

Abstract
The intrinsic growth ability of all the neurons declines during development although some may grow better than others. Numerous intracellular signaling proteins and transcription factors have been shown to regulate the intrinsic growth capacity in mature neurons. Among them, PI3 kinase/Akt pathway is important for controlling axon elongation. As a negative regulator of this pathway, the tumor suppressor phosphatase and tensin homolog (PTEN) appears critical to control the regenerative ability of young and adult neurons. This review will focus on recent research progress in axon regeneration and neural repair by PTEN inhibition and therapeutic potential of blocking this phosphatase for neurological disorders. Inhibition of PTEN by deletion in conditional knockout mice, knockdown by short-hairpin RNA, or blockade by pharmacological approaches, including administration of selective PTEN antagonist peptides, stimulates various degrees of axon regrowth in juvenile or adult rodents with central nervous system injuries. Importantly, post-injury PTEN suppression could enhance axonal growth and functional recovery in adult central nervous system after injury.
Keywords: PTEN inhibition, antagonist peptide, spinal cord injury, intrinsic growth capacity, axon regeneration, functional recovery
Introduction
Following central nervous system (CNS) injuries, the incapability of axons to regenerate is due to the combination of a non-permissive extrinsic CNS environment (McGee and Strittmatter, 2003; Liu et al., 2006; Busch and Silver, 2007) and a loss of intrinsic regenerative ability during development (Goldberg et al., 2002; Lu et al., 2014). A great number of studies identified various cell types and molecules that inhibit neuronal elongation in adult CNS. The components of CNS myelin, glial scar tissues and certain guidance cues inhibit axon elongation and partially contribute to the CNS regeneration failure. Removing or blocking inhibitory activities of negative molecules, such as myelin or scar associated inhibitors, induces limited degree of axon regeneration in vivo. On the other hand, the intrinsic growth ability of all the neurons declines during development although some neuronal types may grow better than others. Numerous factors could regulate the intrinsic growth capacity, including certain transcription factors, such as cAMP-responsive element binding protein (CREB), signal transducer and activator of transcription 3 (STAT3), nuclear factor of activated T cell (NFAT), c-Jun activating transcription factor 3 (AFT3) and Krüppel-like factors (KLFs), and intracellular signaling proteins, such as PI3 kinase, Akt, phosphatase and tensin homolog (PTEN), suppressor of cytokine signaling 3 (SOCS3), B-RAF, dual leucine zipper kinase (DLK), and insulin/insulin-like growth factor-1 (IGF-1) signaling (Moore and Goldberg, 2011; Byrne et al., 2014; Lu et al., 2014). Although targeting each of these signals could enhance axon growth (Moore and Goldberg, 2011), deletion of the tumor suppressor PTEN in conditional knockout (KO) mice appears to result in most dramatic regrowth of CNS axons after injuries (Park et al., 2008; Liu et al., 2010), suggesting that PTEN/mammalian target of rapamycin (mTOR) signaling is critical to regulate the intrinsic regenerative ability of young and adult neurons (Figure 1) (Park et al., 2010; Lu et al., 2014). Further studies on PTEN knockdown by short-hairpin RNA (shRNA) or blockade by pharmacological approaches demonstrate various degrees of axon regrowth in adult rodents with CNS injuries (Zukor et al., 2013; Lewandowski and Steward, 2014; Ohtake et al., 2014). This review will update recent research progress in axon regeneration and neural repair by PTEN inhibition (Table 1) and discuss the therapeutic potential of blocking this phosphatase for neurological disorders.
Figure 1.
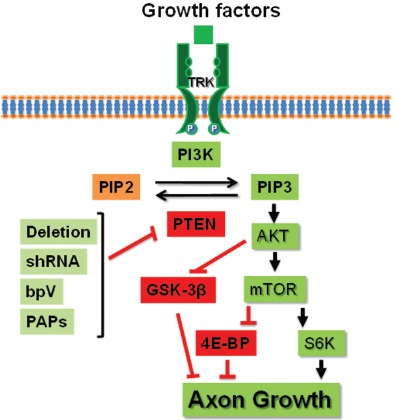
Schematic of Akt/PTEN pathway that regulates neuronal growth and axon regeneration by PTEN inhibition.
Various growth factors, such as nerve growth factor, activate their receptors (especially the tyrosine receptor kinases (TRK)) and PI3K pathway and stimulate neuronal growth by enchaining mTOR activity and suppressing GSK-3β signal. In contrast, intracellular PTEN phos-phatase blocks axon growth by inactivating Akt signal and mTOR path-way. PTEN inhibition by a number of approaches, including deletion in knockout (KO) mice, knockdown by shRNA, or pharmacological blockade with phosphatase inhibitor bpV or selective antagonists PTEN antagonist peptides, promotes neuronal regeneration.
PTEN: Phosphatase and tensin homolog; 4E-BP: 4E-binding protein; bpV: bisperoxovanadium; PIP2: phosphatidylinositol 4,5-bisphosphate; PIP3: phosphatidylinositol 3,4,5 trisphosphate; PI3K: phosphoinositide 3-kinase; GSK-3β: glycogen synthase kinase 3β; S6K: S6 kinase; PAPs: PTEN antagonist peptides.
