

Abstract
Background
Theory-of-mind (ToM), the ability to infer people’s thoughts and feelings, is a pivotal skill in effective social interactions. Individuals with autism spectrum disorders (ASD) have been found to have altered ToM skills, which significantly impacts the quality of their social interactions. Neuroimaging studies have reported altered activation of the ToM cortical network, especially in adults with autism, yet little is known about the brain responses underlying ToM in younger individuals with ASD. This functional magnetic resonance imaging (fMRI) study investigated the neural mechanisms underlying ToM in high-functioning children and adolescents with ASD and matched typically developing (TD) peers.
Methods
fMRI data were acquired from 13 participants with ASD and 13 TD control participants while they watched animations involving two “interacting” geometrical shapes.
Results
Participants with ASD showed significantly reduced activation, relative to TD controls, in regions considered part of the ToM network, the mirror network, and the cerebellum. Functional connectivity analyses revealed underconnectivity between frontal and posterior regions during task performance in the ASD participants.
Conclusions
Overall, the findings of this study reveal disruptions in the brain circuitry underlying ToM in ASD at multiple levels, including decreased activation and decreased functional connectivity.
Electronic supplementary material
The online version of this article (doi:10.1186/s13229-015-0052-x) contains supplementary material, which is available to authorized users.
Keywords: Theory-of-mind; Autism; fMRI; Functional connectivity MRI, mentalizing
Background
Theory-of-mind (ToM) underlies the ability to explain and predict the behavior of others by attributing to them specific mental states such as their intentions, beliefs, desires, or emotions [1]. ToM also influences the quality of social interactions by enabling people to effectively navigate the interpersonal world and make common sense explanations of others’ behavior. Functional magnetic resonance imaging (fMRI) studies in typically developing (TD) individuals have identified a frontal-posterior network that activates during ToM tasks, which includes the medial prefrontal cortex (MPFC), the posterior cingulate cortex (PCC), and bilateral temporoparietal junction (LTPJ and RTPJ) [2, 3]. It should be noted that while these regions have been central to the ToM network, regions such as the right anterior superior temporal sulcus and medial precuneus have also been implicated in processing ToM [4, 5]. Each of these core regions (MPFC, PCC, LTPJ, RTPJ) has been implicated in several specific social cognitive processes. For instance, the MPFC has been associated with reflective reasoning about actions and judgments, including goals and intentions [6, 7], the PCC with visual imagery, retrieval of episodic information, and self-projection [8, 9], and bilateral TPJ with making transient mental inferences (goals, beliefs) about people [10].
Impairment in social interactions is one of the defining characteristics of autism spectrum disorders (ASD) [11, 12], which may underlie a deficit in social cognition in general, and a deficit in ToM in particular [13–17]. Deficits in ToM can have detrimental effects on social cognition and can carry its effects into other domains of cognitive functioning in individuals with ASD [18]. Functional MRI studies have complemented the behavioral findings of poor ToM skills by reporting altered brain activity during ToM tasks in ASD [4, 10, 19–27]. More recently, functional connectivity MRI (fcMRI), which assesses the correlation of the fMRI blood-oxygen-level dependent (BOLD) time-series of a pair of brain areas, has been used by some of these studies to characterize the neural circuitry underlying ToM in ASD [22–25, 27]. Overall, there is an emerging consensus that aberrant activity and connectivity among different brain regions during ToM and other social-cognitive functions can be considered a strong neurobiological feature of brain functioning in ASD [18, 28–32].
Many previous fMRI studies examining connectivity of the ToM network have found reduced functional connectivity between frontal and more posterior regions in adults with ASD, which may entail disrupted functional integration among core regions underlying such tasks [28]. Since ToM is a complex function mediated by the topographically distributed ToM network including four key nodes (PCC, MPFC, LTPJ, RTPJ), coordination within this network and across other centers may be critical in accomplishing social tasks. While weaker synchronization among these regions has been previously found in individuals with ASD during ToM tasks [10, 21, 23, 24], reduced connectivity may not be a universal feature of autism due to its heterogeneity [33, 34]. In addition, children with ASD may show a different connectivity profile than adults with ASD [35, 36], though this still remains a topic of debate [29]. It should be noted that methodological differences across neuroimaging studies have affected the consistency, reliability, and replicability of functional connectivity findings in ASD [37]. While functional connectivity patterns in adults with ASD may provide a snapshot of how the disorder has progressed and affected the brain over several years, it is equally important to understand the nature of connectivity during childhood and adolescence for a better characterization of the developmental profile of this disorder.
The current study examines ToM network connectivity in children and adolescents with ASD using animations involving the complex movement of geometrical shapes in ways that often evoke a social interpretation of their interaction. These animations have been used successfully in a growing number of studies to investigate ToM and attribution of agency in individuals with and without ASD (see [38] for an extensive review). In autism, such stimuli have been used in two previous neuroimaging studies of ToM in adults, both of which found underconnectivity in participants with autism [21, 23]. The purpose of the current study is to assess the brain responses involved in processing ToM in children and pre-adolescents with ASD using region of interest (ROI) analysis as well as whole-brain analysis. Thus, activation and functional connectivity patterns across different levels of analyses are examined. ROI analyses involved regions from the ToM network, as well as additional regions such as the superior temporal sulcus (STS) due to its role in social perception [39], regions considered part of the substrate for action understanding, such as the inferior frontal gyrus [6], angular gyrus [40], and cerebellum, a region that has increasingly shown, of late, to be involved in social cognition [41]. We hypothesize atypical brain activation and functional connectivity in children and adolescents with ASD. This study is novel in that it uses dynamic non-verbal stimuli to examine the integrity of the ToM network in pre-adult ASD, a relatively under-studied topic. The findings of this study will provide valuable insight into understanding the functional makeup of an important neural network that is critical in characterizing the social symptoms in individuals with ASD.
Methods
Participants
Thirteen high-functioning children and adolescents with autism (mean age = 12.6 years, range = 10–16 years) and thirteen TD control individuals (mean age = 12.7 years, range = 10–15 years) were included in this study (all with full-scale and verbal IQ scores of 75 or above). Participants were matched as a group on the basis of age, gender, handedness, and IQ (Table 1). IQ was assessed in all participants using the Wechsler Abbreviated Scale of Intelligence (WASI-I) [42]. The diagnosis of autism, as per DSM-IV guidelines [43], was established using two structured research diagnostic instruments, the Autism Diagnostic Interview-Revised (ADI-R) [44] and the Autism Diagnostic Observation Schedule-Generic (ADOS-G) [45], supplemented with confirmation by expert opinion. Potential participants with autism were excluded on the basis of an associated infectious or genetic disorder, such as fragile-X syndrome or tuberous sclerosis. Potential control participants and participants with autism were also excluded if found to have evidence of birth asphyxia, head injury, or a seizure disorder. Exclusionary criteria were based on neurologic history, examination, and chromosomal analysis. Potential control participants were also screened to exclude those with medical illnesses or a family history of autism, developmental cognitive disorder, affective disorder, anxiety disorder, schizophrenia, obsessive compulsive disorder, or other neurologic or psychiatric disorders thought to have a genetic component (in first-degree relatives or self). They were also excluded based on medications that affect the central nervous system, hypertension, diabetes, substance abuse (self or first-degree relative), steroid use (extreme use such as steroids used in inhalers for asthmatics), and autism in first-, second-, or third-degree relatives. Each participant’s parents signed an informed consent, and written assent was obtained from all minor participants. These documents and procedures had been approved by the University of Pittsburgh and Carnegie Mellon University Institutional Review Boards (Table 1).
Table 1.
