

Abstract
A feedback between decomposition and litter chemical composition occurs with decomposition altering composition that in turn influences the decomposition rate. Elucidating the temporal pattern of chemical composition is vital to understand this feedback, but the effects of plant species and climate on chemical changes remain poorly understood, especially over multiple years. In a 10-year decomposition experiment with litter of four species (Acer saccharum, Drypetes glauca, Pinus resinosa, and Thuja plicata) from four sites that range from the arctic to tropics, we determined the abundance of 11 litter chemical constituents that were grouped into waxes, carbohydrates, lignin/tannins, and proteins/peptides using advanced 13C solid-state NMR techniques. Decomposition generally led to an enrichment of waxes and a depletion of carbohydrates, whereas the changes of other chemical constituents were inconsistent. Inconsistent convergence in chemical compositions during decomposition was observed among different litter species across a range of site conditions, whereas one litter species converged under different climate conditions. Our data clearly demonstrate that plant species rather than climate greatly alters the temporal pattern of litter chemical composition, suggesting the decomposition-chemistry feedback varies among different plant species.
Litter decomposition plays an important role in the terrestrial carbon (C) cycle1 and hence the factors controlling its rate are important to understand. Much of the previous work on this process has examined the effect of initial substrate quality2,3. However, during the course of decomposition, which takes years to decades, there are potential feedbacks between decomposition and litter chemical composition with decomposition altering chemical composition that in turn alters the decomposition rate. These feedbacks lead to a general slowing of the decomposition rate4 and the enrichment of certain chemical constituents such as lignin and waxes which may lead to an eventual convergence of chemical composition3,5,6,7,8,9.
The controls of short- and long-term litter decomposition have been extensively studied at various spatial scales1,10,11,12,13,14. At the global scale, the primary controls of litter decomposition are climate variables such as temperature and moisture, the secondary controls include initial litter quality (physical properties, nutrient contents and organic composition), and the tertiary controls are site chemistry and soil biota1,2,3,12,15,16. At finer spatial scales within a particular climatic region where climate is less variable, initial litter chemistry becomes the primary control of litter decomposition, with site properties such as soil type, biochemical status and soil biota potentially becoming more important13,17,18.
Given the potential feedback between decomposition and litter chemistry, studies on the changes in chemical composition during litter decomposition are needed to fully understand the decomposition process and associated controlling mechanisms. Litter chemistry is generally regarded to be closely associated with the structure, turnover and stability of soil organic mater (SOM)17,19, decomposer community structure and microbial activity20, and soil nutrient cycling21. In contrast to effects of initial litter quality on decomposition, there is less work on the long-term decomposition-chemistry feedback.
Inconsistent results regarding the changes in chemical composition during litter decomposition have been previously reported. Some researchers suggested that the chemistry of different litter types converged during decomposition7, which has been supported by many results of litter bag studies3,6,8,9. Others argued that the difference in initial chemical composition among different litter types would have persistent effects on the litter chemical composition after decomposition13,22. Moreover, different chemical composition pathways could occur, even within one litter type, due to the effects of different decomposition communities and enzyme activities13,16. The discrepancy among different studies could be attributed to the variations in litter type, soil type, site environmental factors, duration of litter decomposition, or the analytical method used13,14,18.
As noted above, one of the major challenges in understanding changes in litter chemistry during decomposition is related to methodology. Although proximate analysis has been routinely used to determine the chemical compositions of litters23,24, the correspondence between these operationally determined forms of C (i.e., resistance to hydrolysis in acid) and the actual forms (i.e., lignin) is often indirect and sometimes misleading. For instance, lignin determined on the basis of resistance to hydrolysis in heated acid solutions may potentially include or actually be other materials, such as cutin and tannin25. Recently, some high-resolution analytical techniques, such as solid-state nuclear magnetic resonance (NMR), pyrolysis-gas chromatography/mass spectrometry (py-GCMS) and near-infrared spectrometry, have been adopted to characterize the chemical composition of litter samples3,13,14. Organic chemical structures of litter have been characterized primarily through 13C cross polarization/magic angle spinning (CP/MAS)6,26,27, and occasionally by13C direct polarization/magic angle spinning (DP/MAS)3 NMR techniques. Some indices, such as the alkyl C to O-alkyl C (A/O-A) ratio and carbohydrate C to methoxy C (CC/MC) ratio, have been calculated from the NMR spectra to describe the extent of chemical changes in litter decomposition6,26,28.
Despite the potential of NMR, the broad peaks typically associated with 13C CP/MAS or DP/MAS spectra of litter present a challenge in identifying specific types of C involved3,17. Recently a series of advanced spectral editing techniques have been developed and applied for systematically elucidating the structure of complex organic matter29. For example, the13C chemical shift anisotropy (CSA) filter technique is capable of selecting sp3-hybridized C, hence overlapping signals in the 90–110 ppm region attributed to anomeric groups (representing carbohydrates)30. In addition, dipolar dephasing technique can separate methoxyl C (characteristic of lignin) from NCH C (characteristic of proteins and peptides), despite that their signals present in the same spectral region (60–45 ppm). These spectral editing techniques enable one to obtain more details in structural changes associated with litter decomposition, and more accurate estimates of how decomposition alters litter chemical composition.
In the present study, we employed several advanced 13C NMR spectral editing techniques to investigate the changes in chemical composition in foliar litters during long-term decomposition. Undecomposed and decomposed litter samples were collected from the Long-term Intersite Decomposition Experiment Team (LIDET) project in which the decomposition of 30 species of litter was investigated at 27 sites by a standardized method4. The litter samples of four species decomposed for up to 10 years at four sites with different mean annual temperatures (MAT) and mean annual precipitation (MAP) (from Arctic to tropical forest) were used in the present study. The objectives of this study were to study the effects of plant species and climate on the changes of litter chemical composition during long-term decomposition. Specifically, we tested the following hypotheses: (1) both plant species and climate influence the changes in litter chemical composition during decomposition; (2) litter chemical convergence occurs during decomposition due to the recalcitrance of certain compounds and the creation of others by decomposers; and (3) the cumulative mass loss of decomposed litters is related to the degree of change in litter chemical composition.
Results
NMR spectroscopy of undecomposed litter samples
The NMR peaks were assigned to the major components in undecomposed litters based on spectral-editing spectra and previous NMR studies of litter29,33,34. The CP/TOSS spectra (Fig. 1a–d) of four undecomposed litters were all dominated by signals from oxygenated aliphatic C from carbohydrates and assigned as follows: ~72 ppm, OCH groups (C2, C3 and C5 of pyranoside rings in cellulose and hemicellulose); ~62 ppm, OCH2 groups (C6-C); ~82 ppm, OCH groups (C4-C) and ~103 ppm, anomeric (O-C-O) C (C1-C). Anomeric C was separated from the aromatic C at 109 ppm in the 13C CSA-filtered spectra (Fig. 1a2–d2). These assignments can also be assisted by the spectra after 13C CSA filter and dipolar dephasing (Fig. 1a3–d3)29 as well as spectra after 13C CSA filter and dipolar dephasing and short CP (Fig. 1a4–d4).
