

Abstract
Hypoxic preconditioning activates endogenous mechanisms that protect against cerebral ischemic and hypoxic injury. To better understand these protective mechanisms, adult rats were housed in a hypoxic environment (8% O2/92% N2) for 3 hours, and then in a normal oxygen environment for 12 hours. Their cerebrospinal fluid was obtained to culture cortical neurons from newborn rats for 1 day, and then the neurons were exposed to oxygen-glucose deprivation for 1.5 hours. The cerebrospinal fluid from rats subjected to hypoxic preconditioning reduced oxygen-glucose deprivation-induced injury, increased survival rate, upregulated Bcl-2 expression and downregulated Bax expression in the cultured cortical neurons, compared with control. These results indicate that cerebrospinal fluid from rats given hypoxic preconditioning protects against oxygen-glucose deprivation-induced injury by affecting apoptosis-related protein expression in neurons from newborn rats.
Keywords: nerve regeneration, hypoxic preconditioning, cerebrospinal fluid, cerebral cortex, oxygen-glucose deprivation, neurons, apoptosis, Bcl-2/Bax, neural regeneration
Introduction
Ischemic cerebrovascular disease-induced vascular dementia substantially affects the health and quality of life of patients. Therefore, it is necessary to develop neuroprotective strategies that relieve cortical neuronal injury after ischemic cerebrovascular events and improve the prognosis of vascular dementia (Meguro et al., 2013; Kwon et al., 2014; Liu et al., 2014; Sakr et al., 2014; Sun et al., 2014). Hypoxic preconditioning (HPC) activates a complex series of endogenous protective mechanisms that reduce hypoxic injury (Benitez et al., 2014; Monson et al., 2014; Sheldon et al., 2014; Suryana and Jones, 2014; Liu et al., 2015). HPC has protective effects against ischemic/hypoxic brain injury and ischemic cerebrovascular disease (Gidday, 2006; Benitez et al., 2014; Monson et al., 2014; Sheldon et al., 2014; Suryana and Jones, 2014; Liu et al., 2015). HPC has been demonstrated to protect against ischemic brain injury induced by acute cerebral infarction as evaluated by measuring infarct volume, neurological function assessment and apoptosis assay (Niu et al., 2009). However, it is difficult to implement HPC in the clinical setting. Nonetheless, HPC is currently a hot topic in translational medicine research.
Cerebrospinal fluid (CSF) is primarily secreted by the choroid plexus of the lateral ventricles, and contains a variety of electrolytes, proteins, glucose and growth factors (Cui et al., 2001; Lehtinen et al., 2011; Niu et al., 2011). CSF has been shown to induce the differentiation of neural stem cells into neurons and astrocytes (Lehtinen et al., 2011; Niu et al., 2011). CSF has also been shown to be neuroprotective for neurons and PC12 cells (Niu et al., 2011). In the present study, we investigate the potential neuroprotective effects of CSF obtained from rats given HPC on cortical neurons exposed to hypoxia, and we examine the underlying mechanisms of action.
Materials and Methods
Experimental animals
A total of 345 healthy adult Wistar rats weighing 200–250 g, irrespective of gender, and 48 neonatal Wistar rats (born within 12 hours) were included in this study. All rats were provided by the Lukang Experimental Animal Center in Jining City, Shandong Province, China (animal lot No. SCXK20080002). Procedures for the use of laboratory animals were approved by the Institutional Animal Care and Use Committee of Affiliated Hospital of Taishan Medical University, China.
Culture and identification of neurons
The cortices were removed from neonatal Wistar rats under sterile conditions, sliced into 1-mm3 blocks in cold D-Hanks buffer, digested with 0.125% trypsin at 37°C in a 5% CO2 incubator for 10 minutes, and filtered with a 200-mesh sieve. Cells at 5 × 105/mL were incubated in a 0.01% poly-L-lysine (Sigma, St. Louis, MO, USA)-coated plate. Then, 24 hours later, the medium was completely replaced with fresh medium. From then on, half of the medium was replaced every 3 days. At 8 days, primary cortical neurons were identified as follows: cells were washed twice with 0.1 M PBS, fixed with 4% paraformaldehyde for 30 minutes, washed twice with PBS, and air-dried. Neurons were stained according to instructions provided by the Neuron Specific Enolase kit (Sigma), and observed under the fluorescence microscope (Radiance 2100; Bio-Rad, Hercules, CA, USA) (Yang et al., 2009; Niu et al., 2011).