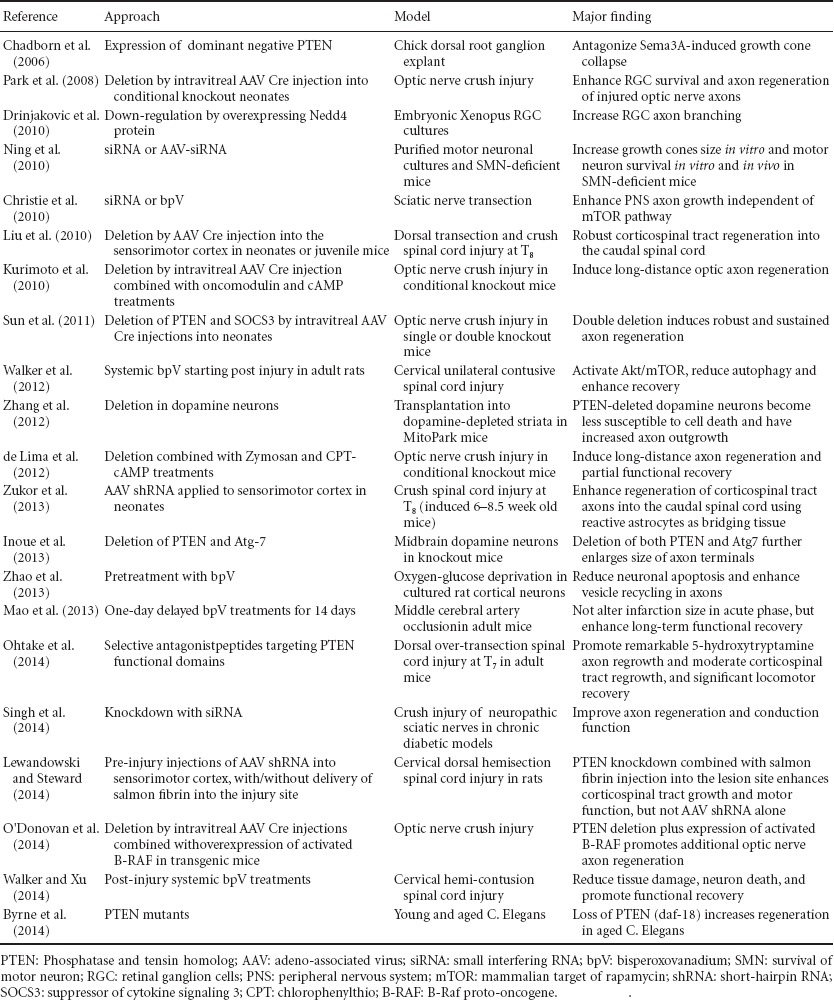
Table 1.
Summary of PTEN inhibition on axon growth and neuroprotection
PTEN Deletion and CNS Regeneration and Neural Repair
PI3K/Akt pathway plays a critical role in regulating axon formation and extension (Zhou and Snider, 2006). Expression of constitutively active Akt in embryonic chick dorsal root ganglion (DRG) neurons increases axon branching, cell hypertrophy and growth cone expansion (Grider et al., 2009). Overexpression of active Akt by adeno-associated virus (AAV) vector remarkably promotes regrowth and reinnervation of lesioned dopaminergic axons and partial behavioral recovery in vivo (Kim et al., 2011). Activating Akt signaling also enhances axon regeneration of Drosophila CNS neurons (Song et al., 2012).
Given that PTEN negatively mediates Akt activity by dephosphorylating phosphoinositide substrates, PTEN suppression is likely to increase axon growth by enhancing activity of PI3K/Akt signaling. Recent studies on neuronal PTEN inactivation by transgenic deletion demonstrate enhanced regeneration of lesioned CNS axons. Intravitreal injection of AAV Cre recombinase enhanced survival of retinal ganglion cells (RGCs) and promoted considerable regeneration of injured optic nerve axons in juvenile mice (Park et al., 2008). Deletion of PTEN by injection of AAV-Cre into the sensorimotor cortex in conditional KO mice induces substantial regrowth of lesioned corticospinal tract (CST) axons and formation of synapse-like structures in the caudal spinal cord of juvenile or adult mice with spinal cord injury (SCI) (Liu et al., 2010). Because treatment with rapamycin, an mTOR inhibitor, abolishes the growth promoting-effect of PTEN deletion (Park et al., 2010), mTOR activation appears critical to control axon growth downstream of PTEN. Simultaneous deletion of PTEN and SOCS3, a negative regulator of Janus kinase (JAK)/STAT pathway, results in more robust and sustained axon regeneration, suggesting that two proteins regulate regenerative programs through distinct mechanisms (Sun et al., 2011). PTEN and SOCS3 double deletion upregulates mTOR activators, such as small GTPaseRheb and IGF-1, in injured RGCs. PTEN deletion combined with overexpression of an active form of B-RAF kinase, a known signal downstream of neurotrophic factors, stimulates additive regeneration of lesioned optic axons (O’Donovan et al., 2014). In addition, simultaneous deletion of PTEN with autophagy-related protein 7 (Atg7), which regulates vacuole transport and autophagy in cytoplasm, increases axon terminal enlargement in midbrain dopamine neurons compared to Atg7 deletion alone (Inoue et al., 2013). Transplanted PTEN-deficient dopamine neurons into mice with Parkinson's disease models were less susceptible to cell death and extended longer axons than control grafts (Zhang et al., 2012). Together, PTEN appears important to restrict regeneration of mature neurons and its inactivation may have therapeutic potential for CNS disorders characterized by axonal damages.