Figure 1. Spectral editing for undecomposed Acer saccharum (ACSA), Drypetes glauca (DRGL), Pinus resinosa (PIRE) and Thuja plicata (THPL);
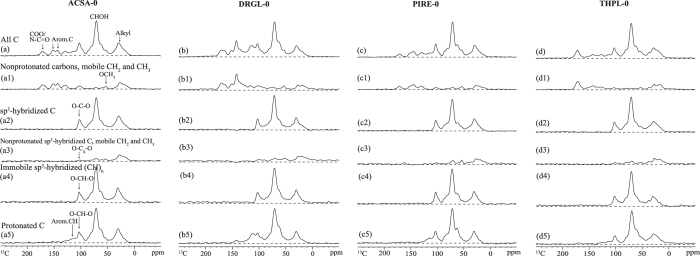
(a–d) full CP/TOSS spectra for reference with a contact time of 1 ms; (a1–d1) dipolar dephasing spectra showing nonprotonated C and mobile segments such as CH3 and OCH3 with 40 μs dephasing time; (a2–d2) selection of alkyl C with a 13C CSA filter, which in particular identifies OCO-C typical of sugar rings (CSA filter time = 47 μs); (a3–d3) selection of nonprotonated and mobile alkyl C with a CSA filter and dipolar dephasing, which in particular identifies OC (RR’)O-C (CSA filter time = 47 μs; dipolar dephasing time = 40 μs); (a4–d4) selection of protonated alkyl C with a CSA filter and short CP, in particular OCHO around 100 ppm (CSA filter time = 47 μs; CP time = 50 μs); (a5–d5) selection of protonated-C-only groups.
The signal intensity from alkyl C region (46–0 ppm) was the second highest for four undecomposed litter species, with the peak at 29 ppm assigned to CCH2C and the signals below 24 ppm due to CH3 groups, as well as shoulders at 35 ppm attributed to CH or quaternary C. These signals mainly arose from waxes such as cutin3. Note that the shoulder at 22 ppm was due to terminal -CH3 of CH3COO of hemicelluloses. The signals in the region (60–46 ppm) included the contributions from both methoxyl C (OCH3) from lignin at 56 ppm, and N-alkyl C (NCH) (partially from peptides or proteins). The signals of the former group were selectively retained in the dipolar dephasing spectra (Fig. 1a1–d1).
Lignin/tannins were main contributors to signals in regions of aromatic C-C+/H (i.e. non-oxygenated olefinic/aromatic C, 138–109 ppm), and the oxygenated aromatic C-O (140–160 ppm). The broad peaks in aromatic region were characteristic features of guaiacyl lignin: 115 ppm, aromatic CH (G5); and 131 ppm, nonprotonated aromatics G1(ne). Note that the “ne” subscript referred to “non-etherified”. Immobile protonated carbon selection spectra (Fig. 1a5–d5) which exhibit the signals of aromatic CH, OCHO, and CHOH also confirmed the presence of aromatic CH (G5) at 115 ppm. Assisted by dipolar dephasing spectra, aromaticC-C+/H could be divided into nonprotonated aromatics (aromatic C-C) and protonated aromatics (aromatic C-H). The predominance of condensed tannins was indicated by a sharp aromatic peak at 131 ppm, and more importantly, by the split peak at 145 and 155 ppm in the aromatic C-O region3. For lignin, a dominance of guaiacyl lignin was indicated by a peak at 148 ppm together with a shoulder at 153 ppm; a peak at 153 ppm with higher intensity than 148 ppm would suggest the presence of syringyl lignin25.
The peak with a maximum at 172 ppm was attributed to carboxyl/carboxylate/amide C (COO/N-C=O) groups. The presence of NCH and amide C were partly associated with proteins/peptides; carboxyl (COO) groups plus CH3 were associated with CH3COO in hemicellulose. In addition, some COO groups were partially due to fatty acids.
Comparison of chemical composition of undecomposed litter samples
The 13C DP/MAS and DP/MAS dipolar dephasing (DD) spectra of undecomposed Drypetes glauca provided quantitative information of chemical constituents (Supplementary Fig. 1). Pinus resinosa had the highest abundances of O-alkyl C (i.e., carbohydrates), while Drypetes glauca had the lowest amount; the other two species had slightly lower amounts than Pinus resinosa (Supplementary Table 2). The amounts of anomeric C (also associated with carbohydrates) for four species were very similar (Supplementary Table 2). The small content of nonprotonated anomeric C indicated that anomeric C of four species was primarily in a protonated form (O-CH-O). This was also confirmed from Fig. 1a3–d3 in which O-Cq-O peaks (quaternary anomeric C) were inconspicuous. Thuja plicata had the largest amount of alkyl C (waxes), whereas Drypetes glauca had the smallest amount, 10% lower than Thuja plicata. All four undecomposed litters contained methoxyl C (Fig. 1a1–d1); however, the amounts were very small. Drypetes glauca had the highest amount of aromatic C-C+/H (Supplementary Table 2) as well as aromatic CH (G5) (Fig. 1a5–d5). For all four litter species, nonprotonated aromatic C accounted for a larger fraction than protonated ones (Supplementary Table 2). The peaks for Pinus resinosa and Thuja plicata in the aromatic C-O region were consistent with the presence of guaiacyl lignin, confirming that they contained primarily the G unit of lignin found in gymonosperms. The low intensity of aromatic signal for Thuja plicata was consistent with its lowest amount of methoxyl C (Supplementary Table 2), suggesting that the lignin content of this species may be low.
On the other hand, the partially split peaks at ~144 and ~154 ppm for Acer saccharum indicated the presence of tannins. These split peaks for Drypetes glauca were not as well-resolved as those for Acer saccharum, suggesting that the former may contain smaller amounts of tannins. No split peaks were observed for Pinus resinosa and Thuja plicata. As angiosperms, the higher intensity of peaks at 153 ppm of Acer saccharum and Drypetes glauca suggested that these two species may contain more syringyl lignin than Pinus resinosa and Thuja plicata. Acer saccharum and Drypetes glauca contained higher contents of aromatic C-C and aromatic C-O than Pinus resinosa and Thuja plicata as well (Supplementary Table 2).
Changes in chemical constituents of litters with decomposition
The trends in abundances of chemical functional groups in Pinus resinosa that were decomposed for 5 and 10 years at ARC, HRF and AND, and for 0.8 and 1.8 years at LUQ were shown in Fig. 2a. Decomposed Pinus resinosa at the coldest site, ARC, showed relatively constant chemical compositions after 5- and 10-year decomposition; the extent of changes in all chemical functional groups were ca. 1% except for a decrease in alkyl C of 3.1% after 5-year decomposition. Specifically, Pinus resinosa exhibited a decrease in O-alkyl as well as alkyl C but an increase of carboxyl/carboxylate/amide C after 5-year decomposition. After 10 years at AND, Pinus resinosa continued to lose O-alkyl C by 8.9% in total, but alkyl C increased by 5.4% relative to its initial abundance in undecomposed litters. Anomeric C started to decrease at this stage, but the change was relatively small. The contents of all other chemical functional groups of this species at this site remained generally constant. At the HRF, Pinus resinosa litter followed a similar decomposition pattern to that at AND, but showed larger changes in O-alkyl and alkylC abundances. Despite only two years of decomposition of Pinus resinosa at LUQ, there was a similar degree of decrease in O-alkyl C and anomeric C to 10 years of decomposition at AND and HRF. In contrast to the AND and HRF sites there appeared to be almost no increase in alkyl C at LUQ. Another difference was that aromatic C-C+/H was more enriched at LUQ than at the other sites.
Figure 2.