Demographic information
| Groups | p value | ||
|---|---|---|---|
| TD (n = 13) | ASD (n = 13) | ||
| Gender | 11 M, 2 F | 11 M, 2 F | – |
| Handedness | 12 R, 1 L | 12 R, 1 L | – |
| Age (in years) | 12.7 (±1.5; 10–15) | 12.6 (±1.9; 10–16) | 0.83 |
| Verbal IQ | 103.7 (±10.1; 87–120) | 98.2 (±13.8; 82–119) | 0.26 |
| Performance IQ | 107.8 (±7.6; 94–120) | 103.2 (±17.8; 89–132) | 0.40 |
| Full-scale IQ | 106.9 (±8.7; 93–119) | 100.8 (±15.2; 83–128) | 0.25 |
| ADOS-G | |||
| - Communication | – | 4.0 (±0.9; 3–5) | – |
| - Social | – | 9.3 (±1.6; 6–11) | – |
| - Socio-communicative | – | 13.3 (±1.7; 3–16) | – |
| ADI-R (in months) | |||
| - Word | – | 24.3 (±10.0; 10–48) | – |
| - Phrase | – | 113.3 (±278; 12–997) | – |
Values are presented as mean (standard deviation; range). The p value is independent t tests for differences between groups. ADOS scores were not available for one individual
TD typically developing, ASD autism spectrum disorder, M male, F female, R right, L left
Experimental paradigm
This experiment compared the brain activation of autism and TD control participants while they were interpreting the behaviors of two interacting animated figures (a large red triangle and a small blue triangle) commonly known as the Frith-Happé animations [38, 46]. For example, if the red triangle’s movement mirrored the blue triangle’s movement but with some delay, the red triangle could be interpreted to be chasing the blue triangle. Three types of animations were used: theory-of-mind (ToM), goal-directed (GD), and random (RD). In the ToM condition, the geometrical figures moved in a way that could be interpreted as driven by an intentional action or interaction involving thoughts and feelings (e.g., coaxing). In the GD condition, the geometrical figures engaged in an interaction with each other in a simple purposeful level (e.g., chasing). In the RD condition, the geometrical figures did not engage with each other at all (e.g., two individual tennis ball movements). There were three epochs each of ToM, GD, and RD stimuli, each containing one animation. The basic visual characteristics depicted in the three types of animations were similar in terms of shape, overall speed of motion, orientation changes, and total duration (between 26 and 47 s).
After the presentation of each animation, four single-word response alternatives that described different actions were presented on the screen and participants were asked to make a forced-choice judgment about which of the words best described each animated action. The correct response was always an accurate description of the animation (determined as the most frequently generated description in a norming study). Another response alternative was an inaccurate description of the animation, but of the appropriate category (e.g., in the ToM condition, this response referred to a mental state, but an incorrect mental state). The other two answers were inaccurate descriptions of the animation that could have applied to animations in the other two conditions (i.e., in the ToM condition, these incorrect response alternatives referred to a random motion and a goal-directed motion; in the GD condition, they referred to a ToM motion and a random motion; and in the RD condition, they referred to a ToM motion and a goal-directed motion). For example, for a ToM animation that depicted “coaxing,” the foils were pushing (GD), surprising (ToM), and spinning (RD). Participants made their responses using two two-button mice, one held in each hand. Each button corresponded to one of the four multiple-choice answers. Responses were accepted for 15 s from the end of each animation. The presentation of each animation constituted a separate event in the experimental design. The animations were presented in blocks of three, one from each condition, with a separation of 6 s between trials within a block. The onset of each animation was synchronized with the beginning of a TR. A 30-s fixation interval was presented between each block, with participants instructed to stare at a centered asterisk and relax.
fMRI acquisition
All imaging data were acquired at the Brain Imaging Research Center (BIRC), jointly administered by Carnegie Mellon University and the University of Pittsburgh on a 3T Siemens Allegra scanner. The stimuli were rear-projected onto a semi-translucent plastic screen, and participants viewed the screen through a mirror attached to the head coil. For the functional imaging, a gradient echo, echo-planar pulse sequence was used with TR = 1000 ms, TE = 30 ms, and a flip angle of 60°. Sixteen adjacent oblique-axial slices were acquired in a single cycle of scanning in an interleaved sequence, with a 5-mm slice thickness, a 1-mm slice gap, a 20 × 20 cm FOV, and a 64 × 64 matrix, resulting in an in-plane resolution of 3.125 × 3.125 mm, for a total of 666 volumes (11 min and 6 s). A 160-slice 3D MPRAGE volume scan with TR = 200 ms, TE = 3.34 ms, flip angle = 7°, FOV = 25.6 cm, 256 × 256 matrix size, and 1-mm slice thickness was acquired at the same orientation as the oblique-axial functional images.
Data preprocessing
Functional images were processed using a combination of Analysis of Functional NeuroImages (AFNI) software [47] and the Oxford Centre for Functional Magnetic Resonance Imaging of the Brain (FMRIB) Software Library (FSL) [48]. Functional images were slice-time corrected, and correction for head motion was performed by registering each functional volume to the first time point of the scan using AFNI’s 3dvolreg. These images were then registered to the anatomical images via FSL’s linear image registration tool FLIRT [49, 50]. Both images were resampled (3 mm isotropic) and standardized to Montreal Neurological Institute (MNI) space via FSL’s nonlinear registration tool (FNIRT), and a Gaussian spatial smoothing filter with a global full width at half maximum (FWHM) of 8 mm was applied using AFNI’s 3dBlurToFWHM.
fMRI activation analysis
Functional images were individually scaled to a mean of 100, and statistical analysis was performed on individual data by using a general linear model (GLM) via AFNI’s 3dDeconvolve with ToM, GD, RD, ToM question (ToMQ), goal-directed question (GDQ), and random question (RDQ) trials as regressors of interest. Each condition was modeled using a duration-modulated (between 26 and 47 s) hemodynamic response function (the dmBLOCK option in 3dDeconvolve). Six additional rigid-body motion parameters acquired from motion estimation were modeled as nuisance covariates in the GLM. The following orthogonal contrasts were computed to assess average differences in brain response, based on the interest of our study: theory-of-mind vs. random animation (ToM + ToMQ vs. RD + RDQ) and goal-directed vs. random Animation (GD + GDQ vs. RD + RDQ).
ROI analysis was first conducted using regions considered part of the ToM network (MPFC, PCC, and bilateral TPJ) along with other regions associated with social perception (bilateral STS and cerebellum) and action understanding (bilateral inferior frontal and angular gyrus). The ROIs for this analysis were obtained using an independent whole-brain activation map (for the contrast social ToM > RD) derived from the Human Connectome Project (HCP), freely available in NeuroVault (http://neurovault.org/images/3180) [51] (see Additional file 1: Table S1 for MNI coordinates for these ROIs). The following 12 ROIs were identified: medial prefrontal cortex (MPFC), posterior cingulate cortex (PCC), temporoparietal junction (LTPJ, RTPJ), superior temporal sulcus (LSTS, RSTS), inferior frontal gyrus (LIFG, RIFG), angular gyrus (LANG, RANG), and cerebellum (LCEREB, RCEREB). These ROIs were created using spherical binary masks (10-mm radius), and mean parameter estimates averaged across all activated voxels within an ROI were extracted from this same contrast on an individual basis for each ROI to inspect individual variability, and were then statistically compared between the groups using a series of two-sample t tests using false discovery rate (FDR) for multiple comparisons correction procedure.