PTEN Knockdown with shRNA and CNS Regeneration
shRNA makes a tight hairpin turn and is frequently used to silence target gene expression by RNA interference. Injections of AAV vector encoding shRNA-PTEN into the motor cortex in neonatal mice significantly reduced expression of PTEN protein and enhanced levels of phosphorylated S-6 kinase, a downstream signal of mTOR in neurons (Zukor et al., 2013). Injections of viral shRNA-PTEN into the sensorimotor cortex of neonates could sufficiently enhance the intrinsic growth of CST neurons and induce CST regrowth in the caudal spinal cord of mice with a crush injury at T8 (induced at 6–8.5 weeks old). Some CST axons crossed the lesion area using reactive astrocytic tissues as the bridging tissue although CST sprouts avoided dense clusters of fibroblasts and macrophages around the lesion. The other group generated a similar viral shRNA-PTEN and efficiently knocked down PTEN protein (Lewandowski and Steward, 2014). Injection of AAV shRNA-PTEN into the motor cortex in adult rats 1 week before a dorsal hemisection injury at C6 did not significantly promote CST regrowth in the caudal spinal cord and locomotor function recovery although some biotinylated dextran amine (BDA)-traced CST axons reached the lesion edge in shRNA-PTEN treated animals. However, shRNA-PTEN plus delivery of salmon fibrin into the injury area significantly increased the number of BDA-labeled CST axons in the caudal spinal cord and forelimb-reaching scores.
Together, PTEN knockdown by pre-injury injection of shRNA stimulates regrowth of injured CST axons in SCI mice, but it has minimal effect in SCI rats. PTEN inhibition combined with other strategies, such as those targeting other intracellular signals or extrinsic factors responsible for regeneration failure, may become more efficient for promoting axon elongation. Notably, it is very important to study whether knockdown of PTEN by viral shRNA-PTEN delivered post-injury stimulates axon regrowth and improves functional recovery after CNS injury because the pre-injury viral vector applications used in previous studies are not clinically translational.
Pharmacological PTEN Inhibition and Neuroprotection and Axon Regeneration
PTEN genetic deletion in KO mice and knockdown by pre-injury injection of shRNA are not feasible for clinical treatment. In contrast, suppression of PTEN by a pharmacological method is highly controllable in initiation time, application period and drug dosage. Bisperoxovanadium (bpV) compounds are inhibitors of several protein tyrosine phosphatases (PTPs) with selectivity for PTEN (IC50 = ~40 nM), but also block other PTPs (such as PTPβ) at higher nM levels. bpV treatment is neuroprotective after different CNS injuries. Systemic bpV initiated immediately after injury (for 7 days), increased spared white matter at the lesion area, numbers of oligodendrocytes and motor neuron area, and improved functional recovery in rats with contusive cervical SCI (Walker et al., 2012; Walker and Xu, 2014). bpV enhanced axon outgrowth of primary cortical neuronal cultures following oxygen-glucose deprivation. Although 1-day delayed treatment with bpV (for 14 days) did not reduce infarction size in mice with middle cerebral artery occlusion in acute stage, they significantly improved long-term functional recovery after ischemia (Mao et al., 2013). Thus, post-injury bpV treatments exhibit beneficial effects on recovery after CNS injuries although the molecular basis for bpV actions is less clear because bpVs target other PTPs as well as PTEN.