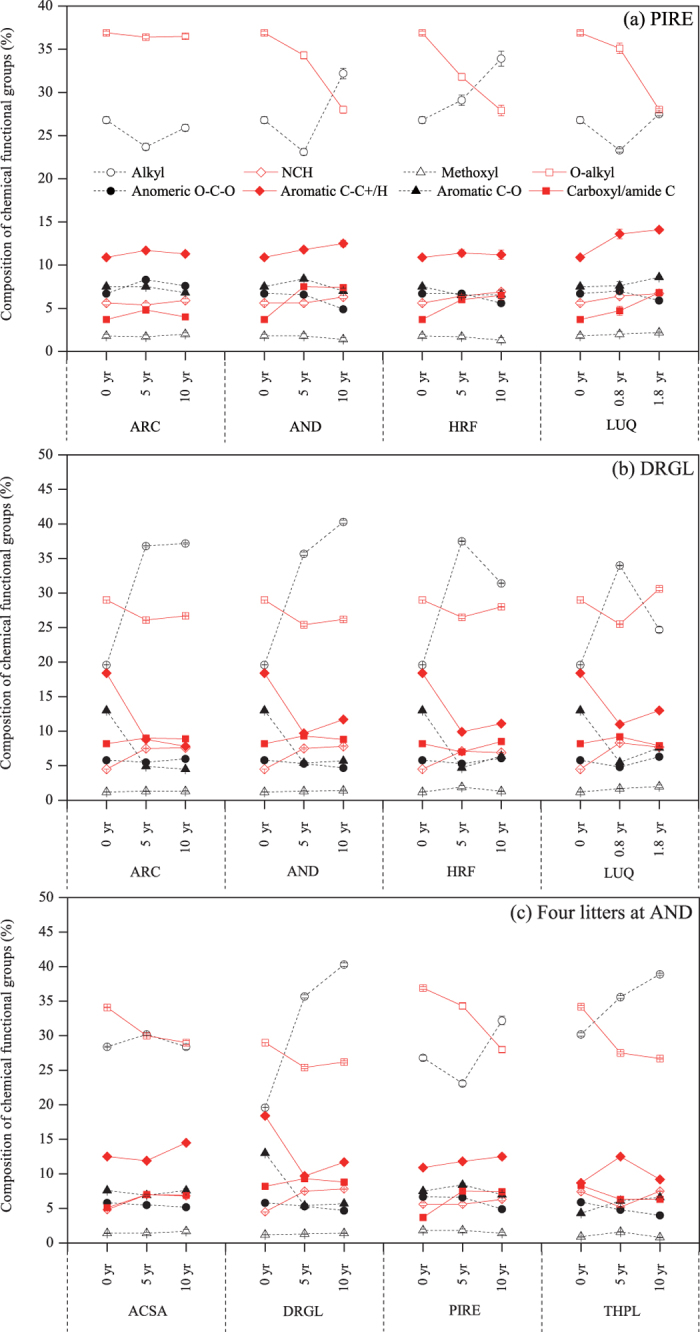
Composition of chemical functional groups (%) in (a) Pinus resinosa (PIRE) at four sites, (b) Drypetes glauca (DRGL) at four sites, and (c) four litters at H. J. Andrews Forest (AND) obtained by 13C CP/TOSS and spectral editing technique. Error bars represent the level of S/N ratio.
The relative C abundance of waxes for Pinus resinosa decreased at year 5 and 10 at ARC, at year 5 at AND, and at year 0.8 at LUQ, but increased in all other samples (Fig. 3a). Carbohydrates decreased in all decomposed samples, except for a slight increase for samples at ARC site (Fig. 3a). Lignins/tannins either did not change or increased as decomposition proceeded. Proteins/peptides were slightly enriched (Fig. 3a).
Figure 3.
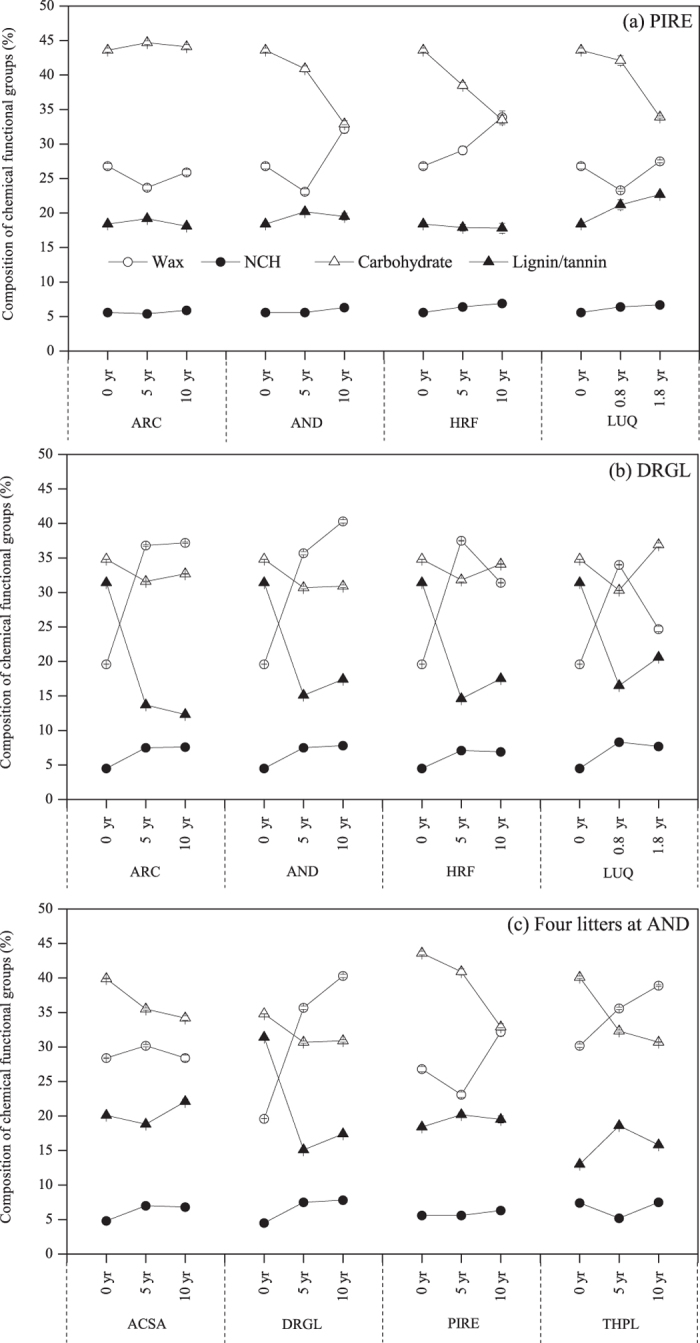
Relative C abundance of different organic compounds (%) in (a) Pinus resinosa (PIRE) at four sites, (b) Drypetes glauca (DRGL) at four sites, and (c) four litters at H. J. Andrews Forest (AND) obtained by 13C CP/TOSS and spectral editing technique. Error bars represent the level of S/N ratio.
Drypetes glauca had a different pattern of compositional change than Pinus resinosa (Fig. 3a,b). Compared with the undecomposed litter, decomposed Drypetes glauca litters exhibited a progressive enrichment in alkyl C and a decline in relative percentages of aromatic C-C+/H and aromatic C-O, indicating the loss of lignin/tannins. The decrease of O-alkyl C and anomeric C associated with carbohydrates was not as remarkable as that of lignin/tannins. Moreover, the relative intensity of methoxyl groups remained unchanged, whereas NCH C was enriched during decomposition. Drypetes glauca became significantly more enriched in waxes and proteins/peptides after decomposition than observed in decomposed Pinus resinosa samples (Fig. 3a,b). Contrary to Pinus resinosa samples, lignin/tannins clearly decreased in Drypetes glauca, with less than half of the initial amount in some samples (Fig. 3b). Carbohydrates generally became less abundant with the exception of a slight increase after 1.8 years at LUQ (Fig. 3b).