Preparation of rat CSF
In accordance with a previous method (Vannucci et al., 1998), adult rats were placed in a low-oxygen chamber (self-made, 8% O2, 92% N2, 37°C) for 3 hours for HPC. Then, after housing under normal oxygen for 12 hours, rats were intraperitoneally anesthetized with 10% chloral hydrate (3–4 mg/kg), and surgically operated. The atlantooccipital membrane was punctured, and the cerebrospinal fluid was extracted from the cisterna magna (approximately 200 μL/rat), filtered using a 0.22-μm microporous membrane, sterilized, and stored at −80°C for subsequent use.
Cell culture
Primary neurons, cultured for 8 days, were randomly assigned to normal control, oxygen-glucose deprivation (OGD), normal CSF and HPC CSF groups. In the normal control group, neurons were cultured with normal high-glucose medium (Life Technologies, San Francisco, CA, USA). In the OGD group, neurons were exposed to OGD for 1.5 hours. In the normal CSF group, neurons were cultured with normal rat CSF and high-glucose Dulbecco's modified Eagle's medium (DMEM) at 1:9 for 1 day, and then exposed to OGD for 1.5 hours. In the HPC CSF group, neurons were cultured with 10% CSF from rats subjected to HPC for 1 day, and then exposed to OGD for 1.5 hours. Each group consisted of six wells. OGD culture was as follows: neurons were washed three times with glucose-free Earle's solution, and then incubated with glucose-free Earle's solution containing 1 mM Na2S2O4 at 37°C, 5% CO2 and saturated humidity (Niu et al., 2011).
Assessment of apoptosis
After removal of the medium, neurons on coverslips were washed twice with PBS, and then incubated with 5 μL annexin-V-FITC and 5 μL propidium iodide (KeyGEN Biotech, Nanjing, China) in 500 μL binding buffer in the dark at room temperature for 5 minutes. After mounting, neurons were observed by confocal laser scanning microscopy (Bio-Rad). In the early stage of apoptosis, the cell membrane displayed green fluorescence, and the nuclei were not stained. In the middle and late stages of apoptosis, the cell membrane displayed green fluorescence, and the nuclei were stained red. Nuclei of dead disintegrated cells were stained red. Fluorescence images were captured using LaserSharp 2000 software (4.5.3; Bio-Rad). A total of 20 images were randomly obtained from each group for measurement of fluorescence intensity (Niu et al., 2011).
Flow cytometry
Neurons were digested with 0.125% trypsin for approximately 4 minutes. Cells were collected and placed in centrifuge tubes and made into a single-cell suspension. Thereafter, 1 mL of cells at 1 × 106/mL was centrifuged at 1,000 r/min for 10 minutes. The supernatant was discarded, and the cells were resuspended in 1 mL of cold PBS. Cells were resuspended in 200 μL binding buffer, and 10 μL annexin V-FITC and 5 μL propidium iodide were added in the dark at room temperature for 20 minutes. For the control, 400 μL of PBS was added instead of dye. Samples were filtered through a 200-mesh sieve and detected immediately with a flow cytometer (FACSCalibur; Bio-Rad) (Yang et al., 2009).