Suppressing PTEN activity is very promising for promoting CNS axon regeneration and neural repair, but transgenic approaches used in previous studies block PTEN before injuries and only target a single neuronal population by local injections of AAV-Cre/AAV-shRNA, not the diffusely-dispersed neurons, such as 5-hydroxytryptamine (5-HT) neurons or propriospinal interneurons known to contribute to functional recovery after SCI (Barbeau and Rossignol, 1991; Ribotta et al., 2000; Li et al., 2004; Courtine et al., 2008). Application of bpV may block PTEN action (Christie et al., 2010), but bpV may result in clinical side effects by interacting with other PTPs (Scrivens et al., 2003), such as lowering blood glucose by activating insulin signaling (Drake and Posner, 1998). It is important to develop a pharmacological method to suppress PTEN activity efficiently and selectively. We recently identified selective antagonist peptides for PTEN by targeting its critical functional domains, including PIP2, ATP-type, PDZ and C-terminal tail domains (Ohtake et al., 2014). We demonstrated that those PTEN antagonist peptides (PAPs) bound COS7 cells that over-expressed PTEN protein, promoted neurite outgrowth in vitro and blocked several signals downstream of PTEN. Systemic PAP treatments by subcutaneous delivery (initiated 2 days after injury, 2-week treatment) stimulated regrowth of descending serotonergic axons in the caudal spinal cord of adult mice with dorsal over-hemisection at T7. Systemic PAPs also enhanced sprouting of CST axons rostral to the lesion and limited regrowth of CST axons in the caudal spinal cord. Importantly, systemic PAPs enhanced recovery of locomotor function several weeks after SCI by increasing BMS locomotor scores and stride length of hindlimbs and reducing grid walk errors. Thus, systemic delivery of small peptides after injury selectively blocks PTEN activity and promotes regrowth of injured multiple CNS axonal tracts. The peptide approach shows benefits in several areas where the transgenic (such as KO) and invasive approaches (such as local chondroitinase and transplants) are highly deficient and may thus facilitate development of a successful therapy for CNS axonal injuries. Constantly, our other studies demonstrated great therapeutic potential of small peptides that selectively block other axon growth inhibitory molecules (GrandPre et al., 2002; Li and Strittmatter, 2003; Fisher et al., 2011).
PTEN Suppression and Peripheral Nervous System (PNS) Regeneration and Axon Myelination
PNS axons exhibit remarked regrowth after injuries, but their regeneration may not be optimal and many patients suffer from long-term or persistent functional impairment after PNS axon lesions, including motor and sensory loss, chronic pain and inappropriate autonomic responses. PTEN is expressed in PNS neurons (such as DRGs) and contributes to their growth cone collapse and reduced axon growth (Chadborn et al., 2006). PTEN knockdown enhanced regeneration of lesioned sciatic nerve and a preconditioning lesion exhibited an additive effect. PTEN knockdown by bpV treatment or local delivery of siRNA accelerated sciatic axon outgrowth in vivo (Christie et al., 2010). PTEN also restricts PNS regeneration in mouse models of diabetes. Following crush of neuropathic sciatic nerves in diabetes models, local delivery of PTEN short interfering RNA increased the number of regenerating myelinated axons distal to injury site, promoted reinnervation of skin by unmyelinated epidermal axons and contributed to recovery of mechanical sensation. Inhibition of PTEN or activation of PI3K enhances growth of DRG neurons probably by mTOR-independent mechanisms because they respond to GSK-3β (also a downstream signal of PI3K) and transcription factor Smad1, but not to mTOR blockade by rapamycin (Zou et al., 2009; Christie et al., 2010; Hur et al., 2011).
PTEN and PI3K activities also regulate function of oligodendrocytes and Schwann cells and myelination during development. PTEN deletion enhanced the numbers of Schwann cells and myelinated small axons with caliber < 1 μm. PTEN deletion in oligodendrocytes induced PIP3-dependent hypermyelination of CNS axons (Goebbels et al., 2010). Interactions between PTEN and trafficking protein Dlg1 (disks large homolog 1) are able to inhibit axonal myelination and this mechanism limits myelin sheath thickness and prevents over-myelination in mouse sciatic nerves (Cotter et al., 2010).
Molecular Targets that Regulate Axon Regeneration along Akt Pathway
Following stimulation by various extracellular growth factors, activation of the receptor tyrosine kinase and/or G-protein coupled receptor activates PI3K and subsequently Akt by increasing the generation of PIP3. Activated Akt phosphorylates tuberous sclerosis protein complex (TSC) and inhibits its activity. Through enhancing Rheb activity, suppression of TSC increases the levels of activated mTOR (Park et al., 2010; Walker et al., 2013), which phosphorylates ribosomal protein p70S6 kinase and 4E binding protein-1 (4E-BP1) and regulatesprotein synthesis and cell growth (Proud, 2002). In particular, phosphorylated p70S6K enhances phosphorylation and activity of ribosomal protein S6 and initiates various translation activities required for cell growth. Phosphorylation of 4E-BP1 reduces its activity and its inhibition on protein synthesis and thus promotes neuronal elongation. Moreover, Akt activation critically regulates GSK-3β signaling by phosphorylating this kinase (at serine 9) and subsequently inactivating it (Figure 1) (Cross et al., 1995). Localized GSK-3β inactivation is required for establishing and maintaining neuronal polarity (Jiang et al., 2005; Yoshimura et al., 2005). GSK-3β inactivation at growth cone is able to stimulate axon formation and extension and to convert dendritic processes into axons in polarized neurons. Consistently, GSK-3β inhibitors enhance neurite outgrowth in vitro and regrowth of injured axons after CNS injury (Zhou et al., 2004; Zhou and Snider, 2005; Dill et al., 2008). Therefore, GSK-3β suppression at growth cone is essential for promoting microtubule assembly in axons (Zhou and Snider, 2006; Hur and Zhou, 2010) although inhibiting GSK-3β may also block axon growth (Alabed et al., 2010; Gobrecht et al., 2014). A further study suggests that GSK-3β inhibition can both enhance and prevent axon growth depending on the substrates involved, enhancing axon elongation if towards primed substrates (such as CRMP2 and adenomatous polyposis coli) and preventing axon growth if toward unprimed substrates (such as microtubule-associated protein 1B) (Kim et al., 2006; Hur and Zhou, 2010). Therefore, GSK-3β activity should be finely tuned to stimulate axon regeneration, including its reduced activity towards one subset of substrates and preserved activity towards other substrates.