For whole-brain analysis, areas of statistically significant activation differences were determined using one- and two-sample t tests using a random-effects model via AFNI’s 3dttest++. To correct for multiple comparisons, 10,000 Monte Carlo simulations were applied via AFNI’s 3dClustSim function to obtain a corrected significance level of p < 0.05 (uncorrected voxelwise threshold of p < 0.025; minimum cluster size of 100 voxels). In summary, there were two levels of analyses to assess brain activation: ROI and whole-brain analysis.
Functional connectivity
To examine functional connectivity during each experimental condition, the activation time-series were extracted using AFNI’s 3dmaskave from the ToM, RD, and GD epochs and were then concatenated to create single time-series for each condition. Several steps of preprocessing attempted to first account for or eliminate several extraneous factors. To minimize signal from the cerebral white matter and lateral ventricles, masks were created at the participant level using FSL’s FAST automated segmentation [52]. Masks were trimmed from the white matter to avoid partial-volume effects, and an average time-series for each region was extracted (described below). Derivatives for head motion, white matter, and ventricular time-series were computed. Following spatial smoothing, sources of noise (head motion, white matter, and lateral ventricles plus derivatives) were modeled and removed using a general linear model, and the residual time-series were used in subsequent functional connectivity analysis.
The ROIs mentioned above, derived from NeuroVault, were used for functional connectivity analysis. Given the large number of pairwise comparisons across all ROIs, increasing the likelihood of type I error, the ROIs were grouped into five sets based on their anatomical locations: frontal (LIFG, RIFG), medial (MPFC, PCC), parietal (LANG, RANG), temporal (LSTS, RSTS, LTPJ, RTPJ), and cerebellum (LCEREB, RCEREB). The average time courses were extracted from these sets from individual participants for all three experimental conditions (ToM, GD, and RD) and were correlated across sets to assess the synchronization between them. Correlation coefficients were Fisher’s z-transformed, using an inverse hyperbolic tangent function. FDR correction was also applied for all connectivity analyses. In summary, overall functional connectivity and the connectivity among these five functionally defined and anatomically grouped sets of ROIs was measured for each participant separately for each of the ToM, GD, and RD conditions.
Accounting for head motion
Because head motion can impact functional connectivity analysis [53, 54], the following precautions were taken. Head motion was quantified as the Euclidean distance calculated from six rigid-body motion parameters (x, y, z, pitch, roll, yaw) for each pair of consecutive time points. For any time point where this measure was >1 mm, which was considered excessive motion, that time point, as well as the immediately preceding and subsequent time points, was modeled out [55]. All participants retained more than 80 % of their time points after censoring, and the number of retained time points did not significantly differ between groups [MTD = 663; MASD = 642, t(24) = 1.61, p = 0.12]. Average head motion over each participant’s session was defined as the root mean square of displacement (RMSD) and did not significantly differ between groups [MTD = 0.13; MASD = 0.15, t(24) = 1.18, p = 0.25].
Results
Overview
The main results of this study are the following: (1) significant activation in regions primarily associated with processing ToM and those associated with action understanding (frontal and parietal regions) in both ASD and TD participants; (2) this pattern of activation closely resembles the ToM activation map from the HCP that used a larger sample size; (3) significantly reduced activation on both ROI analysis and whole-brain analysis during ToM processing in the ASD participants, relative to TD peers, in regions that are part of the ToM network and that of action understanding; (4) no significant between-group differences in activation while processing goal-directed animations; and (5) functional underconnectivity in the ASD group relative to the TD group between anatomical networks (fronto-medial, fronto-parietal, and medial-cerebellum).
Behavioral results
To assess possible group differences in reaction time (RT) and performance accuracy (assessed by error rate) measured during the fMRI task, we conducted two separate two-way (group: TD and ASD) × 3 (condition: ToM, GD, and RD) repeated measures ANOVAs. The first ANOVA revealed a main effect of condition [F(2, 48) = 21.4, p < .001]; however, there was no significant main effect of group [F(1, 24) = 0.6, p = 0.5], nor a group × condition interaction [F(2, 48) = 0.7, p = 0.5], indicating that the groups responded equally in terms of RT.
The second ANOVA testing effects on accuracy of responding revealed a main effect of condition [F(2, 48) = 37.6, p < .001] and a main effect of group [F(1, 24) = 9.5, p < .01]; however, the group × condition interaction was not significant [F(2, 48) = 1.9, p = 0.2]. The fact that the errors in the ASD group were greater than chance in the ToM condition (mean error rate = 81 %) suggests not only that the ASD participants were unable to interpret ToM animations as well as their TD peers (mean error rate = 49 %), but also that they were less likely to select a ToM interpretation than would be expected if they were just guessing. Thus, despite the failure to find a reliable interaction, participants in the ASD group were only outperformed by their TD peers in the ToM condition (see Additional file 1: Table S2) and their bias toward selecting non-ToM descriptions in the ToM condition clearly suggests a deficit in ToM among our sample of children and adolescents with ASD.
Distribution of fMRI activation
Within-group results
The processing of ToM animations, when contrasted with RD animations, showed significant activation in both groups in core areas of the ToM network, namely the precuneus/PCC, medial superior frontal gyrus extending to MPFC, and bilateral angular gyrus extending to TPJ, and in additional regions such as bilateral middle temporal gyrus and bilateral IFG and precentral gyrus (see Fig. 1a, b and Additional file 1: Table S3). Processing GD animations, when contrasted with RD animations, showed significant activation in both groups in the middle temporal gyrus and the right IFG (see Additional file 1: Figure S1 and Additional file 1: Table S4). As an additional feature, the HCP ToM activation map, thresholded at the same t value as our within-group results, is included in Fig. 1 for comparison (see Fig. 1c). Thus, the ToM task in the current study elicited strong activation in both groups in regions that are part of the ToM network and some regions that are considered part of action understanding.
Fig. 1.
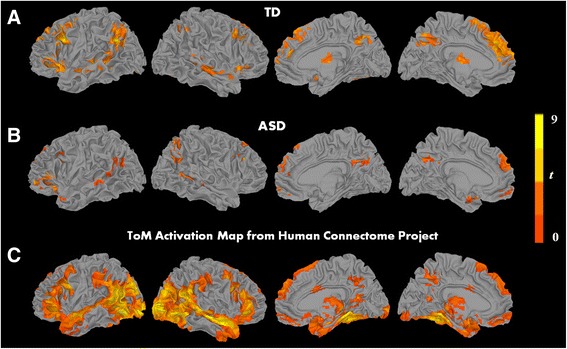
Significant clusters of within-group results for the contrast theory-of-mind vs. random animation (ToM > RD) for the a TD group, b ASD group, and c Human Connectome Project map (p < 0.05, FWE corr.)
Between-group results
Significant group differences were detected when processing ToM was contrasted with processing RD animations, with the ASD participants showing reduced activation when parameter estimates were compared: MPFC [t(24) = 3.30, p = 0.003], PCC [t(24) = 3.89, p = 0.0007], LTPJ [t(24) = 3.05, p = 0.005], LANG [t(24) = 2.27, p = 0.04], LIFG [t(24) = 2.89, p = 0.008], and LCEREB [t(24) = 3.46, p = 0.002], and no inverse effects (ASD > TD) were found (see Additional file 1: Figure S2).
Furthermore, the whole-brain analysis showed a pattern similar to the ROI analysis, where several left hemisphere areas showed reduced activation in the ASD group compared to the TD such as medial superior frontal gyrus extending to MPFC, precentral gyrus, precuneus/PCC, insula extending to IFG, thalamus, and angular gyrus extending to TPJ (see Fig. 2b, c and Table 2). There was also reduced activity in the cerebellum, specifically in left Crus V1. There was no region where the ASD participants showed greater activation relative to the TD participants in this contrast (i.e., there was no evidence of an ASD > TD effect, for the ToM vs. RD contrast). Additionally, there were no significant group differences in the modulation of activation while processing GD animations compared to the RD condition.