Immunofluorescence assay
After removal of the medium, neurons were washed twice with 0.01 M PBS (pH 7.4) at 37°C, fixed with 4% paraformaldehyde for 30 minutes, washed twice with PBS, dried, and then blocked with normal serum (Life Technologies) (diluted with 0.01 M PBS at 1:10) at room temperature for 20 minutes. Excess liquid was shaken off. Neurons were incubated with rabbit anti-Bcl-2 or Bax polyclonal antibody (1:100; Santa Cruz Biotechnology, Santa Cruz, CA, USA) at 4°C overnight. PBS was used as negative control. After washing with PBS, neurons were incubated with goat anti-rabbit IgG-FITC (1:100; Boster, Wuhan, China) at 37°C for 30 minutes. Neurons were then washed, mounted with glycerol in PBS (1:1), and finally observed by confocal laser scanning microscopy (Bio-Rad). Scanning parameters were as follows: excitation wavelength, 554 nm; observation wavelength, 575 nm; 40× objective; point scanning; zoom (1.0). Scanned images were photographed and analyzed using LaserSharp 2000 software (v. 4.5.3; Bio-Rad) (Zhang et al., 2007a, 2008).
Statistical analysis
The data are expressed as the mean ± SD, and were analyzed with one-way analysis of variance and Dunnett's two-tailed t-test using SigmaStat 3.5 (Systat Software, Inc., San Jose, CA, USA). A value of P < 0.05 was considered statistically significant.
Results
CSF from rats given HPC reduced apoptosis in cortical neurond exposed to OGD
In the normal control group, weak red (propidium iodide) and green (annexin V) fluorescence, a few stained cells, and a few early apoptotic neurons were visible. In the OGD group, there was abundant dark green and red fluorescence, and many bare red nuclei, middle and late apoptotic neurons and dead neurons were visible. In the normal CSF group, many cells had a green membrane and a red nucleus. Most were middle and late apoptotic neurons, although a few were early apoptotic neurons. Green and red fluorescence intensity was weaker in the normal CSF group than in the OGD group (P < 0.01). In the HPC CSF group, there were a few fluorescent cells and many cells with green membranes, characteristic of cells undergoing early apoptosis. Green and red fluorescence intensity was significantly weaker in the HPC CSF group than in the OGD and normal CSF groups (P < 0.01; Figure 1, Table 1).
Figure 1.
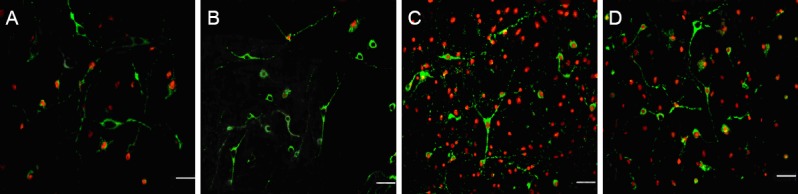
Effect of HPC CSF on apoptosis in cultured cortical neurons exposed to OGD (confocal laser scanning microscopy).
(A) In the normal control group, a few early apoptotic neurons are visible. (B) In the OGD group, middle and late apoptotic neurons and dead neurons are detectable. Green and red fluorescence intensities were weaker in the normal CSF group (C) than in the OGD group. Green and red fluorescence intensities were weaker in the HPC CSF group (D) than in the OGD and normal CSF groups. Green: Annexin V; red: propidium iodide. Scale bars: 50 μm. OGD: Oxygen-glucose deprivation; CSF: cerebrospinal fluid; HPC CSF: CSF from rats given hypoxic preconditioning.
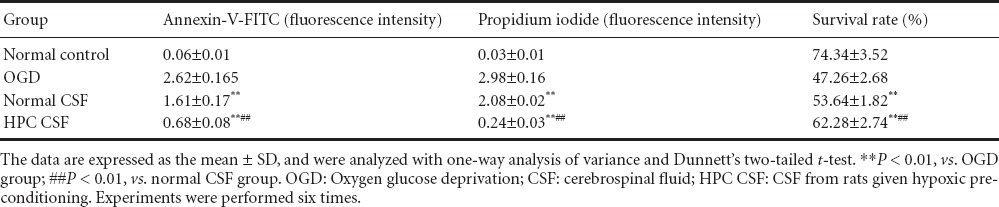
Table 1.
