

Abstract
Most viruses rely heavily on their host machinery to successfully replicate their genome and produce new virus particles. Recently, the interaction of positive-strand RNA viruses with the lipid biosynthetic and transport machinery has been the subject of intense investigation. In this review, we will discuss the contribution of various host lipids and related proteins in RNA virus replication and maturation.
Introduction
Viruses have evolved to utilize cellular membranes for entry, replication of their genome and production of nascent particles. Notably, positive-strand RNA viruses have developed this unique ability to co-opt cytosolic membranes to build a novel RNA replication factory. The factory contains the replicase complex, which includes viral RNA, proteins and host factors [1,2]. Additionally, this platform is thought to protect the virus from host immune defenses. Previous reports indicate that the endoplasmic reticulum (ER), Golgi complex, mitochondria and endosome membranes are reorganized by various RNA viruses to build their replication complex [1-9]. For hepatitis C virus, the replication factory consists of double-membrane vesicles (DMVs) organized in a membranous web (MW) structure [10-15]. Efforts to understand the organization and function of RNA virus replication platforms have initially focused on virus and host factors. However, there is overwhelming evidence that host lipids play a crucial role in the functioning of these replication factories. This review will underscore the recent findings on the roles that lipids play in virus RNA replication and particles production. Lipids are thought to help organize a functional virus replication factory by facilitating membrane curvature or binding to and stimulating the activity of the virus polymerase in the replication complex (Figure 1). Alternatively, lipids recruit core proteins to facilitate virus particles formation (Figure 2).
Figure 1. Lipids in virus RNA synthesis.
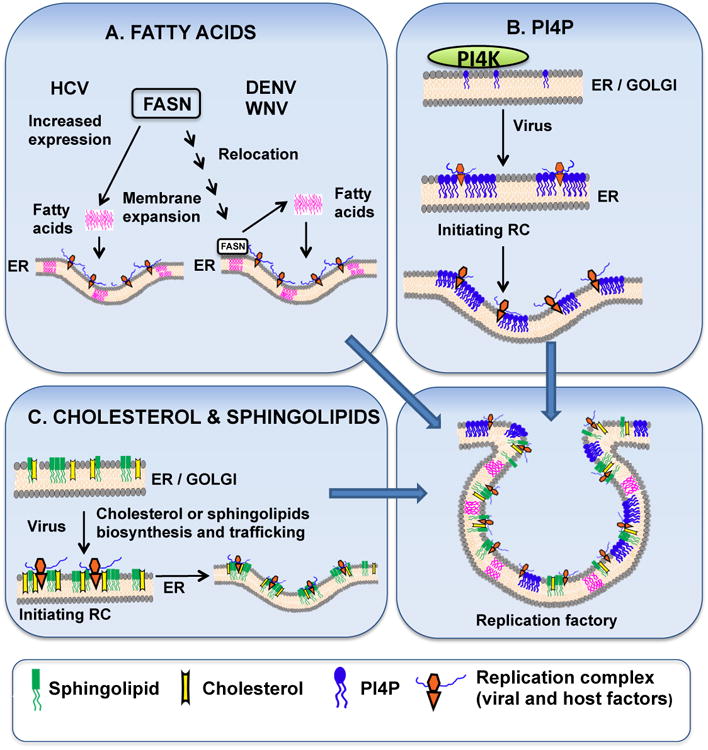
A. Fatty acids. The host protein, FASN, is a key enzyme in the biosynthesis of fatty acids, which are incorporated at sites of viral RNA replication and cause membrane expansion necessary to form the replication factory. FASN expression increases after HCV infection [23] while FASN is recruited to the sites of viral RNA replication after WNV or DENV infection [18,19,28]. B. PI4P. Following virus infection, PI4P is produced on ER membranes by PI4KIIIα or Golgi membranes by PI4KIIIβ. PI4P may bind to virus replicase proteins and contribute to membrane curvature. Alternatively, PI4P can recruit effector proteins via their PH domains, triggering a cascade of events which can lead to the organization of a functional replicase complex [101]. C. Cholesterol and sphingolipids. Biosynthesis and trafficking of cholesterol or sphingolipids are essential for optimal functioning of virus replicase complex. By changing the membrane composition and fluidity, these lipids can contribute to membrane bending and stimulation of the replicase complex.
Figure 2. Lipids in HCV maturation.
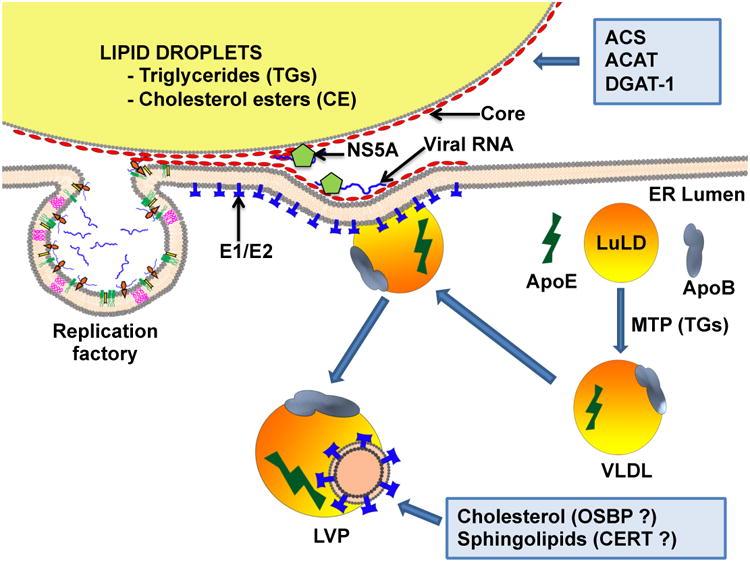
The first step in HCV assembly often takes place on LDs. HCV core localizes to LDs that are in proximity to ER-associated virus replication factory. Following replication, HCV NS5A brings the nascent RNA to LDs via interaction with DGAT1 and core proteins, thus initiating virus assembly [90]. The two enzymes, ACS and ACAT, are crucial for LDs biogenesis and facilitate HCV assembly [84]. Virus particles maturation, in the form of LVP, requires cholesterol and sphingolipids, which may be provided by OSBP and CERT, two PI4P effector proteins [98]. E1/E2: HCV glycoproteins; LuLD: luminal LDs; VLDL: very low density lipoprotein; LVP: lipo-viro-particle.