The changes in composition of chemical functional groups of four litters at AND had a tendency of an enrichment of alkyl C (except for the Acer saccharum) and a decrease of O-alkyl C, while the changes of other functional groups were inconsistent (Fig. 2c). The trends in the change of aromatic C-O were inconsistent for all four species during decomposition, i.e., enrichment for Thuja plicata, reduction for Drypetes glauca, and generally no change for Acer saccharum and Pinus resinosa (Fig. 2c). At AND alkyl C were enriched in all decomposed samples except for Pinus resinosa at the fifth year and Acer saccharum at the tenth year. Drypetes glauca had the largest enrichment of alkyl C after decomposition. The relative intensity of NCH groups increased slightly for all species except for Thuja plicata at the fifth year. The content of methoxyl C remained unchanged. Both O-alkyl and anomeric C, especially protonated anomeric C, became less abundant during decomposition at AND (Fig. 2c), suggesting that carbohydrates may be decomposed (Fig. 3c).
Convergence of litter chemistry during decomposition
PCA was applied to evaluate the effects of litter quality on the degradation of four species at AND. The relative importance of each chemical functional group to the first three principal components (PCs) was obtained (Supplementary Table 3). The first component (PC1) accounted for 52.1% of total variance and was dominated by high negative loadings of alkyl (−0.918) and NCH (−0.851) and positive loadings of anomeric C-C+/H (0.781), aromatic C-O (0.760), O-alkyl (0.673) and aromatic C-C+/H (0.666) (Supplementary Table 2). The largest positive and negative loadings for PC2 (23.0% of variance) were aromatic C-C+/H (0.704) and O-alkyl (–0.663), respectively (Supplementary Table 3).
The enclosed areas of 0-year litters (undecomposed litters) were large, indicating diverse initial litter composition (Fig. 4a). As decomposition proceeded, most sample points approached lower PC1 scores, indicating the enrichment of waxes and proteins/peptides (Fig. 4a). The areas enclosed by the four decomposed litters at 5-and 10-year decomposition became smaller than that enclosed by 0-year litters and the shape of enclosed areas also changed, becoming less variable in the PC2 axis. This indicated that aromatic C-C+/H became less abundant for Drypetes glauca but enriched in the other three litters. In contrast, the enclosed area remained fairly variable on the PC1 axis (Fig. 4a).
Figure 4.
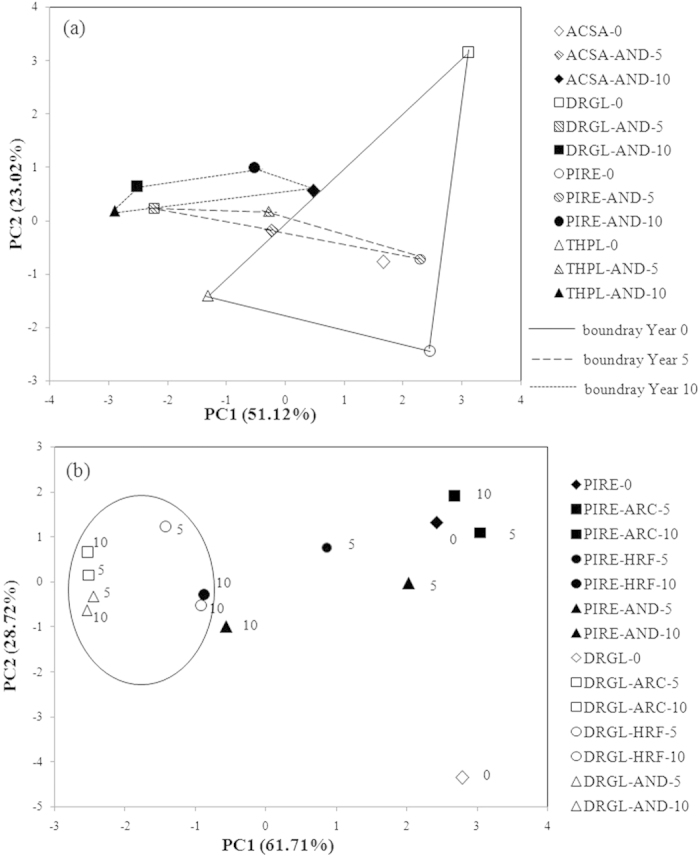
Principle component analysis of the composition of chemical functional groups for (a) four species, including Acer saccharum (ACSA), Drypetes glauca (DRGL), Pinus resinosa (PIRE) and Thuja plicata (THPL), at H. J. Andrews Forest (AND) at decomposition year of 0, 5 and 10 and (b) Pinus resinosa (PIRE) and Drypetes glauca (DRGL) at three sites, including Arctic Lakes (ARC), H. J. Andrews Forest (AND) and Harvard Forest (HRF), at decomposition year of 0, 5 and 10.
PCA was conducted for two species (Pinus resinosa and Drypetes glauca) at three sites (ARC, HRF and AND) at decomposition years of 0, 5 and 10 (Fig. 4b). The first component (PC1) accounted for 61.7% of the total variance and was dominated by a negative loading for alkyl (–0.951) and NCH (–0.943), positive loading for O-alkyl (0.895) and smaller loading for anomeric (0.765), aromatic C-O (0.733) and aromatic C-C+/H (0.616) C (Supplementary Table 4). The largest loadings for PC2 (28.7% of variance) were negative loadings of aromatic C-C+/H and aromatic C-O and positive loadings of methoxyl C (Supplementary Table 4).
Drypetes glauca and Pinus resinosa showed a general convergence in terms of composition. Undecomposed Pinus resinosa and Drypetes glauca litters had relatively high PC1 scores (Fig. 4b) because of their high content of O-alkyl C and low content of alkyl and NCH C. Undecomposed Drypetes glauca was separated from undecomposed Pinus resinosa by the low PC2 scores of Drypetes glauca, due to the high aromatic C-C+/H and aromatic C-O of Drypetes glauca. With increasing decomposition time, the sample points of Drypetes glauca and Pinus resinosa approached lower PC1 score, except for Pinus resinosa at ARC. In addition, except for Pinus resinosa at ARC, Drypetes glauca moved to higher PC2 score while Pinus resinosa moved to lower PC2 score. This result might be due to the significant decrease of aromatic C-C+/H and aromatic C-O for Drypetes glauca but inconsistent changes of aromatic C-C+/H and aromatic C-O for Pinus resinosa. Decomposed Drypetes glauca at AND and ARC at 5-year and 10-year were very close in PCA space, indicating similar compositions. The points of decomposed Drypetes glauca at all sites assembled in the circle indicated that the composition converged over time regardless of the climate. Although Drypetes glauca at HRF at 10-year moved back toward undecomposed sample composition, this may not have been a significant shift. On the other hand, the pattern of decomposed Pinus resinosa samples in PCA space was not the same as for Drypetes glauca. At ARC, Pinus resinosa decomposed at 5- and 10-years were close to their undecomposed sample in PCA space. This was not true for Pinus resinosa at AND and HRF; these samples moved further away from the undecomposed samples after decomposition at a longer time. At AND and HRF, Pinus resinosa at 5-year had relatively large difference on PC1 score, but became closer to each other at 10-year. This indicated the composition of both Pinus resinosa and Drypetes glauca converged regardless of climates if the results for Pinus resinosa at ARC was set aside.