Effects of HPC CSF on cultured cortical neuron apoptosis and survival
CSF from rats given HPC increased the survival rate of cortical neurons exposed to OGD
Flow cytometry revealed that the survival rate was highest in the normal control group, and lowest in the OGD group. Compared with the OGD group, cell survival rate was significantly higher in the normal CSF group (P < 0.01). Furthermore, the survival rate was significantly greater in the HPC CSF group than in the OGD and normal CSF groups (P < 0.01; Figure 2, Table 1).
Figure 2.
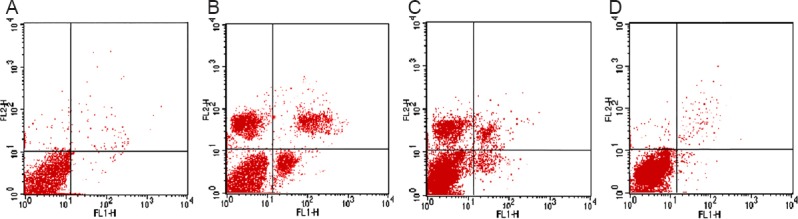
Effects of HPC CSF on cultured cortical neuron survival rate (flow cytometry).
Cell survival rate was highest in the normal control group (A), and lowest in the OGD group (B). Compared with the OGD group, cell survival rate was higher in the normal CSF group (C). The survival rate was greater in the HPC CSF group (D) than in the OGD and normal CSF groups. Lower left: Normal cells; lower right: early apoptotic cells; upper left: necrotic cells: upper right: late apoptotic cells. OGD: Oxygen glucose deprivation; CSF: cerebrospinal fluid; HPC CSF: CSF from rats given hypoxic preconditioning.
CSF from rats given HPC increased Bcl-2 expression and decreased Bax expression in cortical neurons exposed to OGD
The number of Bcl-2-positive cells was low in the normal control and OGD groups. The intensity of Bcl-2 immunofluorescence was significantly greater in the normal CSF group than in the normal control and OGD groups (P < 0.05). No significant difference in the number of Bcl-2-positive cells was detectable between the normal CSF and OGD groups. Bcl-2 immunofluorescence intensity and the number of Bcl-2-positive cells were significantly higher in the HPC CSF group than in the OGD and normal CSF groups (P < 0.01, P < 0.05; Figure 3, Table 2).
Figure 3.
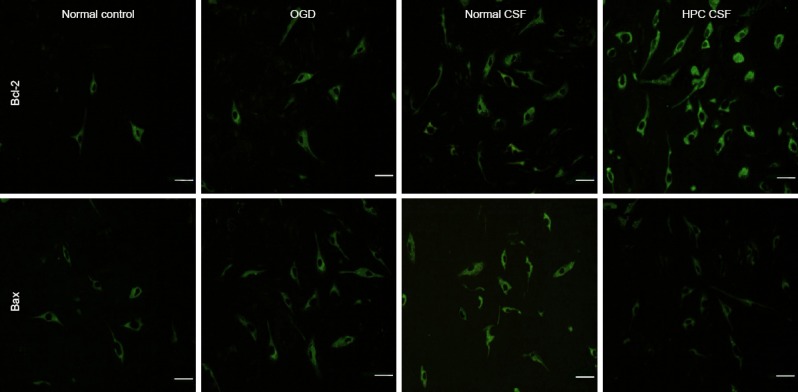
Effects of HPC CSF on Bcl-2 and Bax expression in cultured cortical neurons exposed to OGD (immunofluorescence assay, confocal laser scanning microscopy).
The number of Bcl-2- and Bax-positive cells was less in the normal control and OGD groups. Fluorescence intensities of Bcl-2 and Bax immunolabeling and the numbers of Bcl-2 and Bax-positive cells were greater in the HPC CSF group than in the OGD and normal CSF groups. Scale bars: 50 μm. OGD: Oxygen glucose deprivation; CSF: cerebrospinal fluid; HPC CSF: CSF from rats given hypoxic preconditioning.
Table 2.