Lipids are essential for the integrity of cellular membranes and major alterations in lipid composition can negatively impact cellular homeostasis. However, subtle changes in membrane lipids pose a special challenge for RNA viruses that rely heavily on membranes for efficient genome replication and virus particles production. While these viruses need cytosolic membranes to replicate their genome, they all display specific lipid requirements. Hence, positive-strand RNA viruses employ multifaceted strategies to hijack host machinery involved in lipid biosynthesis and transport. This review will underscore the recent findings on the crucial roles of lipids in virus RNA replication and particles production. Due to space limitation, the review will feature the wealth of knowledge gained from studying the Flaviviridae and Picornaviridae families of viruses
Fatty acids and virus genome replication
Fatty acids are constituents of triglycerides and phospholipids. They contribute to energy production and storage, generation of lipid droplets and structural integrity of membranes. Inhibitors of fatty acids synthesis (e.g. cerulenin, C75) severely impede the replication of enteroviruses (e.g. poliovirus), flaviviruses (e.g. West Nile virus or WNV) or hepatitis C virus (HCV) [16-19]. Fatty acid synthase (FASN) is a key lipogenic enzyme involved in fatty acids synthesis (Table 1; Figure 1A). Indeed, FASN protein levels are increased in HCV-infected patients [20,21] or HCV-infected cells [22,23]. Notably, Nasheri et al. found that HCV NS4B was involved in FASN induction and concomitant increase in fatty acids production [23]. However, NS4B does not appear to bind to FASN protein or the regulatory sequences for FASN expression. Instead, NS4B activates the sterol regulatory element-binding protein (SREBP-1c), a major transcription factor for FASN expression [24,25]. Under this scenario, FASN-derived fatty acids, but not FASN itself, are recruited to build HCV replication factory [23], which consists of double-membrane vesicles (DMVs) organized into a membranous web [5,11,12,14,15,26]. The generation of DMVs likely requires additional phospholipids production to ensure minimal alteration in the lipid content of ER membrane. Indeed, HCV infection leads to increased levels of phosphatidylcholine and phosphatidylethanolamine [22]. Thus, while fatty acids may be directly recruited to the HCV replication factory, they can be converted into phospholipids to generate the DMVs. Alternatively, a recent report shows that FASN binds to, and stimulates HCV NS5B polymerase (RdRp) activity in the replication factory [27]. While this report needs to be confirmed, it implies that FASN plays at least two different roles during HCV replication: providing the building block for phospholipids and stimulating virus polymerase.
Table 1.
Host factors and related lipids in RNA virus replication.
| PROTEIN | LOCATION | FUNCTION | REFERENCES |
|---|---|---|---|
| FASN | Cytosol | Fatty acid synthesis for membrane expansion in virus replication factory. | [23,78] |
| PI4KIIIα | ER | PI4P synthesis; required for membranous web vesicles formation | [39,45] |
| PI4KIIIβ | Golgi | PI4P synthesis and stimulation of viral 3DPol in enteroviruses. | [101,102] |
| OSBP | ER and Golgi | Transports cholesterol to the HCV replication complex. | [67] |
| CERT | ER and Golgi | Transports ceramide from ER to Golgi for sphingomyelin biosynthesis; required for virus production. | [103] |
| DHCR24 | ER | Contributes to cholesterol synthesis; required for HCV genome replication. | [65] |
| DGAT1 | ER | Recruits HCV core and NS5A to lipid droplets: required for virus assembly. | [79,90,104] |
| ACS | ER | Fatty acyl-CoA synthesis for incorporation into triglycerides and cholesterol esters in lipid droplets; required for virus production. | [84] |
| ACAT | ER | Conversion of cholesterol to cholesterol esters in lipid droplets; required for virus production. | [84] |
FASN function is also required for WNV or dengue virus (DENV) RNA replication [18,28]. Nevertheless, these viruses do not induce an increase in FASN protein level. Instead, they cause a redistribution of FASN into their replication platform [18,28]. The DENV NS3 protein which has protease and RNA helicase activity, interacts with, and activates FASN to generate fatty acids, which are utilized to either build or stimulate the activity of DENV replicase complex [28]. There is no report yet that FASN affects poliovirus replication. However, poliovirus 2A protein is reported to stimulate the activity of another lipogenic enzyme, long chain acyl-CoA synthetase 3, thus providing the fatty acids required for poliovirus replication [29].
Phosphatidylinositol 4-phosphate and virus genome replication
Phosphatidylinositol 4-phosphate (PI4P) regulates membrane trafficking via interaction with effector proteins [30,31]. Its role in RNA virus replication (Figure 1B) was revealed in several studies in which phosphatidylinositol 4-kinase (PI4K) knockdown led to a significant decrease in enterovirus or HCV RNA synthesis [32-39]. There are four PI4K isoforms, with PI4KIIIα and PI4KIIIβ functioning in the ER and Golgi complex, respectively [30] (Table 1). HCV replication complex is ER-derived [5,6,10,14,40,41] and favors PI4KIIIα function for replication [34,35,39]. By contrast, enteroviruses build their replication platform on Golgi-derived membranes [3,7,42], hence require PI4KIIIβ for replication [33,43].
Following infection, enteroviruses and HCV recruit PI4KIIIα/β to their replication complex, leading to increased PI4P production [33,34,36,44]. For HCV, several studies have shown that NS5A protein interacts with, and activates PI4KIIIα [34,36,44]. A more recent report by Reiss et al. [45] shows that a conserved sequence in the C-terminal end of NS5A domain 1 is crucial for NS5A interaction with PI4KIIIα. NS5A is a phosphoprotein with two phosphorylation states based on its mobility on SDS-PAGE. The basal NS5A phosphorylation, p56, facilitates HCV genome replication [46] whereas the hyperphosphorylation state, p58, impedes HCV replication but appears to enhance virus production [46-48]. Notably, Reiss et al. [45] reported that PI4KIIIα plays a crucial role in NS5A phosphorylation. Indeed, mutations within the C-terminal end of NS5A domain 1 resulted in both a decrease in HCV genome replication and increased NS5A hyperphosphorylation. Hence, the NS5A-PI4KIIIα interaction is a crucial checkpoint for regulating the transition from HCV genome replication to virus particles production.
The role of enterovirus proteins in the recruitment of PI4KIIIβ remains controversial. Two studies found that enteroviruses 3A protein interacts with PI4KIIIβ [33,49] while coxsackievirus 3A mutation rendered the virus resistant to the inhibitors of PI4KIIIβ, enviroxime and GW5074 [43]. However, Teterina et al. [50] were unable to confirm the interaction between 3A and PI4KIIIβ.
Membranous web (MW) vesicles are formed during HCV infection [10-15,51] and appear as dots or foci in confocal microscopy [6,10,13,41,52]. PI4KIIIα knockdown causes a drastic change in the MW or the disruption of HCV replicase foci [35,36,39,44,45], implying a role for PI4KIIIα in the organization of the HCV replication factory. Alternatively, the PI4P, generated by PI4K, may help to organize or stimulate virus replicase complex (RC) because dephosphorylation of PI4P with SacI impedes HCV or poliovirus RNA synthesis [33,53]. In poliovirus, PI4P binds to the 3D polymerase either to anchor it in the virus RC or stimulate its enzymatic activity [33]. There is also evidence implicating PI4P effector proteins (e.g. oxysterol-binding protein or OSBP; Ceramide transport protein or CERT; four phosphate adaptor protein 2 or FAPP2) function in the transport of cholesterol or sphingolipids required for virus replication (Table 1). These effectors will be discussed below.