Prospective
PI3K/Akt/PTEN pathways play critical roles in regulating neural development and PTEN suppression may have great potential for promoting CNS axon regeneration and neural repair. Many unknown issues and challenges, however, remain regarding PTEN-mediated growth inhibition and its translating potential to clinical applications. Several neuronal populations are known to be sensitive to PTEN blockade, including RGC, CST, 5-HT and DRGs. Do the other types of neurons similarly respond to PTEN inhibition? PTEN is present in both neuronal soma and axonal compartment during axon extension (including the growth cones) (Chadborn et al., 2006). Does PTEN activation in both subcellular structures mediate axon elongation? Sustained and long-distance axon regeneration is crucial for meaningful functional recovery after many CNS injuries, including SCI and optic neuropathy. PTEN deletion induces remarkable axon elongation, but axon regrowth is usually limited to < 1 mm from the lesion. PTEN suppression combined with other strategies, such as regulating inflammatory responses and activities of other signaling pathways (such as SOCS3/STAT/Erk) or transcriptional factors (such as KLFs and CREB), as well as surmounting scar-mediated inhibition, may induce more robust and distant axon regeneration. Obviously, post-injury intervention of PTEN by pharmacological approaches, including selective blockade by small peptides, is promising and highly relevant to future translation into human use. Targeting PTEN-regulating molecules, such as certain miRNAs and E3 ubiquitin protein ligase Nedd4, may also become alternatives for attenuating PTEN activity and promoting CNS regeneration. Because adult CNS appears to lack adequate guidance cues present during neurodevelopment, regulation of growing axons to make direct or indirect connections with their original targets appears also important for promoting functional recovery after axonal injuries (Luo et al., 2013). Moreover, task-specific rehabilitative training is probably required for rewiring appropriate neuronal circuits and reinforcing functionally meaningful synaptic reconnections.
Footnotes
Funding: This work was supported by research grants to SL from NIH (1R21NS066114, 1R01NS079432 and 1R01EY024575), Christopher & Dana Reeve Foundation (LA1-1002-2) and Shriners Research Foundation (86300).
Conflicts of interest: None declared.
References
- Alabed YZ, Pool M, Ong Tone S, Sutherland C, Fournier AE. GSK3 beta regulates myelin-dependent axon outgrowth inhibition through CRMP4. J Neurosci. 2010;30:5635–5643. doi: 10.1523/JNEUROSCI.6154-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barbeau H, Rossignol S. Initiation and modulation of the locomotor pattern in the adult chronic spinal cat by noradrenergic, serotonergic and dopaminergic drugs. Brain Res. 1991;546:250–260. doi: 10.1016/0006-8993(91)91489-n. [DOI] [PubMed] [Google Scholar]
- Busch SA, Silver J. The role of extracellular matrix in CNS regeneration. Curr Opin Neurobiol. 2007;17:120–127. doi: 10.1016/j.conb.2006.09.004. [DOI] [PubMed] [Google Scholar]
- Byrne AB, Walradt T, Gardner KE, Hubbert A, Reinke V, Hammarlund M. Insulin/IGF1 signaling inhibits age-dependent axon regeneration. Neuron. 2014;81:561–573. doi: 10.1016/j.neuron.2013.11.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chadborn NH, Ahmed AI, Holt MR, Prinjha R, Dunn GA, Jones GE, Eickholt BJ. PTEN couples Sema3A signalling to growth cone collapse. J Cell Sci. 2006;119:951–957. doi: 10.1242/jcs.02801. [DOI] [PubMed] [Google Scholar]
- Christie KJ, Webber CA, Martinez JA, Singh B, Zochodne DW. PTEN inhibition to facilitate intrinsic regenerative outgrowth of adult peripheral axons. J Neurosci. 2010;30:9306–9315. doi: 10.1523/JNEUROSCI.6271-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cotter L, Ozcelik M, Jacob C, Pereira JA, Locher V, Baumann R, Relvas JB, Suter U, Tricaud N. Dlg1-PTEN interaction regulates myelin thickness to prevent damaging peripheral nerve overmyelination. Science. 2010;328:1415–1418. doi: 10.1126/science.1187735. [DOI] [PubMed] [Google Scholar]
- Courtine G, Song B, Roy RR, Zhong H, Herrmann JE, Ao Y, Qi J, Edgerton VR, Sofroniew MV. Recovery of supraspinal control of stepping via indirect propriospinal relay connections after spinal cord injury. Nat Med. 2008;14:69–74. doi: 10.1038/nm1682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cross DA, Alessi DR, Cohen P, Andjelkovich M, Hemmings BA. Inhibition of glycogen synthase kinase-3 by insulin mediated by protein kinase B. Nature. 1995;378:785–789. doi: 10.1038/378785a0. [DOI] [PubMed] [Google Scholar]