Effect of HPC CSF on Bcl-2 and Bax expression in cultured cortical neurons
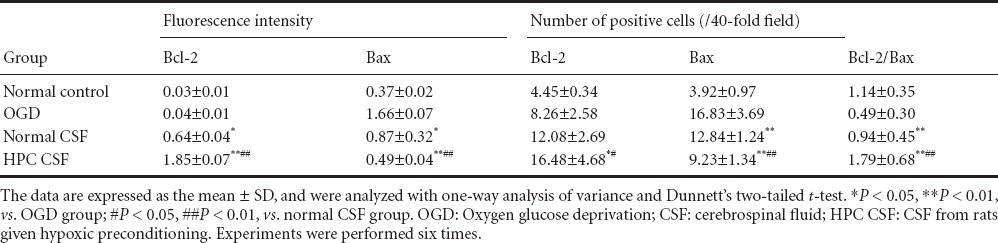
For Bax, in the normal control group, only minor stellate fluorescence was visible, the fluorescence intensity was low, and only a few Bax-positive cells were detectable. The fluorescence intensity of Bax immunolabeling and the number of Bax-positive cells were significantly lower in the HPC CSF and normal CSF groups than in the OGD group (P < 0.05 or P < 0.01). Bax immunofluorescence intensity and the number of Bax-positive cells were significantly lower in the HPC CSF group than in the normal CSF group (P < 0.01; Figure 3, Table 2).
The Bcl-2/Bax ratio was significantly higher in the normal CSF and HPC CSF groups than in the OGD group (P < 0.01). The Bcl-2/Bax ratio was significantly greater in the HPC CSF group than in the normal CSF group (P < 0.01; Table 2).
Discussion
Following HPC (i.e., exposure to a short period of nonfatal hypoxia), cells acquire tolerance to subsequent longer-term ischemic/hypoxic injury (Vannucci et al., 1998; Cui et al., 2001; Lu et al., 2004; Lu et al., 2005; Gidday, 2006; Zhang et al., 2007a, b, 2008); however, the underlying mechanisms are complex and unclear (Vannucci et al., 1998; Cui et al., 2001; Lu et al., 2004; Lu et al., 2005; Gidday, 2006; Zhang et al., 2007a, b, 2008). HPC induces numerous changes, including changes in behavior and metabolism, cellular morphology, as well as neurochemical and molecular biological alterations (Vannucci et al., 1998; Cui et al., 2001; Lu et al., 2004; Lu et al., 2005; Gidday, 2006; Zhang et al., 2007a, b, 2008).
At present, HPC cannot be directly applied in the clinical setting. However, cerebrospinal fluid components can easily traverse the blood-brain barrier. A better understanding of the effects of CSF from animals given HPC may help identify factors that protect against hypoxic/ischemic injury.
In the present study, we found high levels of neuronal apoptosis after 1.5 hours of OGD. Consistent results were obtained using both flow cytometry and confocal laser scanning microscopy. Both normal CSF and CSF from animals given HPC had protective effects on neurons subjected to OGD. The number of apoptotic neurons was reduced in the normal CSF and HPC CSF groups compared with the OGD group. HPC CSF displayed a better neuroprotective effect than normal CSF.