Cholesterol and sphingolipids in virus genome replication
Cholesterol and sphingolipids are major constituents of non-ionic detergent resistant lipid rafts [54,55]. The replicase complexes from HCV, enteroviruses or flaviviruses co-fractionate with lipid rafts while disruption of the lipid rafts interferes with RNA synthesis [56-60]. Consistent with these findings, pharmacological inhibition of the enzymes involved in cholesterol biosynthesis impedes virus RNA synthesis [60-63]. An enzyme involved in cholesterol synthesis, HMGCR (3-hydroxy-methyglutaryl-CoA reductase), relocalizes to WNV replication complex, whereas DHCR24 (24-dehydrocholesterol reductase) level is upregulated during HCV infection [64,65] (Table 1).
How cholesterol is recruited to the viral replicase complex is just beginning to be elucidated. Studies with HCV and enteroviruses show that OSBP (oxysterol-binding protein) is responsible for cholesterol transport [66,67]. Notably, knockdown or pharmacological inhibition of OSBP interferes with RNA replication and disrupts virus replication factory [66-68]. In poliovirus, cholesterol also plays a role in processing precursor 3CD into mature 3D polymerase [60]. Notably, PI4P binds to, and recruits OSBP to the virus replication complex [30]. Additionally, HCV NS5A also binds to OSBP, implying a concerted effort by host and virus factors to bring cholesterol into the virus replication platform [69].
High levels of sphingolipids have been reported in DENV or HCV-infected cells [19,70-74]. Specifically, sphingomyelin levels are elevated in the HCV replicase membrane fraction [70]. Yet, the role of sphingolipids during HCV replication is highlighted by the findings that inhibition of serine palmitoyltransferase (SPT), the first enzyme in the sphingolipid biosynthetic pathway, or knockdown of sphingomyelin synthase, leads to lower HCV replication efficiency [70-73]. Accordingly, sphingomyelin binds to, and activates HCV NS5B polymerase [70,75], implying that sphingomyelin stimulates the function of virus replicase complex. Recently, we have found that lactosylceramide, a glycosphingolipid, colocalizes with HCV replicase complex. Lactosylceramide production requires FAPP2, a PI4P effector protein. Our study suggests that FAPP2, or its transported glycosphingolipid, is crucial for generating functional HCV replication factory [52]. Hence, sphingolipids may contribute to the organization and function of virus replicase complex.
Role of lipids in virus production
The importance of VLDL (Figure 2) in virus particles formation has been reported previously [76-79]. This section will therefore underscore recent developments in the role of other lipids and proteins in regulating the transition from replication to virus assembly. Lipid droplets (LDs) are ER-derived organelles enriched in cholesterol esters and triglycerides [80,81]. LDs are recognized as sites for HCV (e.g. JFH1 strain) and DENV assembly [82,83]. Pharmacological inhibition of triglycerides and cholesterol esters synthesis impedes HCV assembly by decreasing the cellular LDs content or increasing LDs size [84]. Virus core proteins get recruited to LDs via interaction with diacylglycerol acyltransferase-1 (DGAT1) (Table 1) or tail-interacting protein of 47 kD (TIP47) [85,86], two proteins required for LDs biogenesis [87,88]. Knockdown or inhibition of DGAT1 activity hinders HCV assembly [86]. Additionally, the interaction of HCV NS5A with core protein is believed to transfer HCV RNA from replication to assembly sites [82,89] and two LDs-associated proteins, DGAT1 and Rab18, were reported recently to play a crucial role in NS5A-core interaction [90-92]. Nevertheless, the assembly of the highly infectious HCV Jc1 virus takes place on ER membranes and requires p7 function [93-96]. Hence, HCV may utilize LDs to modulate virus infectivity.
As an enveloped virus, host lipids also play a crucial role in the composition of the infectious HCV particles. Recent studies show elevated levels of sphingolipids in cell culture-derived HCV particles [77,97] (Figure 2). Furthermore, inhibition of cholesterol and sphingolipids biosynthesis, or knockdown of proteins involved in the transport of these lipids (OSBP and CERT) hampers HCV production [56,69,98]. Note that OSBP is also required for HCV RNA synthesis as discussed above [67].
Conclusions
Despite the replication factories of RNAs vary widely in structure, positive-strand RNA viruses co-opt similar host lipids and related proteins to amplify their genomes. Since RNA synthesis is a crucial step in viral lifecycle, drugs targeting host lipid metabolic pathways are likely to increase the efficacy of currently available direct acting antivirals. However, several questions remain unanswered. For example, are the discussed lipids and related proteins playing distinct roles during virus replication? Are the lipids involved in membrane curvature required for generating membrane vesicles in the replication factories? How does palmitoylation of HCV proteins contribute to their roles in virus replication and assembly [99,100]? Finally, do most viruses modulate host lipid metabolism to alter the innate or acquired immune response to infection [64]?
Highlights.
Fatty acids provide additional phospholipids in the replicase factory
PI4P stimulates virus proteins and recruits effectors crucial for RNA replication
Cholesterol and sphingolipids help to organize a functional replicase factory
Virus assembly on LDs requires interaction between virus and LDs-bound proteins
Acknowledgments
We are grateful to Israel Benain-Peral for suggestions and technical help with the manuscript. K.V.K is supported by the National Institute of Allergy and Infectious Diseases 1R21AI097858-01 grant.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
(*): papers of special interest
(**): papers of outstanding interest
- 1**.den Boon JA, Ahlquist P. Organelle-like membrane compartmentalization of positive-strand rna virus replication factories. Annual review of microbiology. 2010;64:241–256. doi: 10.1146/annurev.micro.112408.134012. Comprehensive review of the replication complexes from various positive-strand RNA viruses and their untimate connection to host membranes. [DOI] [PubMed] [Google Scholar]
