOI] [PubMed] [Google Scholar]
- 9. Andres F, Coupland G. The genetic basis of flowering responses to seasonal cues. Nat Rev Genet 2012; 13:627-39; PMID:22898651; http://dx.doi.org/ 10.1038/nrg3291 [DOI] [PubMed] [Google Scholar]
- 10. Kim DH, Sung S. Genetic and epigenetic mechanisms underlying vernalization. Arabidopsis Book 2014; 12:e0171; PMID:24653667; http://dx.doi.org/ 10.1199/tab.0171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Song J, Irwin J, Dean C. Remembering the prolonged cold of winter. Curr Biol 2013; 23:R807-11; PMID:24028964; http://dx.doi.org/ 10.1016/j.cub.2013.07.027 [DOI] [PubMed] [Google Scholar]
- 12. Zografos BR, Sung S. Vernalization-mediated chromatin changes. J Exp Bot 2012; 63:4343-8; PMID:22685309; http://dx.doi.org/ 10.1093/jxb/ers157 [DOI] [PubMed] [Google Scholar]
- 13. Ietswaart R, Wu Z, Dean C. Flowering time control: another window to the connection between antisense RNA and chromatin. Trends Genet 2012; 28:445-53; PMID:22785023; http://dx.doi.org/ 10.1016/j.tig.2012.06.002 [DOI] [PubMed] [Google Scholar]
- 14. Gazzani S, Gendall AR, Lister C, Dean C. Analysis of the molecular basis of flowering time variation in Arabidopsis accessions. Plant Physiol 2003; 132:1107-14; PMID:12805638; http://dx.doi.org/ 10.1104/pp.103.021212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Johanson U, West J, Lister C, Michaels S, Amasino R, Dean C. Molecular analysis of FRIGIDA, a major determinant of natural variation in Arabidopsis flowering time. Science 2000; 290:344-7; PMID:11030654; http://dx.doi.org/ 10.1126/science.290.5490.344 [DOI] [PubMed] [Google Scholar]
- 16. Lempe J, Balasubramanian S, Sureshkumar S, Singh A, Schmid M, Weigel D. Diversity of flowering responses in wild Arabidopsis thaliana strains. PLoS Genet 2005; 1:109-18; PMID:16103920; http://dx.doi.org/ 10.1371/journal.pgen.0010006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Michaels SD, He Y, Scortecci KC, Amasino RM. Attenuation of FLOWERING LOCUS C activity as a mechanism for the evolution of summer-annual flowering behavior in Arabidopsis. Proc Natl Acad Sci U S A 2003; 100:10102-7; PMID:12904584; http://dx.doi.org/ 10.1073/pnas.1531467100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Werner JD, Borevitz JO, Uhlenhaut NH, Ecker JR, Chory J, Weigel D. FRIGIDA-independent variation in flowering time of natural Arabidopsis thaliana accessions. Genetics 2005; 170:1197-207; PMID:15911588; http://dx.doi.org/ 10.1534/genetics.104.036533 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Bastow R, Mylne JS, Lister C, Lippman Z, Martienssen RA, Dean C. Vernalization requires epigenetic silencing of FLC by histone methylation. Nature 2004; 427:164-7; PMID:14712277; http://dx.doi.org/ 10.1038/nature02269 [DOI] [PubMed] [Google Scholar]
- 20. Cao Y, Dai Y, Cui S, Ma L. Histone H2B monoubiquitination in the chromatin of FLOWERING LOCUS C regulates flowering time in Arabidopsis. Plant Cell 2008; 20:2586-602; PMID:18849490; http://dx.doi.org/ 10.1105/tpc.108.062760 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. He Y, Doyle MR, Amasino RM. PAF1-complex-mediated histone methylation of FLOWERING LOCUS C chromatin is required for the vernalization-responsive, winter-annual habit in Arabidopsis. Genes Dev 2004; 18:2774-84; PMID:15520273; http://dx.doi.org/ 10.1101/gad.1244504 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Oh S, Park S, van Nocker S. Genic and global functions for Paf1C in chromatin modification and gene expression in Arabidopsis. PLoS