We also sought to clarify the mechanisms mediating the neuroprotective effects of HPC CSF. We examined whether HPC CSF inhibits apoptosis, thereby enhancing cell survival after OGD. Bcl-2 and Bax are critical apoptotic proteins. Bcl-2 suppresses the opening of the mitochondrial permeability transition pore and the start of the apoptotic cascade by regulating caspase-activating factors, reducing intracellular oxidative stress, inhibiting intracellular calcium-activated DNA cleavage, blocking pro-apoptotic pathways, and by maintaining mitochondrial structure and function. In contrast, Bax, which is downstream of p53, is activated by apoptotic stimuli and binds to Bcl-2 on the mitochondrial membrane, forming a heterodimer. Bax counters the anti-apoptotic effects of Bcl-2, and Bax homodimers can directly initiate the apoptotic cascade (Ramalingam and Kim, 2014; Sharifi et al., 2014; Wang et al., 2014a, b; Wu et al., 2014; Yang et al., 2014; Zhao and He, 2015). The Bcl-2/Bax protein ratio determines the ultimate fate of cells. Apoptosis occurs when Bax is dominant, and cell survival is observed when Bcl-2 is dominant (Ramalingam and Kim, 2014; Sharifi et al., 2014; Wang et al., 2014a; Wang et al., 2014b; Wu et al., 2014; Yang et al., 2014; Zhao and He, 2015). We observed that Bcl-2 expression and the Bcl-2/Bax ratio were significantly higher in the HPC CSF group than in the OGD and normal CSF groups. Bax expression was significantly lower in the HPC CSF group than in the OGD and normal CSF groups. Therefore, HPC CSF maintains mitochondrial structure and function, improves hypoxic tolerance, and inhibits apoptosis by upregulating Bcl-2 expression and downregulating Bax expression. HPC CSF may mediate neuroprotection via other mechanisms as well; however, further study is required to identify the neuroprotective factors and their mechanisms of action.
In this study, normal rat CSF was also found to have neuroprotective effects, possibly because it contains factors that enhance neuronal survival, such as nerve growth factor (Cui et al., 2001; Lehtinen et al., 2011; Niu et al., 2011). CSF likely contains numerous neuroprotective factors that easily traverse the blood-brain barrier. CSF also has minimal adverse effects, and protects against ischemic injury. CSF can be used to produce novel drugs that may have therapeutic potential for ischemic brain injury.
In summary, CSF from rats given HPC reduced injury to OGD-exposed cortical neurons and exhibited neuroprotective effects. The neuroprotective mechanism appears to involve the upregulation of Bcl-2 expression, the downregulation of Bax expression, and an increase in the Bcl-2/Bax ratio.
Footnotes
Funding: This study was supported by a grant from the National Science and Technology Support Program of China, No. 2013BAI07B01; and the Natural Science Foundation of Shandong Province in China, No. ZR2012HQ014, ZR2011HM044; a grant from the Open Research Project of Beijing Key Laboratory for Hypoxic Preconditioning and Translational Medicine, No. 2015DYSY02.
Conflicts of interest: None declared.
Copyedited by Patel B, Robens J, Yu J, Qiu Y, Li CH, Song LP, Zhao M
References
- Benitez SG, Castro AE, Patterson SI, Muñoz EM, Seltzer AM. Hypoxic preconditioning differentially affects GABAergic and glutamatergic neuronal cells in the injured cerebellum of the neonatal rat. PLoS One. 2014;9:e102056. doi: 10.1371/journal.pone.0102056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui XY, Li L, An YY, Lu GW. Changes in the contents of glycogen and lactate in the brain and blood during hypoxic preconditioning. Sheng Li Xue Bao. 2001;53:325–328. [PubMed] [Google Scholar]