- 2.den Boon JA, Diaz A, Ahlquist P. Cytoplasmic viral replication complexes. Cell Host Microbe. 2010;8(1):77–85. doi: 10.1016/j.chom.2010.06.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Belov GA, Sztul E. Rewiring of cellular membrane homeostasis by picornaviruses. J Virol. 2014 doi: 10.1128/JVI.00922-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4*.Mackenzie JM, Jones MK, Westaway EG. Markers for trans-golgi membranes and the intermediate compartment localize to induced membranes with distinct replication functions in flavivirus-infected cells. J Virol. 1999;73(11):9555–9567. doi: 10.1128/jvi.73.11.9555-9567.1999. This recent review defines the roles of the retrograde pathway, fatty acids, PI4P and cholesterol in the organiztion and function of poliovirus replication complex. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5*.Romero-Brey I, Merz A, Chiramel A, Lee JY, Chlanda P, Haselman U, Santarella-Mellwig R, Habermann A, Hoppe S, Kallis S, Walther P, et al. Three-dimensional architecture and biogenesis of membrane structures associated with hepatitis c virus replication. PLoS Pathog. 2012;8(12):e1003056. doi: 10.1371/journal.ppat.1003056. This study combines confocal microscopy, electron microscopy and electron tomography to reveal the architecture of HCV replication factory and the crucial role of HCV NS5A in initiating the formation of this factory. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Stone M, Jia S, Heo WD, Meyer T, Konan KV. Participation of rab5, an early endosome protein, in hepatitis c virus rna replication machinery. J Virol. 2007;81(9):4551–4563. doi: 10.1128/JVI.01366-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Suhy DA, Giddings TH, Kirkegaard K. Remodeling the endoplasmic reticulum by poliovirus infection and by individual viral proteins: An autophagy-like origin for virus-induced vesicles. J Virol. 2000;74(19):8953–8965. doi: 10.1128/jvi.74.19.8953-8965.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.van der Meer Y, Snijder EJ, Dobbe JC, Schleich S, Denison MR, Spaan WJ, Locker JK. Localization of mouse hepatitis virus nonstructural proteins and rna synthesis indicates a role for late endosomes in viral replication. J Virol. 1999;73(9):7641–7657. doi: 10.1128/jvi.73.9.7641-7657.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9*.Welsch S, Miller S, Romero-Brey I, Merz A, Bleck CK, Walther P, Fuller SD, Antony C, Krijnse-Locker J, Bartenschlager R. Composition and three-dimensional architecture of the dengue virus replication and assembly sites. Cell Host Microbe. 2009;5(4):365–375. doi: 10.1016/j.chom.2009.03.007. The authors utilize confocal microscopy, electron microscopy and electron tomography to reveal the architecture of DENV replication factory. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Aligo J, Jia S, Manna D, Konan KV. Formation and function of hepatitis c virus replication complexes require residues in the carboxy-terminal domain of ns4b protein. Virology. 2009;393(1):68–83. doi: 10.1016/j.virol.2009.07.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Egger D, Wolk B, Gosert R, Bianchi L, Blum HE, Moradpour D, Bienz K. Expression of hepatitis c virus proteins induces distinct membrane alterations including a candidate viral replication complex. J Virol. 2002;76(12):5974–5984. doi: 10.1128/JVI.76.12.5974-5984.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Gosert R, Egger D, Lohmann V, Bartenschlager R, Blum HE, Bienz K, Moradpour D. Identification of the hepatitis c virus rna replication complex in huh-7 cells harboring subgenomic replicons. J Virol. 2003;77(9):5487–5492. doi: 10.1128/JVI.77.9.5487-5492.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Han Q, Aligo J, Manna D, Belton K, Chintapalli SV, Hong Y, Patterson RL, van Rossum DB, Konan KV. Conserved gxxxg- and s/t-like motifs in the transmembrane domains of ns4b protein are required for hepatitis c virus replication. J Virol. 2011;85(13):6464–6479. doi: 10.1128/JVI.02298-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Konan KV, Giddings TH, Jr, Ikeda M, Li K, Lemon SM, Kirkegaard K. Nonstructural protein precursor ns4a/b from hepatitis c virus alters function and ultrastructure of host secretory apparatus. J Virol. 2003;77(14):7843–7855. doi: 10.1128/JVI.77.14.7843-7855.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15*.Paul D, Romero-Brey I, Gouttenoire J, Stoitsova S, Krijnse-Locker J, Moradpour D, Bartenschlager R. Ns4b self-interaction through conserved c-terminal elements is required for the establishment of functional hepatitis c virus replication complexes. J Virol. 2011;85(14):6963–6976. doi: 10.1128/JVI.00502-11. This study demonstrates the significance of HCV NS4b self-interaction in the functional organiation of HCV replication factory. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Fogg MH, Teterina NL, Ehrenfeld E. Membrane requirements for uridylylation of the poliovirus vpg protein and viral rna synthesis in vitro. J Virol. 2003;77(21):11408–11416. doi: 10.1128/JVI.77.21.11408-11416.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kapadia SB, Chisari FV. Hepatitis c virus rna replication is regulated by host geranylgeranylation and fatty acids. Proc Natl Acad Sci U S A. 2005;102(7):2561–2566. doi: 10.1073/pnas.0409834102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18*.Martin-Acebes MA, Blazquez AB, Jimenez de Oya N, Escribano-Romero E, Saiz JC. West nile virus replication requires fatty acid synthesis but is independent on phosphatidylinositol-4-phosphate lipids. PLoS One. 2011;6(9):e24970. doi: 10.1371/journal.pone.0024970. The authors reveal the importance of fatty acids in WNV replicatoin. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19**.Perera R, Riley C, Isaac G, Hopf-Jannasch AS, Moore RJ, Weitz KW, Pasa-Tolic L, Metz TO, Adamec J, Kuhn RJ. Dengue virus infection perturbs lipid homeostasis in infected mosquito cells. PLoS Pathog. 2012;8(3):e1002584. doi: 10.1371/journal.ppat.1002584. The study reveals the upregulation and redistribution of specific lipids during DENV infection. Fatty acids are enriched in DENV replication complex and crucial for DENV replication. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Aragones G, Alonso-Villaverde C, Oliveras-Ferraros C, Beltran-Debon R, Rull A, Rodriguez-Sanabria F, Camps J, Martin AV, Menendez JA, Joven J. Infection with hiv and hcv enhances the release of fatty acid synthase into circulation: Evidence for a novel indicator of viral infection. BMC gastroenterology. 2010;10:92. doi: 10.1186/1471-230X-10-92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21*.Joven J, Espinel E, Rull A, Beltran-Debon R, Aragones G, Rodriguez-Gallego E, Camps J, Pedro-Botet J, Sans T, Menendez JA, Alonso-Villaverde C. Serum fatty acid synthase concentration is increased in patients with hepatitis viral infection and may assist in the prediction of liver steatosis. Journal of clinical virology : the official publication of the Pan American Society for Clinical Virology. 2011;51(3):199–201. doi: 10.1016/j.jcv.2011.04.003. This study reveals a putative role of FASN in HCV pathogenesis. [DOI] [PubMed] [Google Scholar]