Fig. 2.

Significant clusters of between-group differences (TD > ASD) for the contrast theory-of-mind vs. random (ToM > RD; p < 0.05, FWE corr.), with dot plots showing individual data points for both TD and ASD groups
Table 2.
Group differences in fMRI BOLD activation for ToM vs. random animation and GD vs. random animation (whole-brain analysis)
| Contrast and direction | Region | Hemi. | Cluster vol. (in μl) | Peak coordinates MNI | Peak | ||
|---|---|---|---|---|---|---|---|
| x | y | z | t | ||||
| ToM vs. RD | Superior medial gyrus | L | 28,593 | −4 | 36 | 34 | 5.0 |
| TD > ASD | Insula/inferior frontal | L | 7371 | −24 | 22 | −12 | 4.1 |
| Precentral gyrus | L | 6453 | −40 | 4 | 34 | 5.0 | |
| Angular gyrus | L | 5427 | −42 | −56 | 40 | 4.0 | |
| Cerebellum (Crus VI) | L | 3213 | −22 | −86 | −26 | 4.8 | |
| Precuneus | L | 3024 | −6 | −44 | 40 | 3.7 | |
| Thalamus | R | 2835 | 2 | −8 | 18 | 4.3 | |
| ASD > TD | N/A | N/A | N/A | N/A | N/A | N/A | N/A |
| GD vs. RD | |||||||
| N/A | N/A | N/A | N/A | N/A | N/A | N/A | N/A |
L left, R right, Hemi. hemisphere, vol. volume
Functional connectivity
Functional connectivity analysis revealed a pattern of underconnectivity in the ASD group relative to the TD group in the following way: a two-way (group: TD and ASD) × 3 (condition: ToM, GD, and RD) repeated measures ANOVA was performed to assess overall functional connectivity (by averaging z-scores across all ROIs) and revealed no main effects of group [F(1, 24) = 0.03, p = 0.8] or condition [F(2, 48) = 0.65, p = 0.53]; however, the group × condition interaction was significant [F(2, 48) = 3.43, p = 0.04]. Each condition was examined separately, and overall functional connectivity was found to be significantly reduced in the ASD group compared to the TD group only for the ToM condition [t(24) = 2.8, p = 0.01] but not for GD and RD.
We further examined functional connectivity among anatomical networks (frontal, parietal, temporal, medial, and cerebellum) during the ToM condition (Fig. 3a), revealing significantly decreased connectivity in the ASD group relative to the TD in frontal-medial [t(24) = 3.90, p = .005], frontal-parietal [t(24) = 3.74, p = .001], and medial-cerebellum networks [t(24) = 2.80, p = .01]. It should be noted that this difference was not statistically significant during GD and RD conditions (see Fig. 3b–d), indicating the specificity of our findings of reduced functional connectivity in the ToM condition. We also examined the variability across participants in each group to make sure that the results are not influenced by an outlier or two. This is displayed in Additional file 1: Figure S3.
Fig. 3.

a Correlation matrix showing connectivity among frontal, medial, temporal, parietal, and cerebellum networks during ToM condition. Bar graphs for b frontal-medial, c frontal-parietal, and d medial-cerebellum connectivity during ToM, GD, and RD conditions (error bars represent SEM; *p < 0.1, uncorrected; **p < .05, uncorrected; ***p < .05, FDR corrected, for the test of the difference in connectivity between groups)
Discussion
This study of attributing mental states to animated shapes found robust activation in both ASD and TD groups of participants in regions associated with the ToM network. Additionally, this pattern of activation closely matched the results of an independent and larger dataset from the HCP, where a task similar to ours was used. Analysis of group differences, on the other hand, revealed reduced activation in ASD, relative to TD, participants during the ToM task. This supports the hypothesis of altered/reduced recruitment of the ToM network in ASD, a relatively consistent finding across a range of neuroimaging studies of ToM in autism [10, 20, 21, 23–26]. The activation in the ASD group was reduced in regions associated with processing ToM, including superior frontal gyrus extending to MPFC (SFG/MPFC), angular gyrus extending to TPJ and STS, and precuneus/PCC. Although our cluster of reduced activation in the ASD group in SFG/MPFC was found to be more superior and posterior, and not ventral, it lies very proximate to relatively superior areas in MPFC found in the HCP map. A separate cluster for TD > ASD was found more ventral in MPFC, although this cluster did not survive multiple comparisons correction. These findings of reduced activation in SFG/MPFC, TPJ, STS, and PCC are consistent with neuroimaging studies of ToM using these same animated stimuli in adults with ASD [21, 23]. Additionally, some of the areas of reduced activation in the ASD group for whole-brain analysis overlapped with regions from the HCP map. This was further confirmed as the HCP-based ROI analysis revealed findings similar to our whole-brain results. This underscores the role of these regions in processing ToM, as evidenced from multiple sources of neuroimaging data.
While significant activation in precentral/premotor cortex was observed in the ToM vs. RD contrast (e.g., coaxing) for both groups, this effect was less robust for the GD vs. RD contrast (e.g., chasing), possibly suggesting reliance on motor simulation to assist in attribution of mental states to moving objects. Premotor activations have been reported in many previous studies involving action and event prediction [56, 57]. The motor regions may encode the observable, perceptual-motor properties of others (in this case, the triangles dancing or pushing one another), and the ToM system may interpret those properties in terms of unobservable mental states and traits (the big triangle trying to coerce the little one to come out) [6, 58–60]. Additionally, reduced activation was also seen in this same area in the ASD participants, which may suggest limited understanding of the biological and kinematic properties of the animations presented. One possibility that our task elicited additional activation in regions outside of traditional ToM network is that there might be a functional communication among these regions in understanding ToM at richer and more comprehensive levels. This putative functional communication between the ToM network and regions such as the IFG has been observed in studies targeting mentalizing and mirroring abilities [24, 61], and in resting state fcMRI studies in ASD [62], possibly suggesting limited crosstalk among all these regions. It should also be noted that our observed pattern does not conclusively indicate that the ToM network depends on motor simulation for its inputs during high-level ToM processes. It is possible that these two systems may be functionally independent [63]. Further research is needed to directly test for functional interdependence of these systems during higher-level social cognition.
While our activation findings are largely consistent with previous studies, they are inconsistent with the findings from a recent fMRI study of ToM using verbally presented stories requiring drawing inferences with ASD participants in the same age range studied here [27]. These authors reported increased activation in these same regions (MPFC, TPJ, STS, and PCC) and in two different ASD groups (one with high and one with low ToM ability scores) compared to a TD group. They suggested that the age of the participants, the explicitness of the ToM task, or the mental effort required by the task could explain the hyperactivity of the ToM network in ASD they found. Comparison of our results with those of [27] argues against age being the critical factor affecting whether the participants with ASD show over- or under-activation of the ToM network, but we agree that factors related to the ToM task may explain the discrepancies among studies. Although our participants were explicitly instructed to think about what the triangles were doing, the attribution of mental states to the stimuli used here may still be a more implicit mentalizing process [64] than that necessary for the verbal reasoning required of the task used by [27]. Recent work has shown that performance in ToM tasks in people with ASD is dependent on whether the measure taps implicit or explicit processing and suggests that explicit verbal reasoning about the thoughts and feelings of others may be an important compensatory strategy in individuals with ASD [65]. For example, tasks that tap more into explicit mentalizing processing usually rely more on traditional false-belief tasks [66], where participants are asked to explain the behavior of a story character in a hypothetical scenario. Such processing (explicit) may involve greater recruitment of the ToM network and explain the hyperactivation reported by [27]. We also note that we have previously found increased ToM network activation in adults with ASD when they are required to make inferences about verbally presented stories [25], and that this over-activity in the network occurred regardless of whether the inferences required ToM or not. We suggest that the task used in the present study is sensitive to more implicit processes that are relatively automatic in TD individuals but absent or limited in those with ASD.