- de Lima S, Koriyama Y, Kurimoto T, Oliveira JT, Yin Y, Li Y, Gilbert HY, Fagiolini M, Martinez AM, Benowitz L. Full-length axon regeneration in the adult mouse optic nerve and partial recovery of simple visual behaviors. Proc Natl Acad Sci U S A. 2012;109:9149–9154. doi: 10.1073/pnas.1119449109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dill J, Wang H, Zhou FQ, Li S. Inactivation of glycogen synthase kinase-3 promotes axonal growth and recovery in the CNS. J Neurosci. 2008;28:8914–8928. doi: 10.1523/JNEUROSCI.1178-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drake PG, Posner BI. Insulin receptor-associated protein tyrosine phosphatase(s): role in insulin action. Mol Cell Biochem. 1998;182:79–89. [PubMed] [Google Scholar]
- Drinjakovic J, Jung H, Campbell DS, Strochlic L, Dwivedy A, Holt CE. E3 ligase Nedd4 promotes axon branching by downregulating PTEN. Neuron. 2010;65:341–357. doi: 10.1016/j.neuron.2010.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher D, Xing B, Dill J, Li H, Hoang HH, Zhao Z, Yang XL, Bachoo R, Cannon S, Longo FM, Sheng M, Silver J, Li S. Leukocyte common antigen-related phosphatase is a functional receptor for chondroitin sulfate proteoglycan axon growth inhibitors. J Neurosci. 2011;31:14051–14066. doi: 10.1523/JNEUROSCI.1737-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gobrecht P, Leibinger M, Andreadaki A, Fischer D. Sustained GSK3 activity markedly facilitates nerve regeneration. Nat Commun. 2014;5:4561. doi: 10.1038/ncomms5561. [DOI] [PubMed] [Google Scholar]
- Goebbels S, Oltrogge JH, Kemper R, Heilmann I, Bormuth I, Wolfer S, Wichert SP, Mobius W, Liu X, Lappe-Siefke C, Rossner MJ, Groszer M, Suter U, Frahm J, Boretius S, Nave KA. Elevated phosphatidylinositol 3,4,5-trisphosphate in glia triggers cell-autonomous membrane wrapping and myelination. J Neurosci. 2010;30:8953–8964. doi: 10.1523/JNEUROSCI.0219-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg JL, Klassen MP, Hua Y, Barres BA. Amacrine-signaled loss of intrinsic axon growth ability by retinal ganglion cells. Science. 2002;296:1860–1864. doi: 10.1126/science.1068428. [DOI] [PubMed] [Google Scholar]
- GrandPre T, Li S, Strittmatter SM. Nogo-66 receptor antagonist peptide promotes axonal regeneration. Nature. 2002;417:547–551. doi: 10.1038/417547a. [DOI] [PubMed] [Google Scholar]
- Grider MH, Park D, Spencer DM, Shine HD. Lipid raft-targeted Akt promotes axonal branching and growth cone expansion via mTOR and Rac1 respectively. J Neurosci Res. 2009;87:3033–3042. doi: 10.1002/jnr.22140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hur EM, Zhou FQ. GSK3 signalling in neural development. Nat Rev Neurosci. 2010;11:539–551. doi: 10.1038/nrn2870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hur EM, Saijilafu, Lee BD, Kim SJ, Xu WL, Zhou FQ. GSK3 controls axon growth via CLASP-mediated regulation of growth cone microtubules. Genes Dev. 2011;25:1968–1981. doi: 10.1101/gad.17015911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inoue K, Rispoli J, Yang L, Macleod D, Beal MF, Klann E, Abeliovich A. Coordinate regulation of mature dopaminergic axon morphology by macroautophagy and the PTEN signaling pathway. PLoS Genet. 2013;9:e1003845. doi: 10.1371/journal.pgen.1003845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang H, Guo W, Liang X, Rao Y. Both the establishment and the maintenance of neuronal polarity require active mechanisms: critical roles of GSK-3beta and its upstream regulators. Cell. 2005;120:123–135. doi: 10.1016/j.cell.2004.12.033. [DOI] [PubMed] [Google Scholar]