- 22*.Diamond DL, Syder AJ, Jacobs JM, Sorensen CM, Walters KA, Proll SC, McDermott JE, Gritsenko MA, Zhang Q, Zhao R, Metz TO, et al. Temporal proteome and lipidome profiles reveal hepatitis c virus-associated reprogramming of hepatocellular metabolism and bioenergetics. PLoS Pathog. 2010;6(1):e1000719. doi: 10.1371/journal.ppat.1000719. The authors demonstrate an upregulation of fatty acid synthesis during HCV infection. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Nasheri N, Joyce M, Rouleau Y, Yang P, Yao S, Tyrrell DL, Pezacki JP. Modulation of fatty acid synthase enzyme activity and expression during hepatitis c virus replication. Chemistry & biology. 2013;20(4):570–582. doi: 10.1016/j.chembiol.2013.03.014. [DOI] [PubMed] [Google Scholar]
- 24.Park CY, Jun HJ, Wakita T, Cheong JH, Hwang SB. Hepatitis c virus nonstructural 4b protein modulates sterol regulatory element-binding protein signaling via the akt pathway. J Biol Chem. 2009;284(14):9237–9246. doi: 10.1074/jbc.M808773200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Waris G, Felmlee DJ, Negro F, Siddiqui A. Hepatitis c virus induces proteolytic cleavage of sterol regulatory element binding proteins and stimulates their phosphorylation via oxidative stress. J Virol. 2007;81(15):8122–8130. doi: 10.1128/JVI.00125-07. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 26.Wolk B, Buchele B, Moradpour D, Rice CM. A dynamic view of hepatitis c virus replication complexes. J Virol. 2008;82(21):10519–10531. doi: 10.1128/JVI.00640-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27*.Huang JT, Tseng CP, Liao MH, Lu SC, Yeh WZ, Sakamoto N, Chen CM, Cheng JC. Hepatitis c virus replication is modulated by the interaction of nonstructural protein ns5b and fatty acid synthase. J Virol. 2013;87(9):4994–5004. doi: 10.1128/JVI.02526-12. This study suggests that HCV NS5B recruits FASN to the virus replication factory to facilitate replication. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28*.Heaton NS, Perera R, Berger KL, Khadka S, Lacount DJ, Kuhn RJ, Randall G. Dengue virus nonstructural protein 3 redistributes fatty acid synthase to sites of viral replication and increases cellular fatty acid synthesis. Proc Natl Acad Sci U S A. 2010;107(40):17345–17350. doi: 10.1073/pnas.1010811107. This study demonstrates that DENV NS3 recruits FASN to the virus replication platform to stimulate replication. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29**.Nchoutmboube JA, Viktorova EG, Scott AJ, Ford LA, Pei Z, Watkins PA, Ernst RK, Belov GA. Increased long chain acyl-coa synthetase activity and fatty acid import is linked to membrane synthesis for development of picornavirus replication organelles. PLoS Pathog. 2013;9(6):e1003401. doi: 10.1371/journal.ppat.1003401. The authors show a novel function for poliovirus 2A protein: activattion of acyl-coa synthetase and recruitment of fatty acids to the virus replication platform. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Balla A, Balla T. Phosphatidylinositol 4-kinases: Old enzymes with emerging functions. Trends Cell Biol. 2006;16(7):351–361. doi: 10.1016/j.tcb.2006.05.003. [DOI] [PubMed] [Google Scholar]
- 31.Santiago-Tirado FH, Bretscher A. Membrane-trafficking sorting hubs: Cooperation between pi4p and small gtpases at the trans-golgi network. Trends Cell Biol. 2011;21(9):515–525. doi: 10.1016/j.tcb.2011.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Borawski J, Troke P, Puyang X, Gibaja V, Zhao S, Mickanin C, Leighton-Davies J, Wilson CJ, Myer V, Cornellataracido I, Baryza J, et al. Class iii phosphatidylinositol 4-kinase alpha and beta are novel host factor regulators of hepatitis c virus replication. J Virol. 2009;83(19):10058–10074. doi: 10.1128/JVI.02418-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33**.Hsu NY, Ilnytska O, Belov G, Santiana M, Chen YH, Takvorian PM, Pau C, van der Schaar H, Kaushik-Basu N, Balla T, Cameron CE, et al. Viral reorganization of the secretory pathway generates distinct organelles for rna replication. Cell. 2010;141(5):799–811. doi: 10.1016/j.cell.2010.03.050. The authors show that poliovirus 3A protein recruits PI4KIII beta to the virus replication complex to stimulate poliovirus replication via interaction of the 3D polymerase with PI4P. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34**.Reiss S, Rebhan I, Backes P, Romero-Brey I, Erfle H, Matula P, Kaderali L, Poenisch M, Blankenburg H, Hiet MS, Longerich T, et al. Recruitment and activation of a lipid kinase by hepatitis c virus ns5a is essential for integrity of the membranous replication compartment. Cell Host Microbe. 2011;9(1):32–45. doi: 10.1016/j.chom.2010.12.002. This study shows that HCV NS5A protein binds to and activates PI4KIII alpha to build the functional virus replication fatory. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35*.Tai AW, Benita Y, Peng LF, Kim SS, Sakamoto N, Xavier RJ, Chung RT. A functional genomic screen identifies cellular cofactors of hepatitis c virus replication. Cell Host Microbe. 2009;5(3):298–307. doi: 10.1016/j.chom.2009.02.001. This study utilized siRNA library screen to identify PI4KIII alpha as a crucial factor in HCV replication. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Tai AW, Salloum S. The role of the phosphatidylinositol 4-kinase pi4ka in hepatitis c virus-induced host membrane rearrangement. PLoS One. 2011;6(10):e26300. doi: 10.1371/journal.pone.0026300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Trotard M, Lepere-Douard C, Regeard M, Piquet-Pellorce C, Lavillette D, Cosset FL, Gripon P, Le Seyec J. Kinases required in hepatitis c virus entry and replication highlighted by small interference rna screening. FASEB J. 2009;23(11):3780–3789. doi: 10.1096/fj.09-131920. [DOI] [PubMed] [Google Scholar]
- 38.Vaillancourt FH, Pilote L, Cartier M, Lippens J, Liuzzi M, Bethell RC, Cordingley MG, Kukolj G. Identification of a lipid kinase as a host factor involved in hepatitis c virus rna replication. Virology. 2009;387(1):5–10. doi: 10.1016/j.virol.2009.02.039. [DOI] [PubMed] [Google Scholar]
- 39**.Berger KL, Cooper JD, Heaton NS, Yoon R, Oakland TE, Jordan TX, Mateu G, Grakoui A, Randall G. Roles for endocytic trafficking and phosphatidylinositol 4-kinase iii alpha in hepatitis c virus replication. Proc Natl Acad Sci U S A. 2009;106(18):7577–7582. doi: 10.1073/pnas.0902693106. This study combined siRNA libray screen, electron microsocopy and subcellular farctionation to tidentify PI4KIII alpha as a crucial factor in the organization and functin of the HCV replication platform. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Egger D, Teterina N, Ehrenfeld E, Bienz K. Formation of the poliovirus replication complex requires coupled viral translation, vesicle production, and viral rna synthesis. J Virol. 2000;74(14):6570–6580. doi: 10.1128/jvi.74.14.6570-6580.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Manna D, Aligo J, Xu C, Park WS, Koc H, Heo WD, Konan KV. Endocytic rab proteins are required for hepatitis c virus replication complex formation. Virology. 2010;398(1):21–37. doi: 10.1016/j.virol.2009.11.