Another finding was the reduced activation in the cerebellum, specifically in Crus I, in the ASD group. One of the early accounts that shifted the traditional view of the cerebellum, in sensorimotor processing, was a study by Leiner and colleagues [67] where they presented neuroanatomical, neuroimaging, and behavioral reports of cerebellar involvement in cognitive and language functions. In addition, the cerebellum has been recently proposed to have an important role in social cognition [41]. Examples of tasks that typically trigger activation in this part of the cerebellum involve judgments of intentionality while observing animations of moving shapes [41]. In the current study, these were triangles whose motions evoked interpretations of intentional action/interaction involving thoughts and feelings (e.g., coaxing). The cerebellum has been largely overlooked in social cognition literature in ASD research, despite abnormalities previously reported both structurally and functionally. For example, postmortem studies have revealed abnormalities in Purkinje cells [68–70] and in vivo structural MRI studies have reported hypoplasia of the posterior vermis in ASD [71–73]. Functional MRI studies have found abnormalities in a wide range of tasks in ASD such as language [74], executive functioning paradigms [75], facial and vocal processing [76], and motor performance [77]. More recently, functional connectivity has been explored using both task-based and resting-state targeting low-frequency BOLD signal fluctuations in individuals with ASD [78, 79]. Mostofsky and colleagues found weaker functional connectivity between the cerebellum and motor areas during motor task performance, while Khan and colleagues found weaker functional connectivity between cerebellum and areas associated with higher cognitive functioning (prefrontal, parietal, and temporal) in ASD compared to TD participants. This is similar to the reduced connectivity between medial and cerebellar regions found in our study. The reduced cerebellar activation and connectivity in the ASD participants might underscore the role of the cerebellum in social cognition and suggest a deficit in how the cerebellum is recruited in social processing in ASD.
The finding of reduced functional connectivity in the ASD group is consistent with our previous study with adults using the same experimental paradigm [23] as well as other previous studies [21, 24]. The task-related functional connectivity analysis carried out here revealed significant underconnectivity in children and adolescents with ASD. This is consistent with many studies of people with ASD that have also reported reduced task-induced functional connectivity during processing that included a social or cognitive component (see [28], for an extensive review). As was true of the activation data, group differences in connectivity were observed only for the processing of ToM animations and not during RD or GD animations. This again suggests a fundamental deficit that is unique to ToM in individuals with ASD, and which may result from limited coordination of resources from key regions. Weaker communication among such regions in people with ASD may affect the quality of their ToM abilities, thus affecting the nature and quality of their social interactions [18]. Effective processing of ToM in this task may entail optimal and effective coordination (facilitated by high bandwidth) between relatively distant nodes of the mentalizing network in the TD group. In the ASD group, on the other hand, connectivity may be weaker due to bandwidth constraints between these regions. At the biological level, weaker bandwidth may arise from abnormal white matter trajectories in people with ASD, limiting the degree of synchronization, and this may be more apparent when participants with ASD are asked to perform tasks with high cognitive demand, such as ToM, face perception, language, working memory, and inhibition [28].
One of the limitations of the current study is the relatively smaller sample size, which may have limited our power for detecting stronger effects, in particular, group-by-condition interactions that would lend stronger support to the specificity of the effects to the ToM condition. In addition, the smaller sample size also restricts any analysis of subgroups within the ASD sample. Previous studies have indicated that individuals with higher or lower symptom severity tend to show different connectivity profiles [62, 80]. Nevertheless, the present findings are important, given the dearth of developmental neuroimaging studies in autism [35]. Although there is some evidence of functional over-connectivity in children with ASD [36], most such findings involve low-frequency fluctuations as opposed to an active cognitive task in the current study. In addition, most studies reporting underconnectivity in ASD have addressed domains such as cognitive (working memory, problem-solving, response inhibition), social (ToM, biological motion, face processing), and language (discourse processing, prosody, pun, irony, sentence comprehension, semantic processing), where individuals with ASD tend to show some degree of impairment. Methodological choices have also played a role in the disparity of findings while comparing children vs. adults [37]. Therefore, longitudinal studies tracking the same individuals as they progress from pre-to post-pubertal stages of development are needed to gain a better understanding of the neurodevelopmental trajectory in autism.
Conclusions
In summary, our results showed reduced activation in core ToM regions and non-traditional ToM regions, and reliable underconnectivity across several networks thought to be involved in ToM. The findings of this study provide valuable insights into the neurobiology of social cognition in autism, especially to the complex profile of brain activation and connectivity in children with autism. They also shed light on the disruption in brain functioning in general and connectivity in particular in autism when challenged by complex tasks like ToM.
Acknowledgements
This study was supported by the National Institute of Mental Health (R01-MH-29617 to M.A.J) and the McNulty-Civitan Scientist Award to RKK. The authors would like to thank Dr. Uta Frith, Dr. Francesca Happé, and Dr. Fulvia Castelli (King’s College, London, UK) for generously providing the animation stimuli for our study. We also thank Kaitlyn N. Rody for her comments on an earlier version of this manuscript. Finally, we would like to extend our sincerest appreciation to the individuals and families who generously gave their time and courage to participate in this neuroimaging study.
Abbreviations
- ADI
Autism Diagnostic Interview
- ADOS
Autism Diagnostic Observation Schedule
- ANOVA
analysis of variance
- ASD
autism spectrum disorder
- BOLD
blood-oxygen-level dependent
- fcMRI
functional connectivity MRI
- FDR
false discovery rate
- FMRI
functional magnetic resonance imaging
- FWHM
full width at half maximum
- GD
goal-directed
- GDQ
goal-directed question
- GLM
general linear model
- HCP
human connectome project
- IQ
intelligence quotient
- LANG
left angular gyrus
- LCEREB
left cerebellum
- LIFG
left inferior frontal gyrus
- LSTS
left superior temporal sulcus
- LTPJ
left temporoparietal junction
- MNI
Montreal Neurological Institute
- MPFC
medial prefrontal cortex
- PCC
posterior cingulate cortex
- RANG
right angular gyrus
- RCEREB
right cerebellum
- RD
random
- RDQ
random question
- RIFG
right inferior frontal gyrus
- RMSD
root mean square of displacement
- ROI
region of interest
- RSTS
right superior temporal sulcus
- RT
reaction time
- RTPJ
right temporoparietal junction
- TD
typically developing
- ToM
theory-of-mind
- ToMQ
ToM question
Additional file
Seed ROI coordinates for fcMRI analysis. Table S2. Reaction time and error rate for all experimental conditions. Table S3. fMRI BOLD activation for ToM vs. random animation. Table S4. fMRI BOLD activation for GD vs. random animation. Figure S1. Within-group results for the contrast goal-directed vs. random animation for the a) TD group, and b) ASD group (p < 0.05, FWE corr.). Figure S2. Mean dot plots of each ROI for both TD and ASD groups for the contrast theory-of-mind vs. random (*p < 0.1, uncorrected; **p < .05, uncorrected; ***p < .05, FDR corrected, for the test of the difference in activation between groups). Figure S3. Mean dot plots for B) frontal-medial, C) frontal-parietal, and D) medial-cerebellum connectivity during ToM, GD, and RD conditions (*p < 0.1, uncorrected; **p < .05, uncorrected; ***p < .05, FDR corrected, for the test of the difference in connectivity between groups).