- Kim SR, Chen X, Oo TF, Kareva T, Yarygina O, Wang C, During M, Kholodilov N, Burke RE. Dopaminergic pathway reconstruction by Akt/Rheb-induced axon regeneration. Ann Neurol. 2011;70:110–120. doi: 10.1002/ana.22383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim WY, Zhou FQ, Zhou J, Yokota Y, Wang YM, Yoshimura T, Kaibuchi K, Woodgett JR, Anton ES, Snider WD. Essential roles for GSK-3s and GSK-3-primed substrates in neurotrophin-induced and hippocampal axon growth. Neuron. 2006;52:981–996. doi: 10.1016/j.neuron.2006.10.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurimoto T, Yin Y, Omura K, Gilbert HY, Kim D, Cen LP, Moko L, Kugler S, Benowitz LI. Long-distance axon regeneration in the mature optic nerve: contributions of oncomodulin, cAMP, and pten gene deletion. J Neurosci. 2010;30:15654–15663. doi: 10.1523/JNEUROSCI.4340-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewandowski G, Steward O. AAVshRNA-mediated suppression of PTEN in adult rats in combination with salmon fibrin administration enables regenerative growth of corticospinal axons and enhances recovery of voluntary motor function after cervical spinal cord injury. J Neurosci. 2014;34:9951–9962. doi: 10.1523/JNEUROSCI.1996-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S, Strittmatter SM. Delayed systemic Nogo-66 receptor antagonist promotes recovery from spinal cord injury. J Neurosci. 2003;23:4219–4227. doi: 10.1523/JNEUROSCI.23-10-04219.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S, Liu BP, Budel S, Li M, Ji B, Walus L, Li W, Jirik A, Rabacchi S, Choi E, Worley D, Sah DW, Pepinsky B, Lee D, Relton J, Strittmatter SM. Blockade of Nogo-66, myelin-associated glycoprotein, and oligodendrocyte myelin glycoprotein by soluble Nogo-66 receptor promotes axonal sprouting and recovery after spinal injury. J Neurosci. 2004;24:10511–10520. doi: 10.1523/JNEUROSCI.2828-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu BP, Cafferty WB, Budel SO, Strittmatter SM. Extracellular regulators of axonal growth in the adult central nervous system. Philos Trans R Soc Lond B Biol Sci. 2006;361:1593–1610. doi: 10.1098/rstb.2006.1891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu K, Lu Y, Lee JK, Samara R, Willenberg R, Sears-Kraxberger I, Tedeschi A, Park KK, Jin D, Cai B, Xu B, Connolly L, Steward O, Zheng B, He Z. PTEN deletion enhances the regenerative ability of adult corticospinal neurons. Nat Neurosci. 2010;13:1075–1081. doi: 10.1038/nn.2603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu Y, Belin S, He Z. Signaling regulations of neuronal regenerative ability. Curr Opin Neurobiol. 2014;27:135–142. doi: 10.1016/j.conb.2014.03.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo X, Salgueiro Y, Beckerman SR, Lemmon VP, Tsoulfas P, Park KK. Three-dimensional evaluation of retinal ganglion cell axon regeneration and pathfinding in whole mouse tissue after injury. Exp Neurol. 2013;247:653–662. doi: 10.1016/j.expneurol.2013.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mao L, Jia J, Zhou X, Xiao Y, Wang Y, Mao X, Zhen X, Guan Y, Alkayed NJ, Cheng J. Delayed administration of a PTEN inhibitor BPV improves functional recovery after experimental stroke. Neuroscience. 2013;231:272–281. doi: 10.1016/j.neuroscience.2012.11.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGee AW, Strittmatter SM. The Nogo-66 receptor: focusing myelin inhibition of axon regeneration. Trends Neurosci. 2003;26:193–198. doi: 10.1016/S0166-2236(03)00062-6. [DOI] [PubMed] [Google Scholar]
- Moore DL, Goldberg JL. Multiple transcription factor families regulate axon growth and regeneration. Dev Neurobiol. 2011;71:1186–1211. doi: 10.1002/dneu.20934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ning K, Drepper C, Valori CF, Ahsan M, Wyles M, Higginbottom A, Herrmann T, Shaw P, Azzouz M, Sendtner M. PTEN depletion rescues axonal growth defect and improves survival in SMN-deficient motor neurons. Hum Mol Genet. 2010;19:3159–3168. doi: 10.1093/hmg/ddq226. [DOI] [PubMed] [Google Scholar]
- O’Donovan KJ, Ma K, Guo H, Wang C, Sun F, Han SB, Kim H, Wong JK, Charron J, Zou H, Son YJ, He Z, Zhong J. B-RAF kinase drives developmental axon growth and promotes axon regeneration in the injured mature CNS. J Exp Med. 2014;211:801–814. doi: 10.1084/jem.20131780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohtake Y, Park D, Abdul-Muneer PM, Li H, Xu B, Sharma K, Smith GM, Selzer ME, Li S. The effect of systemic PTEN antagonist peptides on axon growth and functional recovery after spinal cord injury. Biomaterials. 2014;35:4610–4626. doi: 10.1016/j.biomaterials.2014.02.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park KK, Liu K, Hu Y, Kanter JL, He Z. PTEN/mTOR and axon regeneration. Exp Neurol. 2010;223:45–50. doi: 10.1016/j.expneurol.2009.12.032. [DOI] [PubMed] [Google Scholar]