- Gidday JM. Cerebral preconditioning and ischaemic tolerance. Nat Rev Neurosci. 2006;7:437–448. doi: 10.1038/nrn1927. [DOI] [PubMed] [Google Scholar]
- Kwon KJ, Kim MK, Lee EJ, Kim JN, Choi BR, Kim SY, Cho KS, Han JS, Kim HY, Shin CY, Han SH. Effects of donepezil, an acetylcholinesterase inhibitor, on neurogenesis in a rat model of vascular dementia. J Neurol Sci. 2014;347:66–77. doi: 10.1016/j.jns.2014.09.021. [DOI] [PubMed] [Google Scholar]
- Lehtinen MK, Zappaterra MW, Chen X, Yang YJ, Hill A, Lun M, Maynard T, Gonzalez D, Kim S, Ye P, D’Ercole AJ, Wong ET, LaMantia AS, Walsh CA. The cerebrospinal fluid provides a proliferative niche for neural progenitor cells. Neuron. 2011;69:893–905. doi: 10.1016/j.neuron.2011.01.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu X, Zhang J, Sun D, Fan Y, Zhou H, Fu B. Effects of fluoxetine on brain-derived neurotrophic factor serum concentration and cognition in patients with vascular dementia. Clin Interv Aging. 2014;9:411–418. doi: 10.2147/CIA.S58830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Y, Sun Z, Sun S, Duan Y, Shi J, Qi Z, Meng R, Sun Y, Zeng X, Chui D, Ji X. Effects of hypoxic preconditioning on synaptic ultrastructure in mice. Synapse. 2015;69:7–14. doi: 10.1002/syn.21777. [DOI] [PubMed] [Google Scholar]
- Lu GW, Cui XY, Yu S, Li RH, Shao G. Tolerant limit to hypoxia and hypoxic preconditioning. Zhongguo Shenjng Kexue Zazhi. 2004;20:388–393. [Google Scholar]
- Lu GW, Yu S, Li RH, Cui XY, Gao CY. Hypoxic preconditioning: a novel intrinsic cytoprotective strategy. Mol Neurobiol. 2005;31:255–271. doi: 10.1385/MN:31:1-3:255. [DOI] [PubMed] [Google Scholar]
- Meguro K, Akanuma K, Ouchi Y, Meguro M, Nakamura K, Yamaguchi S. Vascular dementia with left thalamic infarction: neuropsychological and behavioral implications suggested by involvement of the thalamic nucleus and the remote effect on cerebral cortex. The Osaki-Tajiri project. Psychiatry Res. 2013;213:56–62. doi: 10.1016/j.pscychresns.2012.12.004. [DOI] [PubMed] [Google Scholar]
- Monson NL, Ortega SB, Ireland SJ, Meeuwissen AJ, Chen D, Plautz EJ, Shubel E, Kong X, Li MK, Freriks LH, Stowe AM. Repetitive hypoxic preconditioning induces an immunosuppressed B cell phenotype during endogenous protection from stroke. J Neuroinflammation. 2014;11:22. doi: 10.1186/1742-2094-11-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niu JZ, Zhang YB, Li MY, Liu LL. Lessening effect of hypoxia-preconditioned rat cerebrospinal fluid on oxygen-glucose deprivation-induced injury of cultured hippocampal neurons in neonate rats and possible mechanism. Sheng Li Xue Bao. 2011;63:491–497. [PubMed] [Google Scholar]
- Niu JZ, Zhang YB, Yang MF, Liu LL, Sun BL. Protective effect of hypoxic preconditioning against cerebral ischemic injury induced by acute cerebral infarction in mice. Zhonghua Shenjing Yixue Zazhi. 2009;8:777–780. [Google Scholar]
- Ramalingam M, Kim SJ. Insulin involved Akt/ERK and Bcl-2/Bax pathways against oxidative damages in C6 glial cells. J Recept Signal Transduct Res. 2014:1–7. doi: 10.3109/10799893.2014.970276. [DOI] [PubMed] [Google Scholar]
- Sakr HF, Khalil KI, Hussein AM, Zaki MS, Eid RA, Alkhateeb M. Effect of dehydroepiandrosterone (DHEA) on memory and brain derived neurotrophic factor (BDNF) in a rat model of vascular dementia. J Physiol Pharmacol. 2014;65:41–53. [PubMed] [Google Scholar]