Footnotes
Rajesh K. Kana and Jose O. Maximo contributed equally to this work.
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
RK participated in the study design, data collection, data analysis, and interpretation and manuscript preparation. JOM participated in the data analysis, interpretation, and manuscript preparation. MAJ, DLW, and TK participated in the study design, data collection, and manuscript preparation. VLC and SES participated in the data collection, data analysis, and manuscript preparation. NJM provided clinical and conceptual support for the project as well as participated in the manuscript preparation. All authors read and approved the final manuscript.
Contributor Information
Rajesh K. Kana, Phone: (205) 934-3171, Email: rkana@uab.edu
Jose O. Maximo, Email: omaximo@uab.edu
Diane L. Williams, Email: williamsd2139@duq.edu
Timothy A. Keller, Email: tk37@andrew.cmu.edu
Sarah E. Schipul, Email: ses@andrew.cmu.edu
Vladimir L. Cherkassky, Email: cherkassky@cmu.edu
Nancy J. Minshew, Email: minshewnj@upmc.edu
Marcel Adam Just, Email: just@cmu.edu.
References
- 1.Gallagher HL, Frith CD. Functional imaging of ‘theory of mind’. Trends Cogn Sci. 2003;7(2):77–83. doi: 10.1016/S1364-6613(02)00025-6. [DOI] [PubMed] [Google Scholar]
- 2.Barch DM, Burgess GC, Harms MP, Petersen SE, Schlaggar BL, Corbetta M, et al. Function in the human connectome: task-fMRI and individual differences in behavior. Neuroimage. 2013;80:169–89. doi: 10.1016/j.neuroimage.2013.05.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Van Overwalle F. Social cognition and the brain: a meta-analysis. Hum Brain Mapp. 2009;30(3):829–58. doi: 10.1002/hbm.20547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dufour N, Redcay E, Young L, Mavros PL, Moran JM, Triantafyllou C, et al. Similar brain activation during false belief tasks in a large sample of adults with and without autism. PLoS One. 2013;8(9):e75468. doi: 10.1371/journal.pone.0075468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Walter H, Schnell K, Erk S, Arnold C, Kirsch P, Esslinger C, et al. Effects of a genome-wide supported psychosis risk variant on neural activation during a theory-of-mind task. Mol Psychiatry. 2011;16(4):462–70. doi: 10.1038/mp.2010.18. [DOI] [PubMed] [Google Scholar]
- 6.de Lange FP, Spronk M, Willems RM, Toni I, Bekkering H. Complementary systems for understanding action intentions. Curr Biol. 2008;18(6):454–7. doi: 10.1016/j.cub.2008.02.057. [DOI] [PubMed] [Google Scholar]
- 7.Van der Cruyssen L, Van Duynslaeger M, Cortoos A, Van Overwalle F. ERP time course and brain areas of spontaneous and intentional goal inferences. Soc Neurosci. 2009;4(2):165–84. doi: 10.1080/17470910802253836. [DOI] [PubMed] [Google Scholar]
- 8.Cavanna AE, Trimble MR. The precuneus: a review of its functional anatomy and behavioural correlates. Brain. 2006;129(Pt 3):564–83. doi: 10.1093/brain/awl004. [DOI] [PubMed] [Google Scholar]
- 9.Buckner RL, Carroll DC. Self-projection and the brain. Trends Cogn Sci. 2007;11(2):49–57. doi: 10.1016/j.tics.2006.11.004. [DOI] [PubMed] [Google Scholar]
- 10.Lombardo MV, Chakrabarti B, Bullmore ET, MRC AIMS Consortium. Baron-Cohen S. Specialization of right temporo-parietal junction for mentalizing and its relation to social impairments in autism. Neuroimage. 2011;56(3):1832–8. doi: 10.1016/j.neuroimage.2011.02.067. [DOI] [PubMed] [Google Scholar]
- 11.CDC Prevalence of autism spectrum disorders—autism and developmental disabilities monitoring network, 11 sites, United States, 2010. MMWR Surveill Summ. 2014;63(2):1–21. [PubMed] [Google Scholar]
- 12.Kim YS, Leventhal BL, Koh YJ, Fombonne E, Laska E, Lim EC, et al. Prevalence of autism spectrum disorders in a total population sample. Am J Psychiatry. 2011;168(9):904–12. doi: 10.1176/appi.ajp.2011.10101532. [DOI] [PubMed] [Google Scholar]
- 13.Baron-Cohen S, Leslie AM, Frith U. Does the autistic child have a “theory of mind”? Cognition. 1985;21(1):37–46. doi: 10.1016/0010-0277(85)90022-8. [DOI] [PubMed] [Google Scholar]
- 14.Chevallier C, Kohls G, Troiani V, Brodkin ES, Schultz RT. The social motivation theory of autism. Trends Cogn Sci. 2012;16(4):231–9. doi: 10.1016/j.tics.2012.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Leekam SR, Perner J. Does the autistic child have a metarepresentational deficit? Cognition. 1991;40(3):203–18. doi: 10.1016/0010-0277(91)90025-Y. [DOI] [PubMed] [Google Scholar]
- 16.Perner J, Frith U, Leslie AM, Leekam SR. Exploration of the autistic child’s theory of mind: knowledge, belief, and communication. Child Dev. 1989;60(3):688–700. doi: 10.2307/1130734. [DOI] [PubMed] [Google Scholar]
- 17.Senju A. Spontaneous theory of mind and its absence in autism spectrum disorders. Neuroscientist. 2012;18(2):108–13. doi: 10.1177/1073858410397208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kana RK, Libero LE, Moore MS. Disrupted cortical connectivity theory as an explanatory model for autism spectrum disorders. Phys Life Rev. 2011;8(4):410–37. doi: 10.1016/j.plrev.2011.10.001. [DOI] [PubMed] [Google Scholar]
- 19.Assaf M, Hyatt CJ, Wong CG, Johnson MR, Schultz RT, Hendler T. Mentalizing and motivation neural function during social interactions in autism spectrum disorders. Neuroimage Clin. 2013;3:321–31. doi: 10.1016/j.nicl.2013.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Baron-Cohen S, Ring HA, Wheelwright S, Bullmore ET, Brammer MJ, Simmons A, et al. Social intelligence in the normal and autistic brain: an fMRI study. Eur J Neurosci. 1999;11(6):1891–8. doi: 10.1046/j.1460-9568.1999.00621.x. [DOI] [PubMed] [Google Scholar]
- 21.Castelli F, Frith C, Happé F, Frith U. Autism, Asperger syndrome and brain mechanisms for the attribution of mental states to animated shapes. Brain. 2002;125(Pt 8):1839–49. doi: 10.1093/brain/awf189. [DOI] [PubMed] [Google Scholar]
- 22.von dem Hagen EA, et al. Direct gaze elicits atypical activation of the theory-of-mind network in autism spectrum conditions. Cereb Cortex. 2013;24(06):1485–92. doi: 10.1093/cercor/bht003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kana RK, Keller TA, Cherkassky VL, Minshew NJ, Just MA. Atypical frontal-posterior synchronization of theory of mind regions in autism during mental state attribution. Soc Neurosci. 2009;4(2):135–52. doi: 10.1080/17470910802198510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kana RK, Libero LE, Hu CP, Deshpande HD, Colburn JS. Functional brain networks and white matter underlying theory-of-mind in autism. Soc Cogn Affect Neurosci. 2014;9(1):98–105. doi: 10.1093/scan/nss106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Mason RA, Williams DL, Kana RK, Minshew N, Just MA. Theory of mind disruption and recruitment of the right hemisphere during narrative comprehension in autism. Neuropsychologia. 2008;46(1):269–80. doi: 10.1016/j.neuropsychologia.2007.07.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Schulte-Ruther M, Greimel E, Markowitsch HJ, Kamp-Becker I, Remschmidt H, Fink GR, et al. Dysfunctions in brain networks supporting empathy: an fMRI study in adults with autism spectrum disorders. Soc Neurosci. 2011;6(1):1–21. doi: 10.1080/17470911003708032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.White SJ, Frith U, Rellecke J, Al-Noor Z, Gilbert SJ. Autistic adolescents show atypical activation of the brain’s mentalizing system even without a prior history of mentalizing problems. Neuropsychologia. 2014;56C:17–25. doi: 10.1016/j.neuropsychologia.2013.12.013. [DOI] [PubMed] [Google Scholar]