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42**.Belov GA, Nair V, Hansen BT, Hoyt FH, Fischer ER, Ehrenfeld E. Complex dynamic development of poliovirus membranous replication complexes. J Virol. 2012;86(1):302–312. doi: 10.1128/JVI.05937-11. This study combined electron microscopy and electron tomography to visualize poliovirus replication platform. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.van der Schaar HM, van der Linden L, Lanke KH, Strating JR, Purstinger G, de Vries E, de Haan CA, Neyts J, van Kuppeveld FJ. Coxsackievirus mutants that can bypass host factor pi4kiiibeta and the need for high levels of pi4p lipids for replication. Cell research. 2012;22(11):1576–1592. doi: 10.1038/cr.2012.129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44*.Berger KL, Kelly SM, Jordan TX, Tartell MA, Randall G. Hepatitis c virus stimulates the phosphatidylinositol 4-kinase iii alpha-dependent phosphatidylinositol 4-phosphate production that is essential for its replication. J Virol. 2011;85(17):8870–8883. doi: 10.1128/JVI.00059-11. This study demonstrates the role of NS5A in the recruitment of PI4KIII alpha to HCV replication factory. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45**.Reiss S, Harak C, Romero-Brey I, Radujkovic D, Klein R, Ruggieri A, Rebhan I, Bartenschlager R, Lohmann V. The lipid kinase phosphatidylinositol-4 kinase iii alpha regulates the phosphorylation status of hepatitis c virus ns5a. PLoS Pathog. 2013;9(5):e1003359. doi: 10.1371/journal.ppat.1003359. The authors show that PI4KIII alpha phosphorylates HCV NS5A and regulates its roles in HCV replication and virus production. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Appel N, Pietschmann T, Bartenschlager R. Mutational analysis of hepatitis c virus nonstructural protein 5a: Potential role of differential phosphorylation in rna replication and identification of a genetically flexible domain. J Virol. 2005;79(5):3187–3194. doi: 10.1128/JVI.79.5.3187-3194.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47*.Han Q, Manna D, Belton K, Cole R, Konan KV. Modulation of hepatitis c virus genome encapsidation by nonstructural protein 4b. J Virol. 2013;87(13):7409–7422. doi: 10.1128/JVI.03523-12. This study indicates that NS4b C-terminal domain regulates the phosphorylation state of HCV NS5A and its role in virus assembly. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Tellinghuisen TL, Foss KL, Treadaway J. Regulation of hepatitis c virion production via phosphorylation of the ns5a protein. PLoS Pathog. 2008;4(3):e1000032. doi: 10.1371/journal.ppat.1000032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Greninger AL, Knudsen GM, Betegon M, Burlingame AL, Derisi JL. The 3a protein from multiple picornaviruses utilizes the golgi adaptor protein acbd3 to recruit pi4kiiibeta. J Virol. 2012;86(7):3605–3616. doi: 10.1128/JVI.06778-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Teterina NL, Pinto Y, Weaver JD, Jensen KS, Ehrenfeld E. Analysis of poliovirus protein 3a interactions with viral and cellular proteins in infected cells. J Virol. 2011;85(9):4284–4296. doi: 10.1128/JVI.02398-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Rouille Y, Helle F, Delgrange D, Roingeard P, Voisset C, Blanchard E, Belouzard S, McKeating J, Patel AH, Maertens G, Wakita T, et al. Subcellular localization of hepatitis c virus structural proteins in a cell culture system that efficiently replicates the virus. J Virol. 2006;80(6):2832–2841. doi: 10.1128/JVI.80.6.2832-2841.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52**.Khan I, Katikaneni DS, Han Q, Sanchez-Felipe L, Hanada K, Ambrose RL, Mackenzie JM, Konan KV. Modulation of hepatitis c virus genome replication by glycosphingolipids and fapp2 protein. J Virol. 2014 doi: 10.1128/JVI.00970-14. This study demonstartes that HCV induces PI4P production in part to hijack fapp2 protein and glycosphingolipids required for virus replication. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Zhang L, Hong Z, Lin W, Shao RX, Goto K, Hsu VW, Chung RT. Arf1 and gbf1 generate a pi4p-enriched environment supportive of hepatitis c virus replication. PLoS One. 2012;7(2):e32135. doi: 10.1371/journal.pone.0032135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Anderson RG, Jacobson K. A role for lipid shells in targeting proteins to caveolae, rafts, and other lipid domains. Science. 2002;296(5574):1821–1825. doi: 10.1126/science.1068886. [DOI] [PubMed] [Google Scholar]
- 55.Simons K, Toomre D. Lipid rafts and signal transduction. Nat Rev Mol Cell Biol. 2000;1(1):31–39. doi: 10.1038/35036052. [DOI] [PubMed] [Google Scholar]
- 56*.Aizaki H, Morikawa K, Fukasawa M, Hara H, Inoue Y, Tani H, Saito K, Nishijima M, Hanada K, Matsuura Y, Lai MM, et al. Critical role of virion-associated cholesterol and sphingolipid in hepatitis c virus infection. J Virol. 2008;82(12):5715–5724. doi: 10.1128/JVI.02530-07. The authors show that cholesterole and shphingolipids are associated with HC particles and are crucial for infectious virus particles production. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Shi ST, Lee KJ, Aizaki H, Hwang SB, Lai MM. Hepatitis c virus rna replication occurs on a detergent-resistant membrane that cofractionates with caveolin-2. J Virol. 2003;77(7):4160–4168. doi: 10.1128/JVI.77.7.4160-4168.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Garcia Cordero J, Leon Juarez M, Gonzalez YMJA, Cedillo Barron L, Gutierrez Castaneda B. Caveolin-1 in lipid rafts interacts with dengue virus ns3 during polyprotein processing and replication in hmec-1 cells. PLoS One. 2014;9(3):e90704. doi: 10.1371/journal.pone.0090704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Sagan SM, Rouleau Y, Leggiadro C, Supekova L, Schultz PG, Su AI, Pezacki JP. The influence of cholesterol and lipid metabolism on host cell structure and hepatitis c virus replication. Biochemistry and cell biology = Biochimie et biologie cellulaire. 2006;84(1):67–79. doi: 10.1139/o05-149. [DOI] [PubMed] [Google Scholar]
- 60**.Ilnytska O, Santiana M, Hsu NY, Du WL, Chen YH, Viktorova EG, Belov G, Brinker A, Storch J, Moore C, Dixon JL, et al. Enteroviruses harness the cellular endocytic machinery to remodel the host cell cholesterol landscape for effective viral replication. Cell Host Microbe. 2013;14(3):281–293. doi: 10.1016/j.chom.2013.08.002. This study shows that enteroviruses hijack cholesterol from lipid rafts, in the plasma membrane, to facilitate polyprotein processing and virus replication. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kapadia SB, Brideau-Andersen A, Chisari FV. Interference of hepatitis c virus rna replication by short interfering rnas. Proc Natl Acad Sci U S A. 2003;100(4):2014–2018. doi: 10.1073/pnas.252783999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ye J, Wang C, Sumpter R, Jr, Brown MS, Goldstein JL, Gale M., Jr Disruption of hepatitis c virus rna replication through inhibition of host protein geranylgeranylation. Proc Natl Acad Sci U S A. 2003;100(26):15865–15870. doi: 10.1073/pnas.2237238100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63*.Paul D, Hoppe S, Saher G, Krijnse-Locker J, Bartenschlager R. Morphological and biochemical characterization of the membranous hepatitis c virus replication compartment. J Virol. 2013;87(19):10612–10627. doi: 10.1128/JVI.01370-13. The authors utlize a novel approach to isolate functional HCV replication factory and validate the double membrane vesicles which constitue the membranous web. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Mackenzie JM, Khromykh AA, Parton RG. Cholesterol manipulation by west nile virus perturbs the cellular immune response. Cell Host Microbe. 2007;2(4):229–239. doi: 10.1016/j.chom.2007.09.003. [DOI] [PubMed] [Google Scholar]