- Park KK, Liu K, Hu Y, Smith PD, Wang C, Cai B, Xu B, Connolly L, Kramvis I, Sahin M, He Z. Promoting axon regeneration in the adult CNS by modulation of the PTEN/mTOR pathway. Science. 2008;322:963–966. doi: 10.1126/science.1161566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Proud CG. Regulation of mammalian translation factors by nutrients. Eur J Biochem. 2002;269:5338–5349. doi: 10.1046/j.1432-1033.2002.03292.x. [DOI] [PubMed] [Google Scholar]
- Ribotta MG, Provencher J, Feraboli-Lohnherr D, Rossignol S, Privat A, Orsal D. Activation of locomotion in adult chronic spinal rats is achieved by transplantation of embryonic raphe cells reinnervating a precise lumbar level. J Neurosci. 2000;20:5144–5152. doi: 10.1523/JNEUROSCI.20-13-05144.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scrivens PJ, Alaoui-Jamali MA, Giannini G, Wang T, Loignon M, Batist G, Sandor VA. Cdc25A-inhibitory properties and antineoplastic activity of bisperoxovanadium analogues. Mol Cancer Ther. 2003;2:1053–1059. [PubMed] [Google Scholar]
- Singh B, Singh V, Krishnan A, Koshy K, Martinez JA, Cheng C, Almquist C, Zochodne DW. Regeneration of diabetic axons is enhanced by selective knockdown of the PTEN gene. Brain. 2014;137:1051–1067. doi: 10.1093/brain/awu031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song Y, Ori-McKenney KM, Zheng Y, Han C, Jan LY, Jan YN. Regeneration of Drosophila sensory neuron axons and dendrites is regulated by the Akt pathway involving Pten and microRNA bantam. Genes Dev. 2012;26:1612–1625. doi: 10.1101/gad.193243.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun F, Park KK, Belin S, Wang D, Lu T, Chen G, Zhang K, Yeung C, Feng G, Yankner BA, He Z. Sustained axon regeneration induced by co-deletion of PTEN and SOCS3. Nature. 2011;480:372–375. doi: 10.1038/nature10594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker CL, Xu XM. PTEN inhibitor bisperoxovanadium protects oligodendrocytes and myelin and prevents neuronal atrophy in adult rats following cervical hemicontusive spinal cord injury. Neurosci Lett. 2014;573:64–68. doi: 10.1016/j.neulet.2014.02.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker CL, Liu NK, Xu XM. PTEN/PI3K and MAPK signaling in protection and pathology following CNS injuries. Front Biol (Beijing) 2013:8. doi: 10.1007/s11515-013-1255-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker CL, Walker MJ, Liu NK, Risberg EC, Gao X, Chen J, Xu XM. Systemic bisperoxovanadium activates Akt/mTOR, reduces autophagy, and enhances recovery following cervical spinal cord injury. PLoS One. 2012;7:e30012. doi: 10.1371/journal.pone.0030012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshimura T, Kawano Y, Arimura N, Kawabata S, Kikuchi A, Kaibuchi K. GSK-3beta regulates phosphorylation of CRMP-2 and neuronal polarity. Cell. 2005;120:137–149. doi: 10.1016/j.cell.2004.11.012. [DOI] [PubMed] [Google Scholar]
- Zhang Y, Granholm AC, Huh K, Shan L, Diaz-Ruiz O, Malik N, Olson L, Hoffer BJ, Lupica CR, Hoffman AF, Backman CM. PTEN deletion enhances survival, neurite outgrowth and function of dopamine neuron grafts to MitoPark mice. Brain. 2012;135:2736–2749. doi: 10.1093/brain/aws196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao J, Qu Y, Wu J, Cao M, Ferriero DM, Zhang L, Mu D. PTEN inhibition prevents rat cortical neuron injury after hypoxia-ischemia. Neuroscience. 2013;238:242–251. doi: 10.1016/j.neuroscience.2013.02.046. [DOI] [PubMed] [Google Scholar]
- Zhou FQ, Snider WD. Cell biology. GSK-3beta and microtubule assembly in axons. Science. 2005;308:211–214. doi: 10.1126/science.1110301. [DOI] [PubMed] [Google Scholar]
- Zhou FQ, Snider WD. Intracellular control of developmental and regenerative axon growth. Philos Trans R Soc Lond B Biol Sci. 2006;361:1575–1592. doi: 10.1098/rstb.2006.1882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou FQ, Zhou J, Dedhar S, Wu YH, Snider WD. NGF-induced axon growth is mediated by localized inactivation of GSK-3beta and functions of the microtubule plus end binding protein APC. Neuron. 2004;42:897–912. doi: 10.1016/j.neuron.2004.05.011. [DOI] [PubMed] [Google Scholar]
- Zou H, Ho C, Wong K, Tessier-Lavigne M. Axotomy-induced Smad1 activation promotes axonal growth in adult sensory neurons. J Neurosci. 2009;29:7116–7123. doi: 10.1523/JNEUROSCI.5397-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zukor K, Belin S, Wang C, Keelan N, Wang X, He Z. Short hairpin RNA against PTEN enhances regenerative growth of corticospinal tract axons after spinal cord injury. J Neurosci. 2013;33:15350–15361. doi: 10.1523/JNEUROSCI.2510-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]