- 28.Just MA, Keller TA, Malave VL, Kana RK, Varma S. Autism as a neural systems disorder: a theory of frontal-posterior underconnectivity. Neurosci Biobehav Rev. 2012;36(4):1292–313. doi: 10.1016/j.neubiorev.2012.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Muller RA, Shih P, Keehn B, Deyoe JR, Leyden KM, Shukla DK. Underconnected, but how? A survey of functional connectivity MRI studies in autism spectrum disorders. Cereb Cortex. 2011;21(10):2233–43. doi: 10.1093/cercor/bhq296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Vissers ME, Cohen MX, Geurts HM. Brain connectivity and high functioning autism: a promising path of research that needs refined models, methodological convergence, and stronger behavioral links. Neurosci Biobehav Rev. 2012;36(1):604–25. doi: 10.1016/j.neubiorev.2011.09.003. [DOI] [PubMed] [Google Scholar]
- 31.Schipul SE, Keller TA, Just MA. Inter-regional brain communication and its disturbance in autism. Front Syst Neurosci. 2011;5:10. doi: 10.3389/fnsys.2011.00010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Maximo JO, Cadena EJ, Kana RK. The implications of brain connectivity in the neuropsychology of autism. Neuropsychol Rev. 2014;24(1):16–31. doi: 10.1007/s11065-014-9250-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Happe F, Ronald A, Plomin R. Time to give up on a single explanation for autism. Nat Neurosci. 2006;9(10):1218–20. doi: 10.1038/nn1770. [DOI] [PubMed] [Google Scholar]
- 34.Hahamy A, Behrmann M, Malach R. The idiosyncratic brain: distortion of spontaneous connectivity patterns in autism spectrum disorder. Nat Neurosci. 2015;18(2):302–9. doi: 10.1038/nn.3919. [DOI] [PubMed] [Google Scholar]
- 35.Uddin LQ, Supekar K, Menon V. Reconceptualizing functional brain connectivity in autism from a developmental perspective. Front Hum Neurosci. 2013;7:458. doi: 10.3389/fnhum.2013.00458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Supekar K, Uddin LQ, Khouzam A, Phillips J, Gaillard WD, Kenworthy LE, et al. Brain hyperconnectivity in children with autism and its links to social deficits. Cell Rep. 2013;5(3):738–47. doi: 10.1016/j.celrep.2013.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Nair A, Keown CL, Datko M, Shih P, Keehn B, Müller RA. Impact of methodological variables on functional connectivity findings in autism spectrum disorders. Hum Brain Mapp. 2014;35(8):4035–48. doi: 10.1002/hbm.22456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.White SJ, Coniston D, Rogers R, Frith U. Developing the frith-happe animations: a quick and objective test of theory of mind for adults with autism. Autism Res. 2011;4(2):149–54. doi: 10.1002/aur.174. [DOI] [PubMed] [Google Scholar]
- 39.Lahnakoski JM, Glerean E, Salmi J, Jääskeläinen IP, Sams M, Hari R, et al. Naturalistic FMRI mapping reveals superior temporal sulcus as the hub for the distributed brain network for social perception. Front Hum Neurosci. 2012;6:233. doi: 10.3389/fnhum.2012.00233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Seghier ML. The angular gyrus: multiple functions and multiple subdivisions. Neuroscientist. 2013;19(1):43–61. doi: 10.1177/1073858412440596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Van Overwalle F, Baetens K, Mariën P, Vandekerckhove M. Social cognition and the cerebellum: a meta-analysis of over 350 fMRI studies. Neuroimage. 2014;86:554–72. doi: 10.1016/j.neuroimage.2013.09.033. [DOI] [PubMed] [Google Scholar]
- 42.Wechsler D. Wechsler abbreviated scale of intelligence. San Antonio, TX: Psychological Corporation; 1999. [Google Scholar]
- 43.American Psychiatric Association . Diagnostic and statistical manual of mental disorders (4th ed., Text Revision) Washington, DC: Author; 2000. [Google Scholar]
- 44.Lord C, Rutter M, Le Couteur A. Autism diagnostic interview-revised: a revised version of a diagnostic interview for caregivers of individuals with possible pervasive developmental disorders. J Autism Dev Disord. 1994;24(5):659–85. doi: 10.1007/BF02172145. [DOI] [PubMed] [Google Scholar]
- 45.Lord C, Risi S, Lambrecht L, Cook EH, Jr, Leventhal BL, DiLavore PC, et al. The autism diagnostic observation schedule-generic: a standard measure of social and communication deficits associated with the spectrum of autism. J Autism Dev Disord. 2000;30(3):205–23. doi: 10.1023/A:1005592401947. [DOI] [PubMed] [Google Scholar]
- 46.Abell F, Happe F, Frith U. Do triangles play tricks? Attribution of mental states to animated shapes in normal and abnormal development. J Cogn Dev. 2000;15:1–20. doi: 10.1016/S0885-2014(00)00014-9. [DOI] [Google Scholar]
- 47.Cox RW. AFNI: software for analysis and visualization of functional magnetic resonance neuroimages. Comput Biomed Res. 1996;29(3):162–73. doi: 10.1006/cbmr.1996.0014. [DOI] [PubMed] [Google Scholar]
- 48.Smith SM, Jenkinson M, Woolrich MW, Beckmann CF, Behrens TE, Johansen-Berg H, et al. Advances in functional and structural MR image analysis and implementation as FSL. Neuroimage. 2004;23(Suppl 1):S208–19. doi: 10.1016/j.neuroimage.2004.07.051. [DOI] [PubMed] [Google Scholar]
- 49.Jenkinson M, Bannister P, Brady M, Smith S. Improved optimization for the robust and accurate linear registration and motion correction of brain images. Neuroimage. 2002;17(2):825–41. doi: 10.1006/nimg.2002.1132. [DOI] [PubMed] [Google Scholar]
- 50.Jenkinson M, Smith S. A global optimisation method for robust affine registration of brain images. Med Image Anal. 2001;5(2):143–56. doi: 10.1016/S1361-8415(01)00036-6. [DOI] [PubMed] [Google Scholar]
- 51.Gorgolewski KJ, Varoquaux G, Rivera G, Schwarz Y, Ghosh SS, Maumet C, et al. NeuroVault.org: a web-based repository for collecting and sharing unthresholded statistical maps of the human brain. Front Neuroinform. 2015;9:8. doi: 10.3389/fninf.2015.00008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zhang Y, Brady M, Smith S. Segmentation of brain MR images through a hidden Markov random field model and the expectation-maximization algorithm. IEEE Trans Med Imaging. 2001;20(1):45–57. doi: 10.1109/42.906424. [DOI] [PubMed] [Google Scholar]