Relationship between cumulative mass losses and compositional changes
The alkyl C and O-alkyl C were two chemical functional groups that have greater changes as compared to other chemical functional groups (Fig. 2) suggesting the relative A/O-A ratio could be used as an index reflecting the extent of chemical composition change. A positive correlation (P < 0.01) between relative A/O-A and mass loss during decomposition was found in Pinus resinosa (Fig. 5a), but not in Drypetes glauca (Fig. 5b). Such correlation was not observed in the four litters at AND (Fig. 5c).
Figure 5.
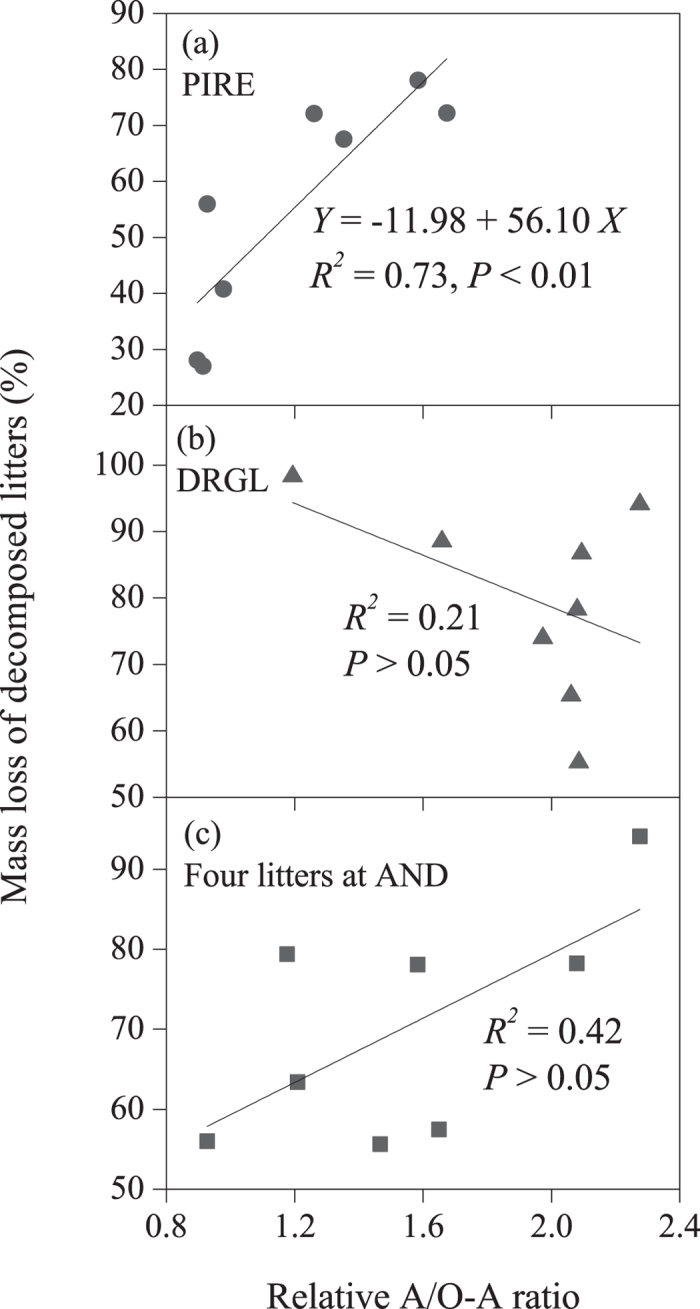
Relationship between the relative alkyl C/O-alkyl C (A/O-A) ratio and mass loss in decomposed litter samples for (a) Pinus resinosa (PIRE) at four sites, (b) Drypetes glauca (DRGL) at four sites, and (c) four litters at H. J. Andrews Forest (AND).
Discussion
Changes of litter chemical composition during decomposition
Solid-state NMR technique has been extensively used to investigate the changes in litter chemical composition during decomposition in the past two decades3,17,27,35. In the present study, we used advanced 13C solid-state NMR technique with several spectral editing techniques to obtain the detailed information of chemical composition of four litter species, which were decomposed in 4 different sites. The most significant changes in litter chemical compositions in the present study were an enrichment of alkyl C and a loss of O-alkyl C after decomposition. Previous studies also showed the increased alkyl C content in bamboo leaf litter36, maize and wheat straws8, black pine needle litter24, buffel grass (Cenchrus ciliaris) and lucerne (Medicago sativa) litters6, which would be mainly attributed to selective preservation of recalcitrant organic compounds such as waxes and cutins or an enrichment in cross-linking of the long chain alkyl compounds3,17,34,36. Numerous previous NMR studies also demonstrated that the O-alkyl C decreased after decomposition3,6,36, because the O-alkyl C compounds in litters mainly include the carbohydrates, which can be easily utilized by microorganisms3. In contrast, Almendros et al.33 and Lemma et al.34 reported that decreased alkyl C content and increased O- alkyl C content in some types of litters were observed after decomposition. In addition, we found that alkyl C decreased during the intermediate stage of decomposition (i.e., the 5-year at ARC and AND and, 0.8-year at LUQ) (Fig. 2a). This could be due to relatively quick loss of either some cuticular material or lipids in the early stages of decomposition3,33.
Unlike previous NMR litter decomposition studies, we were able to distinguish and quantify NCH and methoxyl (region of 60–45 ppm), respectively (Fig. 2, Supplementary Table 2). Our results showed that the content and change of the methoxyl C in litter samples were both very small, whereas NCH was present in comparatively larger amounts and showed larger variations during decomposition (Supplementary Table 2). Previous studies attributed the increase of intensity in the region of 60–45 ppm to preservation of methoxyl C and thus hypothesized that the preservation of lignin occurred during decomposition6,34. However, we found that NCH, instead of methoxyl C, was enriched during decomposition for three of the four species we examined (Fig. 2). We assumed that the enrichment of NCH could be due to formation of new compounds such as bacterial peptidoglycan and fungal chitin in cell walls during the decomposition6,37. Hobara et al.38 also found the yields of total hydrolysable amino acids and total hydrolysable amino sugars increased during litter decomposition, and they suggested that decomposition induced a change in litter chemistry from C-rich plant-derived biopolymers to N-rich microbially-derived biochemicals.
Though it has been assumed that lignin is preserved during decomposition5,39,40, the relative amounts of lignin in the present study did not necessarily increase with decomposition and decreased for Drypetes glauca (Fig. 2b). This was consistent with other NMR studies3,26 and contrasted with what has been reported based on proximate analyses40. In addition, the assumption that decomposition proceeded towards enrichment of lignin was not supported by the study on δ13C changes of litters during decomposition3. These contradictory effects regarding the change of lignin content can depend on differences in litter type, soil type, and site environmental factors, or the analytical method used3,13.
Therefore, our first hypothesis was supported by the results of this study that both plant species and climate markedly influence the changes in litter chemical composition during decomposition.
Litter chemical convergence during decomposition
Numerous litter bag studies have demonstrated that different litter types with various initial chemical compositions eventually converge towards a similar chemistry after extensive decomposition3,8,9,41. However, recently Wickings et al.13 revealed that the chemistry of different litter types diverged, rather than converged, by using the py-GC/MS technique, and they attributed the divergent chemical pathways among different litter types to the differences in decomposer communities13. Our results suggested that divergent initial litter chemistry did not necessarily completely converge as decomposition proceeded, but could reach several endpoints (Fig. 4). For four litters at AND, although the areas enclosed by four decomposed litters at 5- and 10-year decomposition became smaller, the shape of enclosed areas also changed as the boundary that enclosed composition after 5- and 10-year decomposition became less variable in the PC2 axis and remained fairly variable on PC1 axis. In contrast, Preston et al.3 who applied the same method found that chemical composition converged over decomposition with the distance on both PC axes decreasing. In addition, we found that the chemical composition of Pinus resinosa and Drypetes glauca converged regardless of climate conditions (Fig. 4b), with a stronger case of chemistry convergence for Pinus resinosa litter. Therefore, our second hypothesis could be supported by the result of one plant species across different climate conditions. However, it was worth mentioning that inconsistent convergence occurred among different plant litter species across a range of site conditions, particularly when LUQ included. This inconsistency may be attributed to the difference in the soil microbial community, soil invertebrate species, and site environmental factors among different sites13,42. In addition, Bedford et al.42 suggested the inconsistent convergence in chemical trajectory were related to the bag/mesh design in the litter decomposition experiment. They explained that different bag/mesh sizes would cause different chemical trajectories, through changing the water content of the decomposing leaf, creating stable habitats for invertebrates in compressed layers, and consequently causing the selective decomposition of plant tissues42.