- 65*.Takano T, Tsukiyama-Kohara K, Hayashi M, Hirata Y, Satoh M, Tokunaga Y, Tateno C, Hayashi Y, Hishima T, Funata N, Sudoh M, et al. Augmentation of dhcr24 expression by hepatitis c virus infection facilitates viral replication in hepatocytes. J Hepatol. 2011;55(3):512–521. doi: 10.1016/j.jhep.2010.12.011. This study indicates that HCV upreguates cholesterol biosynthetic protein, dhcr24, to enhance virus replication. [DOI] [PubMed] [Google Scholar]
- 66*.Arita M. Phosphatidylinositol-4 kinase iii beta and oxysterol-binding protein accumulate unesterified cholesterol on poliovirus-induced membrane structure. Microbiol Immunol. 2014;58(4):239–256. doi: 10.1111/1348-0421.12144. This study suggests a role for OSBP in the formation of poliovirus replication factory. [DOI] [PubMed] [Google Scholar]
- 67**.Wang H, Perry JW, Lauring AS, Neddermann P, De Francesco R, Tai AW. Oxysterol-binding protein is a phosphatidylinositol 4-kinase effector required for hcv replication membrane integrity and cholesterol trafficking. Gastroenterology. 2014;146(5):1373–1385. e1371–1311. doi: 10.1053/j.gastro.2014.02.002. The authors demontsrate that oxysterol-binding protein transports cholesterol to HCV replication factory factory to faciliate virus replication. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68*.Arita M, Kojima H, Nagano T, Okabe T, Wakita T, Shimizu H. Oxysterol-binding protein family i is the target of minor enviroxime-like compounds. J Virol. 2013;87(8):4252–4260. doi: 10.1128/JVI.03546-12. The authors show that oxysterol-binding protein inhibitorts impede poliovirus replication. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69*.Amako Y, Sarkeshik A, Hotta H, Yates J, 3rd, Siddiqui A. Role of oxysterol binding protein in hepatitis c virus infection. J Virol. 2009;83(18):9237–9246. doi: 10.1128/JVI.00958-09. This study shows that oxysterol-binding protein is required for HCV particles production. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70*.Hirata Y, Ikeda K, Sudoh M, Tokunaga Y, Suzuki A, Weng L, Ohta M, Tobita Y, Okano K, Ozeki K, Kawasaki K, et al. Self-enhancement of hepatitis c virus replication by promotion of specific sphingolipid biosynthesis. PLoS Pathog. 2012;8(8):e1002860. doi: 10.1371/journal.ppat.1002860. This study demonstrates the role of sphingomyelin in the functional organization of HCV replication factory. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Ciesek S, Steinmann E, Manns MP, Wedemeyer H, Pietschmann T. The suppressive effect that myriocin has on hepatitis c virus rna replication is independent of inhibition of serine palmitoyl transferase. The Journal of infectious diseases. 2008;198(7):1091–1093. doi: 10.1086/591463. [DOI] [PubMed] [Google Scholar]
- 72.Sakamoto H, Okamoto K, Aoki M, Kato H, Katsume A, Ohta A, Tsukuda T, Shimma N, Aoki Y, Arisawa M, Kohara M, et al. Host sphingolipid biosynthesis as a target for hepatitis c virus therapy. Nat Chem Biol. 2005;1(6):333–337. doi: 10.1038/nchembio742. [DOI] [PubMed] [Google Scholar]
- 73.Umehara T, Sudoh M, Yasui F, Matsuda C, Hayashi Y, Chayama K, Kohara M. Serine palmitoyltransferase inhibitor suppresses hcv replication in a mouse model. Biochem Biophys Res Commun. 2006;346(1):67–73. doi: 10.1016/j.bbrc.2006.05.085. [DOI] [PubMed] [Google Scholar]
- 74.Yamaji T, Kumagai K, Tomishige N, Hanada K. Two sphingolipid transfer proteins, cert and fapp2: Their roles in sphingolipid metabolism. IUBMB life. 2008;60(8):511–518. doi: 10.1002/iub.83. [DOI] [PubMed] [Google Scholar]
- 75*.Weng L, Hirata Y, Arai M, Kohara M, Wakita T, Watashi K, Shimotohno K, He Y, Zhong J, Toyoda T. Sphingomyelin activates hepatitis c virus rna polymerase in a genotype-specific manner. J Virol. 2010;84(22):11761–11770. doi: 10.1128/JVI.00638-10. This sudy demontsrates that sphingomyelin binds to and enhances HCV polymerase activity. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Alvisi G, Madan V, Bartenschlager R. Hepatitis c virus and host cell lipids: An intimate connection. RNA biology. 2011;8(2):258–269. doi: 10.4161/rna.8.2.15011. [DOI] [PubMed] [Google Scholar]
- 77*.Bartenschlager R, Penin F, Lohmann V, Andre P. Assembly of infectious hepatitis c virus particles. Trends in microbiology. 2011;19(2):95–103. doi: 10.1016/j.tim.2010.11.005. This review analyzes the various roles of lipids in infectious HCV production. [DOI] [PubMed] [Google Scholar]
- 78**.Heaton NS, Randall G. Multifaceted roles for lipids in viral infection. Trends in microbiology. 2011;19(7):368–375. doi: 10.1016/j.tim.2011.03.007. This review highlights the roles of lipids in RNA viruses replication. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Herker E, Ott M. Unique ties between hepatitis c virus replication and intracellular lipids. Trends Endocrinol Metab. 2011;22(6):241–248. doi: 10.1016/j.tem.2011.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Walther TC, Farese RV., Jr The life of lipid droplets. Biochim Biophys Acta. 2009;1791(6):459–466. doi: 10.1016/j.bbalip.2008.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Martin S, Parton RG. Lipid droplets: A unified view of a dynamic organelle. Nat Rev Mol Cell Biol. 2006;7(5):373–378. doi: 10.1038/nrm1912. [DOI] [PubMed] [Google Scholar]
- 82**.Miyanari Y, Atsuzawa K, Usuda N, Watashi K, Hishiki T, Zayas M, Bartenschlager R, Wakita T, Hijikata M, Shimotohno K. The lipid droplet is an important organelle for hepatitis c virus production. Nat Cell Biol. 2007;9(9):1089–1097. doi: 10.1038/ncb1631. This study demonstrates the crucial role of lipid droplets in the assembly of infectious HCV particles. [DOI] [PubMed] [Google Scholar]