- 53.Satterthwaite TD, Elliott MA, Gerraty RT, Ruparel K, Loughead J, Calkins ME, et al. An improved framework for confound regression and filtering for control of motion artifact in the preprocessing of resting-state functional connectivity data. Neuroimage. 2013;64:240–56. doi: 10.1016/j.neuroimage.2012.08.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Van Dijk KR, Sabuncu MR, Buckner RL. The influence of head motion on intrinsic functional connectivity MRI. Neuroimage. 2012;59(1):431–8. doi: 10.1016/j.neuroimage.2011.07.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Power JD, Barnes KA, Snyder AZ, Schlaggar BL, Petersen SE. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. Neuroimage. 2012;59(3):2142–54. doi: 10.1016/j.neuroimage.2011.10.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Schubotz RI. Prediction of external events with our motor system: towards a new framework. Trends Cogn Sci. 2007;11(5):211–8. doi: 10.1016/j.tics.2007.02.006. [DOI] [PubMed] [Google Scholar]
- 57.Schubotz RI, von Cramon DY. Sequences of abstract nonbiological stimuli share ventral premotor cortex with action observation and imagery. J Neurosci. 2004;24(24):5467–74. doi: 10.1523/JNEUROSCI.1169-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Hamilton AF. Reflecting on the mirror neuron system in autism: a systematic review of current theories. Dev Cogn Neurosci. 2013;3:91–105. doi: 10.1016/j.dcn.2012.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Marsh LE, Hamilton AF. Dissociation of mirroring and mentalising systems in autism. Neuroimage. 2011;56(3):1511–9. doi: 10.1016/j.neuroimage.2011.02.003. [DOI] [PubMed] [Google Scholar]
- 60.Thioux M, Gazzola V, Keysers C. Action understanding: how, what and why. Curr Biol. 2008;18(10):R431–4. doi: 10.1016/j.cub.2008.03.018. [DOI] [PubMed] [Google Scholar]
- 61.Libero LE, Maximo JO, Deshpande HD, Klinger LG, Klinger MR, Kana RK. The role of mirroring and mentalizing networks in mediating action intentions in autism. Mol Autism. 2014;5(1):50. doi: 10.1186/2040-2392-5-50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Fishman I, Keown CL, Lincoln AJ, Pineda JA, Müller RA. Atypical cross talk between mentalizing and mirror neuron networks in autism spectrum disorder. JAMA Psychiatry. 2014;71(7):751–60. doi: 10.1001/jamapsychiatry.2014.83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Van Overwalle F, Baetens K. Understanding others’ actions and goals by mirror and mentalizing systems: a meta-analysis. Neuroimage. 2009;48(3):564–84. doi: 10.1016/j.neuroimage.2009.06.009. [DOI] [PubMed] [Google Scholar]
- 64.Frith CD, Frith U. Implicit and explicit processes in social cognition. Neuron. 2008;60(3):503–10. doi: 10.1016/j.neuron.2008.10.032. [DOI] [PubMed] [Google Scholar]
- 65.Schuwerk T, Vuori M, Sodian B. Implicit and explicit theory of mind reasoning in autism spectrum disorders: the impact of experience. Autism. 2014;19:459–68. doi: 10.1177/1362361314526004. [DOI] [PubMed] [Google Scholar]
- 66.Baron-Cohen S, Lombardo M, Tager-Flusberg H. Understanding other minds: perspectives from developmental social neuroscience. Oxford: Oxford University Press; 2013. [Google Scholar]
- 67.Leiner HC, Leiner AL, Dow RS. Cognitive and language functions of the human cerebellum. Trends Neurosci. 1993;16(11):444–7. doi: 10.1016/0166-2236(93)90072-T. [DOI] [PubMed] [Google Scholar]
- 68.Fatemi SH, Halt AR, Realmuto G, Earle J, Kist DA, Thuras P, et al. Purkinje cell size is reduced in cerebellum of patients with autism. Cell Mol Neurobiol. 2002;22(2):171–5. doi: 10.1023/A:1019861721160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Whitney ER, Kemper TL, Rosene DL, Bauman ML, Blatt GJ. Density of cerebellar basket and stellate cells in autism: evidence for a late developmental loss of Purkinje cells. J Neurosci Res. 2009;87(10):2245–54. doi: 10.1002/jnr.22056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Bauman ML, Kemper TL. Neuroanatomic observations of the brain in autism: a review and future directions. Int J Dev Neurosci. 2005;23(2–3):183–7. doi: 10.1016/j.ijdevneu.2004.09.006. [DOI] [PubMed] [Google Scholar]
- 71.Courchesne E, Campbell K, Solso S. Brain growth across the life span in autism: age-specific changes in anatomical pathology. Brain Res. 2011;1380:138–45. doi: 10.1016/j.brainres.2010.09.101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Courchesne E, Yeung-Courchesne R, Press GA, Hesselink JR, Jernigan TL. Hypoplasia of cerebellar vermal lobules VI and VII in autism. N Engl J Med. 1988;318(21):1349–54. doi: 10.1056/NEJM198805263182102. [DOI] [PubMed] [Google Scholar]
- 73.Riva D, Annunziata S, Contarino V, Erbetta A, Aquino D, Bulgheroni S. Gray matter reduction in the vermis and CRUS-II is associated with social and interaction deficits in low-functioning children with autistic spectrum disorders: a VBM-DARTEL Study. Cerebellum. 2013;12(5):676–85. doi: 10.1007/s12311-013-0469-8. [DOI] [PubMed] [Google Scholar]
- 74.Harris GJ, Chabris CF, Clark J, Urban T, Aharon I, Steele S, et al. Brain activation during semantic processing in autism spectrum disorders via functional magnetic resonance imaging. Brain Cogn. 2006;61(1):54–68. doi: 10.1016/j.bandc.2005.12.015. [DOI] [PubMed] [Google Scholar]
- 75.Solomon M, Ozonoff SJ, Ursu S, Ravizza S, Cummings N, Ly S, et al. The neural substrates of cognitive control deficits in autism spectrum disorders. Neuropsychologia. 2009;47(12):2515–26. doi: 10.1016/j.neuropsychologia.2009.04.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Wang AT, Lee SS, Sigman M, Dapretto M. Reading affect in the face and voice: neural correlates of interpreting communicative intent in children and adolescents with autism spectrum disorders. Arch Gen Psychiatry. 2007;64(6):698–708. doi: 10.1001/archpsyc.64.6.698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Allen G, Muller RA, Courchesne E. Cerebellar function in autism: functional magnetic resonance image activation during a simple motor task. Biol Psychiatry. 2004;56(4):269–78. doi: 10.1016/j.biopsych.2004.06.005. [DOI] [PubMed] [Google Scholar]
- 78.Khan AJ, Nair A, Keown CL, Datko MC, Lincoln AJ, Müller RA. Cerebro-cerebellar resting-state functional connectivity in children and adolescents with autism spectrum disorder. Biol Psychiatry. 2015;78(9):625–34. doi: 10.1016/j.biopsych.2015.03.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Mostofsky SH, Powell SK, Simmonds DJ, Goldberg MC, Caffo B, Pekar JJ. Decreased connectivity and cerebellar activity in autism during motor task performance. Brain. 2009;132(Pt 9):2413–25. doi: 10.1093/brain/awp088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Keown CL, Shih P, Nair A, Peterson N, Mulvey ME, Müller RA. Local functional overconnectivity in posterior brain regions is associated with symptom severity in autism spectrum disorders. Cell Rep. 2013;5(3):567–72. doi: 10.1016/j.celrep.2013.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]