- Sharifi S, Barar J, Hejazi MS, Samadi N. Roles of the Bcl-2/Bax ratio, caspase-8 and 9 in resistance of breast cancer cells to paclitaxel. Asian Pac J Cancer Prev. 2014;15:8617–8622. doi: 10.7314/apjcp.2014.15.20.8617. [DOI] [PubMed] [Google Scholar]
- Sheldon RA, Lee CL, Jiang X, Knox RN, Ferriero DM. Hypoxic preconditioning protection is eliminated in HIF-1α knockout mice subjected to neonatal hypoxia-ischemia. Pediatr Res. 2014;76:46–53. doi: 10.1038/pr.2014.53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun ZK, Ma XR, Jia YJ, Liu YR, Zhang JW, Zhang BA. Effects of resveratrol on apoptosis in a rat model of vascular dementia. Exp Ther Med. 2014;7:843–848. doi: 10.3892/etm.2014.1542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suryana E, Jones NM. The effects of hypoxic preconditioning on white matter damage following hypoxic-ischaemic injury in the neonatal rat brain. Int J Dev Neurosci. 2014;37:69–75. doi: 10.1016/j.ijdevneu.2014.06.007. [DOI] [PubMed] [Google Scholar]
- Vannucci RC, Towfighi J, Vannucci SJ. Hypoxic preconditioning and hypoxic-ischemic brain damage in the immature rat: pathologic and metabolic correlates. J Neurochem. 1998;71:1215–1220. doi: 10.1046/j.1471-4159.1998.71031215.x. [DOI] [PubMed] [Google Scholar]
- Wang GH, Lan R, Zhen XD, Zhang W, Xiang J, Cai DF. An-Gong-Niu-Huang Wan protects against cerebral ischemia induced apoptosis in rats: up-regulation of Bcl-2 and down-regulation of Bax and caspase-3. J Ethnopharmacol. 2014a;154:156–162. doi: 10.1016/j.jep.2014.03.057. [DOI] [PubMed] [Google Scholar]
- Wang Y, Zhang H, Chai F, Liu X, Berk M. The effects of escitalopram on myocardial apoptosis and the expression of Bax and Bcl-2 during myocardial ischemia/reperfusion in a model of rats with depression. BMC Psychiatry. 2014b;14:349. doi: 10.1186/s12888-014-0349-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu B, Cui H, Peng X, Fang J, Zuo Z, Deng J, Huang J. Dietary nickel chloride induces oxidative stress, apoptosis and alters Bax/Bcl-2 and caspase-3 mRNA expression in the cecal tonsil of broilers. Food Chem Toxicol. 2014;63:18–29. doi: 10.1016/j.fct.2013.10.033. [DOI] [PubMed] [Google Scholar]
- Yang MF, Zhang YB, Sun BL, Niu JZ, Lu GW. Effect of brain homogenate from hypoxia-preconditioned mice on rat embryonic hippocampal neurons with hypoxia/reoxygenation injury. Zhonghua Shenjing Yixue Zazhi. 2009;8:1094–1097. [Google Scholar]
- Yang SD, Bai ZL, Zhang F, Ma L, Yang DL, Ding WY. Levofloxacin increases the effect of serum deprivation on anoikis of rat nucleus pulposus cells via Bax/Bcl-2/caspase-3 pathway. Toxicol Mech Methods. 2014;24:688–696. doi: 10.3109/15376516.2014.963772. [DOI] [PubMed] [Google Scholar]
- Zhang YB, Lu GW, Yang MF, Niu JZ, Sun BL. Changes in Bcl-2 and Caspase-3 expressions in cortex of hypoxic preconditioning mice. Sheng Li Xue Bao. 2008;60:249–253. [PubMed] [Google Scholar]
- Zhang YB, Lu GW, Yang MF, Wang X, Niu JZ, Sun BL. Changes in the bcl-2 expression and caspase-3 activity in mouse ependyma during hypoxic preconditioning. Zhonghua Shenjing Yixue Zazhi. 2007a;6:986–988. [Google Scholar]
- Zhang YB, Lu GW, Yang MF, Wang X, Sun BL, Niu JZ. Changes in the Bcl-2 expression and Caspase-3 activity in hippocampus of hypoxic preconditioning mice. Zhonghua Shenjing Ke Zazhi. 2007b;40:553–555. [Google Scholar]
- Zhao B, He T. Chidamide, a histone deacetylase inhibitor, functions as a tumor inhibitor by modulating the ratio of Bax/Bcl-2 and P21 in pancreatic cancer. Oncol Rep. 2015;33:304–310. doi: 10.3892/or.2014.3595. [DOI] [PubMed] [Google Scholar]