In addition, we found the different change patterns in the relative C abundance of different organic compounds (including waxes, carbohydtrates, lignins/tannins, and proteins/peptides) among different plant litter species (Fig. 3). Therefore, we can conclude the plant species rather than climate greatly altered the temporal pattern of litter chemical composition during long-term decomposition.
It is worth noting that the difference in site conditions (such as soil types, climate conditions, management practices applied) may partially attribute to the inconsistent convergence or divergence in the chemical pathways8,13,14, and thus, temporal patterns of litter chemical composition over long-period decomposition or across large spatial scales could be quite different in various litter species-site combinations.
Relationship between chemical compositional change and mass losses in decomposed litters
Our results showed that changes in the abundance of alkyl and O-alkyl C were the most consistent and obvious ones during decomposition for all four species, confirming that the A/O-A ratio could be a robust index of the degree of compositional changes26,32. To eliminate the effects of variation in A/O-A ratio among the undecomposed litter types, we used the relative A/O-A ratio as an index to indicate the degree of compositional changes in this study. The positive correlation between the relative A/O-A ratio and mass loss during decomposition was found in Pinus resinosa (Fig. 5a), but not in Drypetes glauca (Fig. 5b), and such correlation was not found in the four litters at AND (Fig. 5c). These results lead to a rejection of our third hypothesis that the more mass lost, the greater changes in chemical compositions that would occur. In combination with the inconsistent convergence of chemical pathways caused by different litter types revealed by the present study and results of Wickings13, the effect of chemical composition changes on the litter decay rate might be more complicated than previously expected3,9,41. To efficiently estimate the mass loss during litter decomposition, Adair et al.1 developed a model with three C pools (a rapidly decomposing labile pool, an intermediate pool, and a recalcitrant pool), which could explain approximately 70% of the variation of data in their study. According to our results, we can assume that the systematic deviations of predictions by such a model would be partially attributed to the various temporal pattern of litter chemical composition during long-term decomposition.
In conclusion, our study has made a detailed investigation on the changes of litter chemical composition during long-term decomposition across broad spatial scales by using advanced solid-state 13C NMR techniques. Our results showed strong evidence that the temporal pattern of litter chemical composition depended largely on the litter species rather than climate. In addition, we also found that the relationship between litter mass loss and changes of chemical composition were affected by plant species and climate conditions. These findings indicated that the decomposition-chemistry feedback would greatly vary among different plant species, and thus parameters representing some plant traits should be incorporated into models that predict the litter chemical change, especially over a long-term period. Further research should be conducted to elucidate the contributions of various chemical pathways of different litter types to the selective preservation, humification, and stability of soil organic matter.
Materials and Methods
The LIDET study and litter sample selection
The LIDET was initiated in 1990 to investigate the impacts of substrate quality and macroclimate on the decomposition of leaf and fine root litters over a 10 year period4,31. A standardized methodology for measuring litter decomposition was used in LIDET at 27 different sites with various climate conditions to study the decomposition patterns of 30 species of litter during the first decade of decomposition4. The undecomposed and decomposed samples in LIDET study provide an opportunity to study the effects of plant species and climate conditions on the temporal pattern of litter chemical composition during the long-term decomposition. Because the analysis using different NMR techniques requires significant instrument time, we selected four plant species from four sites to study the effects of plant species and climate on the temporal pattern of litter chemical composition during long-term decomposition.
Undecomposed and decomposed litter samples of Drypetes glauca (DRGL) and Pinus resinosa (PIRE) from the Arctic Lakes (ARC), H. J. Andrews Forest (AND), Harvard Forest (HRF), and Luquillo Tropical Forest (LUQ) sites were collected to examine interactions between site climate and litter of plant species effects. Previous results of the LIDET study indicated that these two species were significantly different in initial substrate quality (Supplementary Table 1) and exhibited the most consistent temporal patterns of decomposition, with PIRE generally following a single negative exponential decline and DRGL generally following a dual exponential decline4. The four sites represented a strong environmental gradient, with mean annual temperatures (MAT) ranging from −7 to 23 °C (Table 1). While MAT at the AND and HFR were just a few degrees apart (8.6 versus 7.1 °C, respectively), their seasonal distribution of mean annual precipitation (MAP) was quite different, with AND having a distinct dry summer and HFR having a uniform seasonal distribution.
Table 1. Climatic characteristics and biome types of 4 long-term intersite decomposition experiment team (LIDET) sites.
| Site | Code | Latitude | Longitude | Elevation(m) | MAP(cm) | MAT(°C) | AET(cm) | PET(cm) | Biome Type |
|---|---|---|---|---|---|---|---|---|---|
| Arctic Lakes, Alaska | ARC | 63°38’N | 149°34’W | 760 | 32.7 | –7 | 28.4 | 42.3 | Arctic Tundra |
| Harvard Forest, Massachusetts | HRF | 42°40’N | 72°15’W | 335 | 115.2 | 7.1 | 85.1 | 104.1 | Temperate Deciduous Forest |
| H. J Andrews Experimental Forest, Oregon | AND | 44°14’N | 122°11’W | 500 | 230.9 | 8.6 | 76.4 | 98.2 | Temperate Conifer Forest |
| Luquillo Experimental Forest, Puerto Rico | LUQ | 18°19’N | 65°49’W | 350 | 336.3 | 23 | 123.4 | 125.9 | Humid Tropical Forest |
To investigate the effect of plant species on the temporal pattern of litter chemical composition within one climate condition, we also used litter samples of Acer saccharum (ACSA) and Thuja plicata (THPL) from AND, in addition to DRGL and PIRE, because these two species had different initial chemistry (Supplementary Table 1) and represented a wider range of taxa. The site of AND represented an intermediate environmental condition. Four litters examined were decomposed for 5 and 10 years in ARC, AND and HRF, but only decomposed for 0.8 and 1.8 years in LUQ because of the high rate of decomposition at this site. Mass remaining of four litter species after a period of decomposition was determined. For example, after 5 years’ decomposition, the mass remaining (%) for Pinus resinosa in ARC, AND and HRF were 71.9%, 44%, and 27.9%, respectively, and these values were 59.2%, 21.9%, and 27.7%, respectively, after 10 years’ decomposition. In LUQ, after 0.8 and 1.8 years’ decomposition, the mass remaining (%) for Pinus resinosa were 73% and 32.4%, respectively (Supplementary Table 5).
13C NMR spectroscopy
Solid state 13C NMR analyses were performed using a Bruker Avance III 300 spectrometer at 75 MHz for 13C (300 MHz for 1H frequency). Samples were run in a double-resonance probe head using 4-mm rotors. NMR experiments included quantitative 13C DP/MAS and DP/MAS plus recoupled dipolar dephasing (DP/MAS/DD), 13C cross polarization and total suppression of sidebands (CP/TOSS) and 13C CP/TOSS plus dipolar dephasing (DD), 13C chemical-shift-anisotropy (CSA) filter, spectral editing of immobile CH2 + CH.