- 83**.Samsa MM, Mondotte JA, Iglesias NG, Assuncao-Miranda I, Barbosa-Lima G, Da Poian AT, Bozza PT, Gamarnik AV. Dengue virus capsid protein usurps lipid droplets for viral particle formation. PLoS Pathog. 2009;5(10):e1000632. doi: 10.1371/journal.ppat.1000632. The authors demonstrate the crucial role of lipid droplets in the assembly of infectious DENV particles. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84*.Liefhebber JM, Hague CV, Zhang Q, Wakelam MJ, McLauchlan J. Modulation of triglyceride and cholesterol ester synthesis impairs assembly of infectious hepatitis c virus. J Biol Chem. 2014 doi: 10.1074/jbc.M114.582999. This study examines the roles of lipid droplets components in the formation of infectious HCV particles. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Carvalho FA, Carneiro FA, Martins IC, Assuncao-Miranda I, Faustino AF, Pereira RM, Bozza PT, Castanho MA, Mohana-Borges R, Da Poian AT, Santos NC. Dengue virus capsid protein binding to hepatic lipid droplets (ld) is potassium ion dependent and is mediated by ld surface proteins. J Virol. 2012;86(4):2096–2108. doi: 10.1128/JVI.06796-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86**.Herker E, Harris C, Hernandez C, Carpentier A, Kaehlcke K, Rosenberg AR, Farese RV, Jr, Ott M. Efficient hepatitis c virus particle formation requires diacylglycerol acyltransferase-1. Nature medicine. 2010;16(11):1295–1298. doi: 10.1038/nm.2238. The authors demonstrate that the recruitment of HCV core protein to lipid droplets requires its interaction with diacylglycerol acyltransferase-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Harris CA, Haas JT, Streeper RS, Stone SJ, Kumari M, Yang K, Han X, Brownell N, Gross RW, Zechner R, Farese RV., Jr Dgat enzymes are required for triacylglycerol synthesis and lipid droplets in adipocytes. Journal of lipid research. 2011;52(4):657–667. doi: 10.1194/jlr.M013003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Bulankina AV, Deggerich A, Wenzel D, Mutenda K, Wittmann JG, Rudolph MG, Burger KN, Honing S. Tip47 functions in the biogenesis of lipid droplets. J Cell Biol. 2009;185(4):641–655. doi: 10.1083/jcb.200812042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Masaki T, Suzuki R, Murakami K, Aizaki H, Ishii K, Murayama A, Date T, Matsuura Y, Miyamura T, Wakita T, Suzuki T. Interaction of hepatitis c virus nonstructural protein 5a with core protein is critical for the production of infectious virus particles. J Virol. 2008;82(16):7964–7976. doi: 10.1128/JVI.00826-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90*.Camus G, Herker E, Modi AA, Haas JT, Ramage HR, Farese RV, Jr, Ott M. Diacylglycerol acyltransferase-1 localizes hepatitis c virus ns5a protein to lipid droplets and enhances ns5a interaction with the viral capsid core. J Biol Chem. 2013;288(14):9915–9923. doi: 10.1074/jbc.M112.434910. This study shows that diacylglycerol acyltransferase-1 acts a bridge for ns5a-core interaction on lipid droplets. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Salloum S, Wang H, Ferguson C, Parton RG, Tai AW. Rab18 binds to hepatitis c virus ns5a and promotes interaction between sites of viral replication and lipid droplets. PLoS Pathog. 2013;9(8):e1003513. doi: 10.1371/journal.ppat.1003513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Ozeki S, Cheng J, Tauchi-Sato K, Hatano N, Taniguchi H, Fujimoto T. Rab18 localizes to lipid droplets and induces their close apposition to the endoplasmic reticulum-derived membrane. J Cell Sci. 2005;118(Pt 12):2601–2611. doi: 10.1242/jcs.02401. [DOI] [PubMed] [Google Scholar]
- 93.Boson B, Granio O, Bartenschlager R, Cosset FL. A concerted action of hepatitis c virus p7 and nonstructural protein 2 regulates core localization at the endoplasmic reticulum and virus assembly. PLoS Pathog. 2011;7(7):e1002144. doi: 10.1371/journal.ppat.1002144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Lindenbach BD, Evans MJ, Syder AJ, Wolk B, Tellinghuisen TL, Liu CC, Maruyama T, Hynes RO, Burton DR, McKeating JA, Rice CM. Complete replication of hepatitis c virus in cell culture. Science. 2005;309(5734):623–626. doi: 10.1126/science.1114016. [DOI] [PubMed] [Google Scholar]
- 95.Lindenbach BD, Meuleman P, Ploss A, Vanwolleghem T, Syder AJ, McKeating JA, Lanford RE, Feinstone SM, Major ME, Leroux-Roels G, Rice CM. Cell culture-grown hepatitis c virus is infectious in vivo and can be recultured in vitro. Proc Natl Acad Sci U S A. 2006;103(10):3805–3809. doi: 10.1073/pnas.0511218103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Pietschmann T, Kaul A, Koutsoudakis G, Shavinskaya A, Kallis S, Steinmann E, Abid K, Negro F, Dreux M, Cosset FL, Bartenschlager R. Construction and characterization of infectious intragenotypic and intergenotypic hepatitis c virus chimeras. Proc Natl Acad Sci U S A. 2006;103(19):7408–7413. doi: 10.1073/pnas.0504877103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97*.Merz A, Long G, Hiet MS, Brugger B, Chlanda P, Andre P, Wieland F, Krijnse-Locker J, Bartenschlager R. Biochemical and morphological properties of hepatitis c virus particles and determination of their lipidome. J Biol Chem. 2011;286(4):3018–3032. doi: 10.1074/jbc.M110.175018. This study show the the lipid composition of purified HCV particles and underscores the roles of shingolipids and cholesterol in infectious HCV production. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98*.Amako Y, Syed GH, Siddiqui A. Protein kinase d negatively regulates hepatitis c virus secretion through phosphorylation of oxysterol-binding protein and ceramide transfer protein. J Biol Chem. 2011;286(13):11265–11274. doi: 10.1074/jbc.M110.182097. The findings from this study suggest that oxysterol-binding protein and ceramide transfer protein are required for HCV production. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Majeau N, Fromentin R, Savard C, Duval M, Tremblay MJ, Leclerc D. Palmitoylation of hepatitis c virus core protein is important for virion production. J Biol Chem. 2009;284(49):33915–33925. doi: 10.1074/jbc.M109.018549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Yu GY, Lee KJ, Gao L, Lai MM. Palmitoylation and polymerization of hepatitis c virus ns4b protein. J Virol. 2006;80(12):6013–6023. doi: 10.1128/JVI.00053-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Lee C. Roles of phosphoinositides and phosphoinositides kinases in hepatitis c virus rna replication. Archives of pharmacal research. 2012;35(10):1701–1711. doi: 10.1007/s12272-012-1001-2. [DOI] [PubMed] [Google Scholar]
- 102.Nagy PD, Pogany J. The dependence of viral rna replication on co-opted host factors. Nat Rev Microbiol. 2012;10(2):137–149. doi: 10.1038/nrmicro2692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Hanada K, Kumagai K, Tomishige N, Kawano M. Cert and intracellular trafficking of ceramide. Biochim Biophys Acta. 2007;1771(6):644–653. doi: 10.1016/j.bbalip.2007.01.009. [DOI] [PubMed] [Google Scholar]
- 104.Sung PS, Murayama A, Kang W, Kim MS, Yoon SK, Fukasawa M, Kondoh M, Kim JS, Kim H, Kato T, Shin EC. Hepatitis c virus entry is impaired by claudin-1 downregulation in diacylglycerol acyltransferase-1 (dgat1)-deficient cells. J Virol. 2014 doi: 10.1128/JVI.01428-14. [DOI] [PMC free article] [PubMed] [Google Scholar]