Genet 2008; 4:e1000077; PMID:18725930; http://dx.doi.org/ 10.1371/journal.pgen.1000077 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Xu L, Zhao Z, Dong A, Soubigou-Taconnat L, Renou JP, Steinmetz A, Shen WH. Di- and tri- but not monomethylation on histone H3 lysine 36 marks active transcription of genes involved in flowering time regulation and other processes in Arabidopsis thaliana. Mol Cell Biol 2008; 28:1348-60; PMID:18070919; http://dx.doi.org/ 10.1128/MCB.01607-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Zhao Z, Yu Y, Meyer D, Wu C, Shen WH. Prevention of early flowering by expression of FLOWERING LOCUS C requires methylation of histone H3 K36. Nat Cell Biol 2005; 7:1256-60; PMID:16299497; http://dx.doi.org/ 10.1038/ncb1329 [DOI] [PubMed] [Google Scholar]
- 25. Angel A, Song J, Dean C, Howard M. A Polycomb-based switch underlying quantitative epigenetic memory. Nature 2011; 476:105-8; PMID:21785438; http://dx.doi.org/ 10.1038/nature10241 [DOI] [PubMed] [Google Scholar]
- 26. De Lucia F, Crevillen P, Jones AM, Greb T, Dean C. A PHD-polycomb repressive complex 2 triggers the epigenetic silencing of FLC during vernalization. Proc Natl Acad Sci USA 2008; 105:16831-6; PMID:18854416; http://dx.doi.org/ 10.1073/pnas.0808687105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Sung S, Amasino RM. Vernalization in Arabidopsis thaliana is mediated by the PHD finger protein VIN3. Nature 2004; 427:159-64; PMID:14712276; http://dx.doi.org/ 10.1038/nature02195 [DOI] [PubMed] [Google Scholar]
- 28. Périlleux C, Pieltain A, Jacquemin G, Bouché F, Detry N, D'Aloia M, Thiry L, Aljochim P, Delansnay M, Mathieu AS, et al. A root chicory MADS box sequence and the Arabidopsis flowering repressor FLC share common features that suggest conserved function in vernalization and de-vernalization responses. Plant J 2013; 75:390-402; http://dx.doi.org/ 10.1111/tpj.12208 [DOI] [PubMed] [Google Scholar]
- 29. Sheldon CC, Rouse DT, Finnegan EJ, Peacock WJ, Dennis ES. The molecular basis of vernalization: the central role of FLOWERING LOCUS C ( FLC). Proc Natl Acad Sci USA 2000; 97:3753-8; PMID:10716723; http://dx.doi.org/ 10.1073/pnas.97.7.3753 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Satake A, Iwasa Y. A stochastic model of chromatin modification: cell population coding of winter memory in plants. J Theor Biol 2012; 302:6-17; PMID:22381539; http://dx.doi.org/ 10.1016/j.jtbi.2012.02.009 [DOI] [PubMed] [Google Scholar]
- 31. Finnegan EJ, Dennis ES. Vernalization-induced trimethylation of histone H3 lysine 27 at FLC is not maintained in mitotically quiescent cells. Curr Biol 2007; 17:1978-83; PMID:17980595; http://dx.doi.org/ 10.1016/j.cub.2007.10.026 [DOI] [PubMed] [Google Scholar]
- 32. Bernier G, Kinet JM, Sachs RM. The Physiology of Flowering Vol. I : Initiation of Flowers. CRC Press, 1981. [Google Scholar]
- 33. Chintraruck B, Ketellapper HJ. Interaction of vernalization, photoperiod and high temperature in flowering of Arabidopsis thaliana (L.) HEYNH. Plant Cell Physiol 1969; 10:271-6. [Google Scholar]
- 34. Napp-Zinn K. Untersuchungen über das Vernalisationsverhalten einer winterannuellen Rasse von Arabidopsis thaliana. Planta 1957; 50:177-210; http://dx.doi.org/ 10.1007/BF01930342 [DOI] [Google Scholar]
- 35. Lee I, Amasino RM. Effect of vernalization, photoperiod, and light quality on the flowering phenotype of Arabidopsis plants containing the FRIGIDA gene. Plant Physiol 1995; 108:157-62; PMID:12228459 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.