The quantitative 13C DP/MAS NMR experiments were conducted at a spinning speed of 13 kHz with a 200 s recycle delay and 1024 scans. The 90° 13C pulse-length was 4 μs. The recycle delay was determined by the cross polarization/spin-lattice relaxation time/total sideband suppression (CP/T1-TOSS) technique to ensure that all C nuclei were more than 95% relaxed29. Nonprotonated C and mobile C fractions such as methoxyl (OCH3) C and CCH3 were quantified after DP/MAS technique with a recoupled dipolar-dephasing delay of 68 μs29. The recycle delay and number of scans used were the same as for DP/MAS experiment. Considering the long measuring time and similar types of components for four litter species, DP/MAS and DP/MAS/DD spectra were only collected for undecomposed DRGL. The quantitative data of other samples were obtained by calculation of CP efficiency of specific chemical shift regions.
The 13C CP/TOSS/DD was conducted using a 13C CP/MAS NMR technique with a spinning speed of 5 kHz, a contact time of 1 ms and a 1H 90° pulse-length of ca. 4 μs. Four-pulse total suppression of sidebands (TOSS) was employed before detection. Two-pulse phase-modulated (TPPM) decoupling was applied for optimum resolution during detection. Sub-spectra for nonprotonated and mobile C groups were obtained by combining the 13C CP/TOSS sequence with a 40-μs DD. The number of scans of 13C CP/TOSS and 13C CP/TOSS/DD spectra was 6144 for all litter samples. The recycle delays for 13C CP/TOSS and 13C CP/TOSS/DD was 1 s.
The 13C CSA filter inserted into 13C CP/TOSS to select the signals of sp3-hybridized C. Specifically, this technique separated signals of anomeric (O-C-O) C in carbohydrate rings from those of aromatic C-C+/H by selectively suppressing the latter with a five-pulse 13C CSA filter using a CSA-filter time of 47 μs30. Non-protonated anomeric C were obtained by combining this filter with a dipolar dephasing time of 40 μs. The CSA filter was also combined with short CP to give selective spectra of protonated anomeric C. A total of 6144 scans were averaged for spectra of CSA filter, CSA filter coupled with short CP, and CSA filter coupled with DD, with a recycle delay of 1 s.
The spectra for immobile CH2 + CH groups were obtained from the difference of two spectra. The first one was a 13C CP/TOSS spectrum with a short CP time of 50 μs to emphasize protonated C in immobile segments, and the second 13C CP/TOSS spectrum was recorded using a short CP of 50 μs coupled with a 40 μs DD, which only contained the residual signals of quaternary C or mobile segments. The second spectrum was subtracted from the first one, and the difference spectrum represented immobile CH2 and CH C, with a small CH3 contribution29. The number of scans was 6144 for all analyzed samples.
Following our previous publications and related literature3,17,29, the NMR spectra obtained in this study were divided into eight regions representing different chemical environments of a 13C nucleus: alkyl C (46–0 ppm), methoxyl C/NCH (60–46 ppm), O-alkyl C (92–60 ppm), anomatic C-C/anomatic C-H (109–92 ppm), aromatic C-C/aromatic C-H (138–109 ppm), aromatic C-O (162–138 ppm), carboxyl/amide (190–162 ppm) and aldehyde/ketone (220–190 ppm). The area in each region was calculated by integration, and the relative contents of different C fractions were obtained. On the basis that one (or more) particular chemical functional group is often characteristic of a specific compound, some chemical functional groups identified by NMR analysis can represent major classes of organic compounds. Alkyl C, O-alkyl C and anomatic C, aromatic C-C+/H and aromatic C-O, NCH were characteristic functional groups of waxes, carbohydrates, lignins/tannins, and proteins/peptides, respectively. Thus, the relative C abundance of waxes, carbohydrates, lignin/tannins, and proteins/peptides were calculated by the corresponding C function groups. The alkyl C/O-alkyl C (A/O-A) ratio has been extensively used to represent the extents of changes in chemical composition6,26,32. To eliminate the effects of variation in A/O-A in the undecomposed litters, the relative A/O-A was used in this study. The relative A/O-A ratio was defined as:
Relative A/O-A ratio = (A/O-A value of the sample after a period of degradation) / (A/O-A value of the undecomposed sample).
Statistical analysis
All of the statistical analyses were performed using the SPSS software (SPSS 13.0 for windows, SPSS Inc., Chicago, USA). Principal component analysis (PCA) was performed on the composition of chemical functional groups in litters. Linear regression analyze was conducted to determine the relationship between the relative A/O-A ratio and mass loss in decomposed litter samples.
Additional Information
How to cite this article: Li, Y. et al. Plant species rather than climate greatly alters the temporal pattern of litter chemical composition during long-term decomposition. Sci. Rep. 5, 15783; doi: 10.1038/srep15783 (2015).
Supplementary Material
Acknowledgments
This study would not have been possible without the dedicated efforts of many people, who have participated in the LIDET study. In addition to the numerous grants that supported personnel at the individual sites, this study was supported by grants from the National Science Foundation (DEB-9108329, DEB-9806493) and the Kaye and Ward Richardson Endowment.
Footnotes
Author Contributions Y.F.L. and N.C. performed the experimental work, analyzed the data, and drafted the manuscript. J.D.M. and M.E.H. proposed the research, supervised the projects and edited the manuscript. Y.L., X.Y.C. and M.A.C. collected the data. All the authors discussed the results and commented on the contents of the manuscript.
References
- Adair E. C. et al. Simple three-pool model accurately describes patterns of long-term litter decomposition in diverse climates. Global Change Biol. 14, 2636–2660 (2008). [Google Scholar]
- Prescott C. E., Vesterdal L., Preston C. M. & Simard S. W. Influence of initial chemistry on decomposition of foliar litter in contrasting forest types in British Columbia. Can. J. Forest Res. 34, 1714–1729 (2004). [Google Scholar]
- Preston C. M., Nault J. R. & Trofymow J. A. Chemical changes during 6 years of decomposition of 11 litters in some canadian forest sites. part 2. C-13 abundance, solid-state C-13 NMR spectroscopy and the meaning of “lignin”. Ecosystems 12, 1078–1102 (2009). [Google Scholar]
- Harmon M. E. et al. Long-term patterns of mass loss during the decomposition of leaf and fine root litter: an intersite comparison. Global Change Biol. 15, 1320–1338 (2009). [Google Scholar]
- Berg B. & Staaf H. Decomposition rate and chemical changes of scots pine needle litter. I. influence of stand age. Ecol. Bull. 32, 363–372 (1980). [Google Scholar]
- Mathers N. J., Jalota R. K., Dalal R. C. & Boyd S. E. C-13-NMR analysis of decomposing litter and fine roots in the semi-arid Mulga Lands of southern Queensland. Soil Biol. Biochem. 39, 993–1006 (2007). [Google Scholar]
- Fierer N., Grandy A. S., Six J. & Paul E. A. Searching for unifying principles in soil ecology. Soil Biol. Biochem. 41, 2249–2256 (2009). [Google Scholar]
- Wang X., Sun B., Mao J., Sui Y. & Cao X. Structural convergence of maize and wheat straw during two-year decomposition under different climate conditions. Environmen. Sci. Technol. 46, 7159–7165 (2012). [DOI] [PubMed] [Google Scholar]
- Wallenstein M. D. et al. Litter chemistry changes more rapidly when decomposed at home but converges during decomposition–transformation. Soil Biol. Biochem. 57, 311–319 (2013). [Google Scholar]
- Sjoberg G., Nilsson S. I., Persson T. & Karlsson P. Degradation of hemicellulose, cellulose and lignin in decomposing spruce needle litter in relation to N. Soil Biol. Biochem. 36, 1761–1768 (2004). [Google Scholar]
- Moorhead D. L. & Sinsabaugh R. L. A theoretical model of litter decay and microbial interaction. Ecol. Monogr. 76, 151–174 (2006). [Google Scholar]
- Cornwell W. K. et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 11, 1065–1071 (2008). [DOI] [PubMed] [Google Scholar]
- Wickings K., Grandy A. S., Reed S. C. & Cleveland C. C. The origin of litter chemical complexity during decomposition. Ecol. Lett. 15, 1180–1188 (2012). [DOI] [PubMed] [Google Scholar]
- Parsons S. A., Congdon R. A. & Lawler I. R. Determinants of the pathways of litter chemical decomposition in a tropical region. New Phytol. 10.1111/nph.12852 (2014). [DOI] [PubMed] [Google Scholar]
- Valachovic Y. S., Caldwell B. A., Cromack K. & Griffiths R. P. Leaf litter chemistry controls on decomposition of Pacific Northwest trees and woody shrubs. Can. J. Forest Res. 34, 2131–2147 (2004). [Google Scholar]
- Garcia-Palacios P., Maestre F. T., Kattge J. & Wall D. H. Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecol. Lett. 16, 1045–1053 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kogel-Knabner I. The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter. Soil Biol.Biochem. 34, 139–162 (2002). [Google Scholar]
- Currie W. S. et al. Cross-biome transplants of plant litter show decomposition models extend to a broader climatic range but lose predictability at the decadal time scale. Global Change Biol. 16, 1744–1761 (2010). [Google Scholar]
- Schmidt M. W. et al. Persistence of soil organic matter as an ecosystem property. Nature 478, 49–56 (2011). [DOI] [PubMed] [Google Scholar]
- Gul S., Whalen J. K., Ellis B. E. & Grayston S. J. Plant residue chemistry impacts soil processes and microbial community structure: A study with Arabidopsis thaliana cell wall mutants. Appl. Soil Ecol. 60, 84–91 (2012). [Google Scholar]
- Bansal S., Sheley R. L., Blank B. & Vasquez E. A. Plant litter effects on soil nutrient availability and vegetation dynamics: changes that occur when annual grasses invade shrub-steppe communities. Plant Ecol. 215, 367–378 (2014). [Google Scholar]
- Wang H. et al. Dynamics and speciation of organic carbon during decomposition of leaf litter and fine roots in four subtropical plantations of China. Forest Ecol. Manag. 300, 43–52 (2013). [Google Scholar]
- Fioretto A., Papa S., Pellegrino A. & Fuggi A. Decomposition dynamics of Myrtus communis and Quercus ilex leaf litter: Mass loss, microbial activity and quality change. Appl. Soil Ecol. 36, 32–40 (2007). [Google Scholar]
- De Marco A. et al. Decomposition of black locust and black pine leaf litter in two coeval forest stands on Mount Vesuvius and dynamics of organic components assessed through proximate analysis and NMR spectroscopy. Soil Biol. Biochem. 51, 1–15 (2012). [Google Scholar]
- Preston C. M., Trofymow J. A., Sayer B. G. & Niu J. N. C-13 nuclear magnetic resonance spectroscopy with cross-polarization and magic-angle spinning investigation of the proximate-analysis fractions used to assess litter quality in decomposition studies. Can. J. Bot. 75, 1601–1613 (1997). [Google Scholar]
- Baldock J. A. et al. Assessing the extent of decomposition of natural organic materials using solid-state C-13 NMR spectroscopy. Aust. J. Soil Res. 35, 1061–1083 (1997). [Google Scholar]
- Bonanomi G. et al. Litter quality assessed by solid state 13C NMR spectroscopy predicts decay rate better than C/N and Lignin/N ratios. Soil Biol. Biochem. 56, 40–48 (2013). [Google Scholar]
- Bonanomi G. et al. Phytotoxicity, not nitrogen immobilization, explains plant litter inhibitory effects: evidence from solid-state 13C NMR spectroscopy. New Phytol. 191, 1018–1030 (2011). [DOI] [PubMed] [Google Scholar]
- Mao J. D., Chen N. & Cao X. Y. Characterization of humic substances by advanced solid state NMR spectroscopy: Demonstration of a systematic approach. Org. Geochem. 42, 891–902 (2011). [Google Scholar]
- Mao J. D. & Schmidt-Rohr K. Separation of aromatic-carbon C-13 NMR signals from di-oxygenated alkyl bands by a chemical-shift-anisotropy filter. Solid State Nucl. Mag. 26, 36–45 (2004). [DOI] [PubMed] [Google Scholar]
- Long-term Intersite Decomposition Experiment Team (LIDET). Meeting the Challenges of Long-Term, Broad Scale Ecological Experiments. U.S. LTER Network Office, Seattle, WA (1995). [Google Scholar]
- Preston C. M. Carbon-13 solid-state NMR of soil organic matter - using the technique effectively. Can. J. Soil Sci. 81, 255–270 (2001). [Google Scholar]
- Almendros G., Dorado J., González-Vila F. J., Blanco M. J. & Lankes U. 13C NMR assessment of decomposition patterns during composting of forest and shrub biomass. Soil Biol. Biochem. 32, 793–804 (2000). [Google Scholar]
- Lemma B., Nilsson I., Kleja D. B., Olsson M. & Knicker H. Decomposition and substrate quality of leaf litters and fine roots from three exotic plantations and a native forest in the southwestern highlands of Ethiopia. Soil Biol. Biochem. 39, 2317–2328 (2007). [Google Scholar]
- Wang G. A., Zhang L. L., Zhang X. Y., Wang Y. H. & Xu Y. P. Chemical and carbon isotopic dynamics of grass organic matter during litter decompositions: A litterbag experiment. Org. Geochem. 69, 106–113 (2014). [Google Scholar]
- Zhang Y. et al. Chemistry of decomposing mulching materials and the effect on soil carbon dynamics under a Phyllostachys praecox bamboo stand. J. Soil. Sediment. 13, 24–33 (2013). [Google Scholar]
- Ganjegunte G. K., Condron L. M., Clinton P. W., Davis M. R. & Mahieu N. Decomposition and nutrient release from radiata pine (Pinus radiata) coarse woody debris. Forest Ecol. Manag. 187, 197–211 (2004). [Google Scholar]
- Hobara S. et al. The roles of microorganisms in litter decomposition and soil formation. Biogeochemistry 118, 471–486 (2014). [Google Scholar]
- Couteaux M. M., Bottner P. & Berg B. Litter decomposition, climate and litter quality. Trends Ecol. Evol. 10, 63–66 (1995). [DOI] [PubMed] [Google Scholar]
- Berg B. Litter decomposition and organic matter turnover in northern forest soils. Forest Ecol. Manag. 133, 13–22 (2000). [Google Scholar]
- Moore T. R., Trofymow J. A., Prescott C. E., Titus B. D. & Grp C. W. Nature and nurture in the dynamics of C, N and P during litter decomposition in Canadian forests. Plant Soil 339, 163–175 (2011). [Google Scholar]
- Bedford A. P. A modified litter bag design for use in lentic habitats. Hydrobiologia 529, 187–193 (2004). [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.